
Effect of High-Protein Diets on Integrated Myofibrillar Protein Synthesis before Anterior Cruciate Ligament Reconstruction: A Randomized Controlled Pilot Study

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Experimental Design
2.3. Dietary Intake
2.4. Deuterium Oxide Labeling
2.5. Tissue Sampling during ACL Reconstruction
2.6. Myofibrillar Protein Synthesis
2.7. mRNA Expression
2.8. Statistical Analysis
3. Results
3.1. Participants
3.2. Dietary Intake
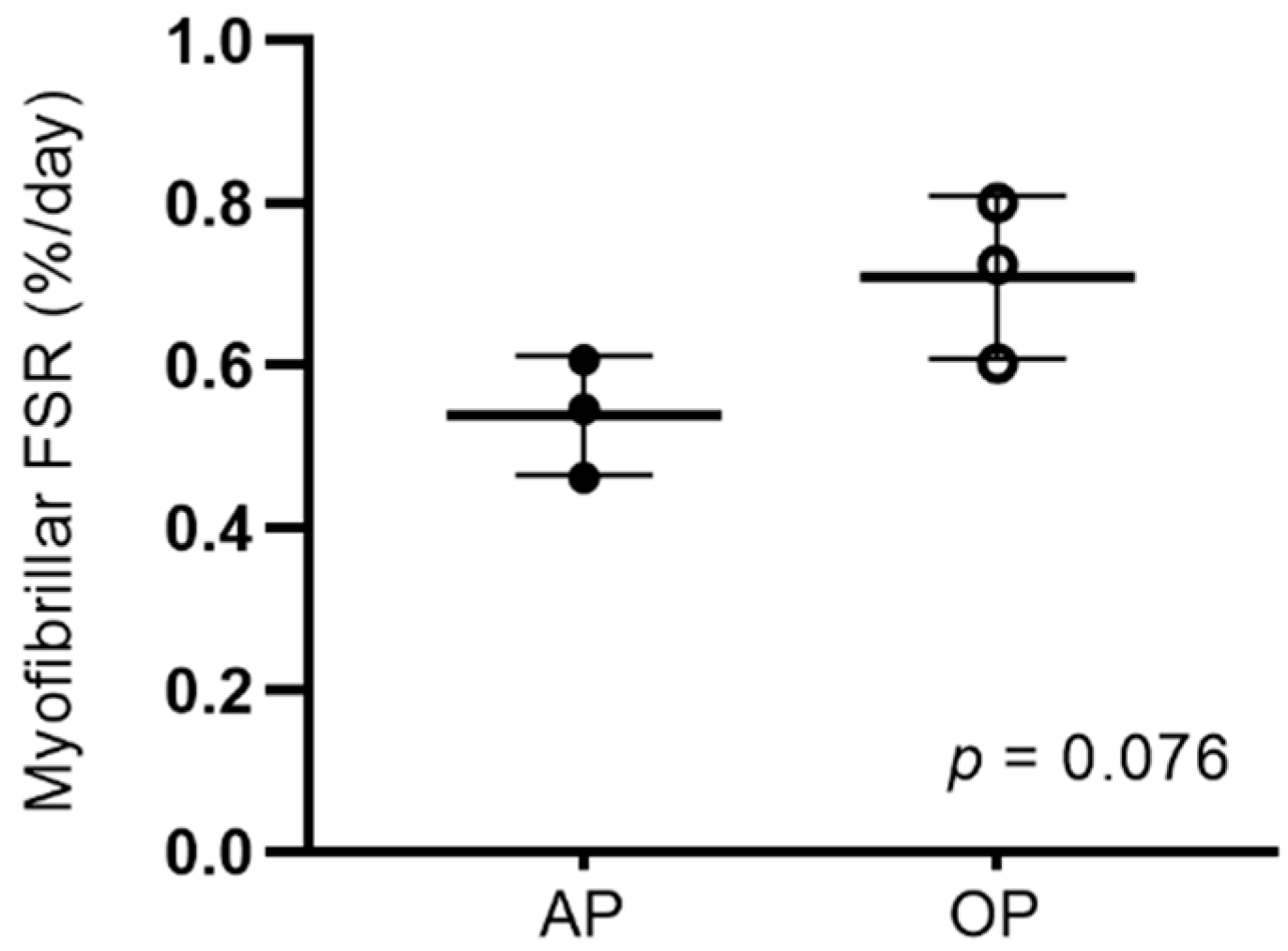
3.3. Myofibrillar Protein Synthesis
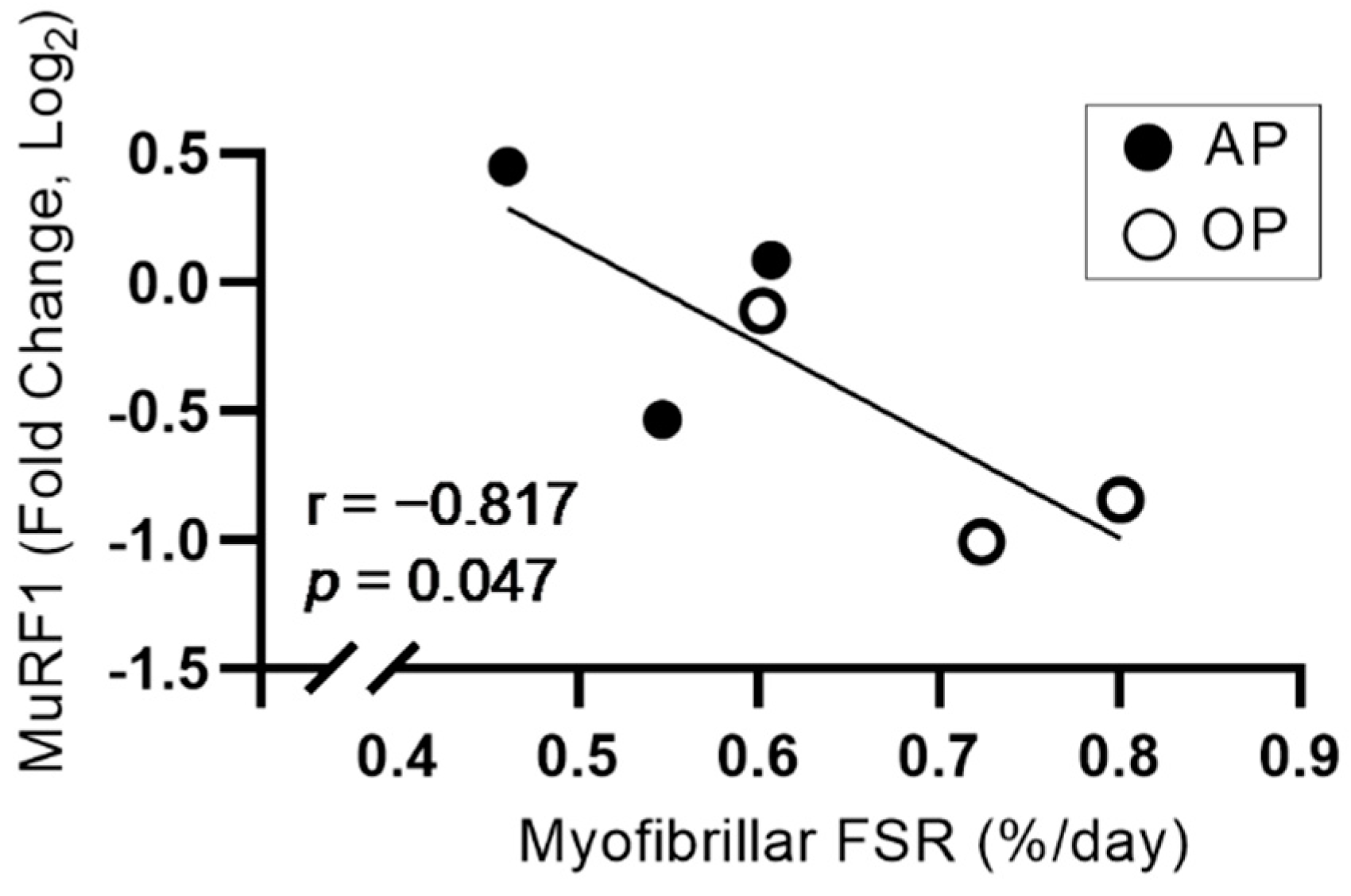
3.4. mRNA Expression
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Norte, G.E.; Knaus, K.R.; Kuenze, C.; Handsfield, G.G.; Meyer, C.H.; Blemker, S.S.; Hart, J.M. MRI-Based Assessment of Lower-Extremity Muscle Volumes in Patients Before and After ACL Reconstruction. J. Sport Rehabil. 2018, 27, 201–212. [Google Scholar] [CrossRef] [PubMed]
- Thomas, A.C.; Wojtys, E.M.; Brandon, C.; Palmieri-Smith, R.M. Muscle atrophy contributes to quadriceps weakness after anterior cruciate ligament reconstruction. J. Sci. Med. Sport 2015, 19, 7–11. [Google Scholar] [CrossRef] [PubMed]
- Rice, D.A.; McNair, P.J. Quadriceps Arthrogenic Muscle Inhibition: Neural Mechanisms and Treatment Perspectives. Semin. Arthritis Rheum. 2010, 40, 250–266. [Google Scholar] [CrossRef] [PubMed]
- Spencer, J.D.; Hayes, K.C.; Alexander, I.J. Knee joint effusion and quadriceps reflex inhibition in man. Arch. Phys. Med. Rehabil. 1984, 65, 171–177. [Google Scholar]
- Kilroe, S.P.; Fulford, J.; Jackman, S.R.; VAN Loon, L.J.C.; Wall, B.T. Temporal Muscle-specific Disuse Atrophy during One Week of Leg Immobilization. Med. Sci. Sports Exerc. 2019, 52, 944–954. [Google Scholar] [CrossRef]
- Wall, B.T.; Dirks, M.; Snijders, T.; Senden, J.M.G.; Dolmans, J.; Van Loon, L.J.C. Substantial skeletal muscle loss occurs during only 5 days of disuse. Acta Physiol. 2013, 210, 600–611. [Google Scholar] [CrossRef]
- Phillips, S.M.; Glover, E.I.; Rennie, M.J. Alterations of protein turnover underlying disuse atrophy in human skeletal muscle. J. Appl. Physiol. 2009, 107, 645–654. [Google Scholar] [CrossRef]
