
Exploring the Modulatory Effect of High-Fat Nutrition on Lipopolysaccharide-Induced Acute Lung Injury in Vagotomized Rats and the Role of the Vagus Nerve

 , , , , ,
, , , , , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
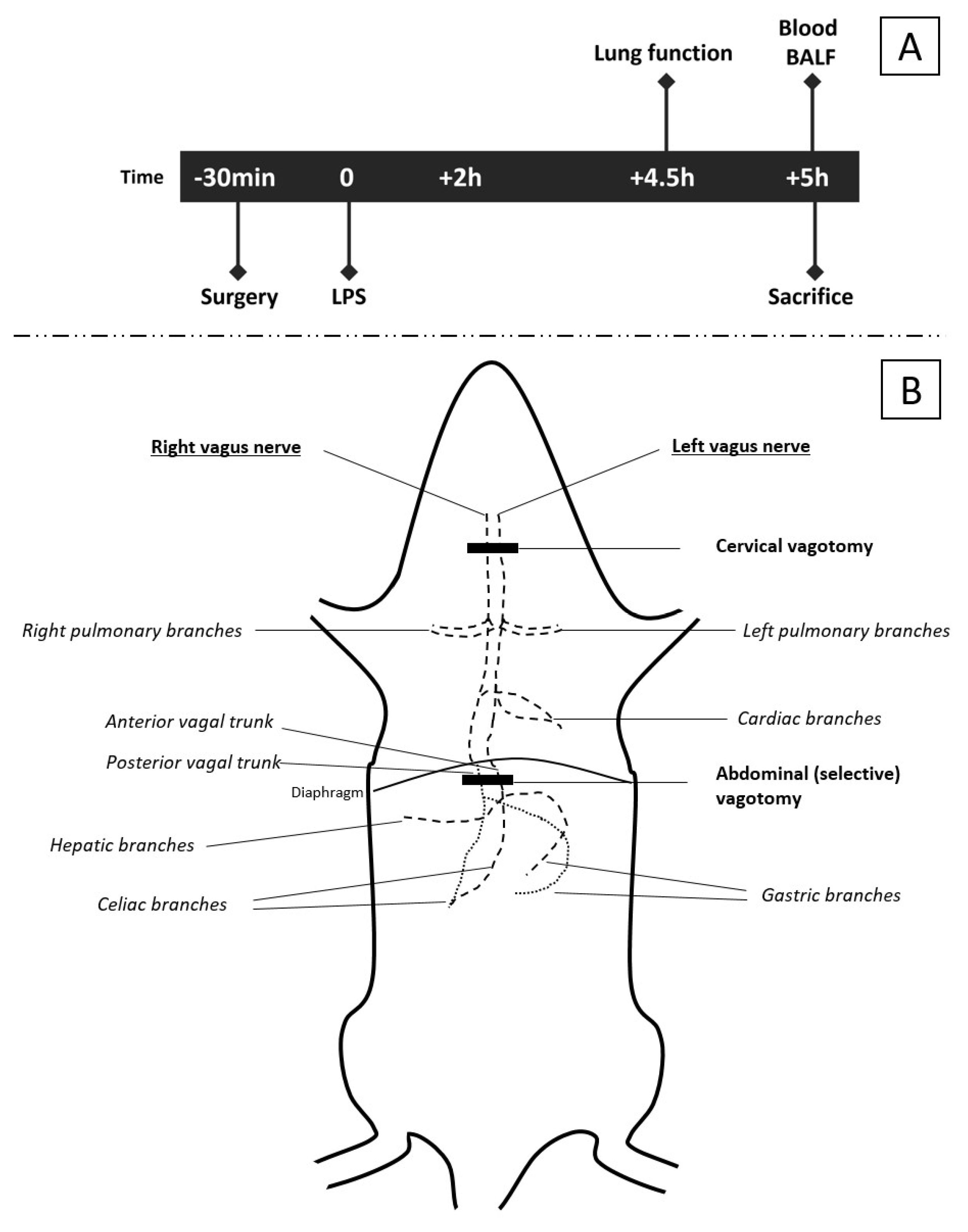
2.1. Animals
2.2. Surgical Procedure
2.3. Experimental Models
2.3.1. Model A: Effect of Selectively Sparing Vagus Nerve on Lung Injury and Function
2.3.2. Model B: Further Exploring the Role of the α7nAChR
2.3.3. Model C: Vagus Nerve Stimulation through High-Fat Enteral Nutrition
2.4. Pulmonary Function Measurement
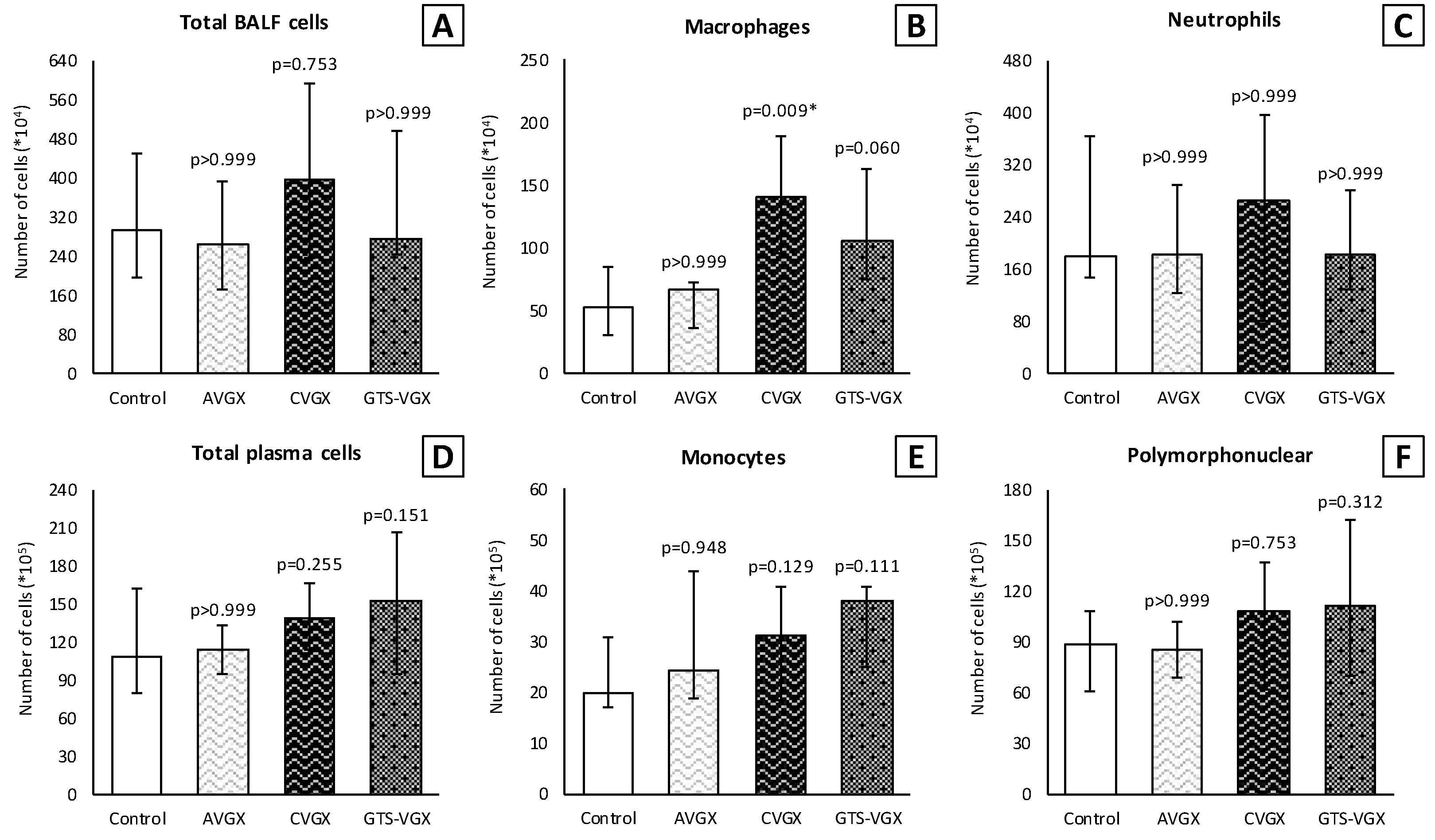
2.5. Bronchoalveolar Lavage (BAL)
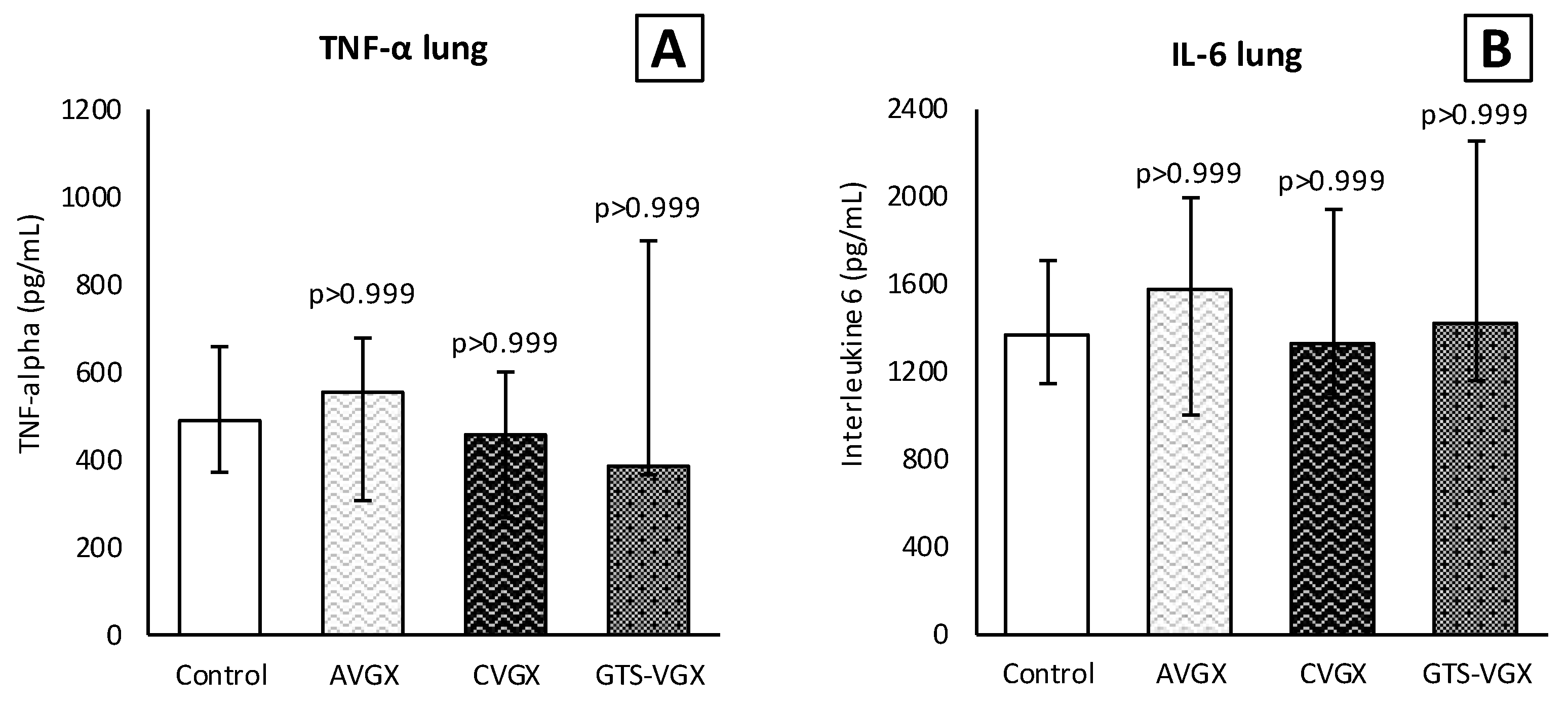
2.6. Preparation of Lung Homogenates
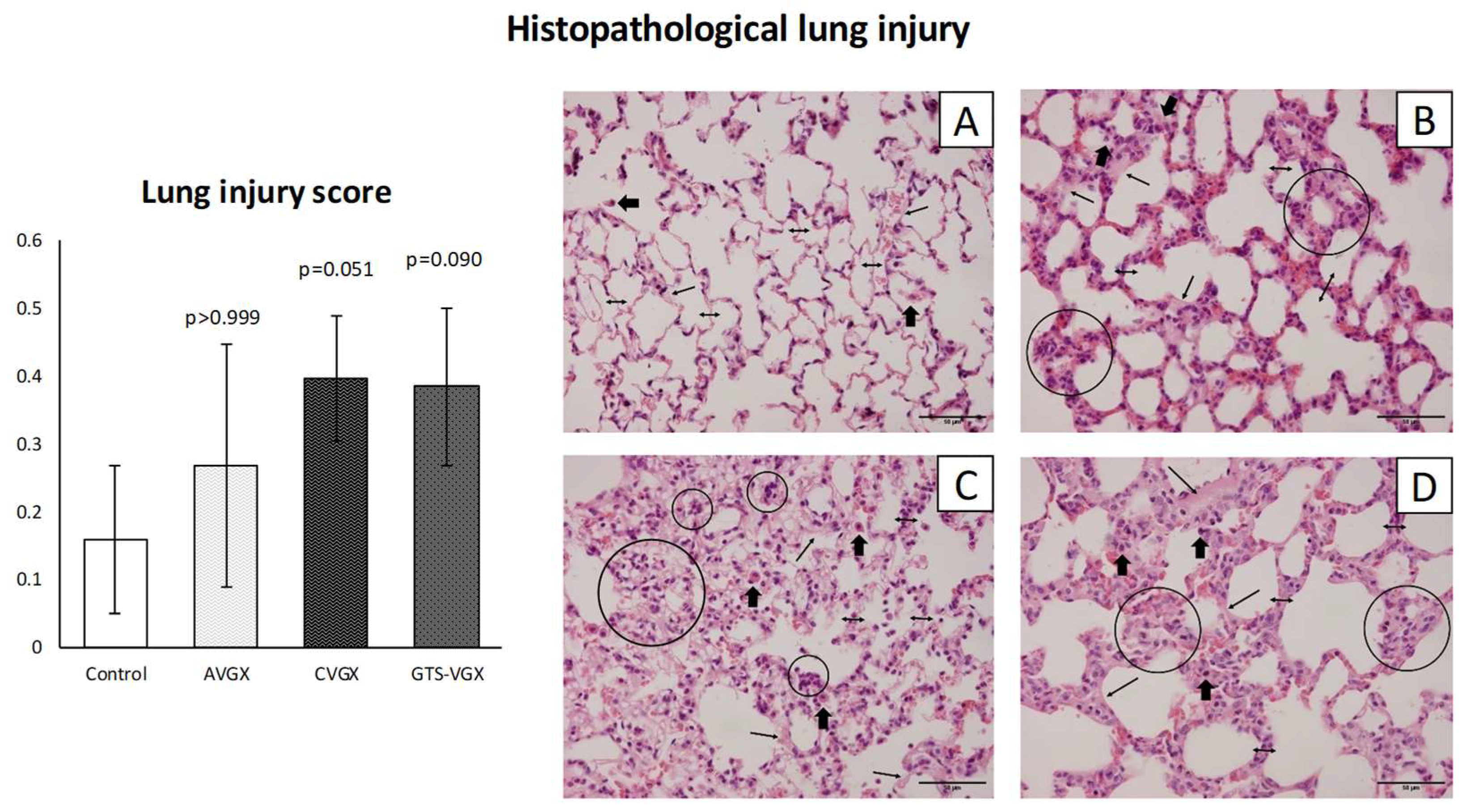
2.7. Lung Histology
2.8. ELISA
2.9. Statistics
3. Results
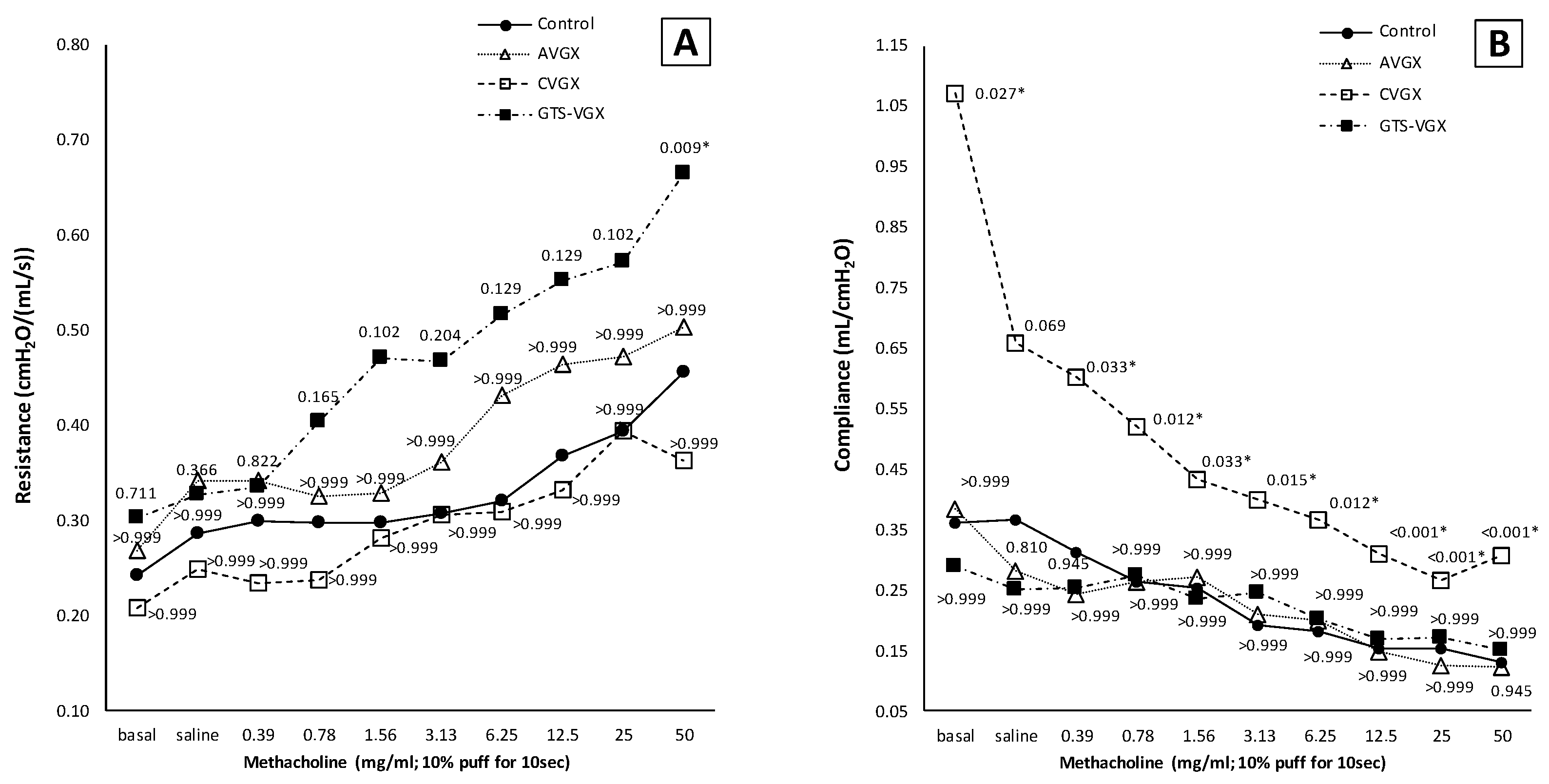
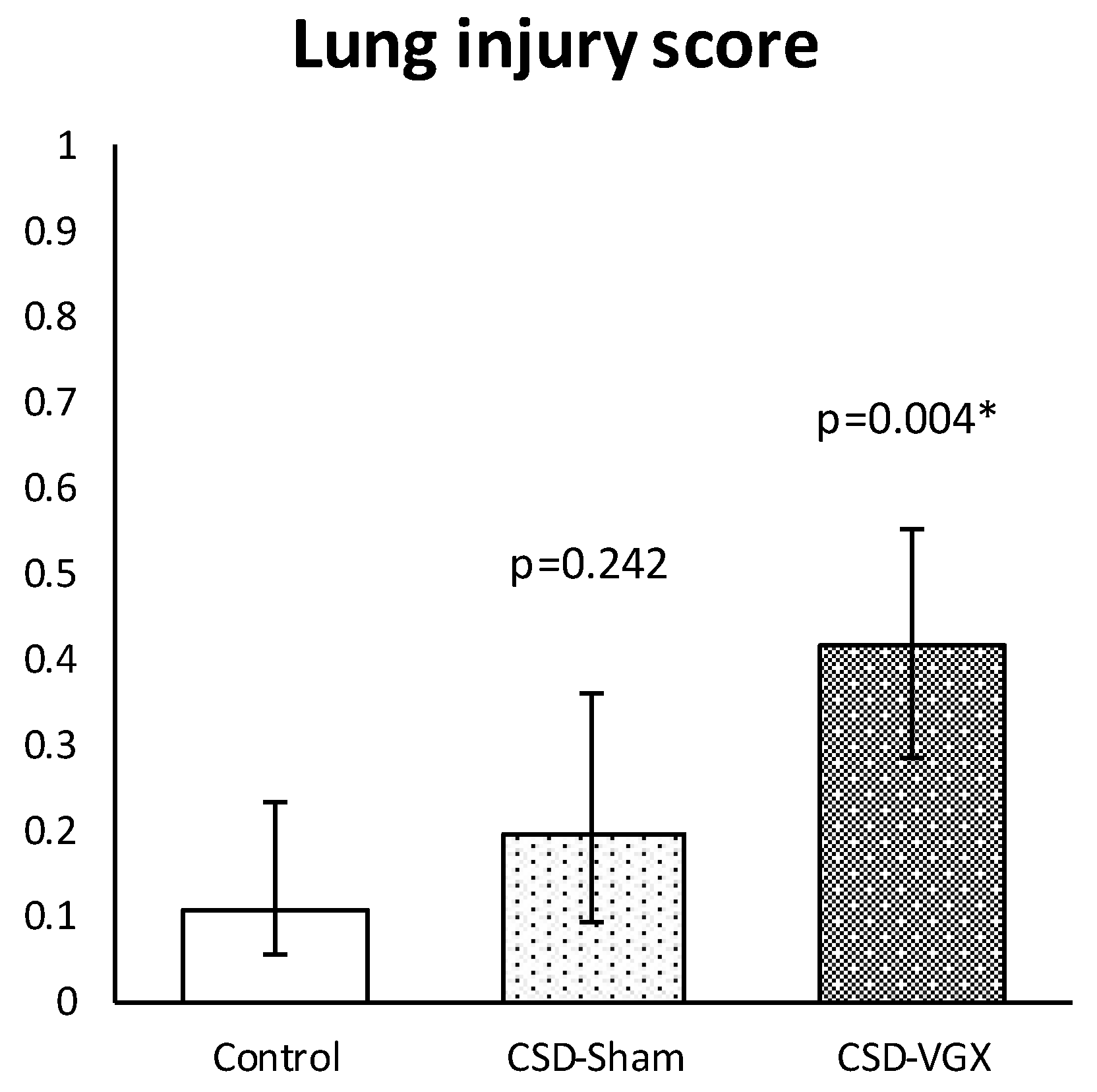
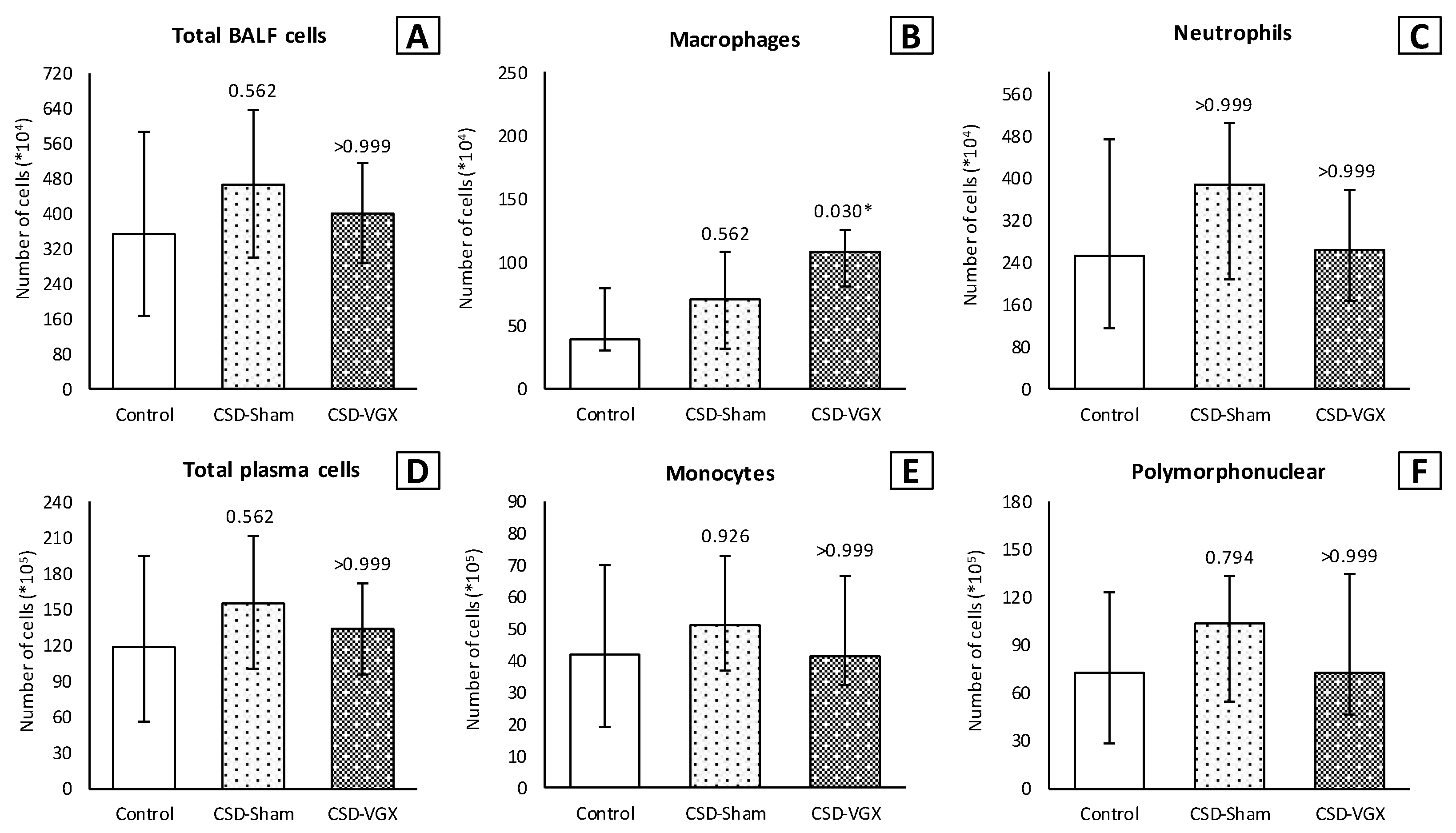
3.1. Model A: Selectively Sparing Vagus Nerve
3.2. Model B: Role of the α7nACh Receptor
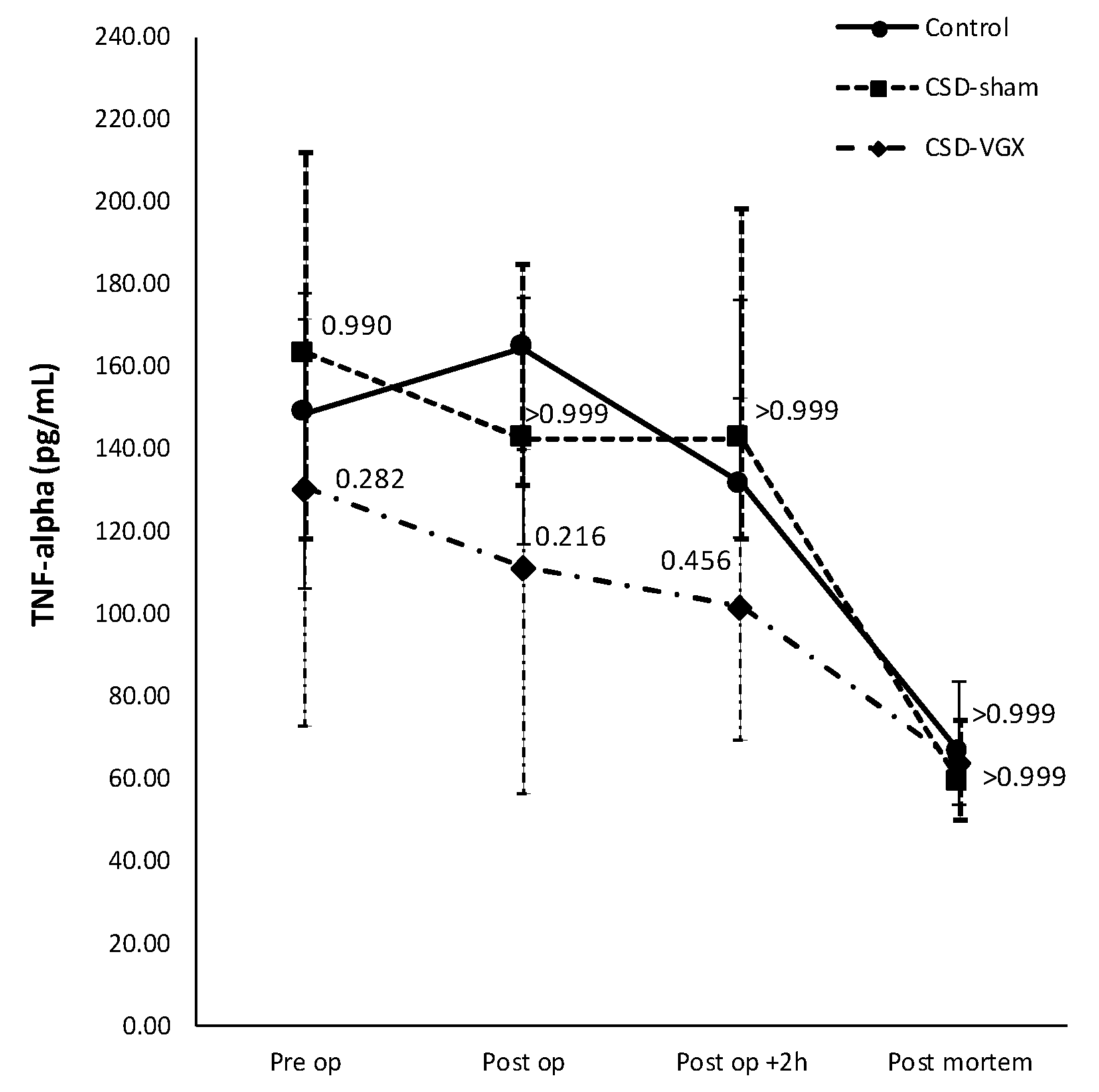
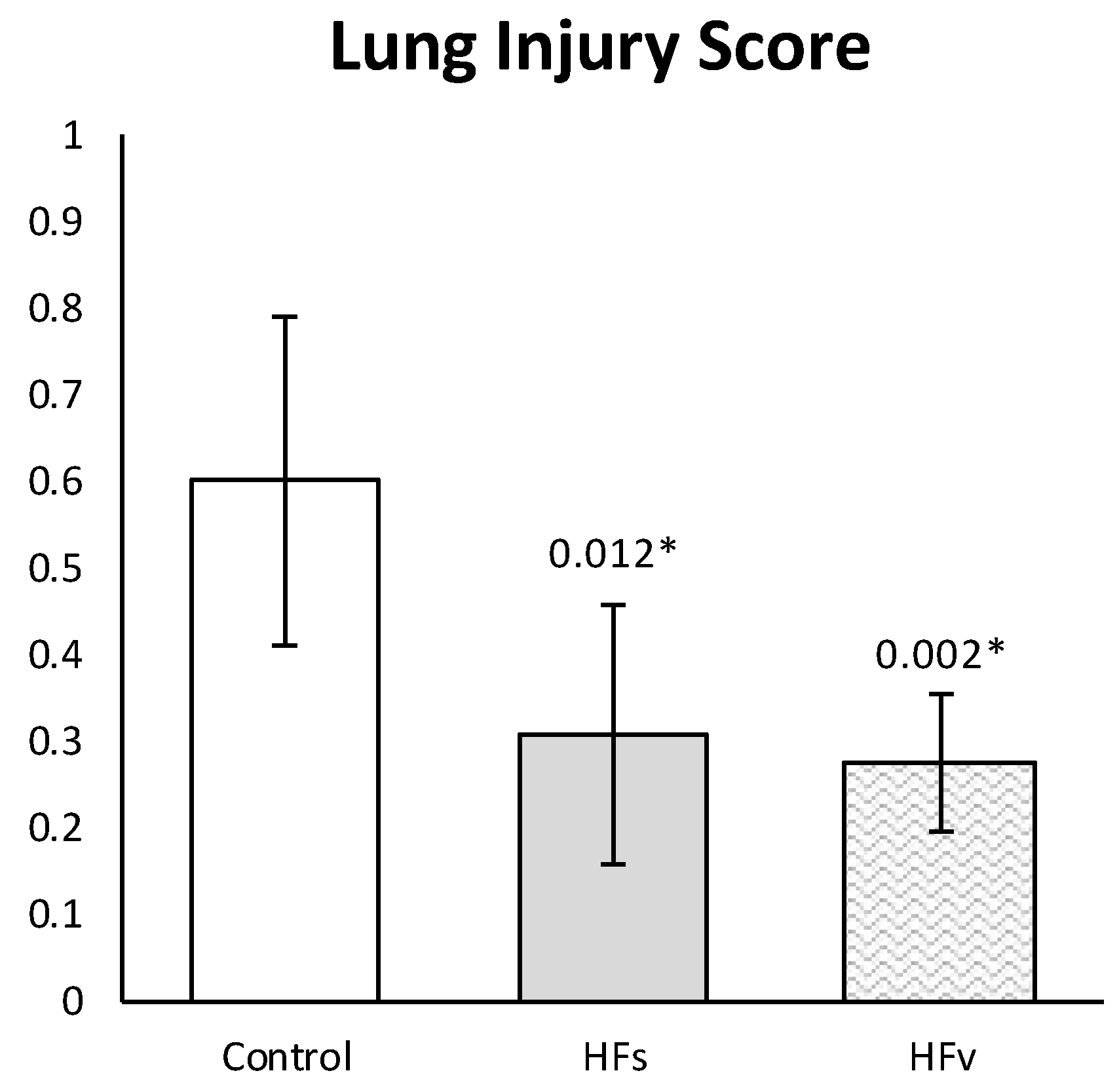
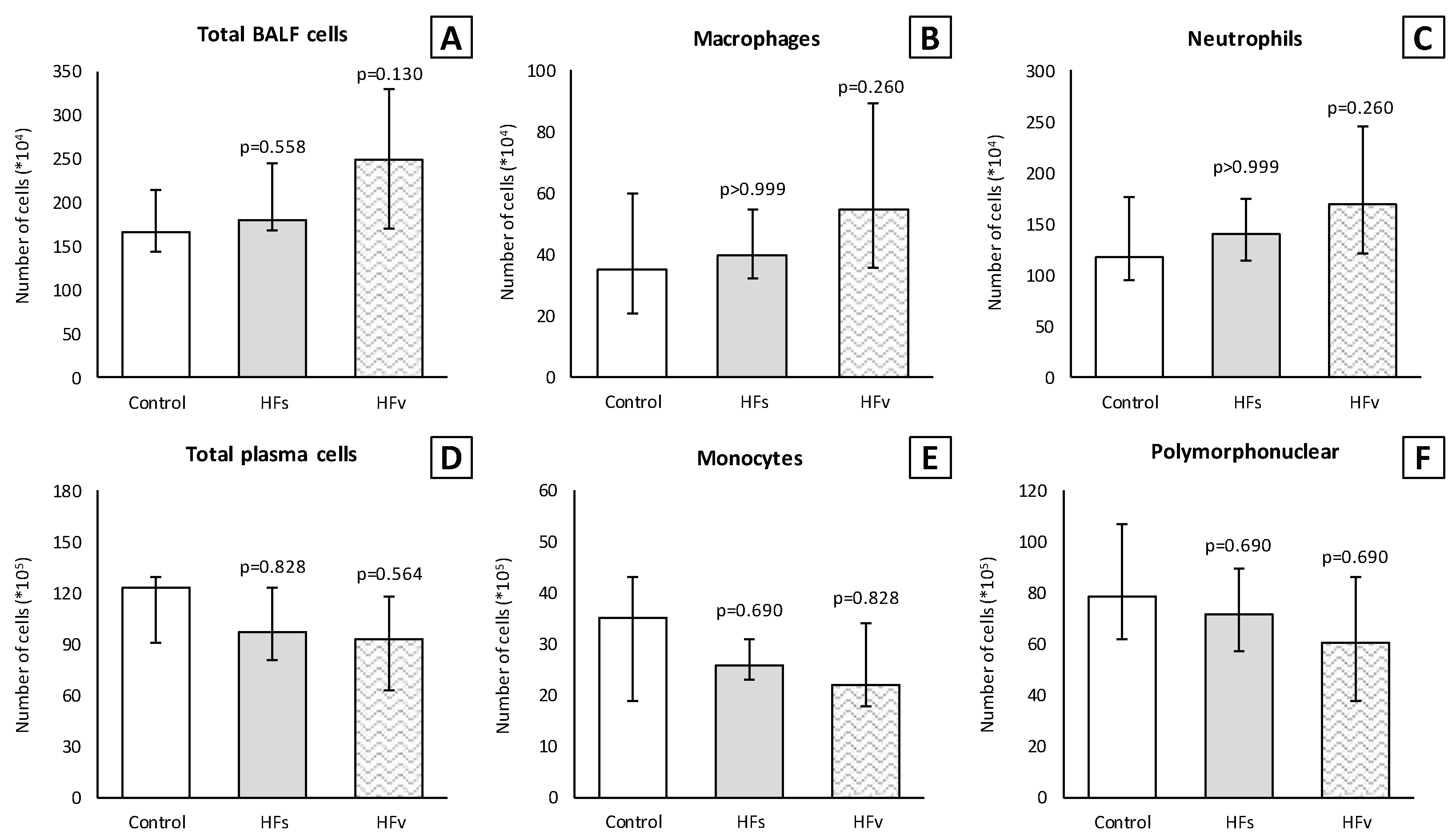
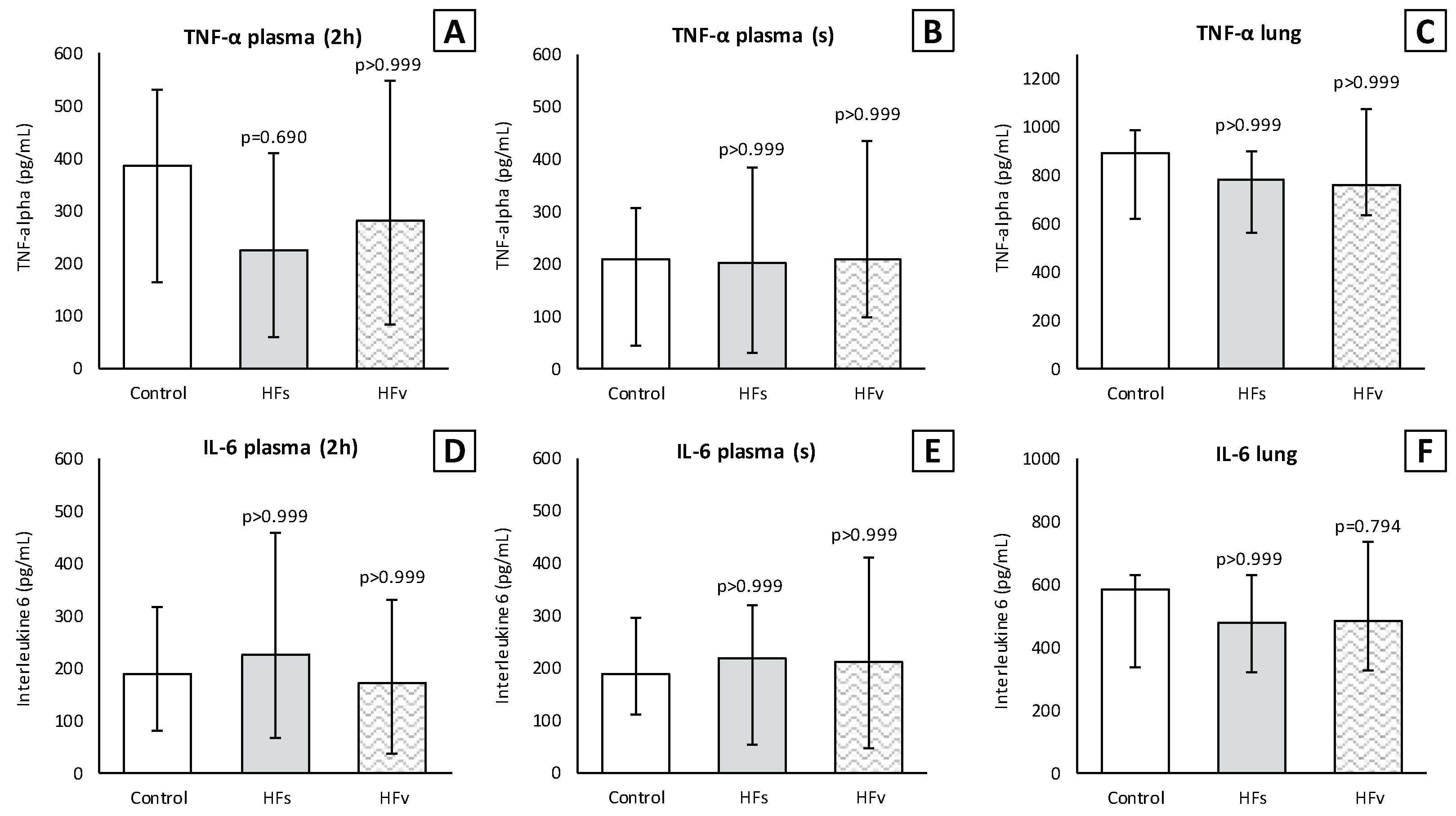
3.3. Model C: Effect of High-Fat Enteral Nutrition on LPS-Induced Lung Injury
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- De Jonge, W.J.; van der Zanden, E.P.; The, F.O.; Bijlsma, M.F.; van Westerloo, D.J.; Bennink, R.J.; Berthoud, H.R.; Uematsu, S.; Akira, S.; van den Wijngaard, R.M.; et al. Stimulation of the vagus nerve attenuates macrophage activation by activating the Jak2-STAT3 signaling pathway. Nat. Immunol. 2005, 6, 844–851. [Google Scholar] [CrossRef] [PubMed]
- Giebelen, I.A.; van Westerloo, D.J.; LaRosa, G.J.; de Vos, A.F.; van der Poll, T. Local stimulation of alpha7 cholinergic receptors inhibits LPS-induced TNF-alpha release in the mouse lung. Shock 2007, 28, 700–703. [Google Scholar] [CrossRef] [PubMed]
- Luyer, M.D.; Greve, J.W.; Hadfoune, M.; Jacobs, J.A.; Dejong, C.H.; Buurman, W.A. Nutritional stimulation of cholecystokinin receptors inhibits inflammation via the vagus nerve. J. Exp. Med. 2005, 202, 1023–1029. [Google Scholar] [CrossRef] [PubMed]
