
Reduced Abundance of Nitrate-Reducing Bacteria in the Oral Microbiota of Women with Future Preeclampsia



Abstract
:1. Introduction
2. Methods
2.1. Study Population
2.2. Oral DNA Extraction
2.3. 16S rRNA Gene Amplicon Library Preparation and Sequencing
2.4. Bioinformatics Analysis for 16S rRNA
2.4.1. Quality Control (QC)
2.4.2. Reads Processing and Taxonomy Assignment
2.4.3. QPCR Analysis of Species Abundance and Functional Capacity
2.4.4. Statistical Analysis
3. Results
3.1. Clinical Characteristic of Participants
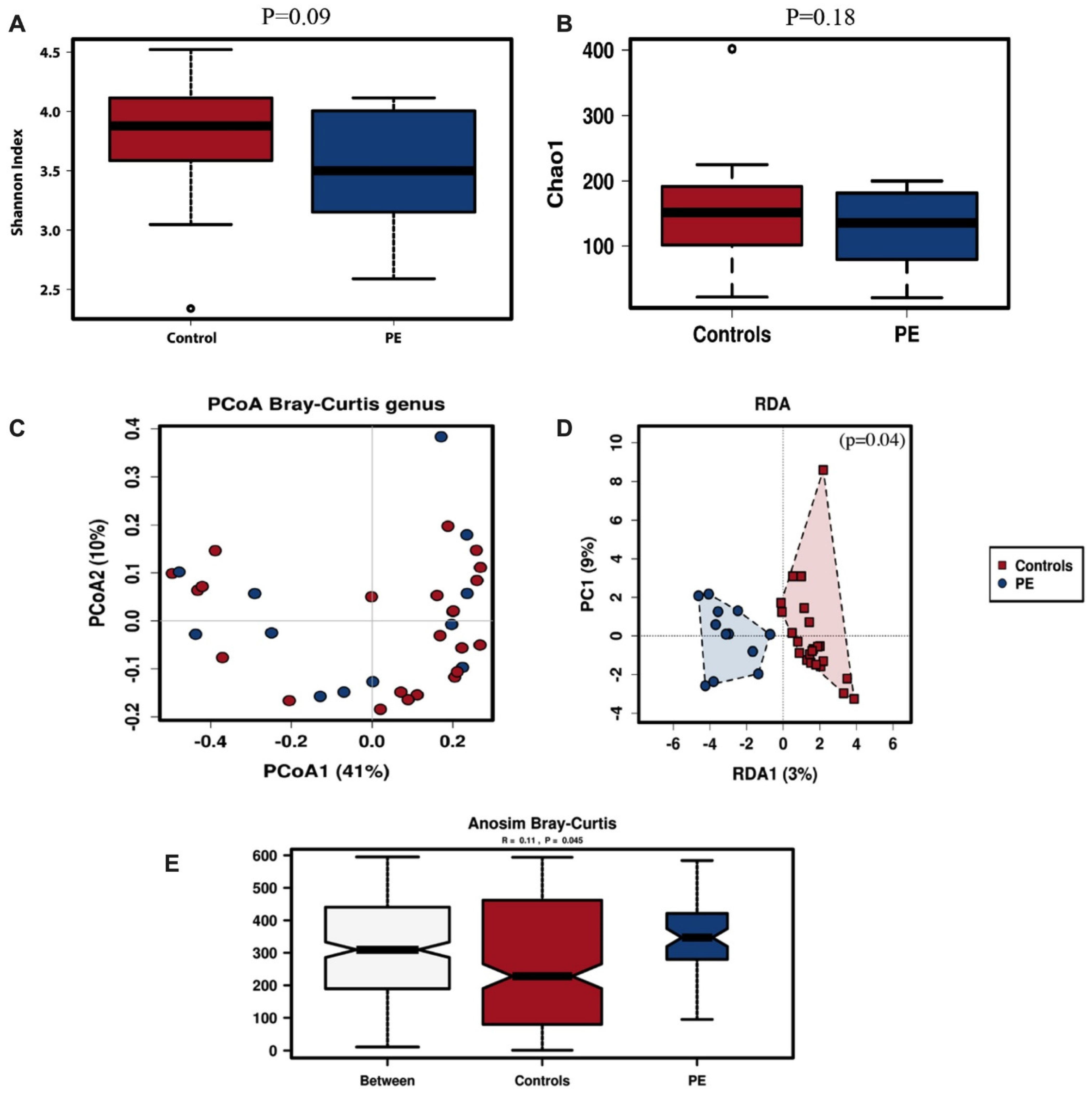
3.2. Oral Microbiota Diversity
3.3. Differences in Oral Microbiota Composition between Normotensive Women and Those Who Developed Preeclampsia
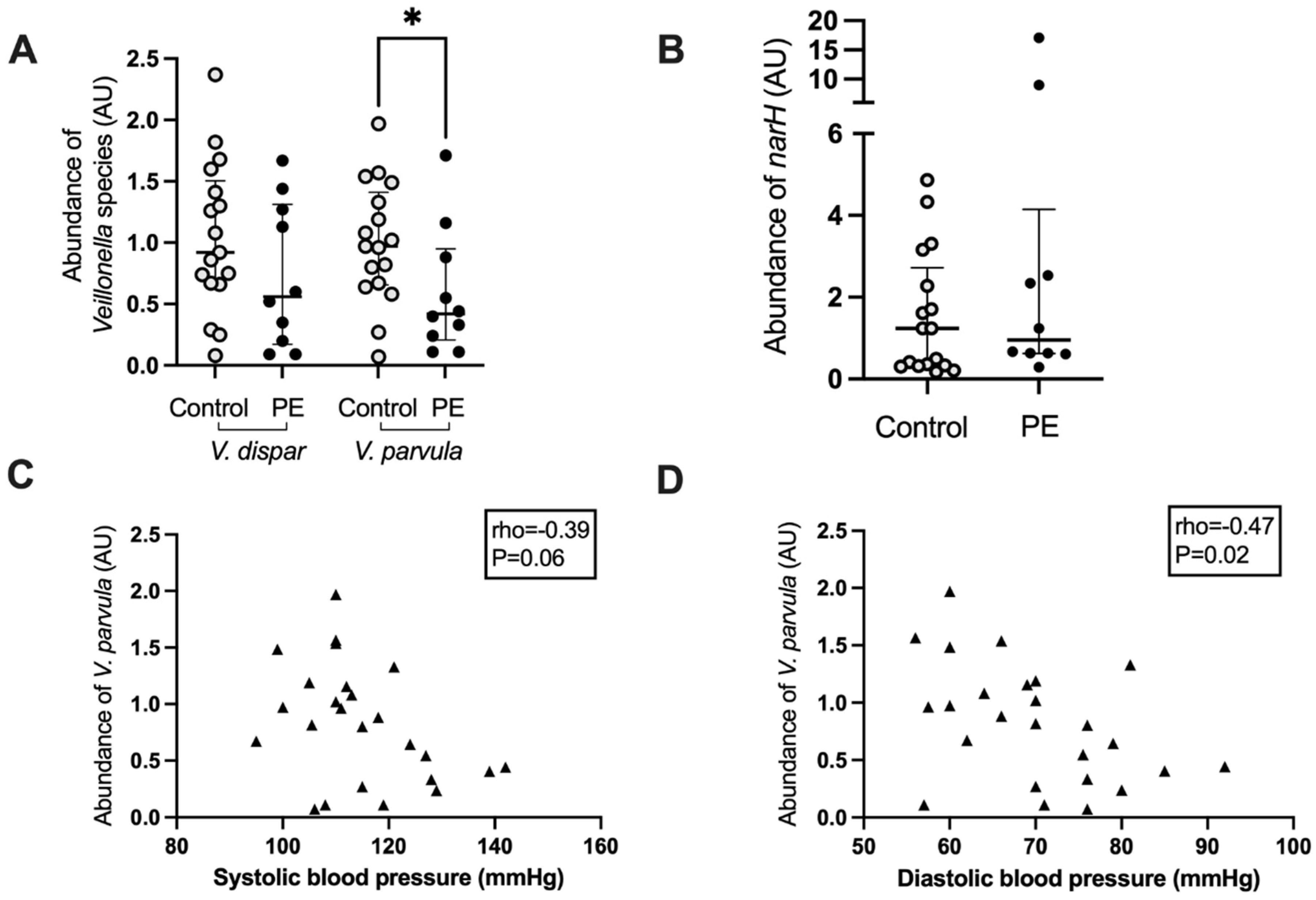
3.4. Correlations between Maternal Blood Pressure and Bacterial Abundance
3.5. Veillonella Species Abundance and Correlation with Blood Pressure
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yu, G.; Phillips, S.; Gail, M.H.; Goedert, J.J.; Humphrys, M.; Ravel, J.; Ren, Y.; Caporaso, N.E. Evaluation of Buccal Cell Samples for Studies of Oral Microbiota. Cancer Epidemiol. Biomark. Prev. 2016, 26, 249–253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, N. Oral Microbiome Metabolism. J. Dent. Res. 2015, 94, 1628–1637. [Google Scholar] [CrossRef] [PubMed]
- Bondonno, C.P.; Liu, A.H.; Croft, K.; Considine, M.; Puddey, I.B.; Woodman, R.; Hodgson, J.M. Antibacterial Mouthwash Blunts Oral Nitrate Reduction and Increases Blood Pressure in Treated Hypertensive Men and Women. Am. J. Hypertens. 2014, 28, 572–575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willmott, T.; McBain, A.J.; Humphreys, G.; Myers, J.; Cottrell, E. Does the Oral Microbiome Play a Role in Hypertensive Pregnancies? Front. Cell. Infect. Microbiol. 2020, 10, 389. [Google Scholar] [CrossRef]
- Begum, S.; Yamasaki, M.; Mochizuki, M. Urinary Levels of Nitric Oxide Metabolites in Normal Pregnancy and Preeclampsia. J. Obstet. Gynaecol. Res. 1996, 22, 551–559. [Google Scholar] [CrossRef]
- Choi, J.W.; Im, M.W.; Pai, S.H. Nitric oxide production increases during normal pregnancy and decreases in preeclampsia. Ann. Clin. Lab. Sci. 2002, 32, 257–263. [Google Scholar]
