
Neither Incretin or Amino Acid Responses, nor Casein Content, Account for the Equal Insulin Response Following Iso-Lactose Loads of Natural Human and Cow Milk in Healthy Young Adults



 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Type and Composition of Tested Milks
2.3. Milk Provision and Preparation
2.3.1. Cow Milk
2.3.2. Human Milk
2.3.3. Low Casein Cow-Milk
2.3.4. Casein-Added Human Milk
2.4. Experimental Procedures
2.5. Biochemical Analyses
2.6. Statistical Analysis
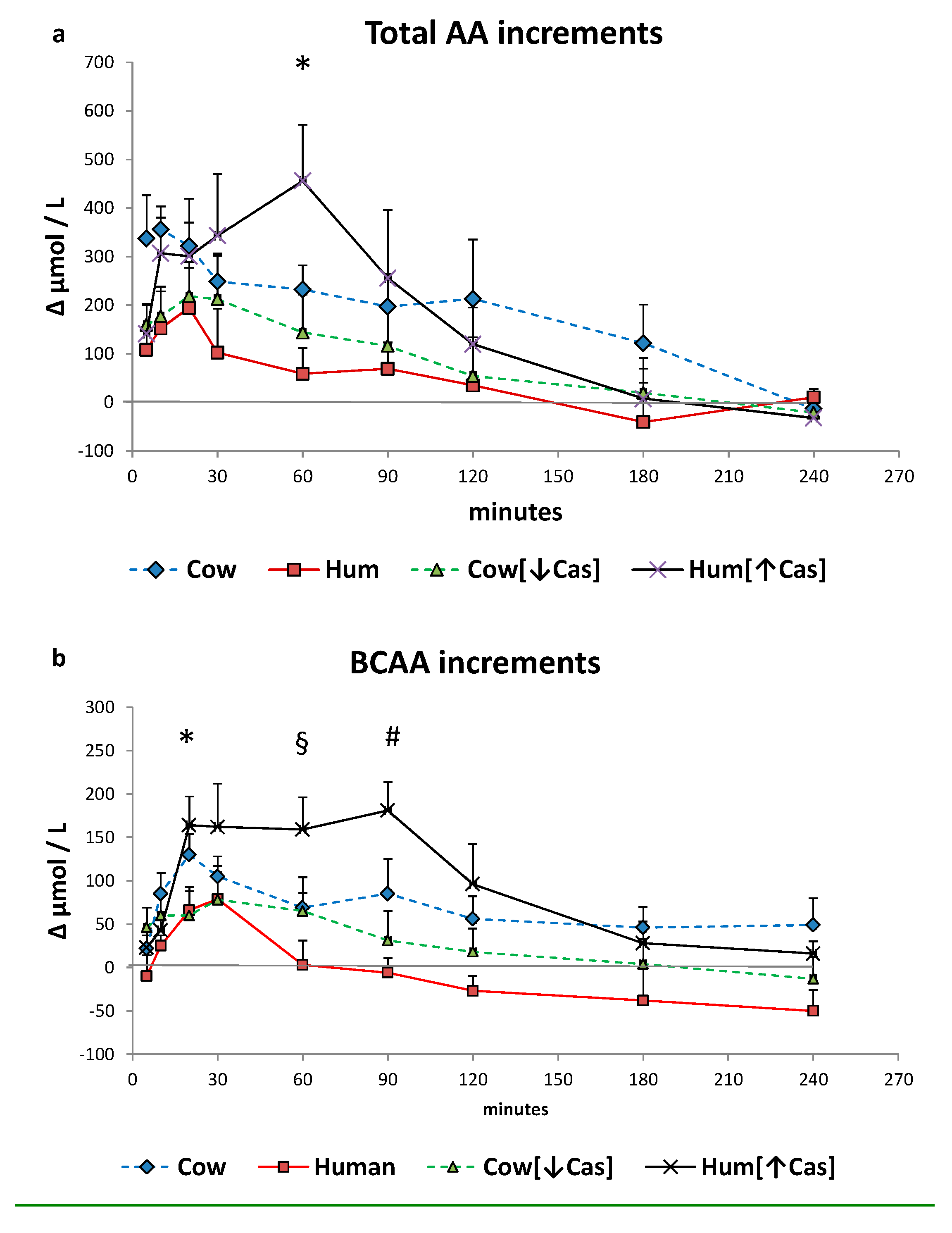
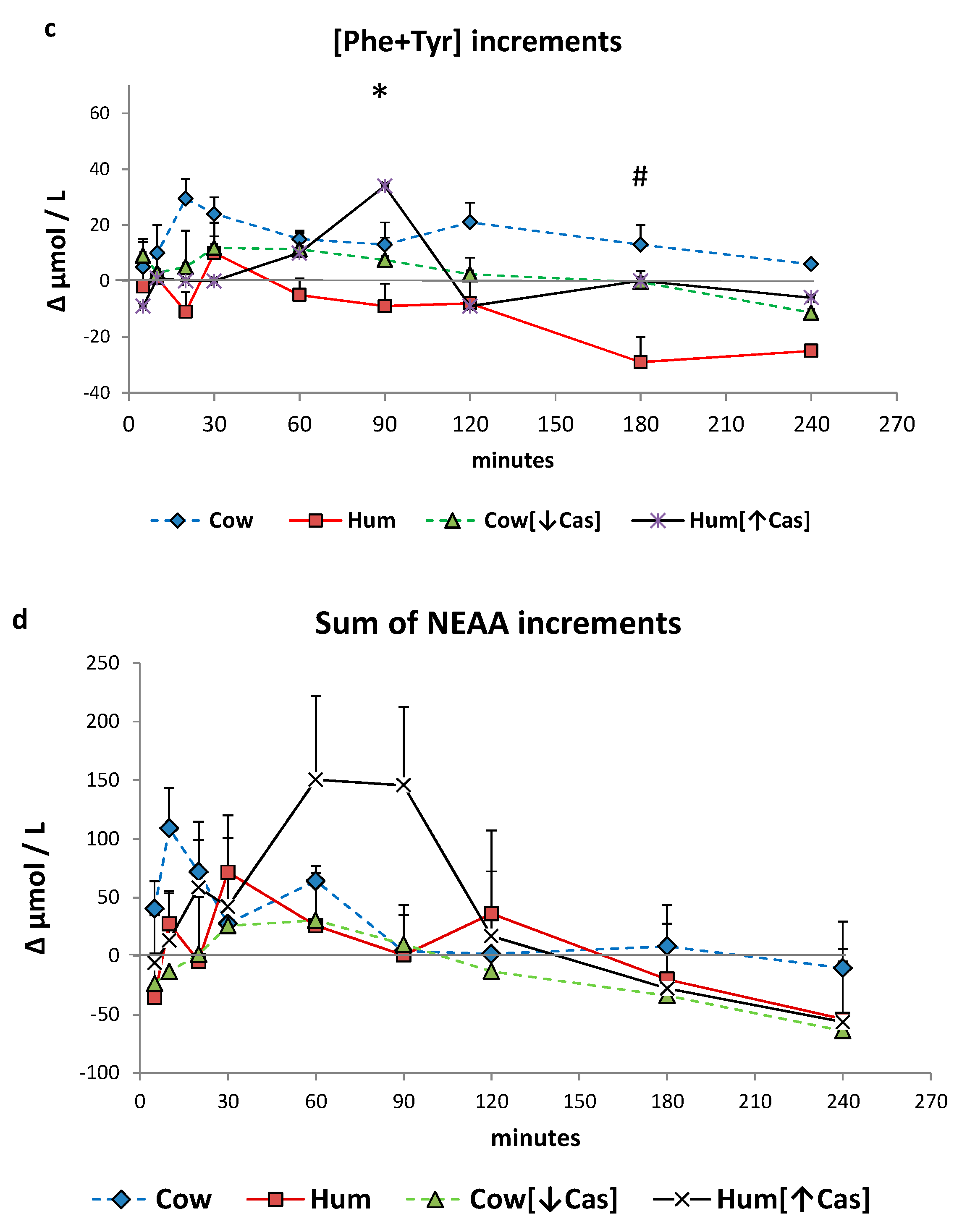
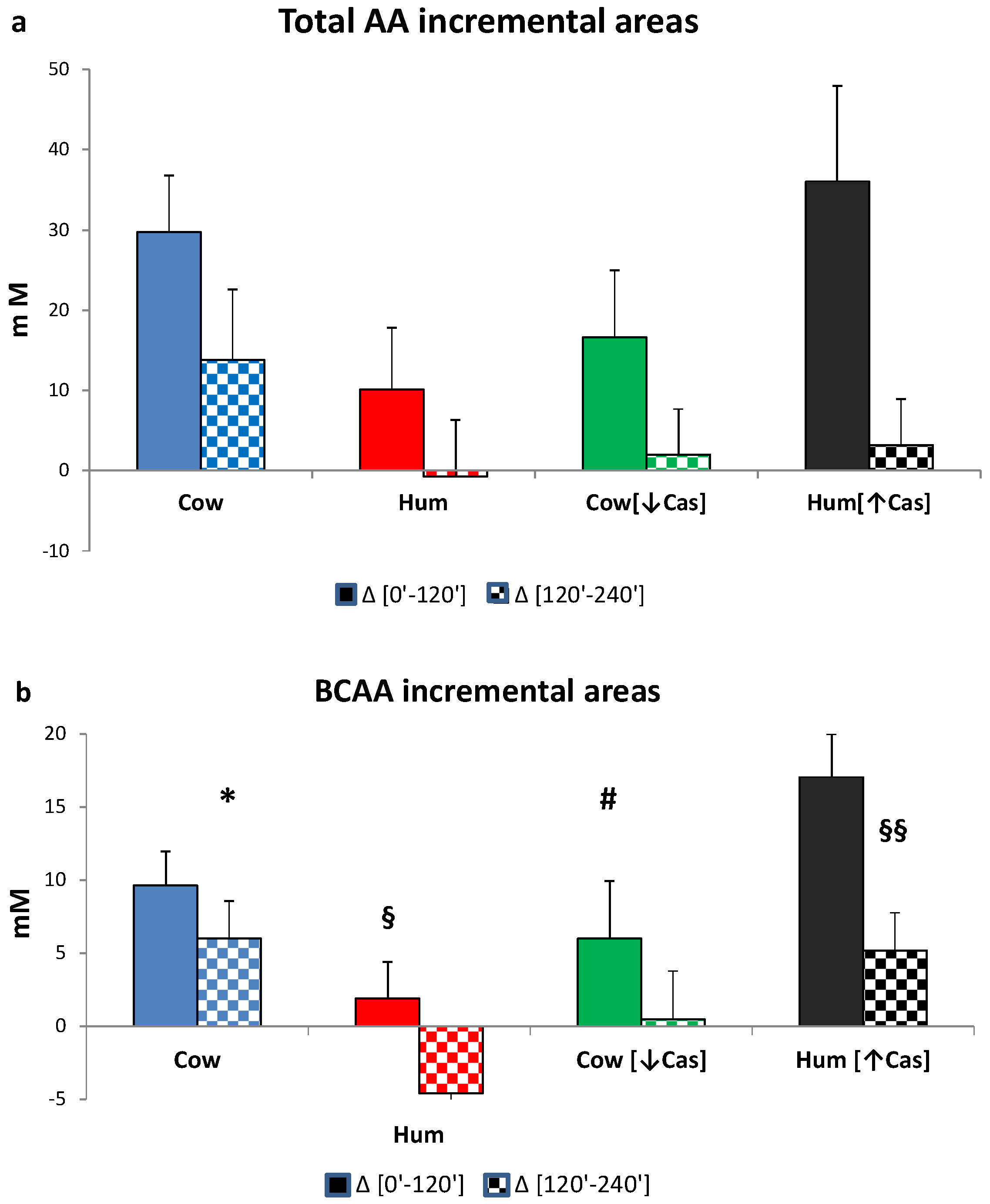
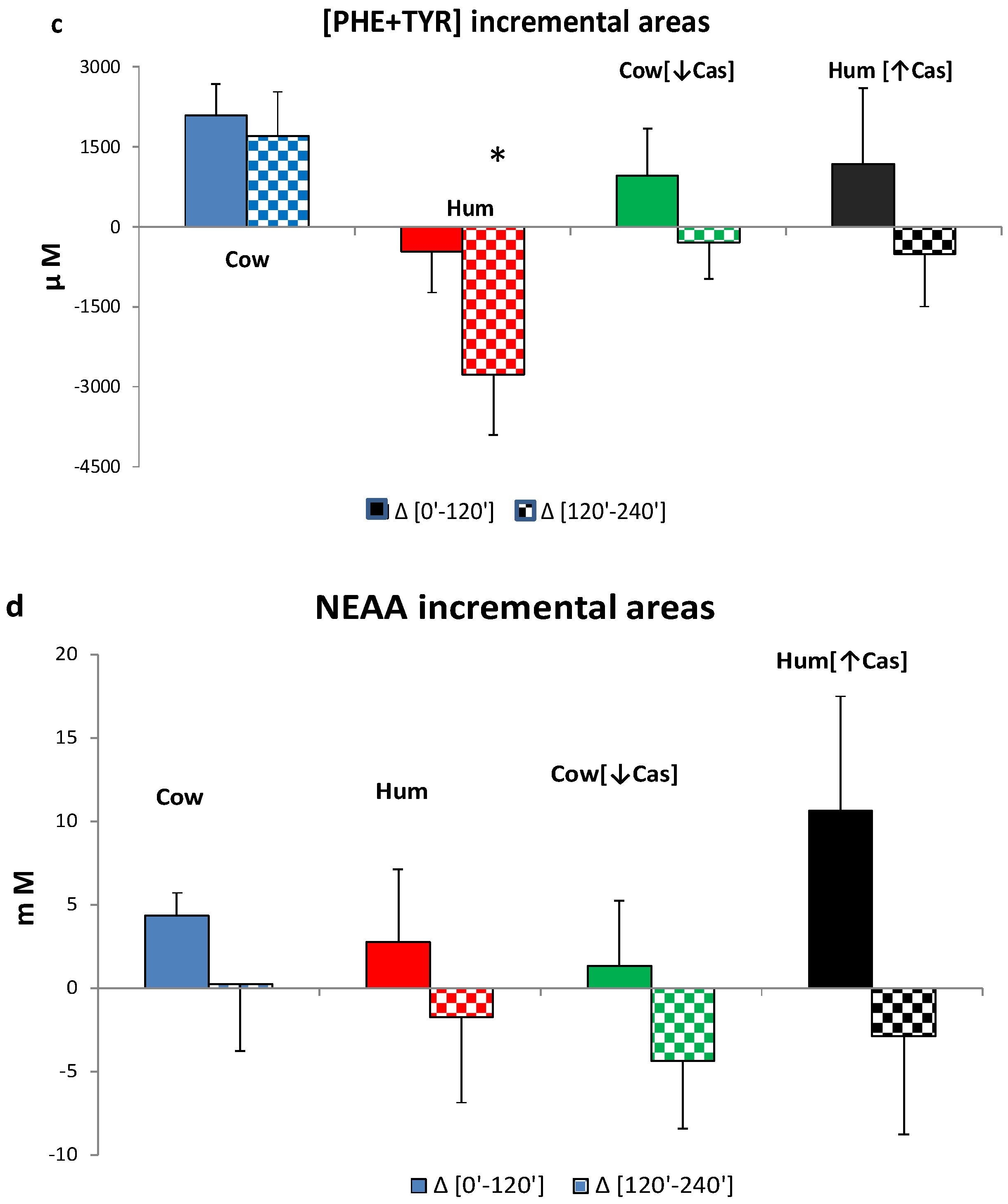
3. Results
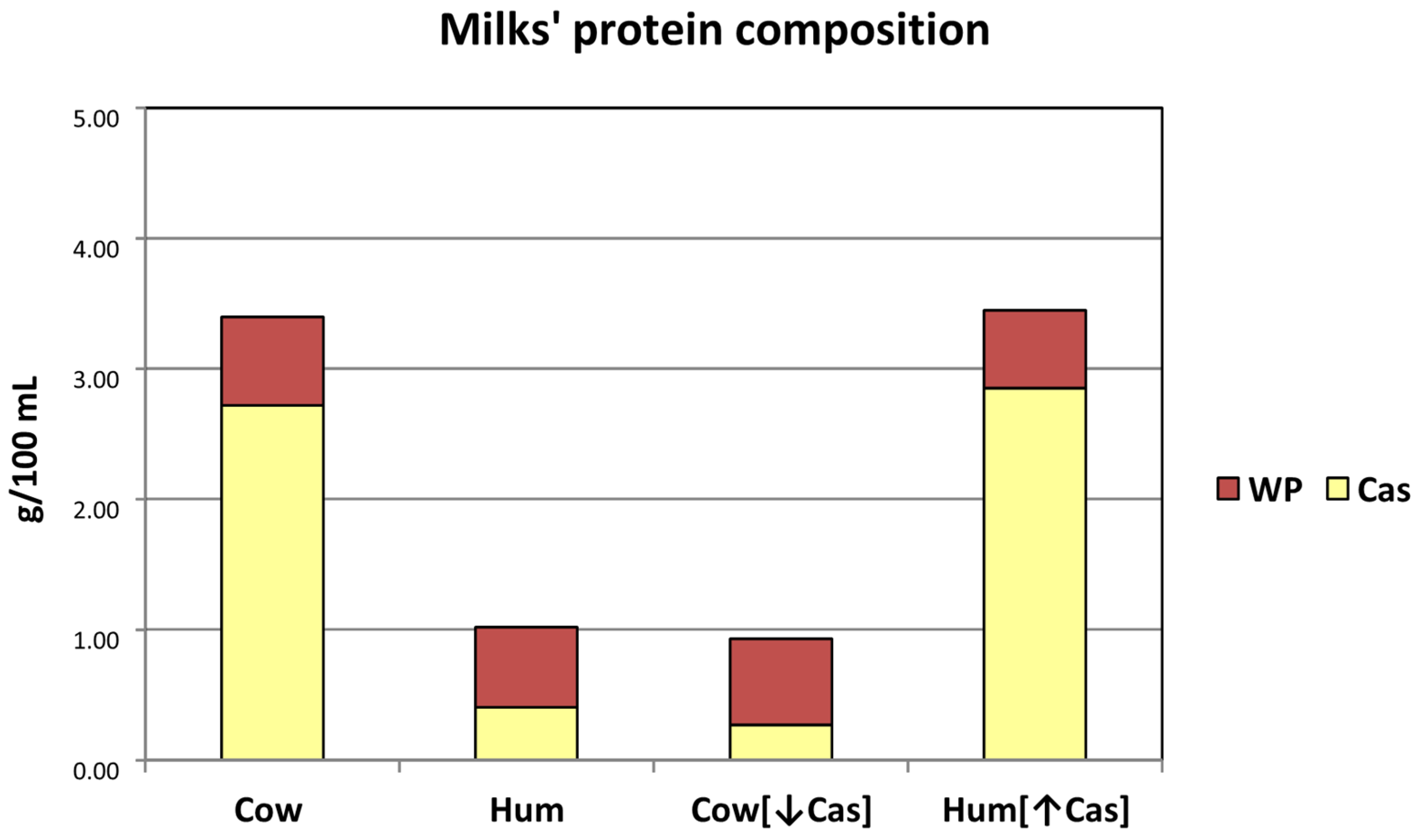