- Yamakawa, K.; Matsumoto, N.; Imamura, Y.; Muroya, T.; Yamada, T.; Nakagawa, J.; Shimazaki, J.; Ogura, H.; Kuwagata, Y.; Shimazu, T. Electrical vagus nerve stimulation attenuates systemic inflammation and improves survival in a rat heatstroke model. PLoS ONE 2013, 8, e56728. [Google Scholar] [CrossRef]
- Chen, C.; Zhang, Y.; Du, Z.; Zhang, M.; Niu, L.; Wang, Y.; Li, J. Vagal efferent fiber stimulation ameliorates pulmonary microvascular endothelial cell injury by downregulating inflammatory responses. Inflammation 2013, 36, 1567–1575. [Google Scholar] [CrossRef]
- Reys, L.G.; Ortiz-Pomales, Y.T.; Lopez, N.; Cheadle, G.; de Oliveira, P.G.; Eliceiri, B.; Bansal, V.; Costantini, T.W.; Coimbra, R. Uncovering the neuroenteric-pulmonary axis: Vagal nerve stimulation prevents acute lung injury following hemorrhagic shock. Life Sci. 2013, 92, 783–792. [Google Scholar] [CrossRef]
- Pavlov, V.A.; Wang, H.; Czura, C.J.; Friedman, S.G.; Tracey, K.J. The cholinergic anti-inflammatory pathway: A missing link in neuroimmunomodulation. Mol. Med. 2003, 9, 125–134. [Google Scholar] [CrossRef]
- Pavlov, V.A.; Tracey, K.J. The vagus nerve and the inflammatory reflex--linking immunity and metabolism. Nat. Rev. Endocrinol. 2012, 8, 743–754. [Google Scholar] [CrossRef]
- Matteoli, G.; Gomez-Pinilla, P.J.; Nemethova, A.; Di Giovangiulio, M.; Cailotto, C.; van Bree, S.H.; Michel, K.; Tracey, K.J.; Schemann, M.; Boesmans, W.; et al. A distinct vagal anti-inflammatory pathway modulates intestinal muscularis resident macrophages independent of the spleen. Gut 2014, 63, 938–948. [Google Scholar] [CrossRef]
- Wu, H.; Li, L.; Su, X. Vagus nerve through alpha7 nAChR modulates lung infection and inflammation: Models, cells, and signals. Biomed. Res. Int. 2014, 2014, 283525. [Google Scholar] [CrossRef]
- Costello, R.W.; Jacoby, D.B.; Fryer, A.D. Pulmonary neuronal M2 muscarinic receptor function in asthma and animal models of hyperreactivity. Thorax 1998, 53, 613–616. [Google Scholar] [CrossRef] [PubMed]
- Mazzone, S.B.; Undem, B.J. Vagal Afferent Innervation of the Airways in Health and Disease. Physiol. Rev. 2016, 96, 975–1024. [Google Scholar] [CrossRef] [PubMed]
