
Procyanidin B2 Attenuates Nicotine-Induced Hepatocyte Pyroptosis through a PPARγ-Dependent Mechanism

 ,
,  and
and
Abstract
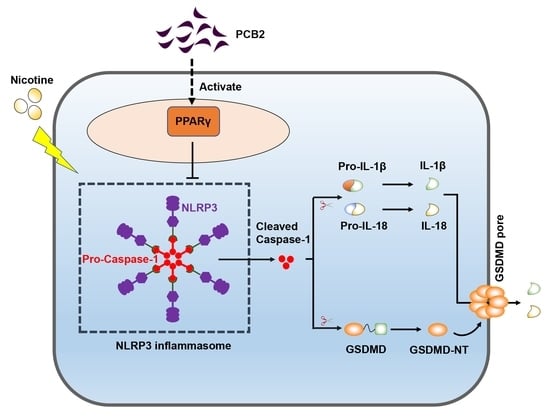
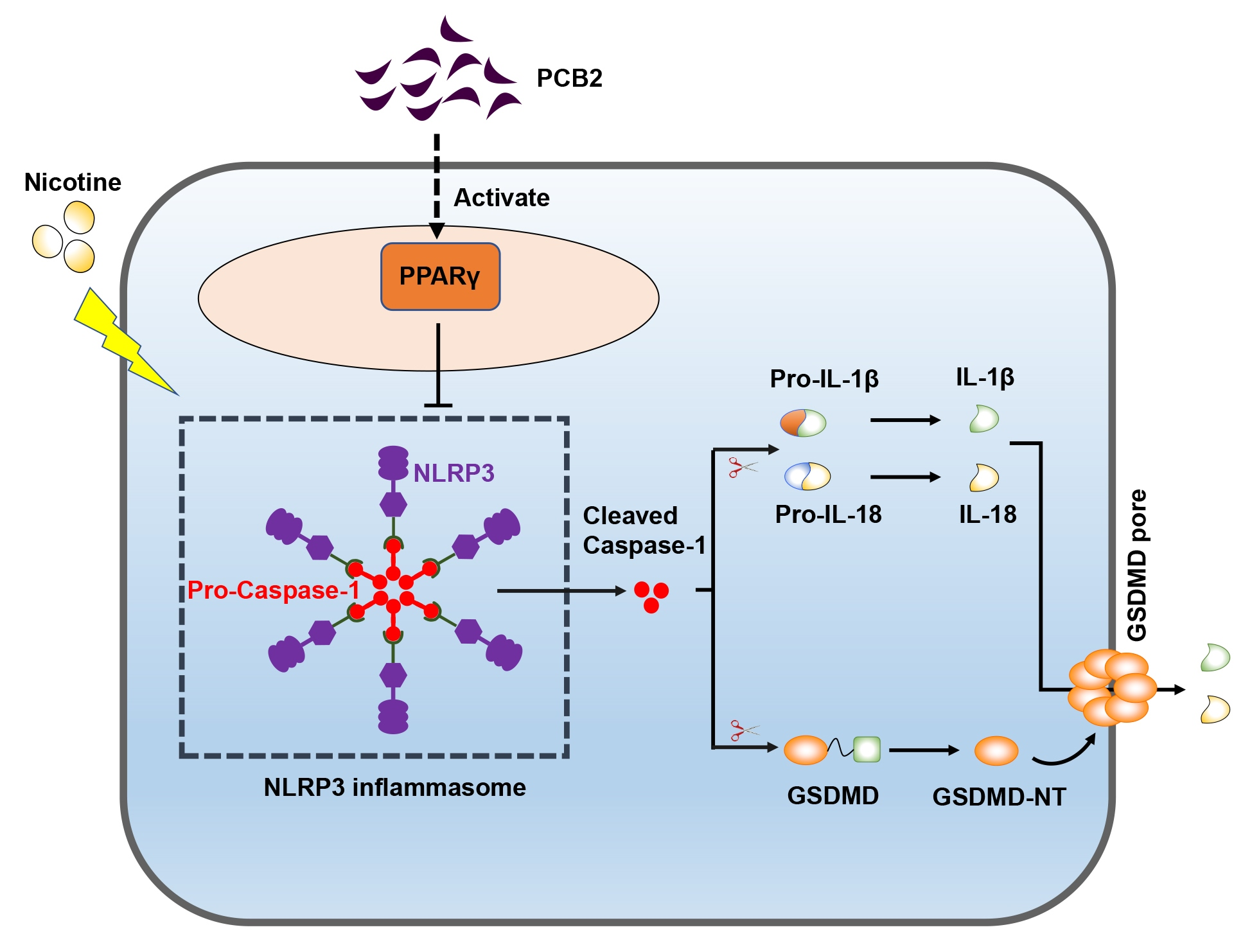
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Reagents, Antibodies and Plasmids
2.2. Animal Procedure
2.3. Cell Culture, Transfection and RNA Interference
2.4. Immunoblotting
2.5. Caspase-1 Activity Assay
2.6. LDH Release Assay
2.7. Cell Death Assay
2.8. Enzyme-Linked Immunosorbent Assay (ELISA)
2.9. Cell Viability Assay
2.10. Statistical Analysis
3. Results
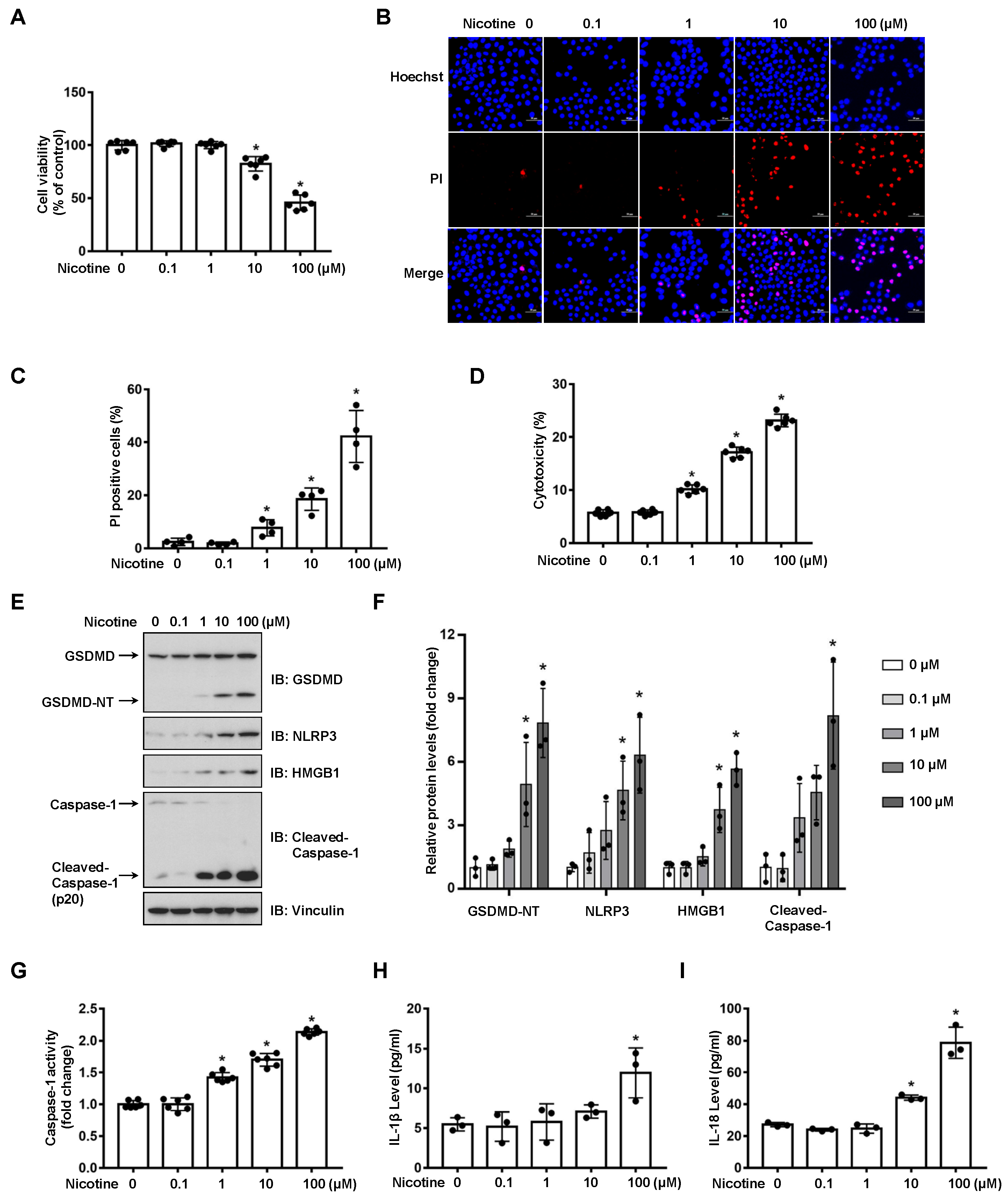
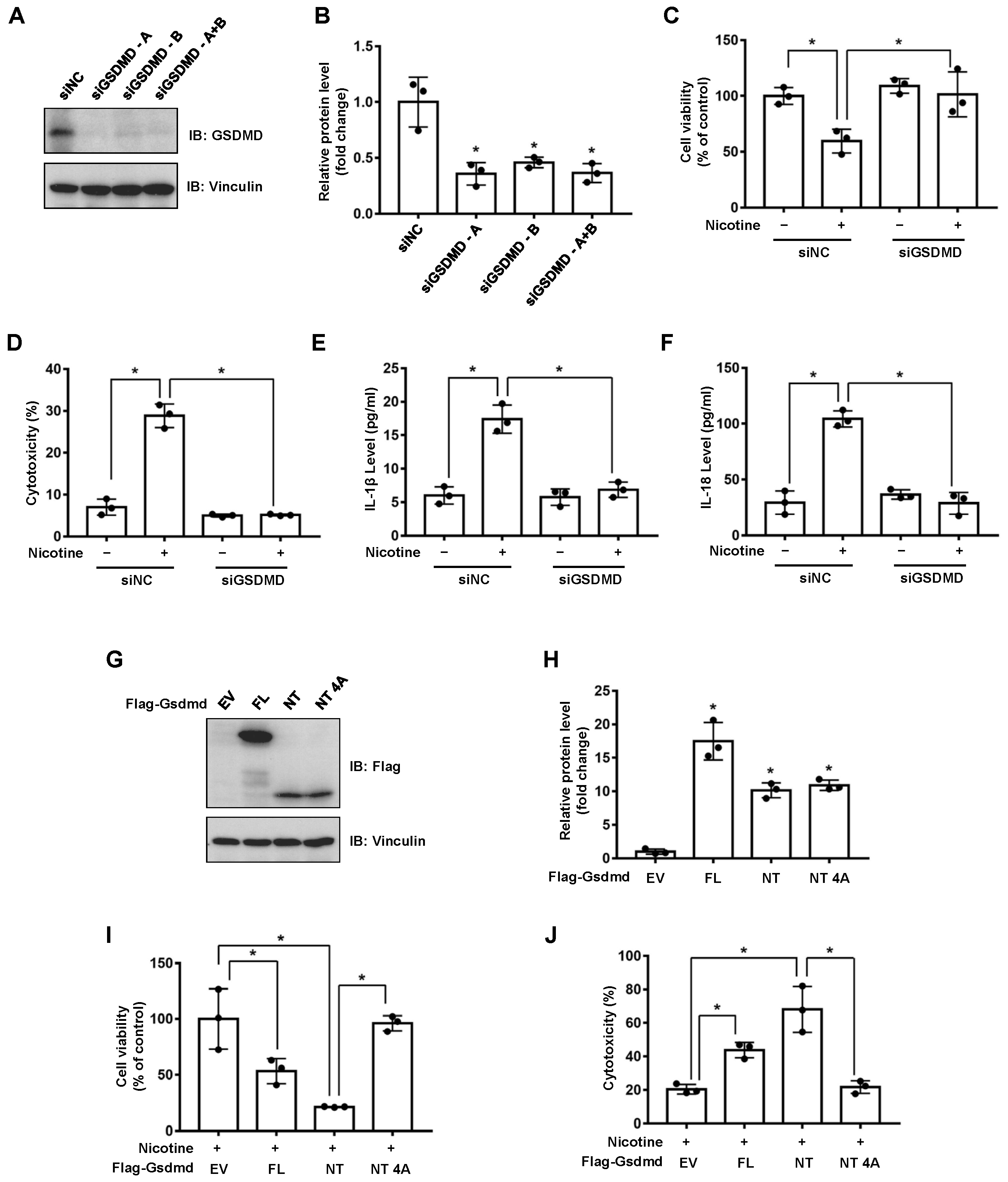
3.1. Nicotine Triggers Hepatocyte Pyroptosis through the Cleavage of GSDMD
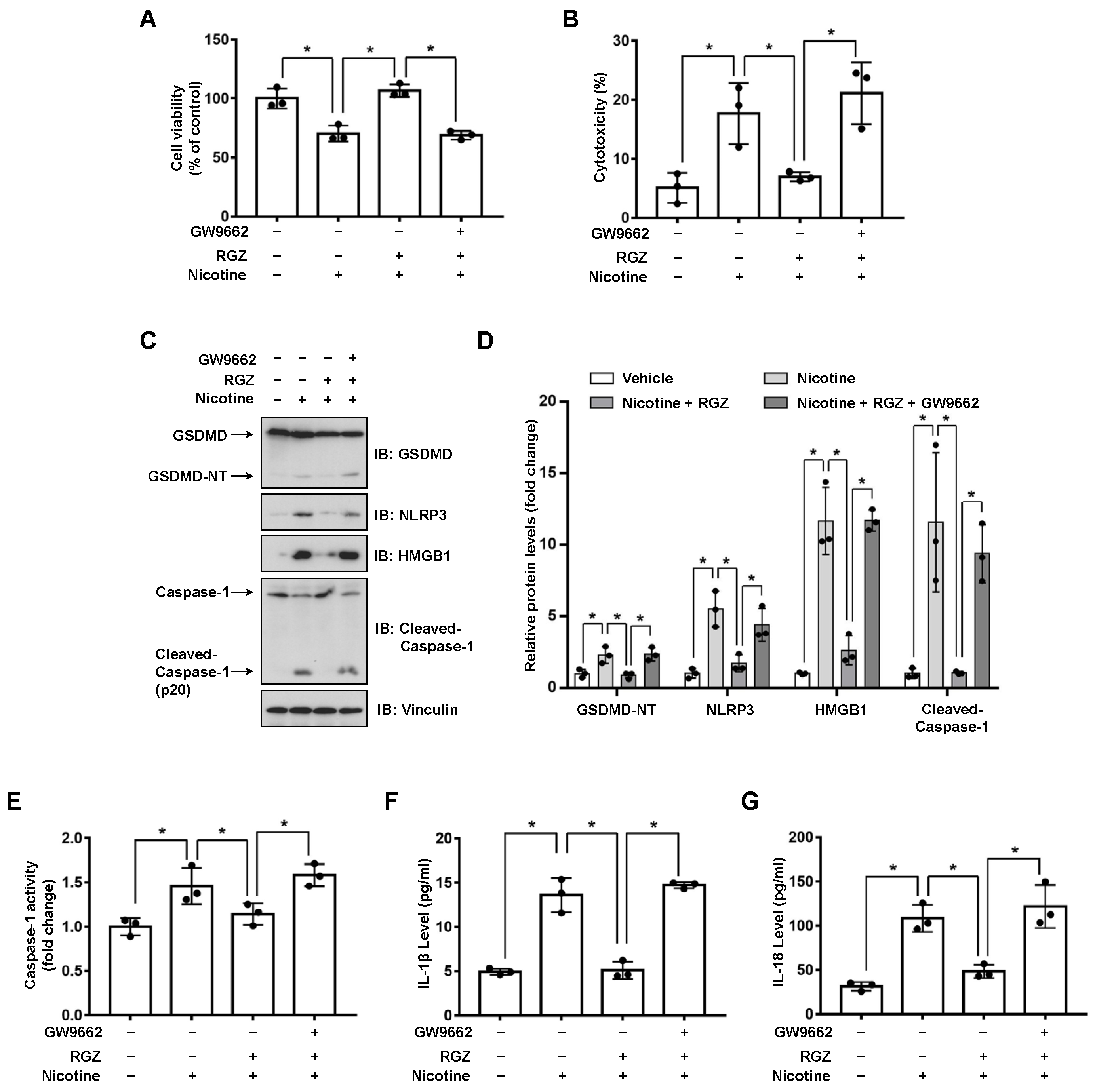
3.2. Activation of PPARγ Prevents Nicotine-Triggered Pyroptosis
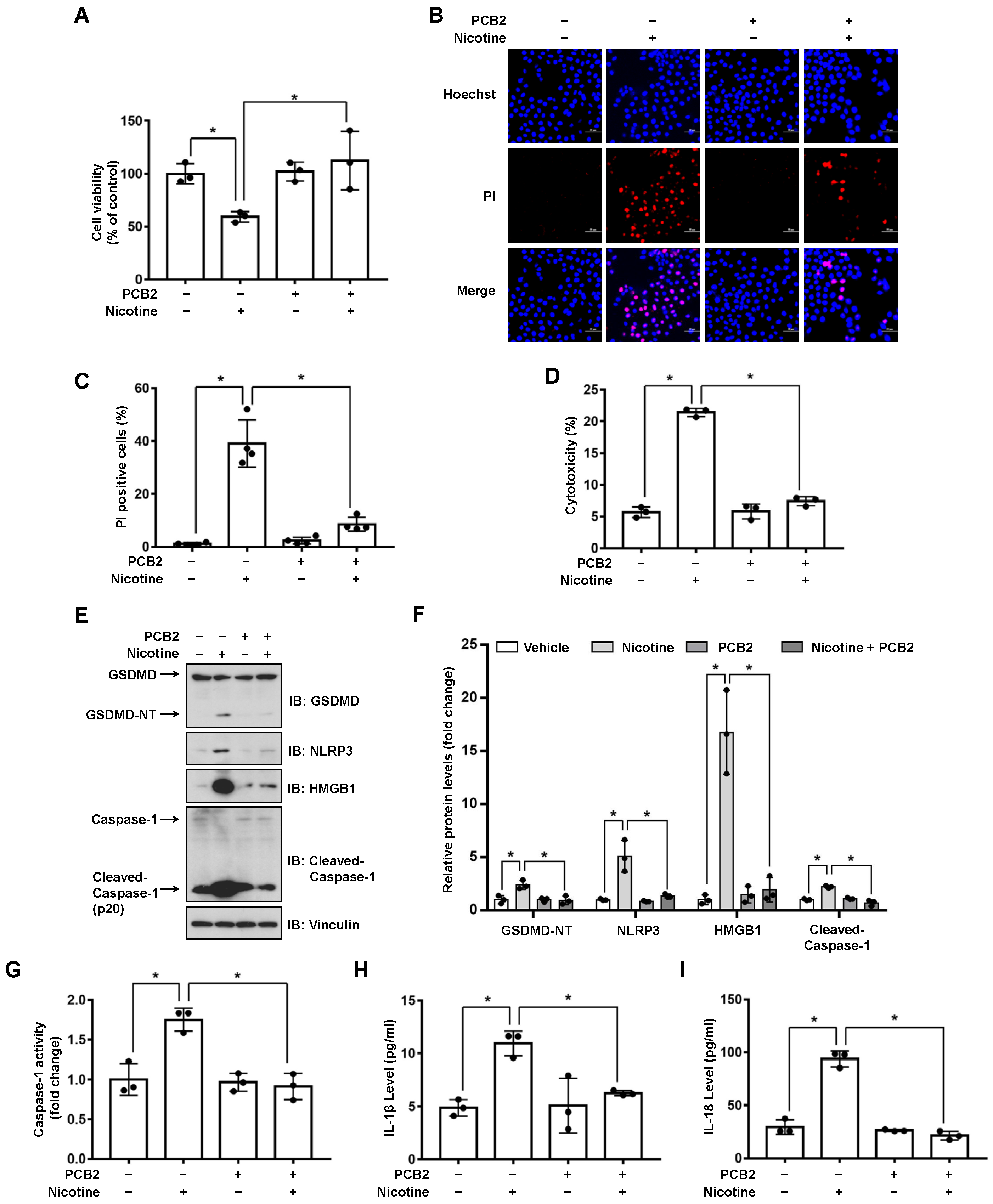
3.3. PCB2 Attenuates Nicotine-Induced Pyroptosis in Hepatocytes
3.4. PPARγ Antagonist Abrogates the Effects of PCB2 on Pyroptosis
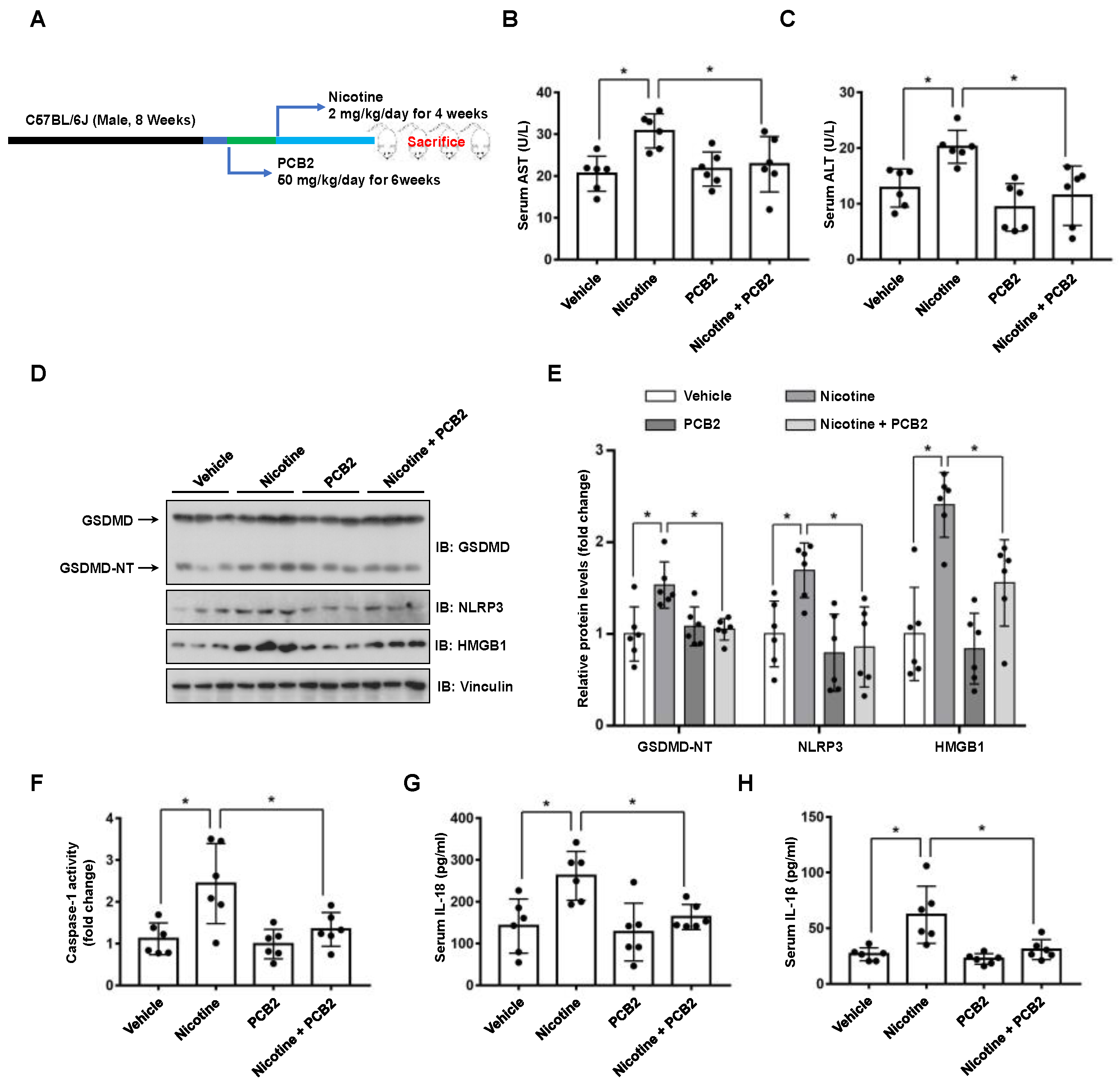
3.5. PCB2 Attenuates Liver Injury and Hepatocyte Pyroptosis in Mice
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Frieden, T.R. A Safer, Healthier U.S.: The Centers for Disease Control and Prevention, 2009–2016. Am. J. Prev. Med. 2017, 52, 263–275. [Google Scholar] [CrossRef] [Green Version]
- Long, G.A. Comparison of select analytes in exhaled aerosol from e-cigarettes with exhaled smoke from a conventional cigarette and exhaled breaths. Int. J. Environ. Res. Public Health 2014, 11, 11177–11191. [Google Scholar] [CrossRef]
- Hasan, K.M.; Friedman, T.C.; Shao, X.; Parveen, M.; Sims, C.; Lee, D.L.; Espinoza-Derout, J.; Sinha-Hikim, I.; Sinha-Hikim, A.P. E-cigarettes and Western Diet: Important Metabolic Risk Factors for Hepatic Diseases. Hepatology 2019, 69, 2442–2454. [Google Scholar] [CrossRef]