3.1. Test Milk Composition and Loads
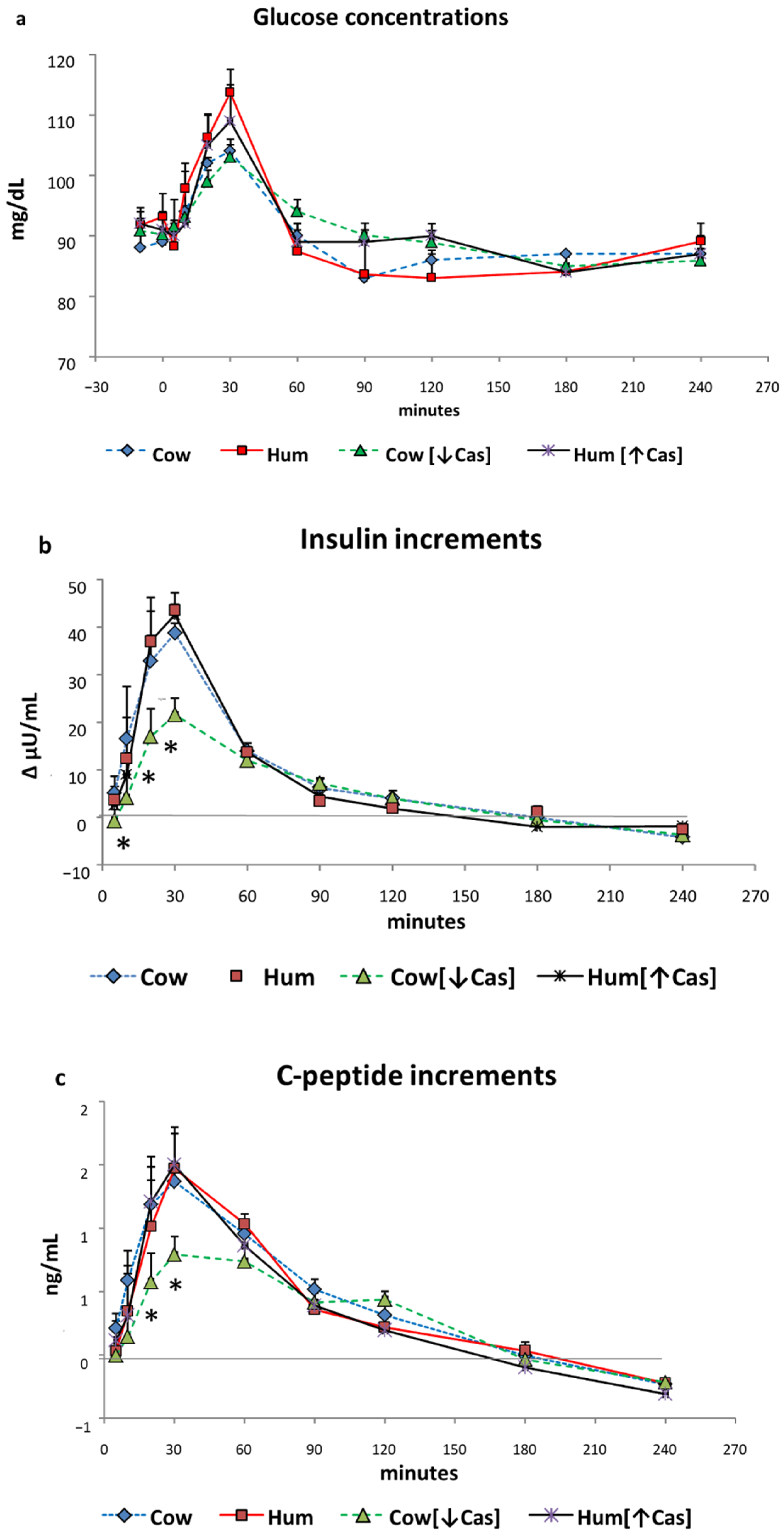
3.2. Glucose
3.3. Insulin
3.4. C-Peptide
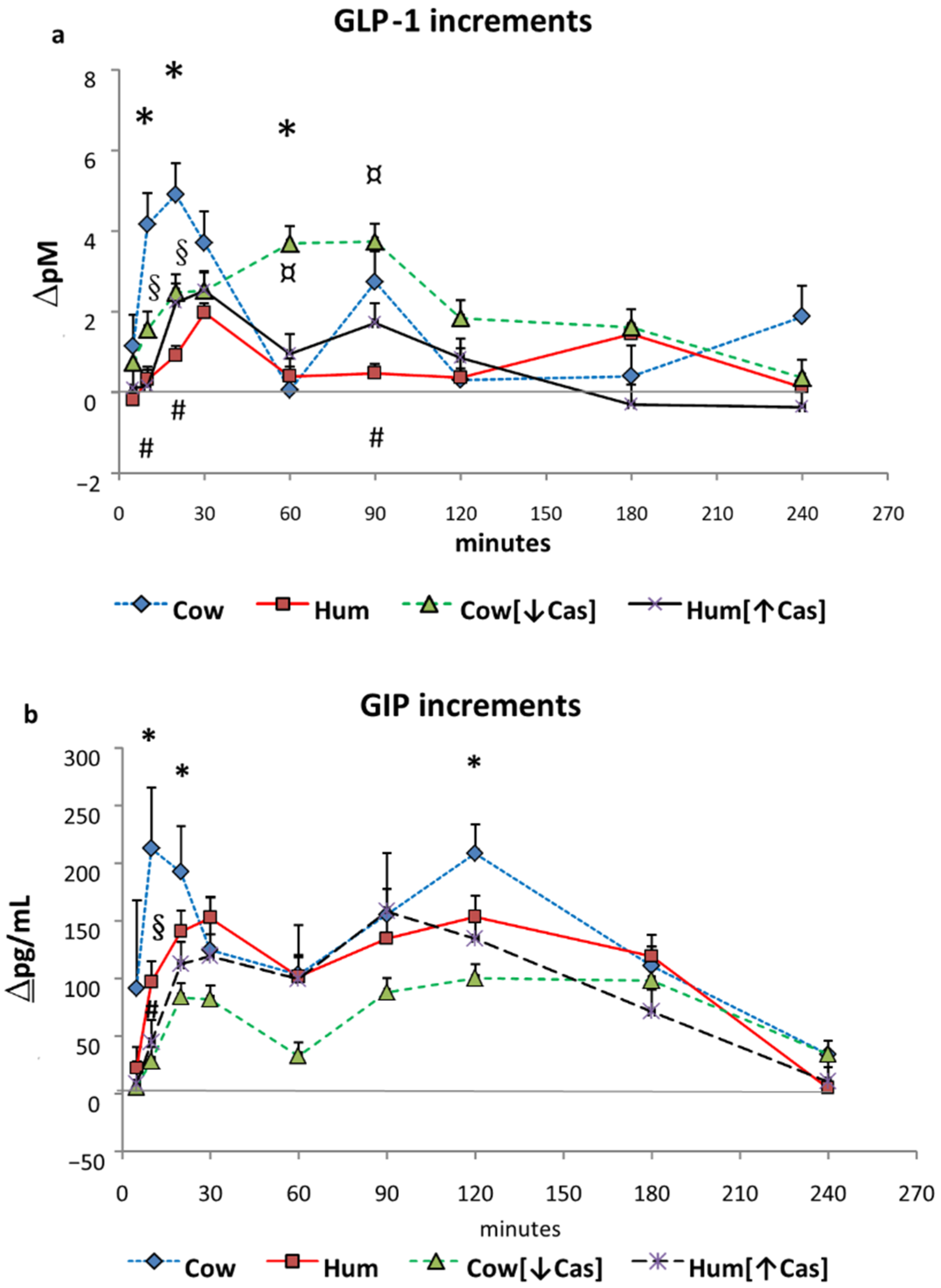
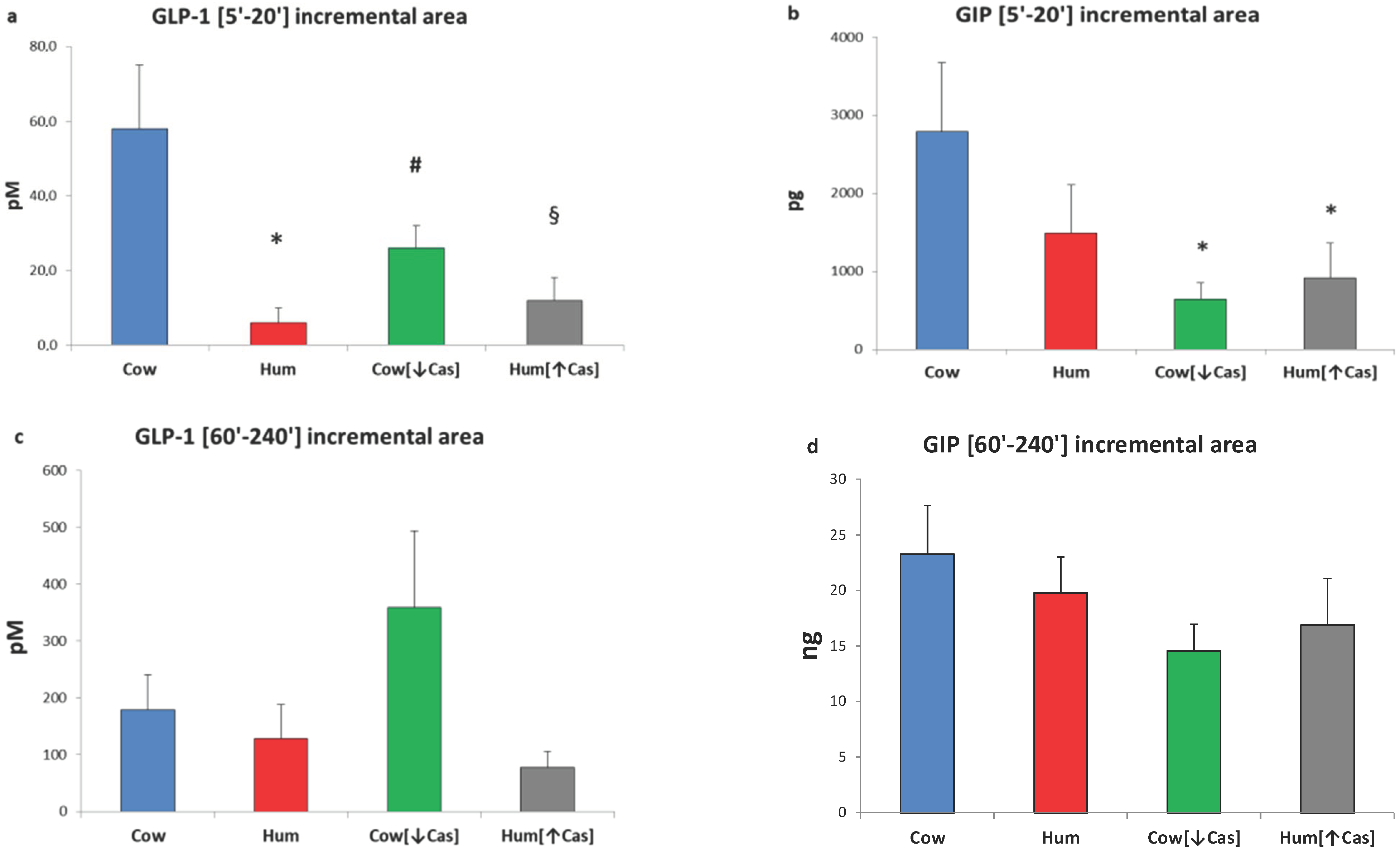
3.5. GLP-1 and GIP
3.6. Plasma Amino Acids
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
Appendix A.1. Participants
Appendix A.2. Participants’ Flow Chart

Appendix A.3. Steps of Recruitment
- Advertisements/invitation to participate in the study were placed at Students’ university college(s).
- Personal data and accurate familiar and personal medical history from potential candidates of both sexes, were collected by the investigators. An in-depth enquiry about present or past pathologies