- Weijs, T.J.; Ruurda, J.P.; Luyer, M.D.; Nieuwenhuijzen, G.A.; van der Horst, S.; Bleys, R.L.; van Hillegersberg, R. Preserving the pulmonary vagus nerve branches during thoracoscopic esophagectomy. Surg. Endosc. 2016, 30, 3816–3822. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Zhang, J.F.; Roberts, A.M.; Collins, L.C.; Fletcher, E.C. Pulmonary rapidly adapting receptor stimulation does not increase airway resistance in anesthetized rabbits. Am. J. Respir. Crit. Care Med. 1999, 160, 906–912. [Google Scholar] [CrossRef]
- Kageyama-Yahara, N.; Suehiro, Y.; Yamamoto, T.; Kadowaki, M. IgE-induced degranulation of mucosal mast cells is negatively regulated via nicotinic acetylcholine receptors. Biochem. Biophys. Res. Commun. 2008, 377, 321–325. [Google Scholar] [CrossRef] [PubMed]
- Fox, A.J.; Barnes, P.J.; Urban, L.; Dray, A. An in vitro study of the properties of single vagal afferents innervating guinea-pig airways. J. Physiol. 1993, 469, 21–35. [Google Scholar] [CrossRef] [PubMed]
- Borovikova, L.V.; Ivanova, S.; Zhang, M.; Yang, H.; Botchkina, G.I.; Watkins, L.R.; Wang, H.; Abumrad, N.; Eaton, J.W.; Tracey, K.J. Vagus nerve stimulation attenuates the systemic inflammatory response to endotoxin. Nature 2000, 405, 458–462. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, F.; Coles, S.K.; McCrimmon, D.R. Respiratory neurons mediating the Breuer-Hering reflex prolongation of expiration in rat. J. Neurosci. 1996, 16, 6526–6536. [Google Scholar] [CrossRef]
- Pack, A.I.; Ogilvie, M.D.; Davies, R.O.; Galante, R.J. Responses of pulmonary stretch receptors during ramp inflations of the lung. J. Appl. Physiol. 1986, 61, 344–352. [Google Scholar] [CrossRef]
- Cong, B.; Li, S.J.; Ling, Y.L.; Yao, Y.X.; Gu, Z.Y.; Wang, J.X.; You, H.Y. Expression and cell-specific localization of cholecystokinin receptors in rat lung. World J. Gastroenterol. 2003, 9, 1273–1277. [Google Scholar] [CrossRef]
- Yu, J.; Lin, S.; Zhang, J.; Otmishi, P.; Guardiola, J.J. Airway nociceptors activated by pro-inflammatory cytokines. Respir. Physiol. Neurobiol. 2007, 156, 116–119. [Google Scholar] [CrossRef] [PubMed]