- Thorgeirsson, T.E.; Geller, F.; Sulem, P.; Rafnar, T.; Wiste, A.; Magnusson, K.P.; Manolescu, A.; Thorleifsson, G.; Stefansson, H.; Ingason, A.; et al. A variant associated with nicotine dependence, lung cancer and peripheral arterial disease. Nature 2008, 452, 638–642. [Google Scholar] [CrossRef] [Green Version]
- Zimmermann, T.; Hueppe, D.; Mauss, S.; Buggisch, P.; Pfeiffer-Vornkahl, H.; Grimm, D.; Galle, P.R.; Alshuth, U. Effects of Smoking on Pegylated Interferon alpha 2a and First Generation Protease Inhibitor-based Antiviral Therapy in Naive Patients Infected with Hepatitis C Virus Genotype 1. J. Gastrointest. Liver Dis. JGLD 2016, 25, 15–24. [Google Scholar] [CrossRef] [Green Version]
- Zein, C.O.; Unalp, A.; Colvin, R.; Liu, Y.C.; McCullough, A.J.; Nonalcoholic Steatohepatitis Clinical Research, N. Smoking and severity of hepatic fibrosis in nonalcoholic fatty liver disease. J. Hepatol. 2011, 54, 753–759. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.K.; Wu, M.; Kim, C.H.; Jin, Z.Y.; Zhou, J.Y.; Han, R.Q.; Yang, J.; Zhang, X.F.; Wang, X.S.; Liu, A.M.; et al. Jiangsu Four Cancers Study: A large case-control study of lung, liver, stomach, and esophageal cancers in Jiangsu Province, China. Eur. J. Cancer Prev. Off. J. Eur. Cancer Prev. Organ. 2017, 26, 357–364. [Google Scholar] [CrossRef]
- Karagozian, R.; Baker, E.; Houranieh, A.; Leavitt, D.; Baffy, G. Risk profile of hepatocellular carcinoma reveals dichotomy among US veterans. J. Gastrointest. Cancer 2013, 44, 318–324. [Google Scholar] [CrossRef]
- El-Zayadi, A.; Selim, O.; Hamdy, H.; El-Tawil, A.; Badran, H.M.; Attia, M.; Saeed, A. Impact of cigarette smoking on response to interferon therapy in chronic hepatitis C Egyptian patients. World J. Gastroenterol. 2004, 10, 2963–2966. [Google Scholar] [CrossRef]