- Wall, B.T.; Snijders, T.; Senden, J.M.G.; Ottenbros, C.L.P.; Gijsen, A.P.; Verdijk, L.; van Loon, L.J. Disuse Impairs the Muscle Protein Synthetic Response to Protein Ingestion in Healthy Men. J. Clin. Endocrinol. Metab. 2013, 98, 4872–4881. [Google Scholar] [CrossRef]
- Wall, B.T.; Dirks, M.; Snijders, T.; van Dijk, J.-W.; Fritisch, M.; Verdijk, L.; Van Loon, L.J.C. Short-term muscle disuse lowers myofibrillar protein synthesis rates and induces anabolic resistance to protein ingestion. Am. J. Physiol. Metab. 2016, 310, E137–E147. [Google Scholar] [CrossRef]
- Kilroe, S.P.; Fulford, J.; Holwerda, A.; Jackman, S.R.; Lee, B.P.; Gijsen, A.P.; van Loon, L.J.; Wall, B.T. Short-term muscle disuse induces a rapid and sustained decline in daily myofibrillar protein synthesis rates. Am. J. Physiol. Metab. 2020, 318, E117–E130. [Google Scholar] [CrossRef]
- Gwin, J.A.; Church, D.D.; Wolfe, R.R.; Ferrando, A.A.; Pasiakos, S.M. Muscle Protein Synthesis and Whole-Body Protein Turnover Responses to Ingesting Essential Amino Acids, Intact Protein, and Protein-Containing Mixed Meals with Considerations for Energy Deficit. Nutrients 2020, 12, 2457. [Google Scholar] [CrossRef] [PubMed]
- Devries, M.C.; McGlory, C.; Bolster, D.R.; Kamil, A.; Rahn, M.; Harkness, L.; Baker, S.K.; Phillips, S. Protein leucine content is a determinant of shorter- and longer-term muscle protein synthetic responses at rest and following resistance exercise in healthy older women: A randomized, controlled trial. Am. J. Clin. Nutr. 2018, 107, 217–226. [Google Scholar] [CrossRef] [PubMed]
- Casperson, S.; Sheffield-Moore, M.; Hewlings, S.J.; Paddon-Jones, D. Leucine supplementation chronically improves muscle protein synthesis in older adults consuming the RDA for protein. Clin. Nutr. 2012, 31, 512–519. [Google Scholar] [CrossRef]
- Murphy, C.H.; I Saddler, N.; Devries, M.C.; McGlory, C.; Baker, S.K.; Phillips, S.M. Leucine supplementation enhances integrative myofibrillar protein synthesis in free-living older men consuming lower- and higher-protein diets: A parallel-group crossover study. Am. J. Clin. Nutr. 2016, 104, 1594–1606. [Google Scholar] [CrossRef] [PubMed]