- Voeten, D.M.; Busweiler, L.A.D.; van der Werf, L.R.; Wijnhoven, B.P.L.; Verhoeven, R.H.A.; van Sandick, J.W.; van Hillegersberg, R.; van Berge Henegouwen, M.I.; Dutch Upper Gastrointestinal Cancer Audit, G. Outcomes of Esophagogastric Cancer Surgery During Eight Years of Surgical Auditing by the Dutch Upper Gastrointestinal Cancer Audit (DUCA). Ann. Surg. 2021, 274, 866–873. [Google Scholar] [CrossRef] [PubMed]
- Muller-Stich, B.P.; Probst, P.; Nienhuser, H.; Fazeli, S.; Senft, J.; Kalkum, E.; Heger, P.; Warschkow, R.; Nickel, F.; Billeter, A.T.; et al. Meta-analysis of randomized controlled trials and individual patient data comparing minimally invasive with open oesophagectomy for cancer. Br. J. Surg. 2021, 108, 1026–1033. [Google Scholar] [CrossRef]
- Janssen, T.; Fransen, L.F.C.; Heesakkers, F.; Dolmans-Zwartjes, A.C.P.; Moorthy, K.; Nieuwenhuijzen, G.A.P.; Luyer, M.D.P. Effect of a multimodal prehabilitation program on postoperative recovery and morbidity in patients undergoing a totally minimally invasive esophagectomy. Dis. Esophagus 2021, 35, doab082. [Google Scholar] [CrossRef] [PubMed]
- Fransen, L.F.C.; Janssen, T.; Aarnoudse, M.; Nieuwenhuijzen, G.A.P.; Luyer, M.D.P. Direct Oral Feeding After a Minimally Invasive Esophagectomy: A Single-Center Prospective Cohort Study. Ann. Surg. 2022, 275, 919–923. [Google Scholar] [CrossRef]
- Puccetti, F.; Wijnhoven, B.P.L.; Kuppusamy, M.; Hubka, M.; Low, D.E. Impact of standardized clinical pathways on esophagectomy: A systematic review and meta-analysis. Dis. Esophagus 2022, 35, doab027. [Google Scholar] [CrossRef]
- Okamura, A.; Takeuchi, H.; Matsuda, S.; Ogura, M.; Miyasho, T.; Nakamura, R.; Takahashi, T.; Wada, N.; Kawakubo, H.; Saikawa, Y.; et al. Factors affecting cytokine change after esophagectomy for esophageal cancer. Ann. Surg. Oncol. 2015, 22, 3130–3135. [Google Scholar] [CrossRef]
- Shinozaki, H.; Matsuoka, T.; Ozawa, S. Pharmacological treatment to reduce pulmonary morbidity after esophagectomy. Ann. Gastroenterol. Surg. 2021, 5, 614–622. [Google Scholar] [CrossRef]
- D’Journo, X.B.; Michelet, P.; Marin, V.; Diesnis, I.; Blayac, D.; Doddoli, C.; Bongrand, P.; Thomas, P.A. An early inflammatory response to oesophagectomy predicts the occurrence of pulmonary complications. Eur. J. Cardiothorac. Surg. 2010, 37, 1144–1151. [Google Scholar] [CrossRef]