- Schiessl, B.; Strasburger, C.; Bidlingmaier, M.; Mylonas, I.; Jeschke, U.; Kainer, F.; Friese, K. Plasma- and urine concentrations of nitrite/nitrate and cyclic Guanosinemonophosphate in intrauterine growth restricted and preeclamptic pregnancies. Arch. Gynecol. Obstet. 2006, 274, 150–154. [Google Scholar] [CrossRef]
- Burleigh, M.C.; Liddle, L.; Monaghan, C.; Muggeridge, D.; Sculthorpe, N.; Butcher, J.P.; Henriquez, F.L.; Allen, J.; Easton, C. Salivary nitrite production is elevated in individuals with a higher abundance of oral nitrate-reducing bacteria. Free Radic. Biol. Med. 2018, 120, 80–88. [Google Scholar] [CrossRef] [Green Version]
- Lundberg, J.O.; Weitzberg, E.; Gladwin, M.T. The nitrate–nitrite–nitric oxide pathway in physiology and therapeutics. Nat. Rev. Drug Discov. 2008, 7, 156–167. [Google Scholar] [CrossRef]
- Koopman, J.E.; Buijs, M.J.; Brandt, B.W.; Keijser, B.J.F.; Crielaard, W.; Zaura, E. Nitrate and the Origin of Saliva Influence Composition and Short Chain Fatty Acid Production of Oral Microcosms. Microb. Ecol. 2016, 72, 479–492. [Google Scholar] [CrossRef] [Green Version]
- Bryan, N.S.; Tribble, G.; Angelov, N. Oral Microbiome and Nitric Oxide: The Missing Link in the Management of Blood Pressure. Curr. Hypertens. Rep. 2017, 19, 33. [Google Scholar] [CrossRef] [PubMed]
- Barbadoro, P.; Ponzio, E.; Coccia, E.; Prospero, E.; Santarelli, A.; Rappelli, G.G.; D’Errico, M.M. Association between hypertension, oral microbiome and salivary nitric oxide: A case-control study. Nitric Oxide 2021, 106, 66–71. [Google Scholar] [CrossRef] [PubMed]
- Goh, C.; Trinh, P.; Colombo, P.C.; Genkinger, J.M.; Mathema, B.; Uhlemann, A.; LeDuc, C.; Leibel, R.; Rosenbaum, M.; Paster, B.J.; et al. Association Between Nitrate-Reducing Oral Bacteria and Cardiometabolic Outcomes: Results From ORIGINS. J. Am. Hear. Assoc. 2019, 8, e013324. [Google Scholar] [CrossRef] [PubMed]
- Jang, H.; Patoine, A.; Wu, T.T.; Castillo, D.A.; Xiao, J. Oral microflora and pregnancy: A systematic review and meta-analysis. Sci. Rep. 2021, 11, 16870. [Google Scholar] [CrossRef]
- Herrera, J.A.; Parra, B.; Herrera, E.; Botero, J.E.; Arce, R.M.; Contreras, A.; López-Jaramillo, P. Periodontal disease severity is related to high levels of C-reactive protein in pre-eclampsia. J. Hypertens. 2007, 25, 1459–1464. [Google Scholar] [CrossRef]