- Bandyopadhyaya, G.; Sinha, S.; Chattopadhyay, B.D.; Chakraborty, A. Protective role of curcumin against nicotine-induced genotoxicity on rat liver under restricted dietary protein. Eur. J. Pharmacol. 2008, 588, 151–157. [Google Scholar] [CrossRef]
- El-Sokkary, G.H.; Cuzzocrea, S.; Reiter, R.J. Effect of chronic nicotine administration on the rat lung and liver: Beneficial role of melatonin. Toxicology 2007, 239, 60–67. [Google Scholar] [CrossRef] [PubMed]
- Khaled, S.; Makled, M.N.; Nader, M.A. Tiron protects against nicotine-induced lung and liver injury through antioxidant and anti-inflammatory actions in rats in vivo. Life Sci. 2020, 260, 118426. [Google Scholar] [CrossRef] [PubMed]
- Brennan, M.A.; Cookson, B.T. Salmonella induces macrophage death by caspase-1-dependent necrosis. Mol. Microbiol. 2000, 38, 31–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Broz, P.; Pelegrin, P.; Shao, F. The gasdermins, a protein family executing cell death and inflammation. Nat. Rev. Immunol. 2020, 20, 143–157. [Google Scholar] [CrossRef]
- Shi, J.; Gao, W.; Shao, F. Pyroptosis: Gasdermin-Mediated Programmed Necrotic Cell Death. Trends Biochem. Sci. 2017, 42, 245–254. [Google Scholar] [CrossRef]
- Wree, A.; Eguchi, A.; McGeough, M.D.; Pena, C.A.; Johnson, C.D.; Canbay, A.; Hoffman, H.M.; Feldstein, A.E. NLRP3 inflammasome activation results in hepatocyte pyroptosis, liver inflammation, and fibrosis in mice. Hepatology 2014, 59, 898–910. [Google Scholar] [CrossRef] [Green Version]
- Khanova, E.; Wu, R.; Wang, W.; Yan, R.; Chen, Y.; French, S.W.; Llorente, C.; Pan, S.Q.; Yang, Q.; Li, Y.; et al. Pyroptosis by caspase11/4-gasdermin-D pathway in alcoholic hepatitis in mice and patients. Hepatology 2018, 67, 1737–1753. [Google Scholar] [CrossRef] [Green Version]
- Xu, B.; Jiang, M.Z.; Chu, Y.; Wang, W.J.; Chen, D.; Li, X.W.; Zhang, Z.; Zhang, D.; Fan, D.M.; Nie, Y.Z.; et al. Gasdermin D plays a key role as a pyroptosis executor of non-alcoholic steatohepatitis in humans and mice. J. Hepatol. 2018, 68, 773–782. [Google Scholar] [CrossRef]
