
The Effect of the Ketogenic Diet on Adiponectin, Omentin and Vaspin in Children with Drug-Resistant Epilepsy

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
Statistical Analysis
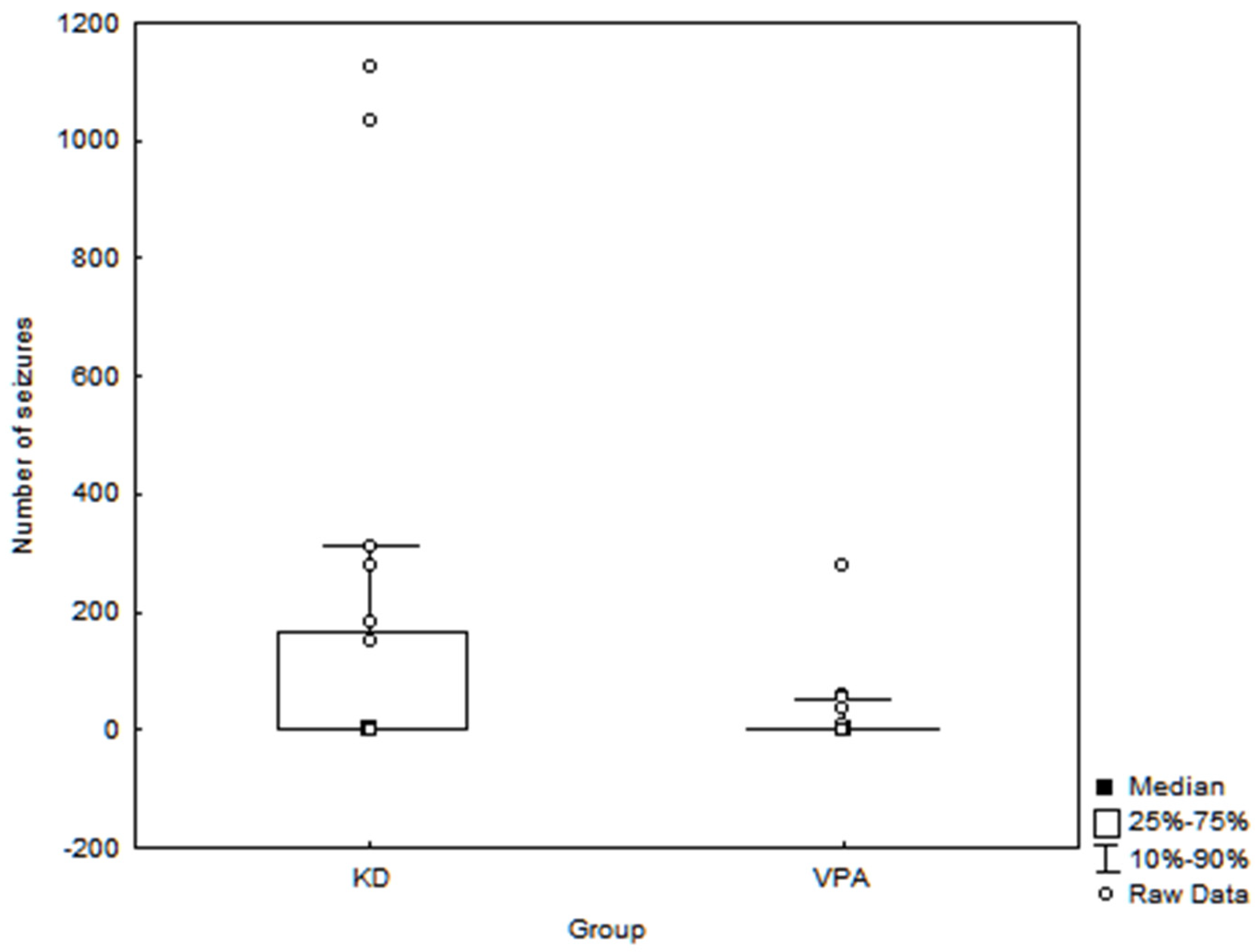
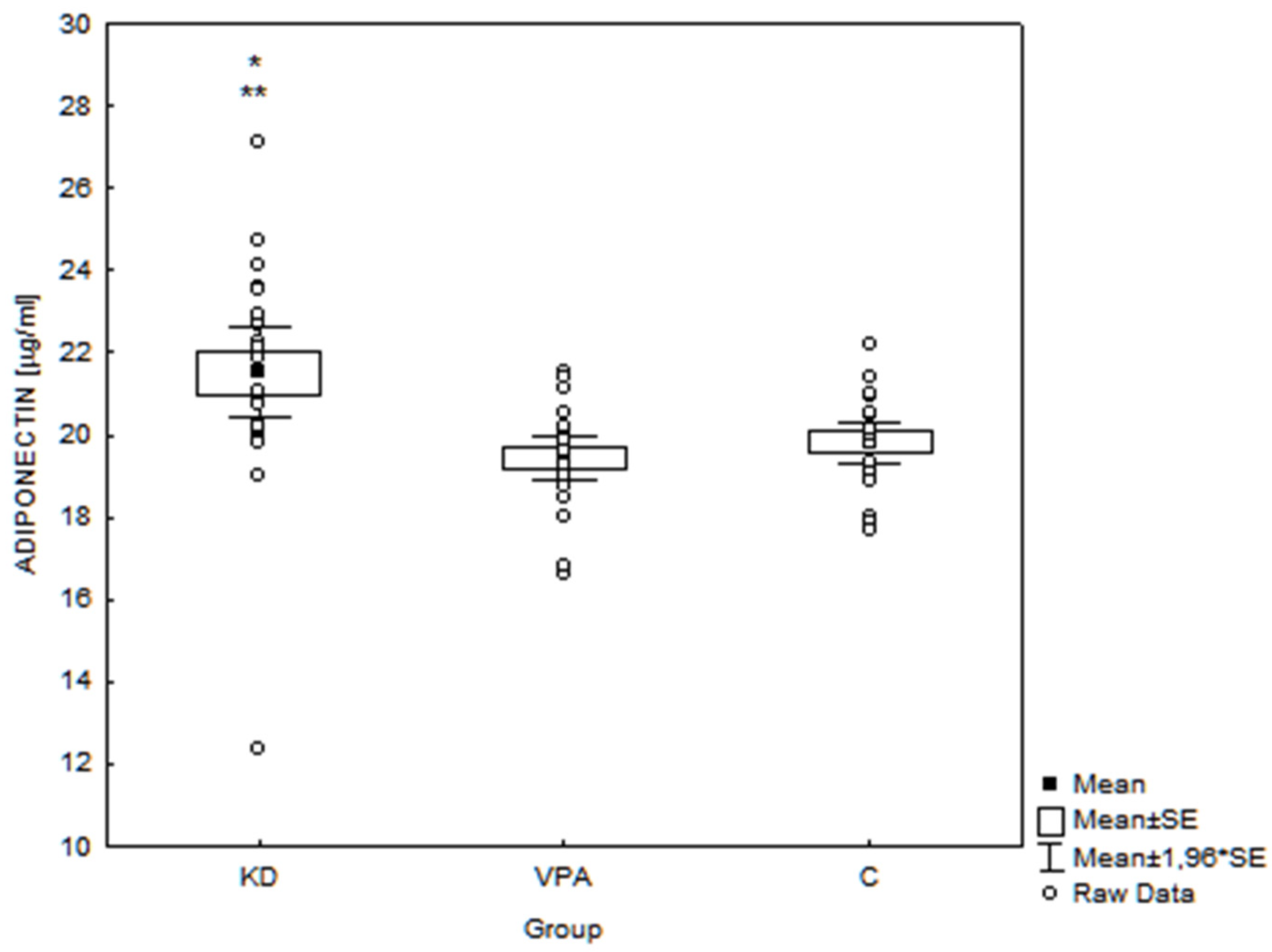
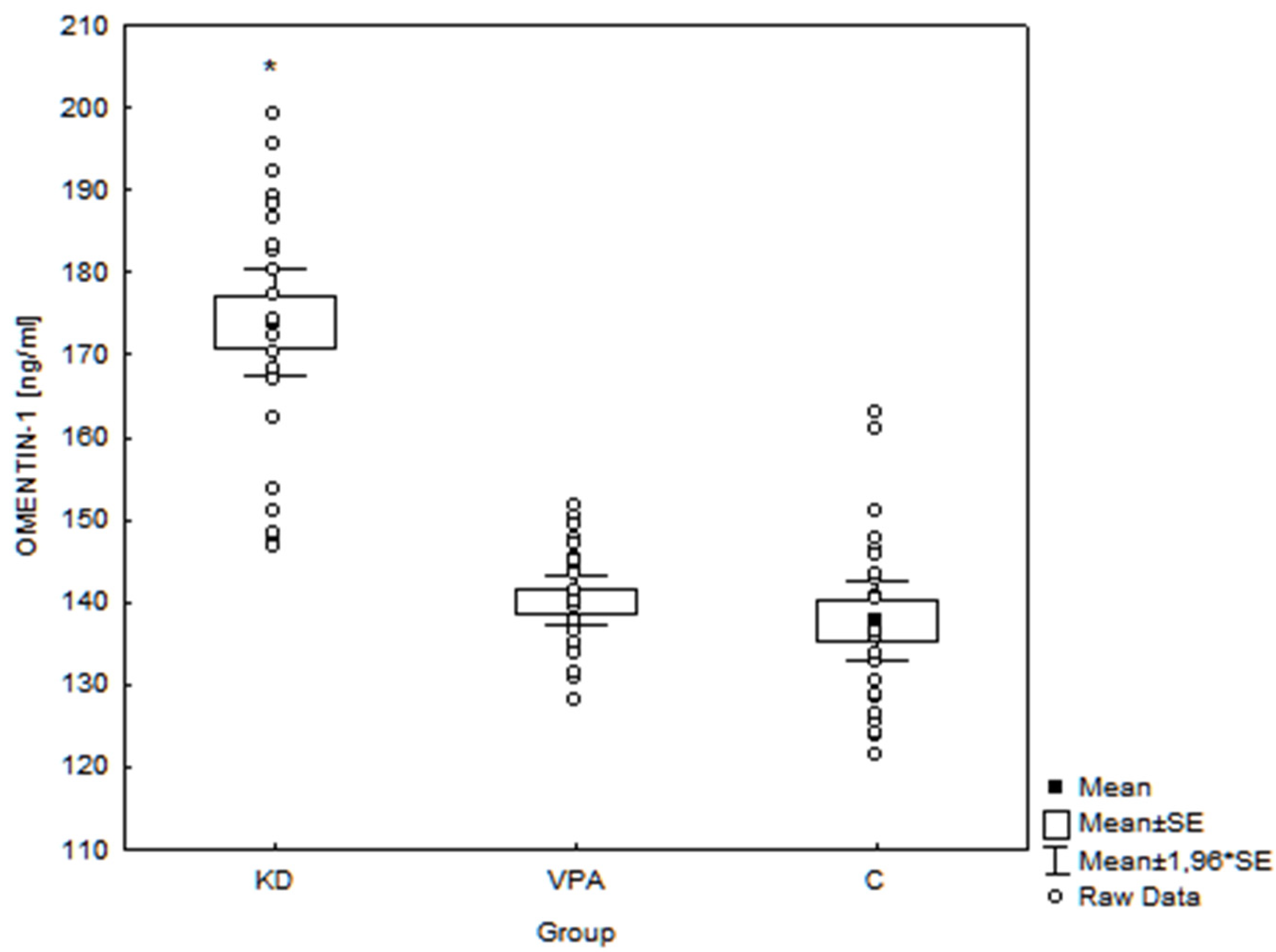
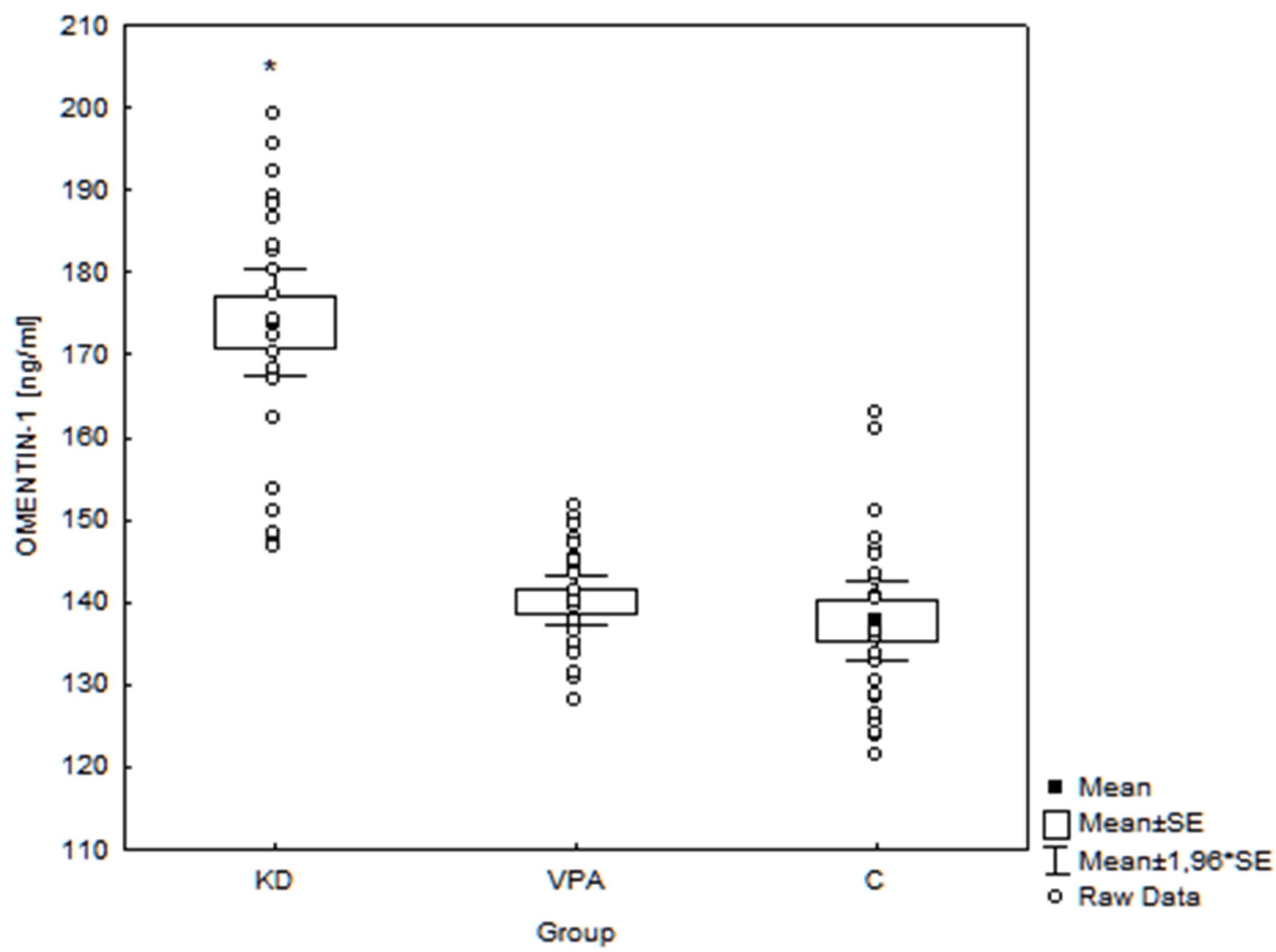
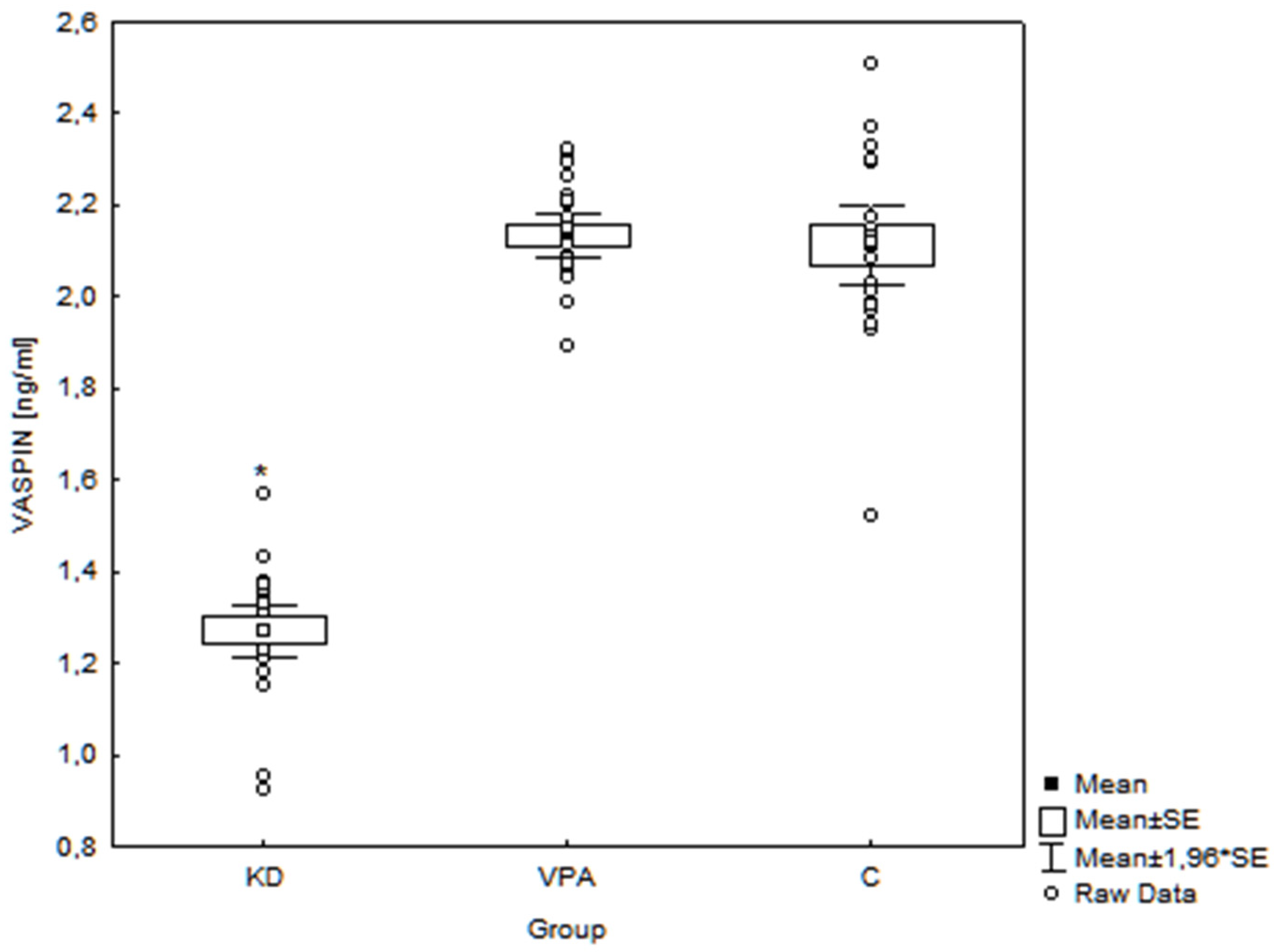
3. Results
4. Discussion
4.1. Ketogenic Diet and Adipokines
4.1.1. Adiponectin
4.1.2. Omentin-1
4.1.3. Vaspin
4.2. Study Limitations and Advantages
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Ethics Approval
Consent to Participation
References
- Ułamek-Kozioł, M.; Czuczwar, S.J.; Januszewski, S.; Pluta, R. Ketogenic diet and epilepsy. Nutrients 2019, 11, 2510. [Google Scholar] [CrossRef] [Green Version]
- Kossoff, E.H.; Zupec-Kania, B.A.; Auvin, S.; Ballaban-Gil, K.R.; Bergqvist, A.G.C.; Blackford, R.; Buchhalter, J.R.; Caraballo, R.H.; Cross, J.H.; Dahlin, M.G.; et al. Optimal Clinical management of children receiving dietary therapies for epilepsy: Updated recommendations of International Ketogenic Diet Study Group. Epilepsia Open 2018, 3, 175–192. [Google Scholar] [CrossRef]
- van der Louw, E.J.T.M.; van den Hurk, D.; Neal, E.; Leiendecker, B.; Fitzsimmon, G.; Dority, L.; Thompson, L.; Marchiò, M.; Dudzińska, M.; Dressler, A.; et al. Ketogenic diet guidelines for infants with refractory epilepsy. Eur. J. Paediatr. Neurol. 2016, 20, 798–809. [Google Scholar] [CrossRef] [PubMed]
- Li, R.-J.; Liu, Y.; Liu, H.-Q.; Li, J. Ketogenic diets and protective mechanisms in epilepsy, metabolic disorders, cancer, neuronal loss, and muscle and nerve degeneration. J. Food Biochem. 2020, 44, e13140. [Google Scholar] [CrossRef] [PubMed]
- Paoli, A.; Bosco, G.; Camporesi, E.M.; Mangar, D. Ketosis, ketogenic diet and food intake control: A complex relationship. Front. Psychol. 2015, 6, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Amicis, R.; Leone, A.; Lessa, C.; Foppiani, A.; Ravella, S.; Ravasenghi, S.; Trentani, C.; Ferraris, C.; Veggiotti, P.; De Giorgis, V.; et al. Long-Term Effects of a Classic Ketogenic Diet on Ghrelin and Leptin Concentration: A 12-Month Prospective Study in a Cohort of Italian Children and Adults with GLUT1-Deficiency Syndrome and Drug Resistant Epilepsy. Nutrients 2019, 11, 1716. [Google Scholar] [CrossRef] [Green Version]
- Trayhurn, P.; Wood, I.S. Adipokines: Inflammation and the pleiotropic role of white adipose tissue. Br. J. Nutr. 2004, 92, 347–355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paudel, Y.N.; Shaikh, M.F.; Shah, S.; Kumari, Y.; Othman, I. Role of inflammation in epilepsy and neurobehavioral comorbidities: Implication for therapy. Eur. J. Pharmacol. 2018, 837, 145–155. [Google Scholar] [CrossRef]
- Krasnodębski, P.; Dęmbe, K.; Kmiecik, M.; Wójcik-Sosonowska, E.; Jasik, M.; Mrozikiewicz-Rakowska, B.; Karnafel, W. Fizjologiczne mechanizmy działania adiponektyny. Med. Metab. 2014, 18, 65–69. [Google Scholar]
- Zhou, J.Y.; Chan, L.; Zhou, S.W. Omentin: Linking metabolic syndrome and cardiovascular disease. Curr. Vasc. Pharmacol. 2014, 12, 136–143. [Google Scholar] [CrossRef]
- Dimova, R.; Tankova, T. The role of vaspin in the development of metabolic and glucose tolerance disorders and atherosclerosis. Biomed. Res. Int. 2015, 2015, 823481. [Google Scholar] [CrossRef]
- Lee, E.B.; Warmann, G.; Dhir, R.; Ahima, R.S. Metabolic dysfunction associated with adiponectin deficiency enhances kainic acid-induced seizure severity. J. Neurosci. 2011, 31, 14361–14366. [Google Scholar] [CrossRef] [Green Version]