- Contreras, A.; Herrera, J.; Soto, J.; Arce, R.M.; Jaramillo, A.; Botero, J. Periodontitis Is Associated With Preeclampsia in Pregnant Women. J. Periodontol. 2006, 77, 182–188. [Google Scholar] [CrossRef] [Green Version]
- Nitert, M.D.; Barrett, H.L.; Foxcroft, K.; Tremellen, A.; Wilkinson, S.; Lingwood, B.; Tobin, J.M.; McSweeney, C.; O’Rourke, P.; McIntyre, H.; et al. SPRING: An RCT study of probiotics in the prevention of gestational diabetes mellitus in overweight and obese women. BMC Pregnancy Childbirth. 2013, 13, 50. [Google Scholar] [CrossRef] [Green Version]
- Lowe, S.A.; Bowyer, L.; Lust, K.; McMahon, L.P.; Morton, M.; North, R.A.; Paech, M.; Said, J. SOMANZ guidelines for the management of hypertensive disorders of pregnancy 2014. Aust. N. Z. J. Obstet. Gynaecol. 2015, 55, e1–e29. [Google Scholar] [CrossRef]
- Hodge, A.; Patterson, A.J.; Brown, W.J.; Ireland, P.; Giles, G. The Anti Cancer Council of Victoria FFQ: Relative validity of nutrient intakes compared with weighed food records in young to middle-aged women in a study of iron supplementation. Aust. N. Z. J. Public Health 2000, 24, 576–583. [Google Scholar] [CrossRef]
- Babraham Bioinformatics-FastQC a Quality Control Tool for High throughput Sequence Data. Available online: https://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 3 November 2020).
- Ewels, P.; Magnusson, M.; Lundin, S.; Käller, M. MultiQC: Summarize analysis results for multiple tools and samples in a single report. Bioinformatics 2016, 32, 3047–3048. [Google Scholar] [CrossRef] [Green Version]
- Martin, M. CUDAPT removes adapter sequences from high-throughput sequencing reads. EMBnet J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef]
- Zakrzewski, M.; Proietti, C.; Ellis, J.; Hasan, S.; Brion, M.-J.; Berger, B.; Krause, L. Calypso: A user-friendly web-server for mining and visualizing microbiome–environment interactions. Bioinformatics 2016, 33, 782–783. [Google Scholar] [CrossRef] [Green Version]
- Doel, J.J.; Benjamin, N.; Hector, M.P.; Rogers, M.; Allaker, R.P. Evaluation of bacterial nitrate reduction in the human oral cavity. Eur. J. Oral Sci. 2005, 113, 14–19. [Google Scholar] [CrossRef]
