
The Influence of Ketone Bodies on Circadian Processes Regarding Appetite, Sleep and Hormone Release: A Systematic Review of the Literature

 ,
,  , ,
, ,  , ,
, ,  , , , ,
, , , , 
Abstract
:1. Introduction
2. Materials and Methods
3. Results
3.1. KD and Chrononutrition
3.2. Effect of the KD on Appetite Regulation
3.3. Effect of the KD on Sleep Quality and Duration
3.4. Effect of the KD on Hormone Release
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kuhlman, S.J.; Craig, L.M.; Duffy, J.F. Introduction to Chronobiology. Cold Spring Harb. Perspect. Biol. 2018, 10, a033613. [Google Scholar] [CrossRef] [PubMed]
- Daan, S.; Aschoff, J. The Entrainment of Circadian Systems. In Handbook of Behavioral Neurobiology; Takahashi, J.S., Turek, F.W., Moore, R.Y., Eds.; Springer: Boston, MA, USA, 2001; Volume 12, pp. 7–43. ISBN 978-0-306-46504-8. [Google Scholar]
- Inouye, S.T.; Kawamura, H. Persistence of Circadian Rhythmicity in a Mammalian Hypothalamic “Island” Containing the Suprachiasmatic Nucleus. Proc. Natl. Acad. Sci. USA 1979, 76, 5962–5966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Challet, E. Minireview: Entrainment of the Suprachiasmatic Clockwork in Diurnal and Nocturnal Mammals. Endocrinology 2007, 148, 5648–5655. [Google Scholar] [CrossRef] [Green Version]
- Daan, S.; Aschoff, J. Circadian Contributions to Survival. In Vertebrate Circadian Systems; Aschoff, J., Daan, S., Groos, G.A., Eds.; Proceedings in Life Sciences; Springer: Berlin/Heidelberg, Germany, 1982; pp. 305–321. ISBN 978-3-642-68653-5. [Google Scholar]
- Aschoff, J. Exogenous and Endogenous Components in Circadian Rhythms. Cold Spring Harb. Symp. Quant. Biol. 1960, 25, 11–28. [Google Scholar] [CrossRef] [PubMed]
- Partch, C.L.; Green, C.B.; Takahashi, J.S. Molecular Architecture of the Mammalian Circadian Clock. Trends Cell Biol. 2014, 24, 90–99. [Google Scholar] [CrossRef] [Green Version]
- Johnston, J.D.; Ordovás, J.M.; Scheer, F.A.; Turek, F.W. Circadian Rhythms, Metabolism, and Chrononutrition in Rodents and Humans. Adv. Nutr. 2016, 7, 399–406. [Google Scholar] [CrossRef] [Green Version]
- Sadacca, L.A.; Lamia, K.A.; de Lemos, A.S.; Blum, B.; Weitz, C.J. An Intrinsic Circadian Clock of the Pancreas Is Required for Normal Insulin Release and Glucose Homeostasis in Mice. Diabetologia 2011, 54, 120–124. [Google Scholar] [CrossRef] [Green Version]
- Lamia, K.A.; Storch, K.-F.; Weitz, C.J. Physiological Significance of a Peripheral Tissue Circadian Clock. Proc. Natl. Acad. Sci. USA 2008, 105, 15172–15177. [Google Scholar] [CrossRef] [Green Version]
- Shostak, A.; Meyer-Kovac, J.; Oster, H. Circadian Regulation of Lipid Mobilization in White Adipose Tissues. Diabetes 2013, 62, 2195–2203. [Google Scholar] [CrossRef] [Green Version]
- Dyar, K.A.; Ciciliot, S.; Wright, L.E.; Biensø, R.S.; Tagliazucchi, G.M.; Patel, V.R.; Forcato, M.; Paz, M.I.P.; Gudiksen, A.; Solagna, F.; et al. Muscle Insulin Sensitivity and Glucose Metabolism Are Controlled by the Intrinsic Muscle Clock. Mol. Metab. 2014, 3, 29–41. [Google Scholar] [CrossRef]
- Muscogiuri, G.; Barrea, L.; Laudisio, D.; Pugliese, G.; Salzano, C.; Savastano, S.; Colao, A. The Management of Very Low-Calorie Ketogenic Diet in Obesity Outpatient Clinic: A Practical Guide. J. Transl. Med. 2019, 17, 356. [Google Scholar] [CrossRef] [PubMed]
- Güldür, T.; Otlu, H.G. Circadian Rhythm in Mammals: Time to Eat & Time to Sleep. Biol. Rhythm. Res. 2017, 48, 243–261. [Google Scholar] [CrossRef]
- Watanabe, M.; Tozzi, R.; Risi, R.; Tuccinardi, D.; Mariani, S.; Basciani, S.; Spera, G.; Lubrano, C.; Gnessi, L. Beneficial Effects of the Ketogenic Diet on Nonalcoholic Fatty Liver Disease: A Comprehensive Review of the Literature. Obes. Rev. 2020, 21, e13024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basciani, S.; Camajani, E.; Contini, S.; Persichetti, A.; Risi, R.; Bertoldi, L.; Strigari, L.; Prossomariti, G.; Watanabe, M.; Mariani, S.; et al. Very-Low-Calorie Ketogenic Diets With Whey, Vegetable, or Animal Protein in Patients With Obesity: A Randomized Pilot Study. J. Clin. Endocrinol. Metab. 2020, 105, 2939–2949. [Google Scholar] [CrossRef]
- Bruci, A.; Tuccinardi, D.; Tozzi, R.; Balena, A.; Santucci, S.; Frontani, R.; Mariani, S.; Basciani, S.; Spera, G.; Gnessi, L.; et al. Very Low-Calorie Ketogenic Diet: A Safe and Effective Tool for Weight Loss in Patients with Obesity and Mild Kidney Failure. Nutrients 2020, 12, 333. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, M.; Tuccinardi, D.; Ernesti, I.; Basciani, S.; Mariani, S.; Genco, A.; Manfrini, S.; Lubrano, C.; Gnessi, L. Scientific Evidence Underlying Contraindications to the Ketogenic Diet: An Update. Obes. Rev. 2020, 21, e13053. [Google Scholar] [CrossRef]