- Gaul, S.; Leszczynska, A.; Alegre, F.; Kaufmann, B.; Johnson, C.D.; Adams, L.A.; Wree, A.; Damm, G.; Seehofer, D.; Calvente, C.J.; et al. Hepatocyte pyroptosis and release of inflammasome particles induce stellate cell activation and liver fibrosis. J. Hepatol. 2021, 74, 156–167. [Google Scholar] [CrossRef]
- Xu, W.; Che, Y.; Zhang, Q.; Huang, H.; Ding, C.; Wang, Y.; Wang, G.; Cao, L.; Hao, H. Apaf-1 Pyroptosome Senses Mitochondrial Permeability Transition. Cell Metab. 2021, 33, 424–436.e410. [Google Scholar] [CrossRef]
- Mao, C.Y.; Li, D.J.; Zhou, E.; Zhang, J.F.; Wang, C.Q.; Xue, C. Nicotine exacerbates atherosclerosis through a macrophage-mediated endothelial injury pathway. Aging-US 2021, 13, 7627–7643. [Google Scholar] [CrossRef]
- Xu, S.; Chen, H.W.; Ni, H.E.; Dai, Q.Y. Targeting HDAC6 attenuates nicotine-induced macrophage pyroptosis via NF-kappa B/NLRP3 pathway. Atherosclerosis 2021, 317, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Zhang, H.; Qi, W.; Zhang, Y.; Li, J.; Li, Z.; Lin, Y.; Bai, X.; Liu, X.; Chen, X.; et al. Nicotine promotes atherosclerosis via ROS-NLRP3-mediated endothelial cell pyroptosis. Cell Death Dis. 2018, 9, 171. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Tian, X.; Yang, M.; Yu, Y.; Zhou, Y.; Gao, Y.; Zhang, L.; Li, Z.; Xiao, Y.; Moses, R.E.; et al. Procyanidin B2 Promotes Intestinal Injury Repair and Attenuates Colitis-Associated Tumorigenesis via Suppression of Oxidative Stress in Mice. Antioxid. Redox Signal. 2021, 35, 75–92. [Google Scholar] [CrossRef] [PubMed]
- Su, H.M.; Li, Y.T.; Hu, D.W.; Xie, L.H.; Ke, H.H.; Zheng, X.D.; Chen, W. Procyanidin B2 ameliorates free fatty acids-induced hepatic steatosis through regulating TFEB-mediated lysosomal pathway and redox state. Free Radic. Bio. Med. 2018, 126, 269–286. [Google Scholar] [CrossRef]