- Jeon, B.T.; Shin, H.J.; Kim, J.B.; Kim, Y.K.; Lee, D.H.; Kim, K.H.; Kim, H.J.; Kang, S.S.; Cho, G.J.; Choi, W.S.; et al. Adiponectin protects hippocampal neurons against kainic acid-induced excitotoxicity. Brain. Res. Rev. 2009, 61, 81–88. [Google Scholar] [CrossRef]
- Ziora, K.; Oświęcimska, J.; Świętochowska, E.; Stojewska, M.; Ostrowska, Z.; Suwała, A. Ocena stężeń adiponektyny—Hormonu tkanki tłuszczowej w surowicy krwi u dziewcząt z jadłowstrętem psychicznym. Endokrynol. Pediat. 2010, 9, 31–42. [Google Scholar]
- Robinson, K.; Prins, J.; Venkatesh, B. Clinical review: Adiponectin biology and its role in inflammation and critical illness. Crit. Care 2011, 15, 221. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.V.; Scherer, P.E. Adiponectin, the past two decades. J. Mol. Cell Biol. 2016, 8, 93–100. [Google Scholar] [CrossRef]
- Tilg, H.; Moschen, A. Adipocytokines: Mediators linking adipose tissue, inflammation and immunity. Nature 2006, 6, 772–783. [Google Scholar] [CrossRef]
- Wang, Y.; Lam, K.S.; Yau, M.H.; Xu, A. Post-translational modifications of adiponectin: Mechanisms and functional implications. J. Biochem. 2008, 409, 623–633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C. Obesity, inflammation, and lung injury (OILI): The good. Mediators Inflamm. 2014, 2014, 978463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, R.Z.; Lee, M.J.; Hu, H.; Pray, J.; Wu, H.B.; Hansen, B.C.; Shuldiner, A.R.; Fried, S.K.; McLenithan, J.C.; Gong, D.W. Identification of omentin as a o novel depot-specific adipokine in human adipose tissue: Possible role in modulating insulin action. Am. J. Physiol. Endocrinol. Metab. 2006, 290, E1253–E1261. [Google Scholar] [CrossRef]
- Hida, K.; Wada, J.; Eguchi, J.; Zhang, H.; Baba, M.; Seida, A.; Hashimoto, I.; Okada, T.; Yasuhara, A.; Nakatsuka, A.; et al. Visceral adipose tissue-derived serine protease inhibitor: A unique insulin-sensitizing adipocytokine in obesity. Proc. Natl. Acad. Sci. USA 2005, 102, 10610–10615. [Google Scholar] [CrossRef] [Green Version]
- Achari, A.E.; Jain, S.K. Adiponectin, a therapeutic target for obesity, diabetes, and endothelial dysfunction. Int. J. Mol. Sci. 2017, 18, 1321. [Google Scholar] [CrossRef] [Green Version]
- Fonseca-Alaniz, M.H.; Takada, J.; Alonso-Vale, M.I.C.; Lima, F.B. Adipose tissue as an endocrine organ: From theory to practice. J. Pediatr. 2007, 83, S192–S203. [Google Scholar] [CrossRef]
- Meier, U.; Gressner, A.M. Endocrine Regulation of Energy Metabolism: Review of pathobiochemical and clinical chemical aspects of leptin, ghrelin, adiponectin, and resistin. Clin. Chem. 2004, 50, 1511–1525. [Google Scholar] [CrossRef]
- Lesna, J.; Ticha, A.; Hyspler, R.; Sobotka, L.; Zadak, Z.; Smahelova, A. Omentin-1 plasma levels and cholesterol metabolism in obese patients with diabetes mellitus type 1: Impact of weight reduction. Nutr. Diabetes 2015, 5, e183. [Google Scholar] [CrossRef] [Green Version]
- Jung, C.H.; Lee, M.J.; Kang, Y.M.; La Lee, Y.; Yoon, H.K.; Kang, S.-W.; Lee, W.J.; Park, J.-Y. Vaspin inhibits cytokine-induced nuclear factor-kappa B activation and adhesion molecule expression via AMP-activated protein kinase activation in vascular endothelial cells. Cardiovasc. Diabetol. 2014, 13, 41. [Google Scholar] [CrossRef] [Green Version]
- Hardie, D.G. AMPK: Positive and negative regulation, and its role in whole-body energy homeostasis. Curr. Opin. Cell Biol. 2015, 33, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Salminen, A.; Hyttinen, J.M.T.; Kaarniranta, K. AMP-activated protein kinase inhibits NF-κB signaling and inflammation: Impact on healthspan and lifespan. J. Mol. Med. 2011, 89, 667–676. [Google Scholar] [CrossRef] [Green Version]