and/or other exclusion features (lifestyle, addictions, alcohol use or abuse, smoking, pharmacological treatments) was performed.
- A detailed description of the aims of the study, and of potential adverse reactions, including an accurate overview analysis of the consent form to be signed prior to the in vivo test, were submitted to each candidate.
- Of the twenty subjects initially selected, two refused to participate in the study for unforeseen subsequent commitments. One enrolled male subject participated in two studies, but then his data were not included in the experimental group(s), because of post hoc blood glucose values resulting in the range of impaired fasting glucose (IFG) criteria (i.e., 112–116 mg/dL), and also because the investigators had reasonable doubts about his observance of the required fasting period (>12 h overnight) as well as of alcohol abstinence prior to studies.
- Seventeen subjects were finally enrolled (n = 17, 8 males, 9 females, age: 23–28 yrs.). They were sequentially and randomly assigned to either one or more of the four experimental groups. Although the investigators made them the proposal to participate in all the four test types, none of them agreed. Therefore, six participants completed three studies, four two studies, while the remainder six participants underwent one study only. Thus, a total of 32 studies were performed. Repeated studies in the same volunteer were spaced by at least two weeks. Since the volunteers were sequentially enrolled over 2–3 yrs., no randomization could be planned at the start of the protocol, rather it was sequentially applied, as far as possible, when the volunteers became available.
- Following the analyses of plasma samples after the studies, the GLP-1 data of one subject, out of the total ten of the Cow [↓Cas] milk group, as well as those of one subject out of eight subjects of the Hum milk group, were excluded from calculation, because of outlier behavior (values >4-fold greater than the mean ± 1 SD of the group). In one subject of the Hum [↓Cas] milk group, GIP was not determined for lack of plasma sample.
