
The Functional Vision Restorative Effect of Crocin via the BDNF–TrkB Pathway: An In Vivo Study


 and
and {kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Materials
2.3. Experimental Design and Animal Groupings
2.4. Determination of Thresholds of the VA and VCSF
2.5. Histological and Immunohistochemistry Analyses
2.6. Protein Analysis
2.7. Statistical Analysis
3. Results
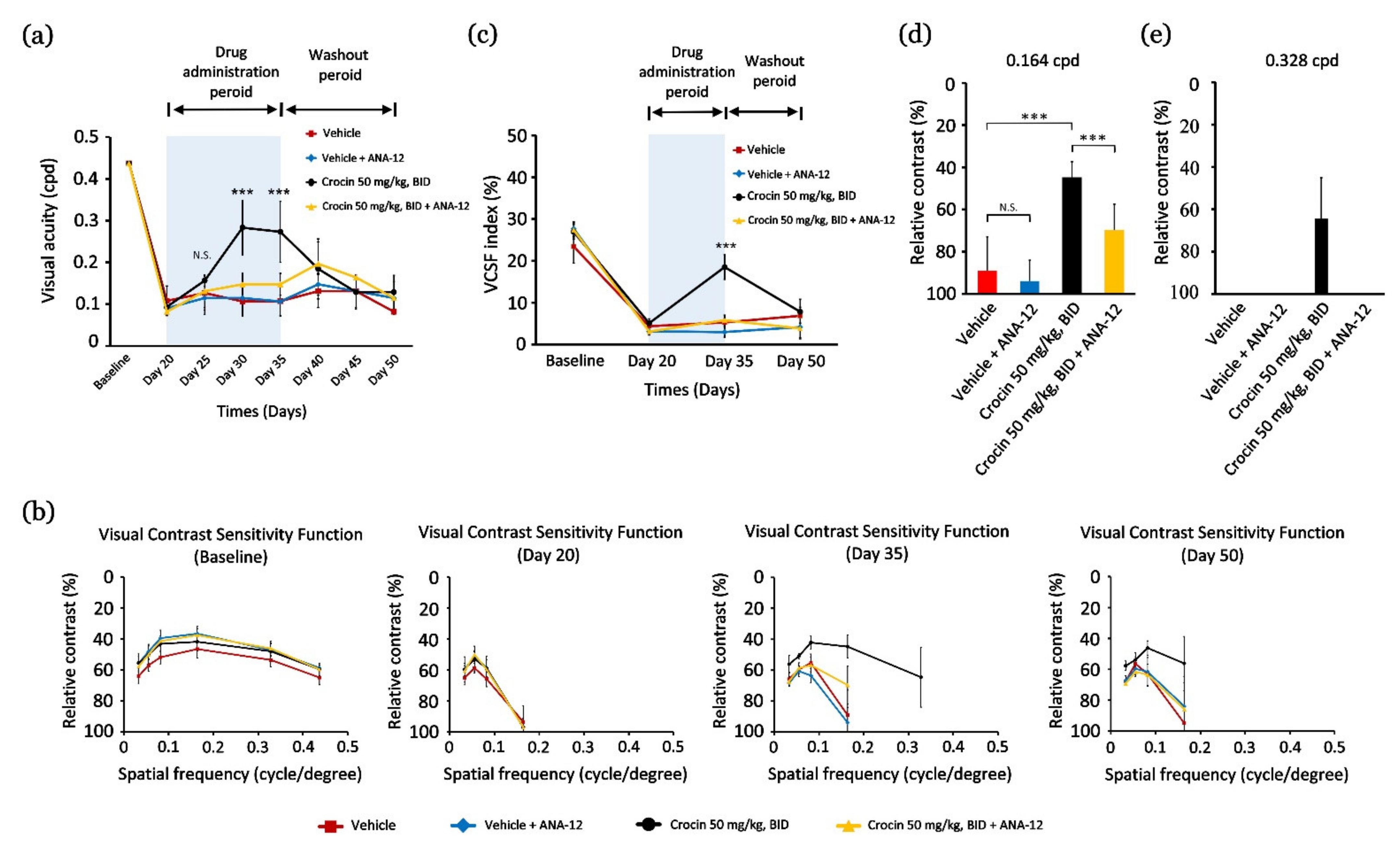
3.1. Crocin Restores the Thresholds of the Residual VA in Light-Evoked Retinal Photodamage Model
3.2. Crocin Promotes VA Restoration via Upregulated Expression of BDNF and TrkB Protein in Retinas
3.3. Crocin Restores and Elevates the Residual Threshold of VA and VCSF via Activating the BDNF–TrkB Pathway
3.4. Crocin via BDNF–TrkB Pathway to Modulate M Opin Protein Localization to Function
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jonas, J.B.; Cheung, C.M.G.; Panda-Jonas, S. Updates on the Epidemiology of Age-Related Macular Degeneration. Asia-Pac. J. Ophthalmol. 2017, 6, 493–497. [Google Scholar]
- Wong, W.L.; Su, X.; Li, X.; Cheung, C.M.; Klein, R.; Cheng, C.Y.; Wong, T.Y. Global prevalence of age-related macular degeneration and disease burden projection for 2020 and 2040: A systematic review and meta-analysis. Lancet Glob. Health 2014, 2, e106–e116. [Google Scholar] [CrossRef] [Green Version]
- Vagge, A.; Ferro Desideri, L.; Del Noce, C.; Di Mola, I.; Sindaco, D.; Traverso, C.E. Blue light filtering ophthalmic lenses: A systematic review. Semin. Ophthalmol. 2021, 36, 541–548. [Google Scholar] [CrossRef] [PubMed]
- Ixcamey, M.; Palma, C. Diabetic macular edema. Disease-a-Month DM 2021, 67, 101138. [Google Scholar] [CrossRef]
- González Martín-Moro, J.; Hernández Verdejo, J.L.; Zarallo Gallardo, J. Photic maculopathy: A review of the literature (I). Arch. Soc. Esp. Oftalmol. 2018, 93, 530–541. [Google Scholar] [CrossRef]
- Elshout, M.; Webers, C.A.; van der Reis, M.I.; de Jong-Hesse, Y.; Schouten, J.S. Tracing the natural course of visual acuity and quality of life in neovascular age-related macular degeneration: A systematic review and quality of life study. BMC Ophthalmol. 2017, 17, 120. [Google Scholar] [CrossRef] [Green Version]
- Ridder, W.H.; Comer, G.; Oquindo, C.; Yoshinaga, P.; Engles, M.; Burke, J. Contrast Sensitivity in Early to Intermediate Age-Related Macular Degeneration (AMD). Curr. Eye Res. 2021, 47, 287–296. [Google Scholar] [CrossRef]
- Maganti, N.; Squires, N.; Mishra, S.; Bomdica, P.; Nigam, D.; Shapiro, A.; Gill, M.K.; Lyon, A.T.; Mirza, R.G. Contrast Sensitivity Testing in Age-Related Macular Degeneration Using Motion Diamond Stimulus. Clin. Ophthalmol. 2022, 16, 507–515. [Google Scholar] [CrossRef]
- Grimm, C.; Remé, C.E. Light Damage Models of Retinal Degeneration. Methods Mol. Biol. 2019, 1834, 167–178. [Google Scholar]