- Paddon-Jones, D.; Sheffield-Moore, M.; Urban, R.J.; Sanford, A.P.; Aarsland, A.; Wolfe, R.R.; Ferrando, A.A. Essential Amino Acid and Carbohydrate Supplementation Ameliorates Muscle Protein Loss in Humans during 28 Days Bedrest. J. Clin. Endocrinol. Metab. 2004, 89, 4351–4358. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, C.; D’Souza, R.F.; Mitchell, S.M.; Figueiredo, V.C.; Miller, B.F.; Hamilton, K.L.; Peelor, F.F.; Coronet, M.; Pileggi, C.A.; Durainayagam, B.; et al. Impact of dairy protein during limb immobilization and recovery on muscle size and protein synthesis; a randomized controlled trial. J. Appl. Physiol. 2018, 124, 717–728. [Google Scholar] [CrossRef] [PubMed]
- Kilroe, S.P.; Fulford, J.; Jackman, S.; Holwerda, A.; Gijsen, A.; van Loon, L.; Wall, B.T. Dietary protein intake does not modulate daily myofibrillar protein synthesis rates or loss of muscle mass and function during short-term immobilization in young men: A randomized controlled trial. Am. J. Clin. Nutr. 2020, 113, 548–561. [Google Scholar] [CrossRef] [PubMed]
- Edwards, S.J.; Smeuninx, B.; McKendry, J.; Nishimura, Y.; Luo, D.; Marshall, R.N.; Perkins, M.; Ramsay, J.; Joanisse, S.; Philp, A.; et al. High-dose leucine supplementation does not prevent muscle atrophy or strength loss over 7 days of immobilization in healthy young males. Am. J. Clin. Nutr. 2020, 112, 1368–1381. [Google Scholar] [CrossRef]
- Gillis, C.; Ljungqvist, O.; Carli, F. Prehabilitation, enhanced recovery after surgery, or both? A narrative review. Br. J. Anaesth. 2022. [Google Scholar] [CrossRef]
- Gillis, C.; Wischmeyer, P.E. Pre-operative nutrition and the elective surgical patient: Why, how and what? Anaesthesia 2019, 74, 27–35. [Google Scholar] [CrossRef]
- Muyskens, J.B.; Foote, D.M.; Bigot, N.J.; Strycker, L.A.; Smolkowski, K.; Kirkpatrick, T.K.; Lantz, B.A.; Shah, S.N.; Mohler, C.G.; Jewett, B.A.; et al. Cellular and morphological changes with EAA supplementation before and after total knee arthroplasty. J. Appl. Physiol. 2019, 127, 531–545. [Google Scholar] [CrossRef] [PubMed]