- Feng, J.; Wang, C.; Liu, T.; Li, J.; Wu, L.; Yu, Q.; Li, S.; Zhou, Y.; Zhang, J.; Chen, J.; et al. Procyanidin B2 inhibits the activation of hepatic stellate cells and angiogenesis via the Hedgehog pathway during liver fibrosis. J. Cell. Mol. Med. 2019, 23, 6479–6493. [Google Scholar] [CrossRef] [Green Version]
- Yang, B.Y.; Zhang, X.Y.; Guan, S.W.; Hua, Z.C. Protective Effect of Procyanidin B2 against CCl4-Induced Acute Liver Injury in Mice. Molecules 2015, 20, 12250–12265. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.; Xiao, L.; Yuan, Y.; Luo, X.; Jiang, M.; Ni, J.; Wang, N. Procyanidin B2 inhibits NLRP3 inflammasome activation in human vascular endothelial cells. Biochem. Pharmacol. 2014, 92, 599–606. [Google Scholar] [CrossRef]
- Frank, D.; Vince, J.E. Pyroptosis versus necroptosis: Similarities, differences, and crosstalk. Cell Death Differ. 2019, 26, 99–114. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, Z.; Ruan, J.; Pan, Y.; Magupalli, V.G.; Wu, H.; Lieberman, J. Inflammasome-activated gasdermin D causes pyroptosis by forming membrane pores. Nature 2016, 535, 153–158. [Google Scholar] [CrossRef] [Green Version]
- Gross, B.; Pawlak, M.; Lefebvre, P.; Staels, B. PPARs in obesity-induced T2DM, dyslipidaemia and NAFLD. Nat. Rev. Endocrinol. 2017, 13, 36–49. [Google Scholar] [CrossRef]
- Azab, A.; Nassar, A.; Azab, A.N. Anti-Inflammatory Activity of Natural Products. Molecules 2016, 21, 1321. [Google Scholar] [CrossRef]
- Tian, Y.; Yang, C.; Yao, Q.; Qian, L.; Liu, J.; Xie, X.; Ma, W.; Nie, X.; Lai, B.; Xiao, L.; et al. Procyanidin B2 Activates PPARgamma to Induce M2 Polarization in Mouse Macrophages. Front. Immunol. 2019, 10, 1895. [Google Scholar] [CrossRef] [Green Version]