- Chen, Y.W.; Huang, Y.P.; Wu, P.C.; Chiang, W.Y.; Wang, P.H.; Chen, B.Y. The Functional Vision Protection Effect of Danshensu via Dopamine D1 Receptors: In Vivo Study. Nutrients 2021, 13, 978. [Google Scholar] [CrossRef]
- Liou, J.-C.; Yang, S.-L.; Wang, P.-H.; Wu, J.-L.; Huang, Y.-P.; Chen, B.-Y.; Lee, M.-C. Protective effect of crocin against the declining of high spatial frequency-based visual performance in mice. J. Funct. Foods 2018, 49, 314–323. [Google Scholar] [CrossRef]
- Hammond, B.R.; Sreenivasan, V.; Suryakumar, R. The Effects of Blue Light-Filtering Intraocular Lenses on the Protection and Function of the Visual System. Clin. Ophthalmol. 2019, 13, 2427–2438. [Google Scholar] [CrossRef] [Green Version]
- Camelo, S.; Latil, M.; Veillet, S.; Dilda, P.J.; Lafont, R. Beyond AREDS Formulations, What Is Next for Intermediate Age-Related Macular Degeneration (iAMD) Treatment? Potential Benefits of Antioxidant and Anti-inflammatory Apocarotenoids as Neuroprotectors. Oxidative Med. Cell. Longev. 2020, 2020, 4984927. [Google Scholar] [CrossRef]
- Heydari, M.; Zare, M.; Badie, M.R.; Watson, R.R.; Talebnejad, M.R.; Afarid, M. Crocin as a vision supplement. Clin. Exp. Optom. 2022, 1, 1–8. [Google Scholar] [CrossRef]
- Broadhead, G.K.; Grigg, J.R.; McCluskey, P.; Hong, T.; Schlub, T.E.; Chang, A.A. Saffron therapy for the treatment of mild/moderate age-related macular degeneration: A randomised clinical trial. Graefe’s Arch. Clin. Exp. Ophthalmol. 2019, 257, 31–40. [Google Scholar] [CrossRef]
- Sepahi, S.; Mohajeri, S.A.; Hosseini, S.M.; Khodaverdi, E.; Shoeibi, N.; Namdari, M.; Tabassi, S.A.S. Effects of Crocin on Diabetic Maculopathy: A Placebo-Controlled Randomized Clinical Trial. Am. J. Ophthalmol. 2018, 190, 89–98. [Google Scholar] [CrossRef]
- Falsini, B.; Piccardi, M.; Minnella, A.; Savastano, C.; Capoluongo, E.; Fadda, A.; Balestrazzi, E.; Maccarone, R.; Bisti, S. Influence of saffron supplementation on retinal flicker sensitivity in early age-related macular degeneration. Investig. Ophthalmol. Vis. Sci. 2010, 51, 6118–6124. [Google Scholar] [CrossRef]
- Yamauchi, M.; Tsuruma, K.; Imai, S.; Nakanishi, T.; Umigai, N.; Shimazawa, M.; Hara, H. Crocetin prevents retinal degeneration induced by oxidative and endoplasmic reticulum stresses via inhibition of caspase activity. Eur. J. Pharmacol. 2011, 650, 110–119. [Google Scholar] [CrossRef]
- Laabich, A.; Vissvesvaran, G.P.; Lieu, K.L.; Murata, K.; McGinn, T.E.; Manmoto, C.C.; Sinclair, J.R.; Karliga, I.; Leung, D.W.; Fawzi, A.; et al. Protective effect of crocin against blue light- and white light-mediated photoreceptor cell death in bovine and primate retinal primary cell culture. Investig. Ophthalmol. Vis. Sci. 2006, 47, 3156–3163. [Google Scholar] [CrossRef]