- Robinson, M.M.; Turner, S.M.; Hellerstein, M.K.; Hamilton, K.L.; Miller, B.F. Long-term synthesis rates of skeletal muscle DNA and protein are higher during aerobic training in older humans than in sedentary young subjects but are not altered by protein supplementation. FASEB J. 2011, 25, 3240–3249. [Google Scholar] [CrossRef]
- Loomba, R.; Decaris, M.; Li, K.W.; Shankaran, M.; Mohammed, H.; Matthews, M.; Richards, L.M.; Nguyen, P.; Rizo, E.; Andrews, B.; et al. Discovery of Half-life of Circulating Hepatitis B Surface Antigen in Patients with Chronic Hepatitis B Infection Using Heavy Water Labeling. Clin. Infect. Dis. 2018, 69, 542–545. [Google Scholar] [CrossRef] [PubMed]
- Damas, F.; Phillips, S.M.; Libardi, C.A.; Vechin, F.C.; Lixandrão, M.E.; Jannig, P.R.; Costa, L.A.R.; Bacurau, A.V.; Snijders, T.; Parise, G.; et al. Resistance training-induced changes in integrated myofibrillar protein synthesis are related to hypertrophy only after attenuation of muscle damage. J. Physiol. 2016, 594, 5209–5222. [Google Scholar] [CrossRef] [PubMed]
- Berman, E.S.F.; Fortson, S.L.; Snaith, S.P.; Gupta, M.; Baer, D.S.; Chery, I.; Blanc, S.; Melanson, E.L.; Thomson, P.J.; Speakman, J.R. Direct Analysis of δ2H and δ18O in Natural and Enriched Human Urine Using Laser-Based, Off-Axis Integrated Cavity Output Spectroscopy. Anal. Chem. 2012, 84, 9768–9773. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Wilkinson, D.J.; Franchi, M.V.; Brook, M.; Narici, M.V.; Williams, J.P.; Mitchell, W.K.; Szewczyk, N.J.; Greenhaff, P.; Atherton, P.J.; Smith, K. A validation of the application of D2O stable isotope tracer techniques for monitoring day-to-day changes in muscle protein subfraction synthesis in humans. Am. J. Physiol. Metab. 2014, 306, E571–E579. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- Oikawa, S.Y.; Bahniwal, R.; Holloway, T.M.; Lim, C.; McLeod, J.C.; McGlory, C.; Baker, S.K.; Phillips, S.M. Potato Protein Isolate Stimulates Muscle Protein Synthesis at Rest and with Resistance Exercise in Young Women. Nutrients 2020, 12, 1235. [Google Scholar] [CrossRef]