- Markar, S.; Gronnier, C.; Duhamel, A.; Bigourdan, J.M.; Badic, B.; du Rieu, M.C.; Lefevre, J.H.; Turner, K.; Luc, G.; Mariette, C. Pattern of Postoperative Mortality After Esophageal Cancer Resection According to Center Volume: Results from a Large European Multicenter Study. Ann. Surg. Oncol. 2015, 22, 2615–2623. [Google Scholar] [CrossRef]
- Seesing, M.F.J.; Kingma, B.F.; Weijs, T.J.; Ruurda, J.P.; van Hillegersberg, R. Reducing pulmonary complications after esophagectomy for cancer. J. Thorac. Dis. 2019, 11, S794–S798. [Google Scholar] [CrossRef] [PubMed]
- Weijs, T.J.; Ruurda, J.P.; Luyer, M.D.; Nieuwenhuijzen, G.A.; van Hillegersberg, R.; Bleys, R.L. Topography and extent of pulmonary vagus nerve supply with respect to transthoracic oesophagectomy. J. Anat. 2015, 227, 431–439. [Google Scholar] [CrossRef] [PubMed]
- Matute-Bello, G.; Downey, G.; Moore, B.B.; Groshong, S.D.; Matthay, M.A.; Slutsky, A.S.; Kuebler, W.M.; Acute Lung Injury in Animals Study, G. An official American Thoracic Society workshop report: Features and measurements of experimental acute lung injury in animals. Am. J. Respir. Cell Mol. Biol. 2011, 44, 725–738. [Google Scholar] [CrossRef] [PubMed]
- Pavlov, V.A.; Tracey, K.J. Neural circuitry and immunity. Immunol. Res. 2015, 63, 38–57. [Google Scholar] [CrossRef]
- Yuan, H.; Silberstein, S.D. Vagus Nerve and Vagus Nerve Stimulation, a Comprehensive Review: Part I. Headache 2016, 56, 71–78. [Google Scholar] [CrossRef]
- Hansen, M.K.; Nguyen, K.T.; Fleshner, M.; Goehler, L.E.; Gaykema, R.P.; Maier, S.F.; Watkins, L.R. Effects of vagotomy on serum endotoxin, cytokines, and corticosterone after intraperitoneal lipopolysaccharide. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2000, 278, R331–R336. [Google Scholar] [CrossRef]
- Kox, M.; Vaneker, M.; van der Hoeven, J.G.; Scheffer, G.J.; Hoedemaekers, C.W.; Pickkers, P. Effects of vagus nerve stimulation and vagotomy on systemic and pulmonary inflammation in a two-hit model in rats. PLoS ONE 2012, 7, e34431. [Google Scholar] [CrossRef]
- Van Westerloo, D.J.; Giebelen, I.A.; Florquin, S.; Daalhuisen, J.; Bruno, M.J.; de Vos, A.F.; Tracey, K.J.; van der Poll, T. The cholinergic anti-inflammatory pathway regulates the host response during septic peritonitis. J. Infect. Dis. 2005, 191, 2138–2148. [Google Scholar] [CrossRef]