- Newman, J.C.; Verdin, E. Ketone Bodies as Signaling Metabolites. Trends Endocrinol. Metab. 2014, 25, 42–52. [Google Scholar] [CrossRef] [Green Version]
- Phinney, S.D.; Bistrian, B.R.; Wolfe, R.R.; Blackburn, G.L. The Human Metabolic Response to Chronic Ketosis without Caloric Restriction: Physical and Biochemical Adaptation. Metabolism 1983, 32, 757–768. [Google Scholar] [CrossRef]
- Nyenwe, E.A.; Kitabchi, A.E. The Evolution of Diabetic Ketoacidosis: An Update of Its Etiology, Pathogenesis and Management. Metabolism 2016, 65, 507–521. [Google Scholar] [CrossRef]
- Regmi, P.; Heilbronn, L.K. Time-Restricted Eating: Benefits, Mechanisms, and Challenges in Translation. iScience 2020, 23, 101161. [Google Scholar] [CrossRef]
- Templeman, I.; Gonzalez, J.T.; Thompson, D.; Betts, J.A. The Role of Intermittent Fasting and Meal Timing in Weight Management and Metabolic Health. Proc. Nutr. Soc. 2020, 79, 76–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Golbidi, S.; Daiber, A.; Korac, B.; Li, H.; Essop, M.F.; Laher, I. Health Benefits of Fasting and Caloric Restriction. Curr. Diabetes Rep. 2017, 17, 123. [Google Scholar] [CrossRef] [PubMed]
- Bostock, E.C.S.; Kirkby, K.C.; Taylor, B.V.M. The Current Status of the Ketogenic Diet in Psychiatry. Front. Psychiatry 2017, 8, 43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paoli, A.; Mancin, L.; Giacona, M.C.; Bianco, A.; Caprio, M. Effects of a Ketogenic Diet in Overweight Women with Polycystic Ovary Syndrome. J. Transl. Med. 2020, 18, 104. [Google Scholar] [CrossRef] [PubMed]
- Basciani, S.; Costantini, D.; Contini, S.; Persichetti, A.; Watanabe, M.; Mariani, S.; Lubrano, C.; Spera, G.; Lenzi, A.; Gnessi, L. Safety and Efficacy of a Multiphase Dietetic Protocol with Meal Replacements Including a Step with Very Low Calorie Diet. Endocrine 2015, 48, 863–870. [Google Scholar] [CrossRef]
- Risi, R.; Tozzi, R.; Watanabe, M. Beyond Weight Loss in Nonalcoholic Fatty Liver Disease: The Role of Carbohydrate Restriction. Curr. Opin. Clin. Nutr. Metab. Care 2021, 24, 349–353. [Google Scholar] [CrossRef]
- Masi, D.; Risi, R.; Basciani, S.; Tuccinardi, D.; Mariani, S.; Lubrano, C.; Gnessi, L.; Watanabe, M. Very Low-Calorie Ketogenic Diets to Treat Patients with Obesity and Chronic Kidney Disease. J. Ren. Nutr. 2021, 31, 340–341. [Google Scholar] [CrossRef]
- Gangitano, E.; Tozzi, R.; Mariani, S.; Lenzi, A.; Gnessi, L.; Lubrano, C. Ketogenic Diet for Obese COVID-19 Patients: Is Respiratory Disease a Contraindication? A Narrative Review of the Literature on Ketogenic Diet and Respiratory Function. Front. Nutr. 2021, 8, 771047. [Google Scholar] [CrossRef]
- Gangitano, E.; Tozzi, R.; Gandini, O.; Watanabe, M.; Basciani, S.; Mariani, S.; Lenzi, A.; Gnessi, L.; Lubrano, C. Ketogenic Diet as a Preventive and Supportive Care for COVID-19 Patients. Nutrients 2021, 13, 1004. [Google Scholar] [CrossRef]
- Liver-Derived Ketone Bodies Are Necessary for Food Anticipation-PubMed. Available online: https://pubmed.ncbi.nlm.nih.gov/26838474/ (accessed on 16 June 2021).
- Tognini, P.; Murakami, M.; Liu, Y.; Eckel-Mahan, K.L.; Newman, J.C.; Verdin, E.; Baldi, P.; Sassone-Corsi, P. Distinct Circadian Signatures in Liver and Gut Clocks Revealed by Ketogenic Diet. Cell Metab. 2017, 26, 523–538.e5. [Google Scholar] [CrossRef]
- Williams, G.; Bing, C.; Cai, X.J.; Harrold, J.A.; King, P.J.; Liu, X.H. The Hypothalamus and the Control of Energy Homeostasis: Different Circuits, Different Purposes. Physiol. Behav. 2001, 74, 683–701. [Google Scholar] [CrossRef]
- Hillebrand, J.J.G.; de Wied, D.; Adan, R.A.H. Neuropeptides, Food Intake and Body Weight Regulation: A Hypothalamic Focus. Peptides 2002, 23, 2283–2306. [Google Scholar] [CrossRef]
- Ueno, H.; Yamaguchi, H.; Kangawa, K.; Nakazato, M. Ghrelin: A Gastric Peptide That Regulates Food Intake and Energy Homeostasis. Regul. Pept. 2005, 126, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Toshinai, K.; Date, Y.; Murakami, N.; Shimada, M.; Mondal, M.S.; Shimbara, T.; Guan, J.-L.; Wang, Q.-P.; Funahashi, H.; Sakurai, T.; et al. Ghrelin-Induced Food Intake Is Mediated via the Orexin Pathway. Endocrinology 2003, 144, 1506–1512. [Google Scholar] [CrossRef] [Green Version]
- Rodin, J.; Wack, J.; Ferrannini, E.; DeFronzo, R.A. Effect of Insulin and Glucose on Feeding Behavior. Metabolism 1985, 34, 826–831. [Google Scholar] [CrossRef]
- Hellström, P.M.; Geliebter, A.; Näslund, E.; Schmidt, P.T.; Yahav, E.K.; Hashim, S.A.; Yeomans, M.R. Peripheral and Central Signals in the Control of Eating in Normal, Obese and Binge-Eating Human Subjects. Br. J. Nutr. 2004, 92, S47–S57. [Google Scholar] [CrossRef] [Green Version]