- Chen, L.; Qi, Y.; Yang, X. Neuroprotective effects of crocin against oxidative stress induced by ischemia/reperfusion injury in rat retina. Ophthalmic Res. 2015, 54, 157–168. [Google Scholar] [CrossRef]
- Ishizuka, F.; Shimazawa, M.; Umigai, N.; Ogishima, H.; Nakamura, S.; Tsuruma, K.; Hara, H. Crocetin, a carotenoid derivative, inhibits retinal ischemic damage in mice. Eur. J. Pharmacol. 2013, 703, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.F.; Ma, J.X.; Shang, Q.L.; An, J.B.; Chen, H.T.; Wang, C.X. Safety, pharmacokinetics, and prevention effect of intraocular crocetin in proliferative vitreoretinopathy. Biomed. Pharmacother. 2019, 109, 1211–1220. [Google Scholar] [CrossRef] [PubMed]
- Nitta, K.; Nishinaka, A.; Hida, Y.; Nakamura, S.; Shimazawa, M.; Hara, H. Oral and ocular administration of crocetin prevents retinal edema in a murine retinal vein occlusion model. Mol. Vis. 2019, 25, 859–868. [Google Scholar] [PubMed]
- Ohno, Y.; Nakanishi, T.; Umigai, N.; Tsuruma, K.; Shimazawa, M.; Hara, H. Oral administration of crocetin prevents inner retinal damage induced by N-methyl-D-aspartate in mice. Eur. J. Pharmacol. 2012, 690, 84–89. [Google Scholar] [CrossRef]
- Lv, B.; Chen, T.; Xu, Z.; Huo, F.; Wei, Y.; Yang, X. Crocin protects retinal ganglion cells against H2O2-induced damage through the mitochondrial pathway and activation of NF-κB. Int. J. Mol. Med. 2016, 37, 225–232. [Google Scholar] [CrossRef] [Green Version]
- Qi, Y.; Chen, L.; Zhang, L.; Liu, W.B.; Chen, X.Y.; Yang, X.G. Crocin prevents retinal ischaemia/reperfusion injury-induced apoptosis in retinal ganglion cells through the PI3K/AKT signalling pathway. Exp. Eye Res. 2013, 107, 44–51. [Google Scholar] [CrossRef]
- Fernández-Albarral, J.A.; Ramírez, A.I.; de Hoz, R.; López-Villarín, N.; Salobrar-García, E.; López-Cuenca, I.; Licastro, E.; Inarejos-García, A.M.; Almodóvar, P.; Pinazo-Durán, M.D.; et al. Neuroprotective and Anti-Inflammatory Effects of a Hydrophilic Saffron Extract in a Model of Glaucoma. Int. J. Mol. Sci. 2019, 20, 4110. [Google Scholar] [CrossRef] [Green Version]
- Xuan, B.; Zhou, Y.H.; Li, N.; Min, Z.D.; Chiou, G.C. Effects of crocin analogs on ocular blood flow and retinal function. J. Ocul. Pharmacol. Ther. Off. J. Assoc. Ocul. Pharmacol. Ther. 1999, 15, 143–152. [Google Scholar] [CrossRef]
- Piccardi, M.; Fadda, A.; Martelli, F.; Marangoni, D.; Magli, A.; Minnella, A.M.; Bertelli, M.; Di Marco, S.; Bisti, S.; Falsini, B. Antioxidant Saffron and Central Retinal Function in ABCA4-Related Stargardt Macular Dystrophy. Nutrients 2019, 11, 2461. [Google Scholar] [CrossRef] [Green Version]
- Owsley, C.; Huisingh, C.; Clark, M.E.; Jackson, G.R.; McGwin, G., Jr. Comparison of Visual Function in Older Eyes in the Earliest Stages of Age-related Macular Degeneration to Those in Normal Macular Health. Curr. Eye Res. 2016, 41, 266–272. [Google Scholar] [CrossRef] [Green Version]