- Shi, J.; Zhao, Y.; Wang, K.; Shi, X.; Wang, Y.; Huang, H.; Zhuang, Y.; Cai, T.; Wang, F.; Shao, F. Cleavage of GSDMD by inflammatory caspases determines pyroptotic cell death. Nature 2015, 526, 660–665. [Google Scholar] [CrossRef]
- Kayagaki, N.; Stowe, I.B.; Lee, B.L.; O’Rourke, K.; Anderson, K.; Warming, S.; Cuellar, T.; Haley, B.; Roose-Girma, M.; Phung, Q.T.; et al. Caspase-11 cleaves gasdermin D for non-canonical inflammasome signalling. Nature 2015, 526, 666–671. [Google Scholar] [CrossRef]
- Chen, K.W.; Demarco, B.; Heilig, R.; Shkarina, K.; Boettcher, A.; Farady, C.J.; Pelczar, P.; Broz, P. Extrinsic and intrinsic apoptosis activate pannexin-1 to drive NLRP3 inflammasome assembly. EMBO J. 2019, 38, 1638. [Google Scholar] [CrossRef]
- Wang, Y.; Gao, W.; Shi, X.; Ding, J.; Liu, W.; He, H.; Wang, K.; Shao, F. Chemotherapy drugs induce pyroptosis through caspase-3 cleavage of a gasdermin. Nature 2017, 547, 99–103. [Google Scholar] [CrossRef]
- Ding, J.; Wang, K.; Liu, W.; She, Y.; Sun, Q.; Shi, J.; Sun, H.; Wang, D.C.; Shao, F. Pore-forming activity and structural autoinhibition of the gasdermin family. Nature 2016, 535, 111–116. [Google Scholar] [CrossRef]
- Hernandez-Gea, V.; Friedman, S.L. Pathogenesis of liver fibrosis. Annu. Rev. Pathol. 2011, 6, 425–456. [Google Scholar] [CrossRef]
- Crotty Alexander, L.E.; Drummond, C.A.; Hepokoski, M.; Mathew, D.; Moshensky, A.; Willeford, A.; Das, S.; Singh, P.; Yong, Z.; Lee, J.H.; et al. Chronic inhalation of e-cigarette vapor containing nicotine disrupts airway barrier function and induces systemic inflammation and multiorgan fibrosis in mice. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2018, 314, R834–R847. [Google Scholar] [CrossRef] [Green Version]
- Kuru, P.; Bilgin, S.; Mentese, S.T.; Tazegul, G.; Ozgur, S.; Cilingir, O.T.; Akakin, D.; Yarat, A.; Kasimay, O. Ameliorative effect of chronic moderate exercise in smoke exposed or nicotine applied rats from acute stress. Nicotine Tob. Res. Off. J. Soc. Res. Nicotine Tob. 2015, 17, 559–565. [Google Scholar] [CrossRef]