- Obici, S.; Feng, Z.; Morgan, K.; Stein, D.; Karkanias, G.; Rossetti, L. Central Administration of Oleic Acid Inhibits Glucose Production and Food Intake. Diabetes 2002, 51, 271–275. [Google Scholar] [CrossRef] [Green Version]
- Gil, K.M.; Skeie, B.; Kvetan, V.; Askanazi, J.; Friedman, M.I. Parenteral Nutrition and Oral Intake: Effect of Glucose and Fat Infusions. JPEN J. Parenter. Enter. Nutr. 1991, 15, 426–432. [Google Scholar] [CrossRef]
- Watanabe, M.; Singhal, G.; Fisher, F.M.; Beck, T.C.; Morgan, D.A.; Socciarelli, F.; Mather, M.L.; Risi, R.; Bourke, J.; Rahmouni, K.; et al. Liver-Derived FGF21 Is Essential for Full Adaptation to Ketogenic Diet but Does Not Regulate Glucose Homeostasis. Endocrine 2020, 67, 95–108. [Google Scholar] [CrossRef]
- Di Mauro, A.; Tuccinardi, D.; Watanabe, M.; Del Toro, R.; Monte, L.; Giorgino, R.; Rampa, L.; Rossini, G.; Kyanvash, S.; Soare, A.; et al. The Mediterranean Diet Increases Glucagon-like Peptide 1 and Oxyntomodulin Compared with a Vegetarian Diet in Patients with Type 2 Diabetes: A Randomized Controlled Cross-over Trial. Diabetes/Metab. Res. Rev. 2021, 37, e3406. [Google Scholar] [CrossRef]
- Tuccinardi, D.; Perakakis, N.; Farr, O.M.; Upadhyay, J.; Mantzoros, C.S. Branched-Chain Amino Acids in Relation to Food Preferences and Insulin Resistance in Obese Subjects Consuming Walnuts: A Cross-over, Randomized, Double-Blind, Placebo-Controlled Inpatient Physiology Study. Clin. Nutr. 2021, 40, 3032–3036. [Google Scholar] [CrossRef] [PubMed]
- Moesgaard, S.G.; Ahrén, B.; Carr, R.D.; Gram, D.X.; Brand, C.L.; Sundler, F. Effects of High-Fat Feeding and Fasting on Ghrelin Expression in the Mouse Stomach. Regul. Pept. 2004, 120, 261–267. [Google Scholar] [CrossRef] [PubMed]
- Vestergaard, E.T.; Zubanovic, N.B.; Rittig, N.; Møller, N.; Kuhre, R.E.; Holst, J.J.; Rehfeld, J.F.; Thomsen, H.H. Acute Ketosis Inhibits Appetite and Decreases Plasma Concentrations of Acyl Ghrelin in Healthy Young Men. Diabetes Obes. Metab. 2021, 23, 1834–1842. [Google Scholar] [CrossRef] [PubMed]
- Stubbs, B.J.; Cox, P.J.; Evans, R.D.; Cyranka, M.; Clarke, K.; de Wet, H. A Ketone Ester Drink Lowers Human Ghrelin and Appetite: Exogenous Ketones and Appetite. Obesity 2018, 26, 269–273. [Google Scholar] [CrossRef]
- Sumithran, P.; Prendergast, L.A.; Delbridge, E.; Purcell, K.; Shulkes, A.; Kriketos, A.; Proietto, J. Ketosis and Appetite-Mediating Nutrients and Hormones after Weight Loss. Eur. J. Clin. Nutr. 2013, 67, 759–764. [Google Scholar] [CrossRef] [Green Version]
- Chearskul, S.; Delbridge, E.; Shulkes, A.; Proietto, J.; Kriketos, A. Effect of Weight Loss and Ketosis on Postprandial Cholecystokinin and Free Fatty Acid Concentrations. Am. J. Clin. Nutr. 2008, 87, 1238–1246. [Google Scholar] [CrossRef] [Green Version]
- Yancy, W.S.; Olsen, M.K.; Guyton, J.R.; Bakst, R.P.; Westman, E.C. A Low-Carbohydrate, Ketogenic Diet versus a Low-Fat Diet To Treat Obesity and Hyperlipidemia: A Randomized, Controlled Trial. Ann. Intern. Med. 2004, 140, 769. [Google Scholar] [CrossRef] [PubMed]
- Johnstone, A.M.; Horgan, G.W.; Murison, S.D.; Bremner, D.M.; Lobley, G.E. Effects of a High-Protein Ketogenic Diet on Hunger, Appetite, and Weight Loss in Obese Men Feeding Ad Libitum. Am. J. Clin. Nutr. 2008, 87, 44–55. [Google Scholar] [CrossRef] [Green Version]
- Nickols-Richardson, S.M.; Coleman, M.D.; Volpe, J.J.; Hosig, K.W. Perceived Hunger Is Lower and Weight Loss Is Greater in Overweight Premenopausal Women Consuming a Low-Carbohydrate/High-Protein vs. High-Carbohydrate/Low-Fat Diet. J. Am. Diet. Assoc. 2005, 105, 1433–1437. [Google Scholar] [CrossRef]
- Burley, V.J.; Kreitzman, S.N.; Hill, A.J.; Blundell, J.E. Across-the-Day Monitoring of Mood and Energy Intake before, during, and after a Very-Low-Calorie Diet. Am. J. Clin. Nutr. 1992, 56, 277S–278S. [Google Scholar] [CrossRef]
- Laing, E.E.; Johnston, J.D.; Möller-Levet, C.S.; Bucca, G.; Smith, C.P.; Dijk, D.-J.; Archer, S.N. Exploiting Human and Mouse Transcriptomic Data: Identification of Circadian Genes and Pathways Influencing Health. BioEssays 2015, 37, 544–556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buxton, O.M.; Cain, S.W.; O’Connor, S.P.; Porter, J.H.; Duffy, J.F.; Wang, W.; Czeisler, C.A.; Shea, S.A. Adverse Metabolic Consequences in Humans of Prolonged Sleep Restriction Combined with Circadian Disruption. Sci. Transl. Med. 2012, 4, 129ra43. [Google Scholar] [CrossRef] [Green Version]
- Morris, C.J.; Yang, J.N.; Garcia, J.I.; Myers, S.; Bozzi, I.; Wang, W.; Buxton, O.M.; Shea, S.A.; Scheer, F.A.J.L. Endogenous Circadian System and Circadian Misalignment Impact Glucose Tolerance via Separate Mechanisms in Humans. Proc. Natl. Acad. Sci. USA 2015, 112, E2225–E2234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dibner, C.; Schibler, U. Circadian Timing of Metabolism in Animal Models and Humans. J. Intern. Med. 2015, 277, 513–527. [Google Scholar] [CrossRef]