- Mitsui, T.; Saito, M.; Harasawa, R. Salivary nitrate-nitrite conversion capacity after nitrate ingestion and incidence of Veillonella spp. in elderly individuals. J. Oral Sci. 2018, 60, 405–410. [Google Scholar] [CrossRef] [Green Version]
- Beighton, D.; Clark, D.; Hanakuka, B.; Gilbert, S.; Do, T. The predominant cultivable Veillonella spp. of the tongue of healthy adults identified using rpoB sequencing. Oral Microbiol. Immunol. 2008, 23, 344–347. [Google Scholar] [CrossRef]
- Ormesher, L.; Myers, J.E.; Chmiel, C.; Wareing, M.; Greenwood, S.L.; Tropea, T.; Lundberg, J.O.; Weitzberg, E.; Nihlen, C.; Sibley, C.P.; et al. Effects of dietary nitrate supplementation, from beetroot juice, on blood pressure in hypertensive pregnant women: A randomised, double-blind, placebo-controlled feasibility trial. Nitric Oxide 2018, 80, 37–44. [Google Scholar] [CrossRef]
- Kapil, V.; Khambata, R.; Robertson, A.; Caulfield, M.; Ahluwalia, A. Dietary Nitrate Provides Sustained Blood Pressure Lowering in Hypertensive Patients. Hypertension 2015, 65, 320–327. [Google Scholar] [CrossRef] [Green Version]
- Agrawal, S. Frequency of consumption of specific food items and symptoms of preeclampsia and eclampsia in Indian women. Int. J. Med. Public Health 2014, 4, 350–353. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic | Preeclampsia | Control | p Value |
|---|---|---|---|
| n | 12 | 24 | |
| Age (years) | 32 (27.5–35.5) | 32 (31.5–35.75) | 0.66 |
| BMI (kg/m2) | 38.05 (35.68–43) | 36.35 (34.50–41) | 0.072 |
| SBP (mmHg) * | 128 (112–130) # | 110 (104–119) | 0.006 |
| DBP (mmHg) * | 75 (67–83) # | 65 (60–73) | 0.026 |
| Glucose (mmol/L) | 4.2 (4.1–4.5) | 4.3 (4.0–4.5) | 0.99 |
| HbA1c (%) | 5.4 (4.9–5.6) | 5.1 (4.9–5.3) | 0.23 |
| C peptide (nmol/L) | 0.6 (0.5–1.0) | 0.5 (0.3–0.6) | 0.068 |
| Insulin (mU/L) | 8.7 (4.5–10.0) | 5.0 (3.4–7.1) | 0.038 |
| Cholesterol (mmol/L) | 6.1 (5.6–7.0) | 6.0 (5.5–6.7) | 0.62 |
| Triglyceride (mmol/L) | 2.2 (1.8–2.9) | 2.1 (1.9–2.9) | 0.86 |
| HDL (mmol/L) | 1.7 (1.3–2.0) | 1.6 (1.4–1.8) | 0.5 |
| LDL (mmol/L) | 3.2 (2.6–4.5) | 3.7 (3.1–4.1) | 0.96 |
| VLDL (mmol/L) | 1.0 (0.9–1.4) | 1.0 (0.8–1.3) | 0.87 |