- Dangana, E.O.; Michael, O.S.; Omolekulo, T.E.; Areola, E.D.; Olatunji, L.A. Enhanced hepatic glycogen synthesis and suppressed adenosine deaminase activity by lithium attenuates hepatic triglyceride accumulation in nicotine-exposed rats. Biomed. Pharmacother. Biomed. Pharmacother. 2019, 109, 1417–1427. [Google Scholar] [CrossRef]
- Wu, L.W.; Guo, C.Y.; Wu, J.Y. Therapeutic potential of PPAR gamma natural agonists in liver diseases. J. Cell. Mol. Med. 2020, 24, 2736–2748. [Google Scholar] [CrossRef]
- Scheen, A.J. Thiazolidinediones and liver toxicity. Diabetes Metab. 2001, 27, 305–313. [Google Scholar] [PubMed]
- Guo, L.; Zhang, L.; Sun, Y.M.; Muskhelishvili, L.; Blann, E.; Dial, S.; Shi, L.M.; Schroth, G.; Dragan, Y.P. Differences in hepatotoxicity and gene expression profiles by anti-diabetic PPAR gamma agonists on rat primary hepatocytes and human HepG2 cells. Mol. Divers. 2006, 10, 349–360. [Google Scholar] [CrossRef]
- Bae, M.A.; Rhee, H.; Song, B.J. Troglitazone but not rosiglitazone induces G1 cell cycle arrest and apoptosis in human and rat hepatoma cell lines. Toxicol. Lett. 2003, 139, 67–75. [Google Scholar] [CrossRef]
- Chen, H.; He, Y.W.; Liu, W.Q.; Zhang, J.H. Rosiglitazone prevents murine hepatic fibrosis induced by Schistosoma japonicum. World J. Gastroenterol. 2008, 14, 2905–2911. [Google Scholar] [CrossRef]
- Yang, C.C.; Wu, C.H.; Lin, T.C.; Cheng, Y.N.; Chang, C.S.; Lee, K.T.; Tsai, P.J.; Tsai, Y.S. Inhibitory effect of PPARgamma on NLRP3 inflammasome activation. Theranostics 2021, 11, 2424–2441. [Google Scholar] [CrossRef]
- Wu, Y.; Li, Y.; Giovannucci, E. Potential Impact of Time Trend of Lifestyle Risk Factors on Burden of Major Gastrointestinal Cancers in China. Gastroenterology 2021, 161, 1830–1841.e1838. [Google Scholar] [CrossRef]