- Poggiogalle, E.; Lubrano, C.; Gnessi, L.; Marocco, C.; Di Lazzaro, L.; Polidoro, G.; Luisi, F.; Merola, G.; Mariani, S.; Migliaccio, S.; et al. Reduced Sleep Duration Affects Body Composition, Dietary Intake and Quality of Life in Obese Subjects. Eat. Weight. Disord. 2016, 21, 501–505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Binks, H.; Vincent, G.E.; Gupta, C.; Irwin, C.; Khalesi, S. Effects of Diet on Sleep: A Narrative Review. Nutrients 2020, 12, 936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Afaghi, A.; O’Connor, H.; Chow, C.M. High-Glycemic-Index Carbohydrate Meals Shorten Sleep Onset. Am. J. Clin. Nutr. 2007, 85, 426–430. [Google Scholar] [CrossRef]
- De Gasquet, P.; Griglio, S.; Pequignot-Planche, E.; Malewiak, M.I. Diurnal Changes in Plasma and Liver Lipids and Lipoprotein Lipase Activity in Heart and Adipose Tissue in Rats Fed a High and Low Fat Diet. J. Nutr. 1977, 107, 199–212. [Google Scholar] [CrossRef]
- Masino, S.A.; Kawamura, M.; Wasser, C.D.; Wasser, C.A.; Pomeroy, L.T.; Ruskin, D.N. Adenosine, Ketogenic Diet and Epilepsy: The Emerging Therapeutic Relationship between Metabolism and Brain Activity. Curr. Neuropharmacol. 2009, 7, 257–268. [Google Scholar] [CrossRef]
- Radulovacki, M.; Miletich, R.S.; Green, R.D. N6 (L-Phenylisopropyl)Adenosine (L-PIA) Increases Slow-Wave Sleep (S2) and Decreases Wakefulness in Rats. Brain Res. 1982, 246, 178–180. [Google Scholar] [CrossRef]
- Halassa, M.M.; Florian, C.; Fellin, T.; Munoz, J.R.; Lee, S.-Y.; Abel, T.; Haydon, P.G.; Frank, M.G. Astrocytic Modulation of Sleep Homeostasis and Cognitive Consequences of Sleep Loss. Neuron 2009, 61, 213–219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, Z.-Y.; Huang, Z.-L.; Qu, W.-M.; Eguchi, N.; Urade, Y.; Hayaishi, O. An Adenosine A2A Receptor Agonist Induces Sleep by Increasing GABA Release in the Tuberomammillary Nucleus to Inhibit Histaminergic Systems in Rats: CGS21680 Inhibits Histaminergic Systems via GABA. J. Neurochem. 2005, 92, 1542–1549. [Google Scholar] [CrossRef] [PubMed]
- Saper, C.B.; Scammell, T.E.; Lu, J. Hypothalamic Regulation of Sleep and Circadian Rhythms. Nature 2005, 437, 1257–1263. [Google Scholar] [CrossRef] [PubMed]
- Laposky, A.D.; Bradley, M.A.; Williams, D.L.; Bass, J.; Turek, F.W. Sleep-Wake Regulation Is Altered in Leptin-Resistant (Db/Db) Genetically Obese and Diabetic Mice. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 2008, 295, R2059–R2066. [Google Scholar] [CrossRef] [Green Version]
- Kondo, Y.; Chikahisa, S.; Shiuchi, T.; Shimizu, N.; Tanioka, D.; Uguisu, H.; Séi, H. Sleep Profile during Fasting in PPAR-Alpha Knockout Mice. Physiol. Behav. 2020, 214, 112760. [Google Scholar] [CrossRef]
- Chikahisa, S.; Tominaga, K.; Kawai, T.; Kitaoka, K.; Oishi, K.; Ishida, N.; Rokutan, K.; Séi, H. Bezafibrate, a Peroxisome Proliferator-Activated Receptors Agonist, Decreases Body Temperature and Enhances Electroencephalogram Delta-Oscillation during Sleep in Mice. Endocrinology 2008, 149, 5262–5271. [Google Scholar] [CrossRef] [Green Version]
- Chikahisa, S.; Shimizu, N.; Shiuchi, T.; Séi, H. Ketone Body Metabolism and Sleep Homeostasis in Mice. Neuropharmacology 2014, 79, 399–404. [Google Scholar] [CrossRef]
- Phillips, F.; Crisp, A.H.; Mcguinness, B.; Kalucy, E.C.; Chen, C.N.; Koval, J.; Kalucy, R.S.; Lacey, J.H. Isocaloric Diet Changes and Electroencephalographic Sleep. Lancet 1975, 306, 723–725. [Google Scholar] [CrossRef]
- Kwan, R.M.F.; Thomas, S.; Mir, M.A. Effects of a Low Carbohydrate Isoenergetic Diet on Sleep Behavior and Pulmonary Functions in Healthy Female Adult Humans. J. Nutr. 1986, 116, 2393–2402. [Google Scholar] [CrossRef] [Green Version]
- Willi, S.M.; Oexmann, M.J.; Wright, N.M.; Collop, N.A.; Key, L.L. The Effects of a High-Protein, Low-Fat, Ketogenic Diet on Adolescents With Morbid Obesity: Body Composition, Blood Chemistries, and Sleep Abnormalities. Pediatrics 1998, 101, 61–67. [Google Scholar] [CrossRef]
- Hallböök, T.; Lundgren, J.; Rosén, I. Ketogenic Diet Improves Sleep Quality in Children with Therapy-Resistant Epilepsy. Epilepsia 2007, 48, 59–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Afaghi, A.; O’Connor, H.; Chow, C.M. Acute Effects of the Very Low Carbohydrate Diet on Sleep Indices. Nutr. Neurosci. 2008, 11, 146–154. [Google Scholar] [CrossRef] [PubMed]
- Guzel, O.; Uysal, U.; Arslan, N. Efficacy and Tolerability of Olive Oil-Based Ketogenic Diet in Children with Drug-Resistant Epilepsy: A Single Center Experience from Turkey. Eur. J. Paediatr. Neurol. 2019, 23, 143–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Husain, A.M.; Yancy, W.S.; Carwile, S.T.; Miller, P.P.; Westman, E.C. Diet Therapy for Narcolepsy. Neurology 2004, 62, 2300–2302. [Google Scholar] [CrossRef]