- Saito, T.; Abe, T.; Wakusawa, R.; Sato, H.; Asai, H.; Tokita-Ishikawa, Y.; Nishida, K. TrkB-T1 receptors on Muller cells play critical role in brain-derived neurotrophic factor-mediated photoreceptor protection against phototoxicity. Curr. Eye Res. 2009, 34, 580–588. [Google Scholar] [CrossRef]
- Daly, C.; Shine, L.; Heffernan, T.; Deeti, S.; Reynolds, A.L.; O’Connor, J.J.; Dillon, E.T.; Duffy, D.J.; Kolch, W.; Cagney, G.; et al. A Brain-Derived Neurotrophic Factor Mimetic Is Sufficient to Restore Cone Photoreceptor Visual Function in an Inherited Blindness Model. Sci. Rep. 2017, 7, 11320. [Google Scholar] [CrossRef] [Green Version]
- Di Polo, A.; Cheng, L.; Bray, G.M.; Aguayo, A.J. Colocalization of TrkB and brain-derived neurotrophic factor proteins in green-red-sensitive cone outer segments. Investig. Ophthalmol. Vis. Sci. 2000, 41, 4014–4021. [Google Scholar]
- Curcio, C.A. Photoreceptor topography in ageing and age-related maculopathy. Eye 2001, 15, 376–383. [Google Scholar] [CrossRef] [Green Version]
- Jackson, G.R.; Owsley, C.; Curcio, C.A. Photoreceptor degeneration and dysfunction in aging and age-related maculopathy. Ageing Res. Rev. 2002, 1, 381–396. [Google Scholar] [CrossRef]
- Lem, D.W.; Davey, P.G.; Gierhart, D.L.; Rosen, R.B. A Systematic Review of Carotenoids in the Management of Age-Related Macular Degeneration. Antioxidants 2021, 10, 1255. [Google Scholar] [CrossRef]
- Hadziahmetovic, M.; Malek, G. Age-Related Macular Degeneration Revisited: From Pathology and Cellular Stress to Potential Therapies. Front. Cell Dev. Biol. 2020, 8, 612812. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, J.-L.; Yang, S.-L.; Ho, Y.-C.; Chen, C.-H.; Tasi, B.-R.; Lee, M.-C.; Chen, B.-Y. The Functional Vision Restorative Effect of Crocin via the BDNF–TrkB Pathway: An In Vivo Study. Nutrients 2022, 14, 1716. https://doi.org/10.3390/nu14091716
Wu J-L, Yang S-L, Ho Y-C, Chen C-H, Tasi B-R, Lee M-C, Chen B-Y. The Functional Vision Restorative Effect of Crocin via the BDNF–TrkB Pathway: An In Vivo Study. Nutrients. 2022; 14(9):1716. https://doi.org/10.3390/nu14091716
Chicago/Turabian StyleWu, Jia-Lain, Shih-Liang Yang, Yung-Chuan Ho, Chao-Hsiang Chen, Bing-Rong Tasi, Meng-Chih Lee, and Bo-Yie Chen. 2022. "The Functional Vision Restorative Effect of Crocin via the BDNF–TrkB Pathway: An In Vivo Study" Nutrients 14, no. 9: 1716. https://doi.org/10.3390/nu14091716
APA StyleWu, J.-L., Yang, S.-L., Ho, Y.-C., Chen, C.-H., Tasi, B.-R., Lee, M.-C., & Chen, B.-Y. (2022). The Functional Vision Restorative Effect of Crocin via the BDNF–TrkB Pathway: An In Vivo Study. Nutrients, 14(9), 1716. https://doi.org/10.3390/nu14091716