- Sherrier, M.; Li, H. The impact of keto-adaptation on exercise performance and the role of metabolic-regulating cytokines. Am. J. Clin. Nutr. 2019, 110, 562–573. [Google Scholar] [CrossRef] [PubMed]
- Kadowaki, T.; Yamauchi, T. Adiponectin and adiponectin receptors. Endocr. Rev. 2005, 26, 439–451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okamoto, M.; Ohara-Imaizumi, M.; Kubota, N.; Hashimoto, S.; Eto, K.; Kanno, T.; Kubota, T.; Wakui, M.; Nagai, R.; Noda, M.; et al. Adiponectin induces insulin secretion in vitro and in vivo at low glucose concentration. Diabetologia 2008, 51, 527–535. [Google Scholar] [CrossRef] [Green Version]
- Kurowska, P.; Mlyczyńska, E.; Dawid, M.; Grzesiak, M.; Dupont, J.; Rak, A. The role of vaspin in porcine corpus luteum. J. Endocrinol. 2020, 247, 283–294. [Google Scholar] [CrossRef] [PubMed]
- Jeong, E.; Youn, B.-S.; Kim, D.W.; Kim, E.H.; Park, J.W.; Namkoong, C.; Jeong, J.Y.; Yoon, S.Y.; Park, J.Y.; Lee, K.-U.; et al. Circadian rhythm of serum vaspin in healthy male volunteers: Relation to meals. J. Clin. Endocrinol. Metab. 2010, 95, 1869–1875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fantuzzi, G. Adiponectin in inflammatory and immune-mediated diseases. Cytokine 2013, 64, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, H.Y.; Guo, L.; Li, Q. Changes of serum omentin-1 levels in normal subjects and in patients with impaired glucose regulation and with newly diagnosed and untreated type 2 diabetes. Diabetes Res. Clin. Pract. 2010, 88, 29–33. [Google Scholar] [CrossRef]
- Ulbricht, D.; Tindall, C.A.; Oertwig, K.; Hanke, S.; Sträter, N.; Heiker, J.T. Kallikrein-related peptidase 14 is the second KLK protease targeted by the serpin vaspin. Biol. Chem. 2018, 399, 1079–1084. [Google Scholar] [CrossRef]
- Kurowska, P.; Mlyczyńska, E.W.; Dawid, M.; Jurek, M.; Klimczyk, D.; Dupont, J.; Rak, A. Review: Vaspin (SERPINA12) expression and function in endocrine cells. Cells 2021, 10, 1710. [Google Scholar] [CrossRef]
- Tan, B.K.; Adya, R.; Farhatullah, S.; Lewandowski, K.C.; O.-Hare, P.; Lehnert, H.; Randeva, H.S. Omentin-1, a novel adipokine, is decreased in overweight insulin-resistant women with polycystic ovary syndrome: Ex vivo and in vivo regulation of omentin-1 by insulin and glucose. Diabetes 2008, 57, 801–808. [Google Scholar] [CrossRef] [Green Version]
- Tan, B.K.; Adya, R.; Randeva, H.S. Omentin: A novel link between inflammation, obesity and cardiovascular disease. Trends Cardiovasc. Med. 2010, 20, 143–148. [Google Scholar] [CrossRef]
- Yamawaki, H.; Kuramoto, J.; Kameshima, S.; Usui, T.; Okada, M.; Hara, Y. Omentin, a novel adipocytokine inhibits TNF-induced vascular inflammation in human endothelial cells. Biochem. Biophys. Res. Commun. 2011, 408, 339–343. [Google Scholar] [CrossRef]
- Rao, S.S.; Hu, Y.; Xie, P.L.; Cao, J.; Wang, Z.-X.; Liu, J.-H.; Yin, H.; Huang, J.; Tan, Y.J.; Luo, J.; et al. Omentin-1 prevents inflammation-induced osteoporosis by downregulating the pro-inflammatory cytokines. Bone Res. 2018, 6, 9. [Google Scholar] [CrossRef] [Green Version]
- Brunetti, L.; Leone, S.; Orlando, G.; Ferrante, C.; Recinella, L.; Chiavaroli, A.; Di Nisio, C.; Shohreh, R.; Manippa, F.; Ricciuti, A.; et al. Hypotensive effects of omentin-1 related to increased adiponectin and decreased interleukin-6 in intra-thoracic pericardial adipose tissue. Pharmacol. Rep. 2014, 66, 991–995. [Google Scholar] [CrossRef]
- Simeone, T.A.; Simeone, K.A.; Rho, J.M. Ketone bodies as anti-seizure agents. Neurochem. Res. 2017, 42, 2011–2018. [Google Scholar] [CrossRef]