- Sofou, K.; Dahlin, M.; Hallböök, T.; Lindefeldt, M.; Viggedal, G.; Darin, N. Ketogenic Diet in Pyruvate Dehydrogenase Complex Deficiency: Short- and Long-Term Outcomes. J. Inherit. Metab. Dis. 2017, 40, 237–245. [Google Scholar] [CrossRef] [Green Version]
- Castro, A.; Gomez-Arbelaez, D.; Crujeiras, A.; Granero, R.; Aguera, Z.; Jimenez-Murcia, S.; Sajoux, I.; Lopez-Jaramillo, P.; Fernandez-Aranda, F.; Casanueva, F. Effect of A Very Low-Calorie Ketogenic Diet on Food and Alcohol Cravings, Physical and Sexual Activity, Sleep Disturbances, and Quality of Life in Obese Patients. Nutrients 2018, 10, 1348. [Google Scholar] [CrossRef] [Green Version]
- Iacovides, S.; Goble, D.; Paterson, B.; Meiring, R.M. Three Consecutive Weeks of Nutritional Ketosis Has No Effect on Cognitive Function, Sleep, and Mood Compared with a High-Carbohydrate, Low-Fat Diet in Healthy Individuals: A Randomized, Crossover, Controlled Trial. Am. J. Clin. Nutr. 2019, 110, 349–357. [Google Scholar] [CrossRef]
- Siegmann, M.J.; Athinarayanan, S.J.; Hallberg, S.J.; McKenzie, A.L.; Bhanpuri, N.H.; Campbell, W.W.; McCarter, J.P.; Phinney, S.D.; Volek, J.S.; Van Dort, C.J. Improvement in Patient-Reported Sleep in Type 2 Diabetes and Prediabetes Participants Receiving a Continuous Care Intervention with Nutritional Ketosis. Sleep Med. 2019, 55, 92–99. [Google Scholar] [CrossRef]
- Klement, R.J.; Weigel, M.M.; Sweeney, R.A. A Ketogenic Diet Consumed during Radiotherapy Improves Several Aspects of Quality of Life and Metabolic Health in Women with Breast Cancer. Clin. Nutr. 2021, 40, 4267–4274. [Google Scholar] [CrossRef]
- Hofstra, W.A.; de Weerd, A.W. How to Assess Circadian Rhythm in Humans: A Review of Literature. Epilepsy Behav. 2008, 13, 438–444. [Google Scholar] [CrossRef]
- Caputo, M.; Pigni, S.; Agosti, E.; Daffara, T.; Ferrero, A.; Filigheddu, N.; Prodam, F. Regulation of GH and GH Signaling by Nutrients. Cells 2021, 10, 1376. [Google Scholar] [CrossRef] [PubMed]
- Mohd Azmi, N.A.S.; Juliana, N.; Azmani, S.; Mohd Effendy, N.; Abu, I.F.; Mohd Fahmi Teng, N.I.; Das, S. Cortisol on Circadian Rhythm and Its Effect on Cardiovascular System. Int. J. Environ. Res. Public Health 2021, 18, 676. [Google Scholar] [CrossRef] [PubMed]
- Ikegami, K.; Refetoff, S.; Van Cauter, E.; Yoshimura, T. Interconnection between Circadian Clocks and Thyroid Function. Nat. Rev. Endocrinol. 2019, 15, 590–600. [Google Scholar] [CrossRef] [PubMed]
- Filicori, M.; Santoro, N.; Merriam, G.R.; Crowley, W.F. Characterization of the Physiological Pattern of Episodic Gonadotropin Secretion throughout the Human Menstrual Cycle. J. Clin. Endocrinol. Metab. 1986, 62, 1136–1144. [Google Scholar] [CrossRef]
- Klingman, K.M.; Marsh, E.E.; Klerman, E.B.; Anderson, E.J.; Hall, J.E. Absence of Circadian Rhythms of Gonadotropin Secretion in Women. J. Clin. Endocrinol. Metab. 2011, 96, 1456–1461. [Google Scholar] [CrossRef]
- Miyatake, A.; Morimoto, Y.; Oishi, T.; Hanasaki, N.; Sugita, Y.; Iijima, S.; Teshima, Y.; Hishikawa, Y.; Yamamura, Y. Circadian Rhythm of Serum Testosterone and Its Relation to Sleep: Comparison with the Variation in Serum Luteinizing Hormone, Prolactin, and Cortisol in Normal Men. J. Clin. Endocrinol. Metab. 1980, 51, 1365–1371. [Google Scholar] [CrossRef]
- Sassin, J.F.; Frantz, A.G.; Weitzman, E.D.; Kapen, S. Human Prolactin: 24-Hour Pattern with Increased Release Doring Sleep. Science 1972, 177, 1205–1207. [Google Scholar] [CrossRef]
- Egli, M.; Leeners, B.; Kruger, T.H.C. Prolactin Secretion Patterns: Basic Mechanisms and Clinical Implications for Reproduction. Reproduction 2010, 140, 643–654. [Google Scholar] [CrossRef] [Green Version]
- Brzezinski, A. Melatonin in Humans. N. Engl. J. Med. 1997, 336, 186–195. [Google Scholar] [CrossRef]
- Hardeland, R. Chronobiology of Melatonin beyond the Feedback to the Suprachiasmatic Nucleus—Consequences to Melatonin Dysfunction. Int. J. Mol. Sci. 2013, 14, 5817–5841. [Google Scholar] [CrossRef] [Green Version]
- Ekmekcioglu, C. Melatonin Receptors in Humans: Biological Role and Clinical Relevance. Biomed. Pharmacother. 2006, 60, 97–108. [Google Scholar] [CrossRef] [PubMed]
- Zisapel, N. New Perspectives on the Role of Melatonin in Human Sleep, Circadian Rhythms and Their Regulation: Melatonin in Human Sleep and Circadian Rhythms. Br. J. Pharmacol. 2018, 175, 3190–3199. [Google Scholar] [CrossRef] [PubMed]