- Howard, E.E.; Pasiakos, S.M.; Blesso, C.N.; Fussell, M.A.; Rodriguez, N.R. Divergent Roles of Inflammation in Skeletal Muscle Recovery from Injury. Front. Physiol. 2020, 11, 87. [Google Scholar] [CrossRef]
- Zeng, N.; D’Souza, R.F.; MacRae, C.L.; Figueiredo, V.C.; Pileggi, C.A.; Markworth, J.F.; Merry, T.L.; Cameron-Smith, D.; Mitchell, C.J. Daily protein supplementation attenuates immobilization-induced blunting of postabsorptive muscle mTORC1 activation in middle-aged men. Am. J. Physiol. Physiol. 2021, 320, C591–C601. [Google Scholar] [CrossRef]
- Fry, C.S.; Johnson, D.L.; Ireland, M.L.; Noehren, B. ACL injury reduces satellite cell abundance and promotes fibrogenic cell expansion within skeletal muscle. J. Orthop. Res. 2016, 35, 1876–1885. [Google Scholar] [CrossRef] [PubMed]
- Møller, A.B.; Vendelbo, M.H.; Schjerling, P.; Couppé, C.; Moller, N.; Kjær, M.; Hansen, M.; Jessen, N. Immobilization Decreases FOXO3a Phosphorylation and Increases Autophagy-Related Gene and Protein Expression in Human Skeletal Muscle. Front. Physiol. 2019, 10, 736. [Google Scholar] [CrossRef] [PubMed]
- Greenhaff, P.L.; Karagounis, L.; Peirce, N.; Simpson, E.J.; Hazell, M.; Layfield, R.; Wackerhage, H.; Smith, K.; Atherton, P.; Selby, A.; et al. Disassociation between the effects of amino acids and insulin on signaling, ubiquitin ligases, and protein turnover in human muscle. Am. J. Physiol. Endocrinol. Metab. 2008, 295, E595–E604. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| AP | OP | p Value | |
|---|---|---|---|
| Age (y) | 32 ± 11 | 30 ± 7 | 0.78 |
| Height (m) | 1.8 ± 0.1 | 1.8 ± 0.1 | 0.75 |
| Weight (kg) | 85.3 ± 8.2 | 76.2 ± 14.5 | 0.45 |
| Body mass index | 26.2 ± 1.6 | 23.9 ± 2.2 | 0.25 |
| AP | OP | p Value | |
|---|---|---|---|
| Energy (kcal/d) | 2367 ± 125 | 2853 ± 382 | 0.10 |
| Carbohydrate (g·kg−1·d−1) | 3.7 ± 0.6 | 4.3 ± 1.8 | 0.60 |
| Fat (g·kg−1·d−1) | 1.0 ± 0.l | 1.4 ± 0.3 | 0.06 |
| Protein (g·kg−1·d−1) | 1.2 ± 0.0 | 1.9 ± 0.2 | 0.01 |
| Leucine (mg·kg−1·d−1) | 73 ± 10 | 104 ± 14 | 0.04 |