- Pack, A.I.; DeLaney, R.G.; Fishman, A.P. Augmentation of phrenic neural activity by increased rates of lung inflation. J. Appl. Physiol. Respir. Environ. Exerc. Physiol. 1981, 50, 149–161. [Google Scholar] [CrossRef]
- Rieger-Fackeldey, E.; Sindelar, R.; Sedin, G.; Jonzon, A. Bronchopulmonary C-fibers modulate the breathing pattern in surfactant-depleted juvenile cats. Respir. Physiol. Neurobiol. 2008, 160, 341–349. [Google Scholar] [CrossRef]
- Malik, A.B. Mechanisms of neurogenic pulmonary edema. Circ. Res. 1985, 57, 1–18. [Google Scholar] [CrossRef]
- Garg, B.K.; Loring, R.H. GTS-21 has cell-specific anti-inflammatory effects independent of alpha7 nicotinic acetylcholine receptors. PLoS ONE 2019, 14, e0214942. [Google Scholar] [CrossRef] [PubMed]
- Giebelen, I.A.; van Westerloo, D.J.; LaRosa, G.J.; de Vos, A.F.; van der Poll, T. Stimulation of alpha 7 cholinergic receptors inhibits lipopolysaccharide-induced neutrophil recruitment by a tumor necrosis factor alpha-independent mechanism. Shock 2007, 27, 443–447. [Google Scholar] [CrossRef]
- Wang, J.; Li, R.; Peng, Z.; Zhou, W.; Hu, B.; Rao, X.; Yang, X.; Li, J. GTS-21 Reduces Inflammation in Acute Lung Injury by Regulating M1 Polarization and Function of Alveolar Macrophages. Shock 2019, 51, 389–400. [Google Scholar] [CrossRef] [PubMed]
- De Haan, J.J.; Lubbers, T.; Hadfoune, M.; Luyer, M.D.; Dejong, C.H.; Buurman, W.A.; Greve, J.W. Postshock intervention with high-lipid enteral nutrition reduces inflammation and tissue damage. Ann. Surg. 2008, 248, 842–848. [Google Scholar] [CrossRef]
- Luyer, M.D.; Jacobs, J.A.; Vreugdenhil, A.C.; Hadfoune, M.; Dejong, C.H.; Buurman, W.A.; Greve, J.W. Enteral administration of high-fat nutrition before and directly after hemorrhagic shock reduces endotoxemia and bacterial translocation. Ann. Surg. 2004, 239, 257–264. [Google Scholar] [CrossRef] [PubMed]
- Luyer, M.D.; Derikx, J.P.; Beyaert, R.; Hadfoune, M.; van Kuppevelt, T.H.; Dejong, C.H.; Heineman, E.; Buurman, W.A.; Greve, J.W. High-fat nutrition reduces hepatic damage following exposure to bacterial DNA and hemorrhagic shock. J. Hepatol. 2009, 50, 342–350. [Google Scholar] [CrossRef]