- Maalouf, M.A.; Rho, J.M.; Mattson, M.P. The neuroprotective properties of calorie restriction, the ketogenic diet, and ketogenic bodies. Brain Res. Rev. 2009, 59, 293–315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giordano, C.; Marchio, M.; Timofeeva, E.; Biagini, G. Neuroactive peptides as putative mediators of antiepileptic ketogenic diets. Front. Neurol. 2014, 5, 63. [Google Scholar] [CrossRef] [PubMed]
- Stafstrom, C.; Rho, J.M. The ketogenic diet as a treatment paradigm for diverse neurological disorders. Front. Pharmacol. 2012, 3, 59. [Google Scholar] [CrossRef] [Green Version]
- Gasior, M.; Rogawski, M.A.; Hartman, A.L. Neuroprotective and disease-modifying effects of ketogenic diet. Behav. Pharmacol. 2006, 17, 431–439. [Google Scholar] [CrossRef] [Green Version]
- Vezzani, A. Innate immunity and inflammation in temporal lobe epilepsy: New emphasis on the role of complement activation. Epilepsy Curr. 2008, 8, 75–77. [Google Scholar] [CrossRef] [Green Version]
- Vezzani, A.; Granata, T. Brain inflammation in epilepsy: Experimental and clinical evidence. Epilepsia 2005, 46, 1724–1743. [Google Scholar] [CrossRef]
- Vezzani, A.; French, J.; Bartfai, T.; Baram, T.Z. The role of inflammation in epilepsy. Nat. Rev. Neurol. 2011, 7, 31–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Damani, M.R.; Zhao, L.; Fontainhas, A.M.; Amaral, J.; Fariss, R.N.; Wong, W.T. Age-related alterations in the dynamic behavior of microglia. Aging Cell 2011, 10, 263–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vezzani, A.; Friedman, A. Brain inflammation as a biomarker in epilepsy. Biomark. Med. 2011, 5, 607–614. [Google Scholar] [CrossRef] [Green Version]
- Aronica, E.; Crino, P.B. Inflammation in epilepsy: Clinical observations. Epilepsia 2011, 52, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Riazi, K.; Galic, M.A.; Pittman, Q.J. Contributions of peripheral inflammation to seizure susceptibility: Cytokines and brain excitability. Epilepsy Res. 2010, 89, 34–42. [Google Scholar] [CrossRef]
- Nielsen, K.A.; Hansen, E.L.; Gille, S. Genotyping of the cytochrome P450 2D6 4469 C>T polymorphism Rusing SimpleProbes. Scand. J. Clin. Lab. Investig. 2007, 67, 280–290. [Google Scholar] [CrossRef] [PubMed]
- Yu, N.; Di, Q.; Liu, H.; Hu, Y.; Jiang, Y.; Yan, Y.; Zhang, Y.-F.; Zhang, Y.-D. Nuclear factor-kappa B activity regulates brain expression of P-glycoprotein in the kainic acid-induced seizure rats. Mediat. Inflamm. 2011, 2011, 670613. [Google Scholar] [CrossRef]
- Kapoor, D.; Sharma, S.; Garg, D. Emerging Role of the Ketogenic Dietary Therapies beyond Epilepsy in Child Neurology. Ann. Indian Acad. Neurol. 2021, 24, 470–480. [Google Scholar]
- Ruskin, D.N.; Sacchettic, P.; Masinob, S.A. A unifying mechanism of ketogenic diet action: The multiple roles of nicotinamide adenine dinucleotide. Epilepsy Res. 2020, 167, 106469. [Google Scholar]
- Franczyk, M.P.; Qi, N.; Stromsdorfer, K.L.; Li, C.; Yamaguchi, S.; Itoh, H.; Yoshino, M.; Sasaki, Y.; Brookheart, R.T.; Finck, B.N.; et al. Importance of Adipose Tissue NAD+ Biology in Regulating Metabolic Flexibility. Endocrinology 2021, 162, bqab006. [Google Scholar] [CrossRef]
- Dracheva, K.V.; Pobozheva, I.A.; Panteleeva, A.A.; Baranova, E.I.; Pchelina, S.N.; Miroshnikova, V.V. Omentin-1 gene expression is gender specific influenced by PPARγ. Atherosclerosis 2021, 331, E190–E191. [Google Scholar] [CrossRef]