| Pregnancy outcomes | |||
| Infant birth weight | 3613 (2915–3782) | 3488 (3215–3704) | 0.72 |
| Infant gender (M/F) | 7/5 | 11/13 | 0.48 |
| Delivery mode Vaginal/Caesarean | 5/7 | 13/11 | 0.47 |
| Gestational age at delivery | 39.4 (37.6–39.8) | 39.7 (38.6–40.9) | 0.15 |
| Genus | Systolic Blood Pressure | Diastolic Blood Pressure | ||
|---|---|---|---|---|
| Rho | p-Value | Rho | p-Value | |
| Methanobrevibacter | 0.48 | 0.007 | 0.53 | 0.002 |
| Candidatus Brocadia | 0.48 | 0.007 | - | - |
| Uncultured organism | 0.48 | 0.007 | 0.34 | 0.044 |
| Veillonella | −0.46 | 0.010 | −0.38 | 0.036 |
| Desulfomicrobium | 0.45 | 0.013 | - | - |
| Mycoplasma | 0.44 | 0.014 | 0.38 | 0.037 |
| Actinophytocola | 0.42 | 0.019 | ||
| Lachnospiraceae NK4A136 | 0.41 | 0.025 | 0.47 | 0.008 |
| Roseomonas | −0.39 | 0.031 | - | - |
| Aggregatibacter | −0.38 | 0.037 | - | - |
| Dechloromonas | 0.37 | 0.042 | - | - |
| Prevotella | −0.16 | 0.088 | −0.14 | 0.091 |
| Actinophytocola | - | - | 0.49 | 0.005 |
| Peptococcus | - | - | 0.42 | 0.02 |
| Lysobacter | - | - | 0.39 | 0.035 |
| Ignavibacterium | - | - | 0.39 | 0.035 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Altemani, F.; Barrett, H.L.; Callaway, L.K.; McIntyre, H.D.; Dekker Nitert, M. Reduced Abundance of Nitrate-Reducing Bacteria in the Oral Microbiota of Women with Future Preeclampsia. Nutrients 2022, 14, 1139. https://doi.org/10.3390/nu14061139
Altemani F, Barrett HL, Callaway LK, McIntyre HD, Dekker Nitert M. Reduced Abundance of Nitrate-Reducing Bacteria in the Oral Microbiota of Women with Future Preeclampsia. Nutrients. 2022; 14(6):1139. https://doi.org/10.3390/nu14061139
Chicago/Turabian StyleAltemani, Faisal, Helen L. Barrett, Leonie K. Callaway, H. David McIntyre, and Marloes Dekker Nitert. 2022. "Reduced Abundance of Nitrate-Reducing Bacteria in the Oral Microbiota of Women with Future Preeclampsia" Nutrients 14, no. 6: 1139. https://doi.org/10.3390/nu14061139
APA StyleAltemani, F., Barrett, H. L., Callaway, L. K., McIntyre, H. D., & Dekker Nitert, M. (2022). Reduced Abundance of Nitrate-Reducing Bacteria in the Oral Microbiota of Women with Future Preeclampsia. Nutrients, 14(6), 1139. https://doi.org/10.3390/nu14061139