- Fischer, C.; Mueller, T.; Pfeffer, M.; Wicht, H.; von Gall, C.; Korf, H.-W. Melatonin Receptor 1 Deficiency Affects Feeding Dynamics and Pro-Opiomelanocortin Expression in the Arcuate Nucleus and Pituitary of Mice. Neuroendocrinology 2017, 105, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Ríos-Lugo, M.J.; Jiménez-Ortega, V.; Cano-Barquilla, P.; Mateos, P.F.; Spinedi, E.J.; Cardinali, D.P.; Esquifino, A.I. Melatonin Counteracts Changes in Hypothalamic Gene Expression of Signals Regulating Feeding Behavior in High-Fat Fed Rats. Horm. Mol. Biol. Clin. Investig. 2015, 21, 175–183. [Google Scholar] [CrossRef] [Green Version]
- Meng, X.; Li, Y.; Li, S.; Zhou, Y.; Gan, R.-Y.; Xu, D.-P.; Li, H.-B. Dietary Sources and Bioactivities of Melatonin. Nutrients 2017, 9, 367. [Google Scholar] [CrossRef] [Green Version]
- Moran, L.J.; Ko, H.; Misso, M.; Marsh, K.; Noakes, M.; Talbot, M.; Frearson, M.; Thondan, M.; Stepto, N.; Teede, H.J. Dietary Composition in the Treatment of Polycystic Ovary Syndrome: A Systematic Review to Inform Evidence-Based Guidelines. J. Acad. Nutr. Diet. 2013, 113, 520–545. [Google Scholar] [CrossRef]
- Mavropoulos, J.; Yancy, W.; Hepburn, J.; Westman, E. The effects of a low-carbohydrate, ketogenic diet on the polycystic ovary syndrome: A pilot study. Nutr. Metab. 2005, 2, 35. [Google Scholar] [CrossRef] [Green Version]
- Palomba, S.; Giallauria, F.; Falbo, A.; Russo, T.; Oppedisano, R.; Tolino, A.; Colao, A.; Vigorito, C.; Zullo, F.; Orio, F. Structured Exercise Training Programme versus Hypocaloric Hyperproteic Diet in Obese Polycystic Ovary Syndrome Patients with Anovulatory Infertility: A 24-Week Pilot Study. Hum. Reprod. 2008, 23, 642–650. [Google Scholar] [CrossRef]
- Volek, J.S.; Freidenreich, D.J.; Saenz, C.; Kunces, L.J.; Creighton, B.C.; Bartley, J.M.; Davitt, P.M.; Munoz, C.X.; Anderson, J.M.; Maresh, C.M.; et al. Metabolic Characteristics of Keto-Adapted Ultra-Endurance Runners. Metabolism 2016, 65, 100–110. [Google Scholar] [CrossRef] [Green Version]
- Wilson, J.M.; Lowery, R.P.; Roberts, M.D.; Sharp, M.H.; Joy, J.M.; Shields, K.A.; Partl, J.M.; Volek, J.S.; D’Agostino, D.P. Effects of Ketogenic Dieting on Body Composition, Strength, Power, and Hormonal Profiles in Resistance Training Men. J. Strength Cond. Res. 2020, 34, 3463–3474. [Google Scholar] [CrossRef]
- Vidić, V.; Ilić, V.; Toskić, L.; Janković, N.; Ugarković, D. Effects of Calorie Restricted Low Carbohydrate High Fat Ketogenic vs. Non-Ketogenic Diet on Strength, Body-Composition, Hormonal and Lipid Profile in Trained Middle-Aged Men. Clin. Nutr. 2021, 40, 1495–1502. [Google Scholar] [CrossRef] [PubMed]
- Volek, J.S.; Sharman, M.J.; Love, D.M.; Avery, N.G.; Goacutemez, A.L.; Scheett, T.P.; Kraemer, W.J. Body Composition and Hormonal Responses to a Carbohydrate-Restricted Diet. Metabolism 2002, 51, 864–870. [Google Scholar] [CrossRef] [PubMed]
- Mongioì, L.M.; Cimino, L.; Condorelli, R.A.; Magagnini, M.C.; Barbagallo, F.; Cannarella, R.; La Vignera, S.; Calogero, A.E. Effectiveness of a Very Low Calorie Ketogenic Diet on Testicular Function in Overweight/Obese Men. Nutrients 2020, 12, 2967. [Google Scholar] [CrossRef]
- Rhodes, M.E.; Talluri, J.; Harney, J.P.; Frye, C.A. Ketogenic Diet Decreases Circulating Concentrations of Neuroactive Steroids of Female Rats. Epilepsy Behav. 2005, 7, 231–239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polito, R.; Messina, G.; Valenzano, A.; Scarinci, A.; Villano, I.; Monda, M.; Cibelli, G.; Porro, C.; Pisanelli, D.; Monda, V.; et al. The Role of Very Low Calorie Ketogenic Diet in Sympathetic Activation through Cortisol Secretion in Male Obese Population. J. Clin. Med. 2021, 10, 4230. [Google Scholar] [CrossRef]
- Langfort, J.; Pilis, W.; Zarzeczny, R.; Nazar, K.; Kaciuba-Uściłko, H. Effect of Low-Carbohydrate-Ketogenic Diet on Metabolic and Hormonal Responses to Graded Exercise in Men. J. Physiol. Pharmacol. 1996, 47, 361–371. [Google Scholar]
- Stimson, R.H.; Johnstone, A.M.; Homer, N.Z.M.; Wake, D.J.; Morton, N.M.; Andrew, R.; Lobley, G.E.; Walker, B.R. Dietary Macronutrient Content Alters Cortisol Metabolism Independently of Body Weight Changes in Obese Men. J. Clin. Endocrinol. Metab. 2007, 92, 4480–4484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hainer, V.; Stich, V.; Kunesová, M.; Parízková, J.; Zák, A.; Wernischová, V.; Hrabák, P. Effect of 4-Wk Treatment of Obesity by Very-Low-Calorie Diet on Anthropometric, Metabolic, and Hormonal Indexes. Am. J. Clin. Nutr. 1992, 56, 281S–282S. [Google Scholar] [CrossRef]
- Kaptein, E.M.; Fisler, J.S.; Duda, M.J.; Nicoloff, J.T.; Drenick, E.J. Relationship Between the Changes in Serum Thyroid Hormone Levels and Protein Status during Prolonged Protein Supplemented Caloric Deprivation. Clin. Endocrinol. 1985, 22, 1–15. [Google Scholar] [CrossRef]