- Kim, S.A.; Shin, S. Fruit and vegetable consumption and non-alcoholic fatty liver disease among Korean adults: A prospective cohort study. J. Epidemiol. Commun. H 2020, 74, 1035–1042. [Google Scholar] [CrossRef]
- Xing, Y.W.; Lei, G.T.; Wu, Q.H.; Jiang, Y.; Huang, M.X. Procyanidin B2 protects against diet-induced obesity and non-alcoholic fatty liver disease via the modulation of the gut microbiota in rabbits. World J. Gastroenterol. 2019, 25, 955–966. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Wang, R.; Xu, R.; Chu, Y.; Gu, W. Procyanidin B2 ameliorates endothelial dysfunction and impaired angiogenesis via the Nrf2/PPARgamma/sFlt-1 axis in preeclampsia. Pharmacol. Res. 2022, 106127. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, J.; Yao, Q.; Xie, X.; Cui, Q.; Jiang, T.; Zhao, Z.; Du, X.; Lai, B.; Xiao, L.; Wang, N. Procyanidin B2 Attenuates Nicotine-Induced Hepatocyte Pyroptosis through a PPARγ-Dependent Mechanism. Nutrients 2022, 14, 1756. https://doi.org/10.3390/nu14091756
Liu J, Yao Q, Xie X, Cui Q, Jiang T, Zhao Z, Du X, Lai B, Xiao L, Wang N. Procyanidin B2 Attenuates Nicotine-Induced Hepatocyte Pyroptosis through a PPARγ-Dependent Mechanism. Nutrients. 2022; 14(9):1756. https://doi.org/10.3390/nu14091756
Chicago/Turabian StyleLiu, Jia, Qinyu Yao, Xinya Xie, Qi Cui, Tingting Jiang, Ziwei Zhao, Xiong Du, Baochang Lai, Lei Xiao, and Nanping Wang. 2022. "Procyanidin B2 Attenuates Nicotine-Induced Hepatocyte Pyroptosis through a PPARγ-Dependent Mechanism" Nutrients 14, no. 9: 1756. https://doi.org/10.3390/nu14091756
APA StyleLiu, J., Yao, Q., Xie, X., Cui, Q., Jiang, T., Zhao, Z., Du, X., Lai, B., Xiao, L., & Wang, N. (2022). Procyanidin B2 Attenuates Nicotine-Induced Hepatocyte Pyroptosis through a PPARγ-Dependent Mechanism. Nutrients, 14(9), 1756. https://doi.org/10.3390/nu14091756