- El-Mesallamy, H.O.; El-Derany, M.O.; Hamdy, N.M. Serum omentin-1 and chemerin levels are interrelated in patients with Type 2 diabetes mellitus with or without ischaemic heart disease. Diabet. Med. 2011, 28, 1194–1200. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Yang, G.; Shi, S.; Yang, M.; Liu, H.; Boden, G. The adipose triglyceride lipase, adiponectin and visfatin are downregulated by tumor necrosis factor-α (TNF-α) in vivo. Cytokine 2009, 45, 12–19. [Google Scholar] [CrossRef]
- Boison, D. New insights into the mechanisms of the ketogenic diet. Curr. Opin. Neurol. 2017, 30, 187–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dowis, K.; Banga, S. The Potential Health Benefits of the Ketogenic Diet: A Narrative Review. Nutrients 2021, 13, 1654. [Google Scholar] [CrossRef] [PubMed]
- Yao, H.; Fan, C.; Lu, Y.; Fan, X.; Xia, L.; Li, P.; Wang, R.; Tang, T.; Wang, Y.; Qi, K. Alteration of gut microbiota affects expression of adiponectin and resistin through modifying DNA methylation in high-fat diet-induced obese mice. Genes Nutr. 2020, 15, 12. [Google Scholar] [CrossRef] [PubMed]
- Poff, A.M.; Rho, J.M.; D’Agostino, D.P. Ketone Administration for Seizure Disorders: History and Rationale for Ketone Esters and Metabolic Alternatives. Front. Neurosci. 2019, 13, 1041. [Google Scholar] [CrossRef] [Green Version]
- Díez, J.J.; Iglesias, P. The role of the morel adipocyte derived hormone adiponectin in human disease. Eur. J. Endocrinol. 2003, 148, 293–300. [Google Scholar] [CrossRef] [Green Version]
- Chen, B.; Liao, W.Q.; Xu, N.; Xu, H.; Wen, J.Y.; Yu, C.A.; Liu, X.-Y.; Li, C.-L.; Shao, S.-M.; Campbell, W. Adiponectin protects against cerebral ischemia-reperfusion injury through anti-inflammatory action. Brain Res. 2009, 1273, 129–137. [Google Scholar] [CrossRef]
- Tang, H.; Sebastian, B.M.; Axhemi, A.; Chen, X.; Hillian, A.D.; Jacobsen, D.W.; Nagy, L.E. Ethanol-induced oxidative stress via the CYP2E1 pathway disrupts adiponectin secretion from adipocytes. Alcohol. Clin. Exp. Res. 2012, 36, 214–222. [Google Scholar] [CrossRef] [Green Version]
- Ethemoglu, O.; Aya, H.; Koyuncub, I.; Gönelb, A. Comparison of cytokines and prooxidants/antioxidants markers among adults with refractory versus well-controlled epilepsy: A cross-sectional study. Seizure 2018, 60, 105–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toscano, E.; Lessa, J.; Goncavales, A.; Rocha, N.P.; Giannetti, A.V.; de Oliveira, G.N.; Rachid, M.A.; Vieira, E.A.M.; Teixeria, A.L. Circulating levels of adipokines are altered in patients with temporal lobe epilepsy. Epilepsy Behav. 2019, 90, 137–141. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.-R.; Jin, M.-F.; Tang, L.; Liu, Y.-Y.; Ni, H. Acute phase serum leptin, adiponectin, interleukin-6, and visfatin are altered in Chinese children with febrile seizures: A cross-sectional study. Front. Endocrinol. 2020, 11, 531. [Google Scholar] [CrossRef]
- Palmio, J.; Vuolteenaho, K.; Lehtimäki, K.; Nieminen, R.; Peltola, J.; Moilanen, E. CSF and plasma adipokines after tonic–clonic seizures. Seizure 2016, 39, 10–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rauchenzauner, M.; Klepper, J.; Leiendecker, B.; Luef, G.; Rostasy, K.; Ebenbichler, C. The Ketogenic Diet in Children with Glut1 Deficiency Syndrome and Epilepsy. J. Pediatr. 2008, 153, 716–718. [Google Scholar] [CrossRef]
- Bertoli, S.; Giulini Neri, I.; Trentani, C.; Ferraris, C.; De Amicis, R.; Battezzati, A.; Veggiotti, P.; De Giorgis, V.; Tagliabue, A. Short-term effects of ketogenic diet on anthropometric parameters, body fat distribution, and inflammatory cytokine production in GLUT1 deficiency syndrome. Nutrition 2015, 31, 981–987. [Google Scholar] [CrossRef]