- Bielohuby, M.; Sawitzky, M.; Stoehr, B.J.M.; Stock, P.; Menhofer, D.; Ebensing, S.; Bjerre, M.; Frystyk, J.; Binder, G.; Strasburger, C.; et al. Lack of Dietary Carbohydrates Induces Hepatic Growth Hormone (GH) Resistance in Rats. Endocrinology 2011, 152, 1948–1960. [Google Scholar] [CrossRef] [Green Version]
- Spulber, G.; Spulber, S.; Hagenäs, L.; Åmark, P.; Dahlin, M. Growth Dependence on Insulin-like Growth Factor-1 during the Ketogenic Diet: Growth and IGF-I During Ketogenic Diet. Epilepsia 2009, 50, 297–303. [Google Scholar] [CrossRef] [PubMed]
- Eckel-Mahan, K.L.; Patel, V.R.; de Mateo, S.; Orozco-Solis, R.; Ceglia, N.J.; Sahar, S.; Dilag-Penilla, S.A.; Dyar, K.A.; Baldi, P.; Sassone-Corsi, P. Reprogramming of the Circadian Clock by Nutritional Challenge. Cell 2013, 155, 1464–1478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hatori, M.; Vollmers, C.; Zarrinpar, A.; DiTacchio, L.; Bushong, E.A.; Gill, S.; Leblanc, M.; Chaix, A.; Joens, M.; Fitzpatrick, J.A.J.; et al. Time-Restricted Feeding without Reducing Caloric Intake Prevents Metabolic Diseases in Mice Fed a High-Fat Diet. Cell Metab. 2012, 15, 848–860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McHill, A.W.; Phillips, A.J.; Czeisler, C.A.; Keating, L.; Yee, K.; Barger, L.K.; Garaulet, M.; Scheer, F.A.; Klerman, E.B. Later Circadian Timing of Food Intake Is Associated with Increased Body Fat. Am. J. Clin. Nutr. 2017, 106, 1213–1219. [Google Scholar] [CrossRef] [PubMed]
- Dashti, H.S.; Gómez-Abellán, P.; Qian, J.; Esteban, A.; Morales, E.; Scheer, F.A.J.L.; Garaulet, M. Late Eating Is Associated with Cardiometabolic Risk Traits, Obesogenic Behaviors, and Impaired Weight Loss. Am. J. Clin. Nutr. 2021, 113, 154–161. [Google Scholar] [CrossRef]
- Gangitano, E.; Gnessi, L.; Lenzi, A.; Ray, D. Chronobiology and Metabolism: Is Ketogenic Diet Able to Influence Circadian Rhythm? Front. Neurosci. 2021, 15, 756970. [Google Scholar] [CrossRef]
- Shimazu, T.; Hirschey, M.D.; Newman, J.; He, W.; Shirakawa, K.; Le Moan, N.; Grueter, C.A.; Lim, H.; Saunders, L.R.; Stevens, R.D.; et al. Suppression of Oxidative Stress by β-Hydroxybutyrate, an Endogenous Histone Deacetylase Inhibitor. Science 2013, 339, 211–214. [Google Scholar] [CrossRef] [Green Version]
- Pivovarova, O.; Jürchott, K.; Rudovich, N.; Hornemann, S.; Ye, L.; Möckel, S.; Murahovschi, V.; Kessler, K.; Seltmann, A.-C.; Maser-Gluth, C.; et al. Changes of Dietary Fat and Carbohydrate Content Alter Central and Peripheral Clock in Humans. J. Clin. Endocrinol. Metab. 2015, 100, 2291–2302. [Google Scholar] [CrossRef] [Green Version]
- Murakami, M.; Tognini, P. Molecular Mechanisms Underlying the Bioactive Properties of a Ketogenic Diet. Nutrients 2022, 14, 782. [Google Scholar] [CrossRef]
- Chaix, A.; Panda, S. Ketone Bodies Signal Opportunistic Food-Seeking Activity. Trends Endocrinol. Metab. 2016, 27, 350–352. [Google Scholar] [CrossRef] [Green Version]
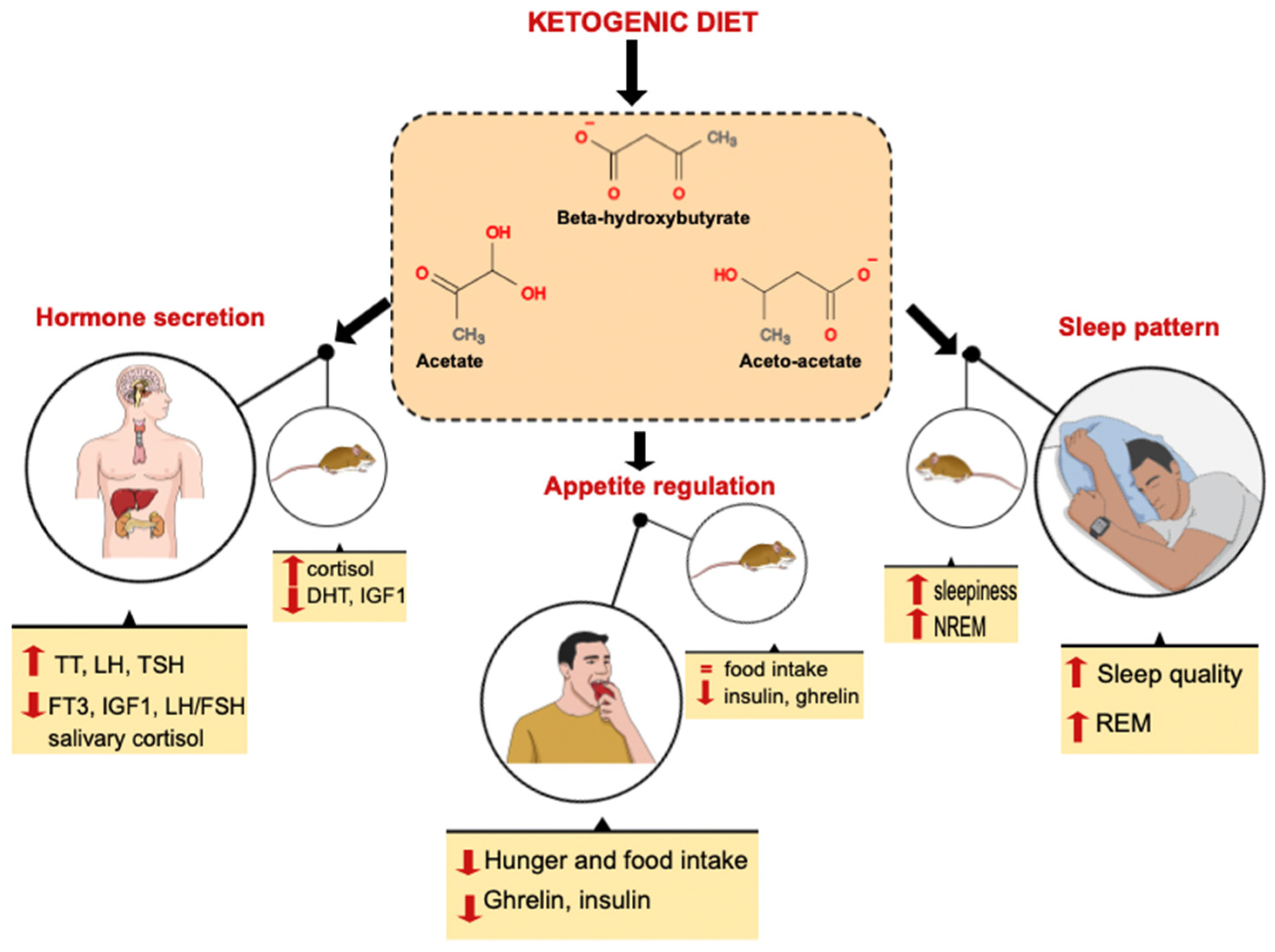

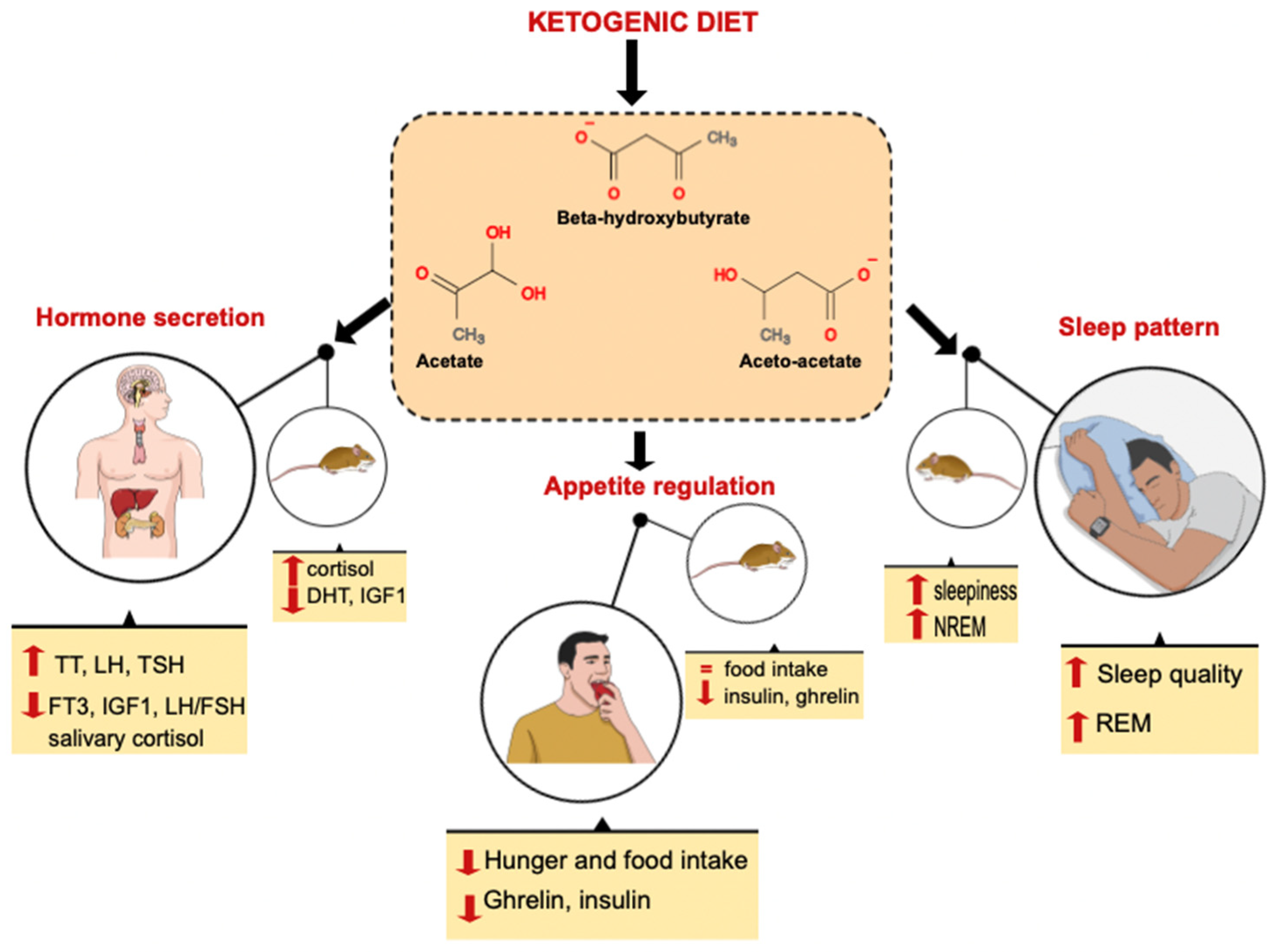{kind=link}
| First Author | Year | Study Design | Species | Duration | Number of Subjects n (f) | Type of Diet | Control | Results |
|---|---|---|---|---|---|---|---|---|
| Burley | 1992 | CT | Humans | 2 wk | 8 (0) | VLCKD | none | ↓ hunger, ↓ irritability, ↓ urge to eat during the second week of VLCKD and the post-diet week |
| Boden | 2005 | CT | Humans | 2 wk | 10 (7) | HFKD | none | spontaneously ↓ energy intake, =level of hunger and satisfaction, ↓leptin, ↑ghrelin, ↓insulin |
| Chearskul | 2008 | CT | Humans | 8 wk | 12 (0) | VLCKD | none | =levels of CCK and FFA during ketosis,↓ postprandial CCK and FFA concentrations after weight loss. ↓ hunger during ketosis, =at week 0 and 1 week after diet |
| Vestergaard | 2021 | RCT | Humans | Single adm | 10 (0) | KE | placebo, glucose | ↓ hunger and prospective food consumption, ghrelin and GLP-1 |
| Sumithran | 2013 | CT | Humans | 8 wk | 39 (n.a.) | VLCKD | none | during ketosis = ghrelin, ↓ leptin and subjective ratings of appetite |
| Johnston | 2006 | RCT | Humans | 6 wk | 19 (15) | VLCKD | non ketogenic low carbohydrate (157 g) | =hunger |
| Johnstone | 2008 | RCT | Humans | 4 wk | 17 (0) | HFKD | medium carbohydrate intake non ketogenic diet (35% carbohydrate) | ↓ hunger and food intake |
| Nickols-Richardson | 2005 | RCT | Humans | 6 wk | 28 (28) | HFKD | high-carbohydrate/low-fat | ↓ hunger |
| Johnston | 2004 | RCT | Human | 6 wk | 16 (n.a.) | HPLFKD | high-carbohydrate/low-fat (280 g) | ↑ satisfaction |
| Vancy | 2004 | RCT | Humans | 24 wk | 79 (n.a.) | HFKD | low fat diet | ↓ energy intake |
| Stubbs | 2017 | RCT | Humans | Single adm | 15 (5) | KE | isocaloric dextrose | ↓ of hunger and plasma insulin, ghrelin, GLP-1, and PYY levels |
| First Author | Year | Study Design | Species | Duration | Number of Subjects n (f) | Type of Diet | Control | Results |