Appendix A.4. Milk Sources and Preparation
Appendix A.5. Milk Protein Composition and Analysis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cow Milk | Human Milk | |
|---|---|---|
| Concentrations (g/100 mL) | ||
| Total protein | 3.40 | 1.02 |
| Serum protein | 0.65 | 0.58 |
| Calculated casein | 2.75 | 0.44 |
| Casein/WP ratio | 81/19 | 43/57 |
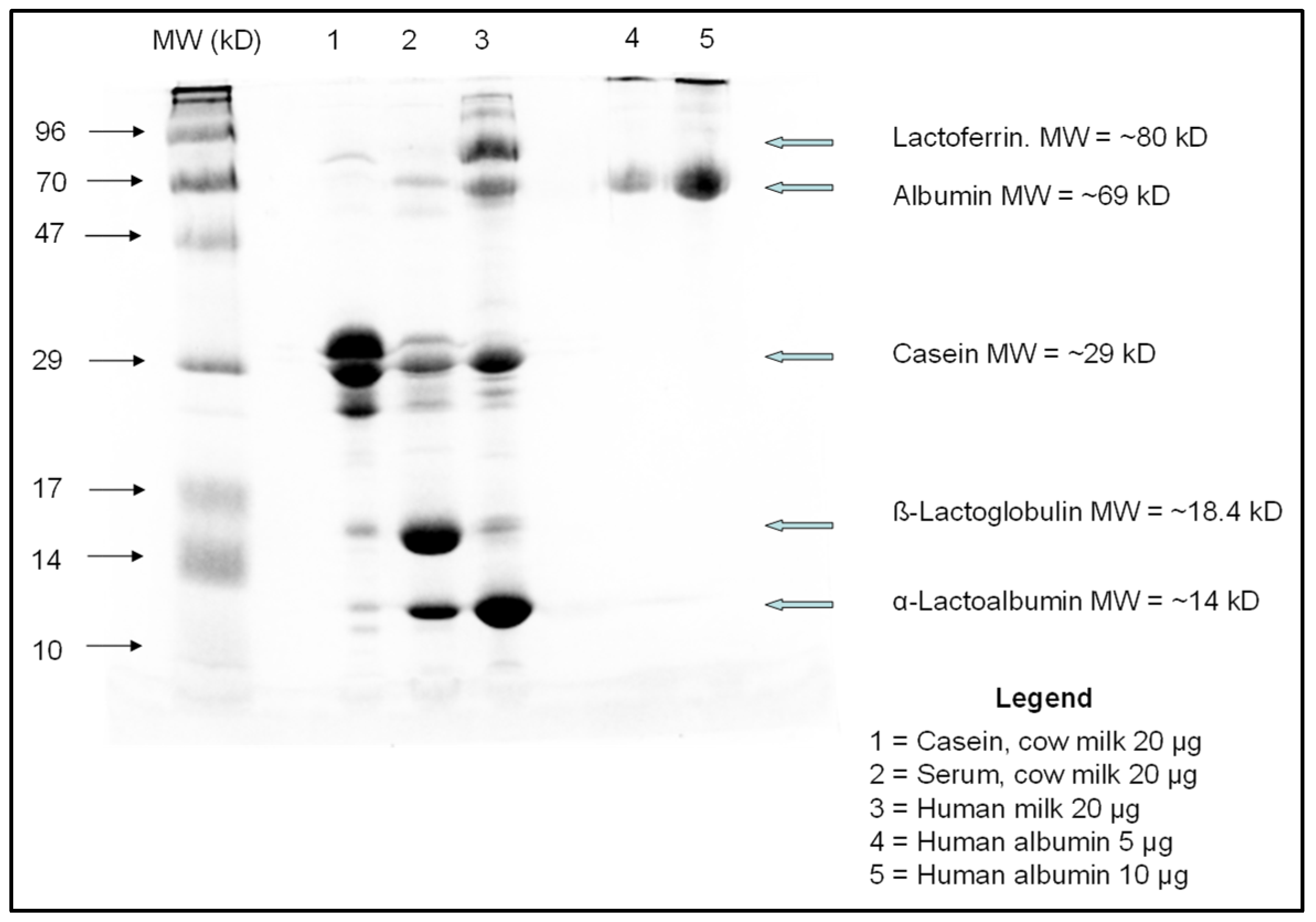
Appendix A.6. SDS PAGE Electrophoresis: Methodology

Appendix A.7. Milk Sources and Preparation
Appendix A.7.1. Cow Milk
Appendix A.7.2. Human Milk
Appendix A.7.3. Low Casein Cow-Milk
Appendix A.7.4. Casein-Added Human Milk
Appendix A.8. Lactose Addition to Cow-Based Milk(s)
Appendix A.9. Administered Milk Volumes
Appendix A.10. Plasma Amino Acid Analysis by Gas Chromatography-Mass Spectrometry (GCMS)
References
- Pereira, P.C. Milk nutritional composition and its role in human health. Nutrition 2014, 30, 619–627. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, R.; Meisel, H. Food-derived peptides with biological activity: From research to food applications. Curr. Opin. Biotechnol. 2007, 18, 163–169. [Google Scholar] [CrossRef] [PubMed]
- Madureira, A.; Tavares, T.; Gomes, A.; Pintado, M.; Malcata, F. Invited review: Physiological properties of bioactive peptides obtained from whey proteins. J. Dairy Sci. 2010, 93, 437–455. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Fan, Y.; Shaikh, A.S.; Wang, Z.; Wang, D.; Tan, H. Dezhou donkey (Equus asinus) milk a potential treatment strategy for type 2 diabetes. J. Ethnopharmacol. 2019, 246, 112221. [Google Scholar] [CrossRef] [PubMed]
- Lionetti, L.; Cavaliere, G.; Bergamo, P.; Trinchese, G.; De Filippo, C.; Gifuni, G.; Gaita, M.; Pignalosa, A.; Donizzetti, I.; Putti, R.; et al. Diet supplementation with donkey milk upregulates liver mitochondrial uncoupling, reduces energy efficiency and improves antioxidant and antiinflammatory defences in rats. Mol. Nutr. Food Res. 2012, 56, 1596–1600. [Google Scholar] [CrossRef] [PubMed]
- Haug, A.; Høstmark, A.T.; Harstad, O.M. Bovine milk in human nutrition—A review. Lipids Health Dis. 2007, 6, 25. [Google Scholar] [CrossRef]
- Andreas, N.J.; Kampmann, B.; Le-Doare, K.M. Human breast milk: A review on its composition and bioactivity. Early Hum. Dev. 2015, 91, 629–635. [Google Scholar] [CrossRef]
- Lönnerdal, B. Nutritional and physiologic significance of human milk proteins. Am. J. Clin. Nutr. 2003, 77, 1537S–1543S. [Google Scholar] [CrossRef]
- Lönnerdal, B.; Erdmann, P.; Thakkar, S.K.; Sauser, J.; Destaillats, F. Longitudinal evolution of true protein, amino acids and bioactive proteins in breast milk: A developmental perspective. J. Nutr. Biochem. 2017, 41, 1–11. [Google Scholar] [CrossRef]
- Savino, F.; Liguori, S.A.; Sorrenti, M.; Fissore, M.F.; Oggero, R. Breast Milk Hormones and Regulation of Glucose Homeostasis. Int. J. Pediatr. 2011, 2011, 1–11. [Google Scholar] [CrossRef]
- Trinchese, G.; Cavaliere, G.; De Filippo, C.; Aceto, S.; Prisco, M.; Chun, J.T.; Penna, E.; Negri, R.; Muredda, L.; Demurtas, A.; et al. Human Milk and Donkey Milk, Compared to Cow Milk, Reduce Inflammatory Mediators and Modulate Glucose and Lipid Metabolism, Acting on Mitochondrial Function and Oleylethanolamide Levels in Rat Skeletal Muscle. Front. Physiol. 2018, 9, 32. [Google Scholar] [CrossRef] [PubMed]
- Hernell, O. Human milk vs. cow’s milk and the evolution of infant formulas. Nestle Nutr. Workshop Ser. Pediatr. Program. 2011, 67, 17–28. [Google Scholar] [CrossRef] [PubMed]
- Castilho, S.D.; Filho, A.D.A.B. The history of infant nutrition. J. Pediatr. 2010, 86, 179–188. [Google Scholar] [CrossRef] [PubMed]
- European Childhood Obesity Trial Study Group; Koletzko, B.; Von Kries, R.; Closa, R.; Escribano, J.; Scaglioni, S.; Giovannini, M.; Beyer, J.; Demmelmair, H.; Gruszfeld, D.; et al. Lower protein in infant formula is associated with lower weight up to age 2 y: A randomized clinical trial. Am. J. Clin. Nutr. 2009, 89, 1836–1845. [Google Scholar] [CrossRef]
- Oddy, W.H. Infant feeding and obesity risk in the child. Breastfeed. Rev. 2012, 20, 7–12. [Google Scholar]
- Liotto, N. Protein content of infant formula for the healthy full-term infant. Am. J. Clin. Nutr. 2020, 111, 946–947. [Google Scholar] [CrossRef]
- European Childhood Obesity Trial Study Group; Socha, P.; Grote, V.; Gruszfeld, D.; Janas, R.; Demmelmair, H.; Closa-Monasterolo, R.; Subías, J.E.; Scaglioni, S.; Verduci, E.; et al. Milk protein intake, the metabolic-endocrine response, and growth in infancy: Data from a randomized clinical trial. Am. J. Clin. Nutr. 2011, 94, 1776S–1784S. [Google Scholar] [CrossRef]
- Gunnerud, U.; Holst, J.J.; Östman, E.; Björck, I. The glycemic, insulinemic and plasma amino acid responses to equi-carbohydrate milk meals, a pilot- study of bovine and human milk. Nutr. J. 2012, 11, 83. [Google Scholar] [CrossRef]
- Floyd, J.C.; Fajans, S.S.; Pek, S.; A Thiffault, C.; Knopf, R.F.; Conn, J.W. Synergistic Effect of Essential Amino Acids and Glucose upon Insulin Secretion in Man. Diabetes 1970, 19, 109–115. [Google Scholar] [CrossRef]