- Partsalaki, I.; Karvela, A.; Spiliotis, B.E. Metabolic impact of a ketogenic diet compared to ahypocaloric diet in obese children and adolescents. J. Pediatr. Endocr. Met. 2012, 25, 697–704. [Google Scholar]
- Rosenbaum, M.; Hall, K.D.; Guo, J.; Ravussin, E.; Mayer, L.S.; Reitman, M.L.; Smith, S.R.; Walsh, B.T.; Leibel, R.L. Glucose and lipid homeostasis and inflammation in humans following an isocaloric ketogenic diet. Obesity 2019, 27, 971–981. [Google Scholar] [CrossRef] [PubMed]
- Cipryan, L.; Dostal, T.; Plews, D.J.; Hofmann, P.; Laursen, P.B. Adiponectin/leptin ratio increases after a 12-week very low-carbohydrate, high-fat diet, and exercise training in healthy individuals: A non-randomized, parallel design study. Nutr. Res. 2021, 87, 22–30. [Google Scholar] [CrossRef]
- Eyzaguirre, F.; Mericq, V. Insulin resistance markers in children. Horm. Res. 2009, 71, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Greco, R.; Latini, G.; Chiarelli, F.; Iannetti, P.; Verrotti, A. Leptin, ghrelin, and adiponectin in epileptic patients treated with valproic acid. Neurology 2005, 65, 1808–1809. [Google Scholar] [CrossRef]
- Rauchenzauner, M.; Haberlandt, E.; Scholl-Bürgi, S.; Barbara, E.; Hoppichler, F.; Karall, D.; Ebenbichler, C.F.; Rostasy, K.; Luef, G. Adiponectin and visfatin concentrations in children treated with valproic acid. Epilepsia 2008, 49, 353–357. [Google Scholar] [CrossRef]
- Meral, C.; Cekmez, F.; Vurucu, S.; Tascılar, E.; Pirgon, O.; Canpolat, F.E.; Ipcioglu, M.M.; Aydemir, G.; Aydınoz, S. New adipocytokines (vaspin, apelin, visfatin, adiponectin) levels in children treated with valproic acid. Eur. Cytokine Netw. 2011, 22, 118–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aly, R.H.; Amr, N.H.; Saad, W.E.; Megahed, A.A. Insulin resistance in patients on valproic acid: Relation to adiponectin. Acta Neurol. Scand. 2015, 131, 169–175. [Google Scholar] [CrossRef] [PubMed]
- Sidhua, H.S.; Srinivasb, R.; Sadhotraa, A. Evaluate the effects of long-term valproic acid treatment on metabolic profiles in newly diagnosed or untreated female epileptic patients: A prospective study. Seizure 2017, 48, 15–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Şimşek, F.; Ceylan, M.; Kızıltunç, A.; İyigün, I. The correlation between carbamazepine and valproic acid monotherapy with serum adiponectin and carnitine. Acta Neurol. Belg. 2021, 121, 1823–1830. [Google Scholar] [CrossRef] [PubMed]
- Qiao, L.; Schaack, J.; Shao, J. Suppression of adiponectin gene expression by histone deacetylase inhibitor valproic acid. Endocrinology 2006, 147, 865–874. [Google Scholar] [CrossRef] [PubMed]
- Nishaa, Y.; Bobbya, Z.; Wadwekarb, V. Biochemical derangements related to metabolic syndrome in epileptic patients on treatment with valproic acid. Seizure Eur. J. Epilepsy 2018, 60, 57–60. [Google Scholar] [CrossRef] [Green Version]
- Sonmez, F.M.; Zaman, D.; Aksoy, A.; Deger, O.; Aliyazicioglu, R.; Karaguzel, G.; Fazlioglu, K. The effects of topiramate and valproate therapy on insulin, c-peptide, leptin, neuropeptide Y, adiponectin, visfatin, and resistin levels in children with epilepsy. Seizure 2013, 22, 856–861. [Google Scholar] [CrossRef] [Green Version]
- de Souza Batista, C.M.; Yang, R.Z.; Lee, M.J.; Glynn, N.M.; Yu, D.Z.; Pray, J.; Ndubuizu, K.; Patil, S.; Schwartz, A.; Kligman, M.; et al. Omentin plasma levels and gene expression are decreased in obesity. Diabetes 2007, 56, 1655–1661. [Google Scholar] [CrossRef] [Green Version]