| Isoleucine (mg·kg−1·d−1) | 42 ± 7 | 58 ± 8 | 0.06 |
| Valine (mg·kg−1·d−1) | 48 ± 7 | 68 ± 9 | 0.04 |
| Total BCAAs (mg·kg−1·d−1) | 162 ± 24 | 230 ± 32 | 0.04 |
| AP | OP | p Value | Effect Size | |
|---|---|---|---|---|
| Myogenesis | ||||
| MyoD | 1.00 ± 0.11 | 1.53 ± 0.94 | 0.39 | 0.63 |
| Myogenin | 1.03 ± 0.31 | 1.12 ± 0.23 | 0.72 | 0.25 |
| Pax7 | 1.20 ± 0.92 | 0.79 ± 0.16 | 0.48 | 0.51 |
| Myf5 | 1.17 ± 0.85 | 0.89 ± 0.14 | 0.60 | 0.35 |
| Myf6 | 1.10 ± 0.52 | 0.96 ± 0.43 | 0.74 | 0.24 |
| Proteolysis | ||||
| MAFbx | 1.10 ± 0.50 | 0.94 ± 0.24 | 0.66 | 0.31 |
| MuRF1 | 1.04 ± 0.34 | 0.66 ± 0.23 | 0.18 | 1.05 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Howard, E.E.; Margolis, L.M.; Fussell, M.A.; Rios, C.G.; Meisterling, E.M.; Lena, C.J.; Pasiakos, S.M.; Rodriguez, N.R. Effect of High-Protein Diets on Integrated Myofibrillar Protein Synthesis before Anterior Cruciate Ligament Reconstruction: A Randomized Controlled Pilot Study. Nutrients 2022, 14, 563. https://doi.org/10.3390/nu14030563
Howard EE, Margolis LM, Fussell MA, Rios CG, Meisterling EM, Lena CJ, Pasiakos SM, Rodriguez NR. Effect of High-Protein Diets on Integrated Myofibrillar Protein Synthesis before Anterior Cruciate Ligament Reconstruction: A Randomized Controlled Pilot Study. Nutrients. 2022; 14(3):563. https://doi.org/10.3390/nu14030563
Chicago/Turabian StyleHoward, Emily E., Lee M. Margolis, Maya A. Fussell, Clifford G. Rios, Eric M. Meisterling, Christopher J. Lena, Stefan M. Pasiakos, and Nancy R. Rodriguez. 2022. "Effect of High-Protein Diets on Integrated Myofibrillar Protein Synthesis before Anterior Cruciate Ligament Reconstruction: A Randomized Controlled Pilot Study" Nutrients 14, no. 3: 563. https://doi.org/10.3390/nu14030563
APA StyleHoward, E. E., Margolis, L. M., Fussell, M. A., Rios, C. G., Meisterling, E. M., Lena, C. J., Pasiakos, S. M., & Rodriguez, N. R. (2022). Effect of High-Protein Diets on Integrated Myofibrillar Protein Synthesis before Anterior Cruciate Ligament Reconstruction: A Randomized Controlled Pilot Study. Nutrients, 14(3), 563. https://doi.org/10.3390/nu14030563