|---|---|---|---|---|---|---|---|---|
| Phillips | 1975 | RCT | human | 4 d | 8 (0) | low-carbohydrate high-fat diet | HCLFD | ↑ REM sleep |
| Kwan | 1986 | CT | human | 1 wk | 6 (6) | low-carbohydrate isoenergetic diet | none | ↑ REM sleep |
| Willi | 1998 | CT | human | 8 wk | 6 (3) adolescents with obesity | HPLFKD | none | ↑ REM sleep, ↓ SWS |
| Husain | 2004 | CT | human | 8 wk | 9 (1) with narcolepsy | LCD | none | improve sleepiness, sleep attacks and sleep paralysis |
| Hallböök | 2007 | CT | human | 3–12 m | 18 (9) children with therapy-resistant epilepsy | HFKD | none | ↓ total sleep, =SWS, ↑ REM sleep, ↓ sleep stage 2, =stage 1 |
| Afaghi | 2008 | CT | human | 48 h | 14 (0) | LCD | none | ↑ SWS, ↓ REM |
| Sofou | 2017 | CT | human | 6 m–6 y | 19 (16) children with PDC deficiency | KD | none | improve nocturnal awakenings and daytime somnolence |
| Guzel | 2018 | single-center, prospective study | human | 3–24 m | 389 (187) children with drug-resistant epilepsy | olive oil-based KD | none | sleep disturbances in 20% |
| Castro | 2018 | LCT | human | 60–90 d | 20 (12) with obesity | VLCKD | none | improvement in sleepiness, =sleep quality, =duration |
| Iacovides | 2019 | RCT crossover, | human | 3 wk | 11 (10) | HFKD | HCLFD | =sleep quality, =morning vigilance |
| Siegmann | 2019 | NRC longitudinal study | human | n.a. | 378 (259) with T2D and obesity | VLCKD | none | ↑ sleep quality |
| Klement | 2021 | CT | human | n.a. | 29 (29) women with early-stage breast cancer undergoing radiotherapy | HFKD | none | ↑ sleep quality |
| First Author | Year | Study Design | Species | Duration | Number of Subjects n(f) | Type of Diet | Control | Results |
|---|---|---|---|---|---|---|---|---|
| Mavropoulos | 2005 | LCT | Human | 24 wk | 11 (11) | VLCKD | none | ↓ percent FT (−22%), ↓ LH/FSH ratio (−36%) |
| Wilson | 2020 | RCT | Human | 10 wk | 25 (0) | Isocaloric KD | WD | ↑ TT |
| Paoli | 2021 | RCT | Human | 8 wk | 19(0) | VLCKD | WD | ↓ TT and IGF1 |
| Mongioì | 2020 | LCT | Human | at least 8 (13.5 ± 0.83 wk) | 40 (0) | VLCKD | none | ↑ TT and LH |
| Rhodes | 2005 | RCT | Dawley rats | 6 wk | 36 (36) | KD | ad libitum chow | ↓DHT |
| Vidić | 2021 | RCT | Human | 8 wk | 20 (0) | HFKD | iso energetic NKLCHFD | ↑ TT and FT |
| Kose | 2017 | CT | Human | 14.7 ± 3.6 wk | 120 (57) | HFKD | none | ↑ TSH, ↓ FT3, ↓ FT4 |
| Iacovides | 2018 | RCT | Human | ongoing | 40 (20) | VLCKD | HCLF | ongoing |
| Volek | 2002 | CT | Human | 6 wk | 12 (0) | VLCKD | ND | ↑ FT4; ≈TT, FT, SHBG, cortisol or FT3 |
| Kaptein | 1985 | LCT | Human | 40 d | 10 (0) | HPKD | none | ≈TSH, ↓ FT3, ↑ FT4, ↑rT3 |
| Van Dam | 2002 | LCT | Human | 7 d | 15 (15) | VLCKD | none | ↑ LH |
| Paoli | 2020 | PCT | Human | 12 wk | 14 (20) | LCKD | none | ↓ LH/FSH ratio, ↓FT, ↑estradiol, ↑progesterone, SHBG |
| Langfort | 1996 | CT | Human | 3 d | 8 (0) | HFKD | mixed diet | ↑ cortisol |
| Stimson | 2007 | RCT | Human | 4 wk | 17 (0) | HFKD | MFMCD | ↑ cortisol; ↓ 5alpha- and 5beta-reduced 24H-cortisol metabolites |
| Bielohuby | 2011 | 3 arms CT | Wistar rats | 4 wk | 16 (0) | HFKD | normal CH and LCNKD | ↓ IGF1, ↓ GHR, ↓ IGFBP-3 |
| Spulber | 2009 | LCT | Human | 3–6 m | 22 (9) | LCKD | none | ↓ IGF1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Masi, D.; Spoltore, M.E.; Rossetti, R.; Watanabe, M.; Tozzi, R.; Caputi, A.; Risi, R.; Balena, A.; Gandini, O.; Mariani, S.; et al. The Influence of Ketone Bodies on Circadian Processes Regarding Appetite, Sleep and Hormone Release: A Systematic Review of the Literature. Nutrients 2022, 14, 1410. https://doi.org/10.3390/nu14071410
Masi D, Spoltore ME, Rossetti R, Watanabe M, Tozzi R, Caputi A, Risi R, Balena A, Gandini O, Mariani S, et al. The Influence of Ketone Bodies on Circadian Processes Regarding Appetite, Sleep and Hormone Release: A Systematic Review of the Literature. Nutrients. 2022; 14(7):1410. https://doi.org/10.3390/nu14071410
Chicago/Turabian StyleMasi, Davide, Maria Elena Spoltore, Rebecca Rossetti, Mikiko Watanabe, Rossella Tozzi, Alessandra Caputi, Renata Risi, Angela Balena, Orietta Gandini, Stefania Mariani, and et al. 2022. "The Influence of Ketone Bodies on Circadian Processes Regarding Appetite, Sleep and Hormone Release: A Systematic Review of the Literature" Nutrients 14, no. 7: 1410. https://doi.org/10.3390/nu14071410
APA StyleMasi, D., Spoltore, M. E., Rossetti, R., Watanabe, M., Tozzi, R., Caputi, A., Risi, R., Balena, A., Gandini, O., Mariani, S., Spera, G., Gnessi, L., & Lubrano, C. (2022). The Influence of Ketone Bodies on Circadian Processes Regarding Appetite, Sleep and Hormone Release: A Systematic Review of the Literature. Nutrients, 14(7), 1410. https://doi.org/10.3390/nu14071410