- Lubbers, T.; de Haan, J.J.; Luyer, M.D.; Verbaeys, I.; Hadfoune, M.; Dejong, C.H.; Buurman, W.A.; Greve, J.W. Cholecystokinin/Cholecystokinin-1 receptor-mediated peripheral activation of the afferent vagus by enteral nutrients attenuates inflammation in rats. Ann. Surg. 2010, 252, 376–382. [Google Scholar] [CrossRef] [PubMed]
- Lubbers, T.; De Haan, J.J.; Hadfoune, M.; Zhang, Y.; Luyer, M.D.; Grundy, D.; Buurman, W.A.; Greve, J.W. Lipid-enriched enteral nutrition controls the inflammatory response in murine Gram-negative sepsis. Crit. Care Med. 2010, 38, 1996–2002. [Google Scholar] [CrossRef] [PubMed]
- Ling, Y.L.; Meng, A.H.; Zhao, X.Y.; Shan, B.E.; Zhang, J.L.; Zhang, X.P. Effect of cholecystokinin on cytokines during endotoxic shock in rats. World J. Gastroenterol. 2001, 7, 667–671. [Google Scholar] [CrossRef]
- Meng, A.H.; Ling, Y.L.; Zhang, X.P.; Zhang, J.L. Anti-inflammatory effect of cholecystokinin and its signal transduction mechanism in endotoxic shock rat. World J. Gastroenterol. 2002, 8, 712–717. [Google Scholar] [CrossRef] [PubMed]
- Tian, F.; Ling, Y.; Chen, Y.; Wang, Z. Effects of CCK-8 and Cystathionine gamma-Lyase/Hydrogen Sulfide System on Acute Lung Injury in Rats. Inflammation 2017, 40, 174–183. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Ni, Z.; Cong, B.; Gao, W.; Xu, S.; Wang, C.; Yao, Y.; Ma, C.; Ling, Y. CCK-8 inhibits LPS-induced IL-1beta production in pulmonary interstitial macrophages by modulating PKA, p38, and NF-kappaB pathway. Shock 2007, 27, 678–686. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.J.; Gao, W.J.; Cong, B.; Yao, Y.X.; Gu, Z.Y. Effect of lipopolysaccharide on expression and characterization of cholecystokinin receptors in rat pulmonary interstitial macrophages. Acta Pharmacol. Sin. 2004, 25, 1347–1353. [Google Scholar]
- Monnikes, H.; Lauer, G.; Bauer, C.; Tebbe, J.; Zittel, T.T.; Arnold, R. Pathways of Fos expression in locus ceruleus, dorsal vagal complex, and PVN in response to intestinal lipid. Am. J. Physiol. 1997, 273, R2059–R2071. [Google Scholar] [CrossRef]
- Glatzle, J.; Kreis, M.E.; Kawano, K.; Raybould, H.E.; Zittel, T.T. Postprandial neuronal activation in the nucleus of the solitary tract is partly mediated by CCK-A receptors. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2001, 281, R222–R229. [Google Scholar] [CrossRef]