- Oświęcimska, J.; Suwała, A.; Świętochowska, E.; Ostrowska, Z.; Gorczyca, P.; Ziora-Jakutowicz, K.; Machura, E.; Szczepańska, M.; Kukla, M.; Stojewska, M.; et al. Serum omentin levels in adolescent girls with anorexia nervosa and obesity. Physiol. Res. 2015, 64, 701–709. [Google Scholar] [CrossRef]
- Yang, J.; Gao, Y. Clinical relevance of serum omentin-1 levels as a biomarker of prognosis in patients with acute cerebral infarction inhibit inflammation, thereby reducing the formation and rupture of unstable plaques and prevent cerebrovascular events. Brain Behav. 2020, 10, e01678. [Google Scholar] [CrossRef]
- Youn, B.S.; Klöting, N.; Kratzsch, J.; Lee, N.; Park, J.W.; Song, E.S.; Ruschke, K.; Oberbach, A.; Fasshauer, M.; Stumvoll, M.; et al. Serum vaspin concentrations in human obesity and type 2 diabetes. Diabetes 2008, 57, 372–377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oberbach, A.; Kirsch, K.; Lehmann, S.; Schlichting, N.; Fasshauer, M.; Zarse, K.; Stumvoll, M.; Ristow, M.; Blüher, M.; Kovacs, P. Serum vaspin concentrations are decreased after exercise-induced oxidative stress. Obes. Facts 2010, 3, 328–331. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.M.; Cho, G.J.; Yannakoulia, M.; Hwang, T.G.; Kim, I.H.; Park, E.K.; Mantzoros, C.S. Lifestyle modification increases circulating adiponectin concentrations but does not change vaspin concentrations. Metabolism 2011, 60, 1294–1299. [Google Scholar] [CrossRef] [PubMed]
- Oświęcimska, J.; Suwała, A.; Świętochowska, E.; Ostrowska, Z.; Gorczyca, P.; Ziora-Jakutowicz, K.; Machura, M.; Szczepańska, M.; Hyla-Klekot, L.; Kukla, M.; et al. Serum vaspin concentrations in girls with anorexia nervosa. J. Pediatr. Endocrinol. Metab. 2016, 29, 681–686. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.; Fouejeu Wamba, P.C.; Lapointe, M.; Poirier, P.; Martin, J.; Bastien, M.; Cianflone, K. Increased vaspin levels are associated with beneficial metabolic outcome pre- and post-bariatric surgery. PLoS ONE 2014, 23, e111002. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Group | |||
|---|---|---|---|---|
| KD (n = 24) | VPA (n = 26) | C (n = 22) | p | |
| Mean ± SD (min–max) | ||||
| Age [years] | 6.2 ± 2.2 (3.0–9.9) | 6.7 ± 2.3 (3.1–9.9) | 7.7 ± 1.6 (3.6–9.7) | 0.84 |
| Body mass [kg] | 19.73 ± 7.59 (9.1–39.0) | 24.76 ± 9.43 (15.5–54.1) | 28.25 ±6.7 (17.0–40.7) | 0.08 |
| Height [cm] | 112.65 ± 15.82 (86.0–138.0) | 120.4 ± 16.41 (95.0–157.5) | 129.17 ± 9.83 (103.0–141.5) | 0.07 |
| BMI [kg/m2] | 15.05 ± 2.27 (11.23–21.84) | 16.55 ± 2.57 (11.36–21.81) | 16.74 ± 2.46 (13.73–23.71) | 0.11 |
| Waist circ. [cm] | 50.58 ± 7.76 (39.5–67.0) | 54.98 ± 6.65 (45.0–74.0) | 58.15 ± 6.45 (48.0–74.0) | 0.05 |
| Hip circ. [cm] | 56.87 ± 10.12 (40.5–79.0) | 62.55 ± 8.32 (51.0–89.0) | 68.67 ± 8.34 (55.0–84.0) | 0.01 * |
| WHR | 0.89 ± 0.05 (0.74–0.98) | 0.88 ± 0.05 (0.78–1.00) | 0.85 ± 0.06 (0.71–0.98) | 0.1 |
| Total cholesterol [mg/dL] | 171.6 ± 42.12 (87.0–279.0) | 162.02 ± 29.92 (112.0–235.0) | 171.07 ± 27.54 (127.0–257.0) | 0.81 |
| LDL cholesterol (mg/dL) | 102.27 ± 34.34 (38.0–180.6) | 96.54 ± 28.03 (58.3–161.3) | 101.76 ± 23.51 (54.3–159.6) | 0.51 |
| HDL cholesterol [mg/dL] | 53.10 ± 17.04 (26.0–102.0) | 51.74 ± 8.5 (35.5–65.0) | 57.28 ± 12.8 (36.6–88.2) | 0.16 |
| Triglycerides [mg/dL] | 103.1 ± 53.34 (29.0–302.0) | 67.03 ± 43.79 (26.0–279.0) | 67.63 ± 37.52 (31.0–204.0) | 0.001 * # |
| AST [IU/L] | 30.2 ± 40.61 (6.0–219.0) | 27.2 ± 7.37 (18.0–43.0) | 26.8 ± 4.57 (19.0–37.0) | 0.19 |
| ALT [IU/L] | 35.37 ± 22.27 (11.0–120.0) | 15.43 ± 8.8 (6.0–54.0) | 15.77 ± 3.86 (9.0–22.0) | 0.04 # |
| Fasting glucose [mg/dL] | 72.03 ± 8.98 (53.0–90.0) | 86.47 ± 7.94 (64.0–101.0) | 88.5 ± 7.05 (74.0–101.0) | 0.001 * # |