- Schmid, R.; Schulte-Frohlinde, E.; Schusdziarra, V.; Neubauer, J.; Stegmann, M.; Maier, V.; Classen, M. Contribution of Postprandial Amino Acid Levels to Stimulation of Insulin, Glucagon, and Pancreatic Polypeptide in Humans. Pancreas 1992, 7, 698–704. [Google Scholar] [CrossRef]
- Nilsson, M.; Stenberg, M.; Frid, A.H.; Holst, J.J.; Björck, I.M. Glycemia and insulinemia in healthy participants after lactose-equivalent meals of milk and other food proteins: The role of plasma amino acids and incretins. Am. J. Clin. Nutr. 2004, 80, 1246–1253. [Google Scholar] [CrossRef] [PubMed]
- Van Loon, L.J.; Kruijshoop, M.; Menheere, P.P.; Wagenmakers, A.J.; Saris, W.H.; Keizer, H.A. Amino acid ingestion strongly enhances insulin secretion in patients with long-term type 2 diabetes. Diabetes Care 2003, 26, 625–630. [Google Scholar] [CrossRef]
- Nauck, M.A.; Meier, J.J. Incretin hormones: Their role in health and disease. Diabetes Obes. Metab. 2018, 20 (Suppl. 1), 5–21. [Google Scholar] [CrossRef] [PubMed]
- Egger, L.; Ménard, O.; Baumann, C.; Duerr, D.; Schlegel, P.; Stoll, P.; Vergères, G.; Dupont, D.; Portmann, R. Digestion of milk proteins: Comparing static and dynamic in vitro digestion systems with in vivo data. Food Res. Int. 2019, 118, 32–39. [Google Scholar] [CrossRef] [PubMed]
- World Medical Association. World Medical Association Declaration of Helsinki: Ethical principles for medical research involving human subjects. JAMA 2013, 310, 2191–2194. [Google Scholar] [CrossRef] [PubMed]
- Livesey, G. A perspective on food energy standards for nutrition labelling. Br. J. Nutr. 2001, 85, 271–287. [Google Scholar] [CrossRef] [PubMed]
- Arslanoglu, S.; Bertino, E.; Tonetto, P.; De Nisi, G.; Ambruzzi, A.M.; Biasini, A.; Profeti, C.; Spreghini, M.R.; Moro, G.E. Guidelines for the establishment and operation of a donor human milk bank. J. Matern. Neonatal Med. 2010, 23, 1–20. [Google Scholar] [CrossRef]
- Strata, A. Milk and derivatives. Functional properties. Progr. Nutr. 2013, 15, 3–32. (In Italian) [Google Scholar]
- Bijl, E.; van Valenberg, H.J.F.; Huppertz, T.; van Hooijdonk, A.C.M. Protein, casein, and micellar salts in milk: Current content and historical perspectives. J. Dairy Sci. 2013, 96, 5455–5464. [Google Scholar] [CrossRef]
- Adams, R.F. Determination of amino acid profiles in biological samples by gas chromatography. J. Chromatogr. A 1974, 95, 189–212. [Google Scholar] [CrossRef]
- Schwenk, W.F.; Berg, P.J.; Beaufrere, B.; Miles, J.M.; Haymond, M.W. Use of t-butyldimethylsilylation in GC/MS analysis of physiologic compounds in plasma using electron impact ionization. Anal. Biochem. 1982, 141, 101–109. [Google Scholar] [CrossRef]
- Csapó, J.; Albert, C.S.; Lók, K.; Csapó-Kiss, Z.S. Separation and determination of the amino acids by ion exchange column chromatography applying post-column derivatization. Acta Univ. Sapientiae Aliment. 2008, 5, 29. [Google Scholar]
- Garber, A. The importance of early insulin secretion and its impact on glycaemic regulation. Int. J. Obes. 2000, 24, S32–S37. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Hall, W.L.; Millward, D.J.; Long, S.J.; Morgan, L.M. Casein and whey exert different effects on plasma amino acid profiles, gastrointestinal hormone secretion and appetite. Br. J. Nutr. 2003, 89, 239–248. [Google Scholar] [CrossRef] [PubMed]
- Boirie, Y.; Dangin, M.; Gachon, P.; Vasson, M.-P.; Maubois, J.-L.; Beaufrère, B. Slow and fast dietary proteins differently modulate postprandial protein accretion. Proc. Natl. Acad. Sci. USA 1997, 94, 14930–14935. [Google Scholar] [CrossRef]
- Salehi, A.; Gunnerud, U.; Muhammed, S.J.; Östman, E.; Holst, J.J.; Björck, I.; Rorsman, P. The insulinogenic effect of whey protein is partially mediated by a direct effect of amino acids and GIP on β-cells. Nutr. Metab. 2012, 9, 48. [Google Scholar] [CrossRef]
- Magnuson, S.J.; Henry, J.F.; Yip, T.T.; Hutchens, W.T. Structural Homology of Human, Bovine, and Porcine Milk Lactoferrins: Evidence for Shared Antigenic Determinants. Pediatr. Res. 1990, 28, 176–181. [Google Scholar] [CrossRef]
- Calbet, J.A.L.; Holst, J.J. Gastric emptying, gastric secretion and enterogastrone response after administration of milk proteins or their peptide hydrolysates in humans. Eur. J. Nutr. 2004, 43, 127–139. [Google Scholar] [CrossRef]
- Stanstrup, J.; Schou, S.S.; Holmer-Jensen, J.; Hermansen, K.; Dragsted, L.O. Whey Protein Delays Gastric Emptying and Suppresses Plasma Fatty Acids and Their Metabolites Compared to Casein, Gluten, and Fish Protein. J. Proteome Res. 2014, 13, 2396–2408. [Google Scholar] [CrossRef]
- Ma, J.; Stevens, J.E.; Cukier, K.; Maddox, A.F.; Wishart, J.M.; Jones, K.L.; Clifton, P.M.; Horowitz, M.; Rayner, C.K. Effects of a Protein Preload on Gastric Emptying, Glycemia, and Gut Hormones After a Carbohydrate Meal in Diet-Controlled Type 2 Diabetes. Diabetes Care 2009, 32, 1600–1602. [Google Scholar] [CrossRef]