- Peters, E.G.; Smeets, B.J.J.; Nors, J.; Back, C.M.; Funder, J.A.; Sommer, T.; Laurberg, S.; Love, U.S.; Leclercq, W.K.G.; Slooter, G.D.; et al. Perioperative lipid-enriched enteral nutrition versus standard care in patients undergoing elective colorectal surgery (SANICS II): A multicentre, double-blind, randomised controlled trial. Lancet Gastroenterol. Hepatol. 2018, 3, 242–251. [Google Scholar] [CrossRef]











Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Seesing, M.F.J.; Janssen, H.J.B.; Geraedts, T.C.M.; Weijs, T.J.; van Ark, I.; Leusink-Muis, T.; Folkerts, G.; Garssen, J.; Ruurda, J.P.; Nieuwenhuijzen, G.A.P.; et al. Exploring the Modulatory Effect of High-Fat Nutrition on Lipopolysaccharide-Induced Acute Lung Injury in Vagotomized Rats and the Role of the Vagus Nerve. Nutrients 2023, 15, 2327. https://doi.org/10.3390/nu15102327
Seesing MFJ, Janssen HJB, Geraedts TCM, Weijs TJ, van Ark I, Leusink-Muis T, Folkerts G, Garssen J, Ruurda JP, Nieuwenhuijzen GAP, et al. Exploring the Modulatory Effect of High-Fat Nutrition on Lipopolysaccharide-Induced Acute Lung Injury in Vagotomized Rats and the Role of the Vagus Nerve. Nutrients. 2023; 15(10):2327. https://doi.org/10.3390/nu15102327
Chicago/Turabian StyleSeesing, Maarten F. J., Henricus J. B. Janssen, Tessa C. M. Geraedts, Teus J. Weijs, Ingrid van Ark, Thea Leusink-Muis, Gert Folkerts, Johan Garssen, Jelle P. Ruurda, Grard A. P. Nieuwenhuijzen, and et al. 2023. "Exploring the Modulatory Effect of High-Fat Nutrition on Lipopolysaccharide-Induced Acute Lung Injury in Vagotomized Rats and the Role of the Vagus Nerve" Nutrients 15, no. 10: 2327. https://doi.org/10.3390/nu15102327
APA StyleSeesing, M. F. J., Janssen, H. J. B., Geraedts, T. C. M., Weijs, T. J., van Ark, I., Leusink-Muis, T., Folkerts, G., Garssen, J., Ruurda, J. P., Nieuwenhuijzen, G. A. P., van Hillegersberg, R., & Luyer, M. D. P. (2023). Exploring the Modulatory Effect of High-Fat Nutrition on Lipopolysaccharide-Induced Acute Lung Injury in Vagotomized Rats and the Role of the Vagus Nerve. Nutrients, 15(10), 2327. https://doi.org/10.3390/nu15102327