| Insulin [μU/mL] | 2.48 ± 0.64 (1.34–3.75) | 4.19 ± 0.64 (3.13–5.3) | 4.55 ± 0.92 (3.09–5.98) | 0.001 * # |
| HOMA-IR | 0.44 ± 0.13 (0.22–0.7) | 0.91 ± 0.18 (0.66–1.29) | 1.0 ± 0.23 (0.65–1.51) | 0.001 * # |
| CRP [mg/L] | 0.9 ± 1.44 (0.2–5.6) | 0.56 ± 0.72 (0.2–3.8) | 1.87 ± 3.16 (0.1–5.7) | 0.42 |
| HCO3− [mmol/L] | 19.95 ± 2.03 (16.5–24.1) | 22.62 ± 1.68 (19.5–25.5) | 21.95 ± 1.08 (19.1–24.3) | 0.001 * # |
| BE [mEq/L] | −4.39 ± 2.14 (−7.9–0.4) | −1.68 ± 1.75 (−4.9–0.9) | −2.35 ± 1.05 (−4.2–(−0.5)) | 0.001 * # |
| BHBA [mmol/L] | 4.31 ± 1.62 (1.0–7.3) | 0.29 ± 0.23 (0.1–1.3) | 0.29 ± 0.25 (0.1–1.3) | 0.001 * # |
| Parameter | Adiponectin [μg/mL] | Omentin-1 [ng/mL] | Vaspin [ng/mL] |
|---|---|---|---|
| Age (years) | r = −0.04 p = 0.72 | r = −0.15 p = 0.22 | r = 0.02 p = 0.86 |
| Number of seizures/month # | r = 0.06 p = 0.69 | r = 0.08 p = 0.61 | r = −0.08 p = 0.60 |
| Body mass (kg) | r = −0.22 p = 0.07 | r = −0.34 p < 0.01 * | r = 0.28 p = 0.02* |
| Height (cm) | r = −0.14 p = 0.24 | r = −0.31 p = 0.01 * | r = 0.19 p = 0.11 |
| BMI (kg/m2) | r = −0.21 p = 0.08 | r = −0.22 p = 0.07 | r = 0.25 p = 0.04 * |
| Waist circ. (cm) | r = −0.19 p = 0.11 | r = −0.21 p = 0.08 | r = 0.21 p = 0.09 |
| Hip circ. (cm) | r = −0.29 p = 0.02 * | r = −0.27 p = 0.03 * | r = 0.21 p = 0.08 |
| WHR | r = −0.34 p = 0.01 * | r = −0.29 p = 0.02 * | r = −0.11 p = 0.38 |
| Total cholesterol (mg/dL) | r = 0.15 p = 0.21 | r = 0.14 p = 0.25 | r = −0.17 p = 0.17 |
| LDL cholesterol (mg/dL) | r = 0.11 p = 0.38 | r = 0.06 p = 0.64 | r = −0.08 p = 0.51 |
| HDL cholesterol (mg/dL) | r = −0.03 p = 0.82 | r = −0.01 p = 0.94 | r = −0.04 p = 0.75 |
| Triglycerides (mg/dL) | r = −0.36 p < 0.01 * | r = −0.39 p < 0.01 * | r = −0.36 p < 0.01 * |
| AST (IU/L) | r = 0.26 p = 0.03 * | r = 0.05 p = 0.69 | r = −0.05 p = 0.69 |
| ALT (IU/L) | r = 0.31 p = 0.01 * | r = 0.12 p = 0.32 | r = −0.15 p = 0.22 |
| Fasting glucose (mg/dL) | r = −0.34 p < 0.01 * | r = −0.49 p < 0.01 * | r = 0.50 p < 0.01 * |
| Insulin (μU/mL) | r = −0.32 p = 0.01 * | r = −0.59 p < 0.01 * | r = 0.66 p < 0.01 * |
| HOMA-IR | r = −0.35 p < 0.01 * | r = −0.62 p < 0.01 * | r = 0.65 p < 0.01 * |
| CRP (mg/L) | r = 0.10 p = 0.42 | r = 0.05 p = 0.72 | r = 0.09 p = 0.49 |
| HCO3− (mmol/L) | r = −0.26 p = 0.03 * | r = −0.41 p < 0.01 * | r = 0.41 p < 0.01 * |
| BE (mEq/L) | r = −0.32 p = 0.01 * | r = −0.41 p < 0.01 * | r = 0.42 p < 0.01 * |
| BHBA (mmol/L) | r = 0.44 p < 0.01 * | r = 0.58 p < 0.01 * | r = −0.76 p < 0.01 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chyra, M.; Roczniak, W.; Świętochowska, E.; Dudzińska, M.; Oświęcimska, J. The Effect of the Ketogenic Diet on Adiponectin, Omentin and Vaspin in Children with Drug-Resistant Epilepsy. Nutrients 2022, 14, 479. https://doi.org/10.3390/nu14030479
Chyra M, Roczniak W, Świętochowska E, Dudzińska M, Oświęcimska J. The Effect of the Ketogenic Diet on Adiponectin, Omentin and Vaspin in Children with Drug-Resistant Epilepsy. Nutrients. 2022; 14(3):479. https://doi.org/10.3390/nu14030479
Chicago/Turabian StyleChyra, Marcin, Wojciech Roczniak, Elżbieta Świętochowska, Magdalena Dudzińska, and Joanna Oświęcimska. 2022. "The Effect of the Ketogenic Diet on Adiponectin, Omentin and Vaspin in Children with Drug-Resistant Epilepsy" Nutrients 14, no. 3: 479. https://doi.org/10.3390/nu14030479
APA StyleChyra, M., Roczniak, W., Świętochowska, E., Dudzińska, M., & Oświęcimska, J. (2022). The Effect of the Ketogenic Diet on Adiponectin, Omentin and Vaspin in Children with Drug-Resistant Epilepsy. Nutrients, 14(3), 479. https://doi.org/10.3390/nu14030479