- Carver, J.A.; Holt, C. Functional and dysfunctional folding, association and aggregation of caseins. Adv. Protein Chem. Struct. Biol. 2019, 118, 163–216. [Google Scholar] [CrossRef] [PubMed]
- Permyakov, E.A.; Permyakov, S.E.; Breydo, L.; Redwan, E.M.; Almehdar, H.A.; Uversky, V.N. Disorder in Milk Proteins: Lactalbumin. Part A. Structural Properties and Conformational Behavior. Curr. Protein Pept. Sci. 2016, 17, 352–367. [Google Scholar] [CrossRef] [PubMed]
- Bourlieu, C.; Michalski, M.-C. Structure–function relationship of the milk fat globule. Curr. Opin. Clin. Nutr. Metab. Care 2015, 18, 118–127. [Google Scholar] [CrossRef] [PubMed]
- Kuczyńska, B.; Puppel, K.; Gołȩbiewski, M.; Metera, E.; Sakowski, T.; Słoniewski, K.; Gołębiewski, M. Differences in whey protein content between cow’s milk collected in late pasture and early indoor feeding season from conventional and organic farms in Poland. J. Sci. Food Agric. 2012, 92, 2899–2904. [Google Scholar] [CrossRef] [PubMed]
- Böttger, I.; Dobbs, R.; Faloona, G.R.; Unger, R.H. The Effects of Triglyceride Absorption upon Glucagon, Insulin, and Gut Glucagon-Like Immunoreactivity. J. Clin. Investig. 1973, 52, 2532–2541. [Google Scholar] [CrossRef]
- Imamura, F.; Micha, R.; Wu, J.H.; de Oliveira Otto, M.C.; Otite, F.O.; Abioye, A.I.; Mozaffarian, D. Effects of Saturated Fat, Polyunsaturated Fat, Monounsaturated Fat, and Carbohydrate on Glucose-Insulin Homeostasis: A Systematic Review and Meta-analysis of Randomised Controlled Feeding Trials. PLoS Med. 2016, 13, e1002087. [Google Scholar] [CrossRef] [PubMed]
- Baggio, L.L.; Drucker, D.J. Biology of incretins: GLP-1 and GIP. Gastroenterology 2007, 132, 2131–2157. [Google Scholar] [CrossRef]
- Krarup, T.; Holst, J.J.; Larsen, K.L. Responses and molecular heterogeneity of IR-GIP after intraduodenal glucose and fat. Am. J. Physiol. Metab. 1985, 249, E195–E200. [Google Scholar] [CrossRef]
- Beysen, C.; Karpe, F.; Fielding, B.A.; Clark, A.; Levy, J.C.; Frayn, K.N. Interaction between specific fatty acids, GLP-1 and insulin secretion in humans. Diabetologia 2002, 45, 1533–1541. [Google Scholar] [CrossRef]
- Rocca, A.S.; LaGreca, J.; Kalitsky, J.; Brubaker, P.L. Monounsaturated Fatty Acid Diets Improve Glycemic Tolerance through Increased Secretion of Glucagon-Like Peptide-1*. Endocrinology 2001, 142, 1148–1155. [Google Scholar] [CrossRef] [PubMed]








| Milk Type | Units | Cow | Hum | Cow [↓Cas] | Hum [↑Cas] |
|---|---|---|---|---|---|
| Milk volumes | mL/Kg BW | ||||
| natural milk | 5.47 ± 0.00 | 4.82 ± 0.07 | 0.50 ± 0.00 | 5.02 ± 0.04 | |
| added water | 0 | 0 | 4.32 ± 0.00 | 0 | |
| Total | 5.47 ± 0.00 | 4.82 ± 0.07 | 4.82 ± 0.00 | 5.16 ± 0.04 | |
| Milk composition | |||||
| Lactose | g/dL | ||||
| endog. | 4.90 ± 0.00 | 7.12 ± 0.19 | 0.48 ± 0.00 | 6.70 ± 0.10 | |
| added | 1.63 ± 0.00 | / | 6.51 ± 0.00 | / | |
| Total | 6.53 ± 0.00 | 7.12 ± 0.19 | 6.99 ± 0.00 | 6.70 ± 0.10 | |
| Protein | g/dL | 3.4 a | 1.02 ± 0.03 | 0.92 ± 0.05 | 3.45 ± 0.02 |
| endog. casein | 2.72 b | 0.41 c | 0.27 ± 0.01 d | 0.40 ± 0.00 c | |
| added casein | 2.45 ± 0.02 | ||||
| Total casein | 2.72 b | 0.41 c | 0.27 ± 0.01 d | 2.85 ± 0.02 c | |
| endog. WP | 0.68 b | 0.61 c | 0.06 ± 0.00 d | 0.08 ± 0.01 c | |
| added WP | 0.59 ± 0.00 | ||||
| Total WP | 0.68 b | 0.61 c | 0.66 ± 0.00 | 0.60 ± 0.01 c | |
| Fat | g/dL | 3.5 ± 0.0 | 2.30 ± 0.26 | 2.20 ± 0.11 | 1.82 ± 0.09 |
| Total energy e | kJ | 226 | 155 | 149 | 166 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tessari, P.; Toffolon, A.; Vettore, M.; Iori, E.; Lante, A.; Feller, E.; Rocco, E.A.; Vedovato, M.; Verlato, G.; Bellettato, M. Neither Incretin or Amino Acid Responses, nor Casein Content, Account for the Equal Insulin Response Following Iso-Lactose Loads of Natural Human and Cow Milk in Healthy Young Adults. Nutrients 2022, 14, 1624. https://doi.org/10.3390/nu14081624
Tessari P, Toffolon A, Vettore M, Iori E, Lante A, Feller E, Rocco EA, Vedovato M, Verlato G, Bellettato M. Neither Incretin or Amino Acid Responses, nor Casein Content, Account for the Equal Insulin Response Following Iso-Lactose Loads of Natural Human and Cow Milk in Healthy Young Adults. Nutrients. 2022; 14(8):1624. https://doi.org/10.3390/nu14081624
Chicago/Turabian StyleTessari, Paolo, Alessandro Toffolon, Monica Vettore, Elisabetta Iori, Anna Lante, Emiliano Feller, Elisabetta Alma Rocco, Monica Vedovato, Giovanna Verlato, and Massimo Bellettato. 2022. "Neither Incretin or Amino Acid Responses, nor Casein Content, Account for the Equal Insulin Response Following Iso-Lactose Loads of Natural Human and Cow Milk in Healthy Young Adults" Nutrients 14, no. 8: 1624. https://doi.org/10.3390/nu14081624
APA StyleTessari, P., Toffolon, A., Vettore, M., Iori, E., Lante, A., Feller, E., Rocco, E. A., Vedovato, M., Verlato, G., & Bellettato, M. (2022). Neither Incretin or Amino Acid Responses, nor Casein Content, Account for the Equal Insulin Response Following Iso-Lactose Loads of Natural Human and Cow Milk in Healthy Young Adults. Nutrients, 14(8), 1624. https://doi.org/10.3390/nu14081624