

Human Milk Oligosaccharides as Potential Antibiofilm Agents: A Review

 , , , , ,
, , , , ,  and
and Highlights
- Biofilms are highly antibiotic-tolerant communities of bacteria that pose serious health concerns in clinical settings.
- The potential application of human milk oligosaccharides as an alternative to antibiotics is discussed.
- Whether human milk oligosaccharides are safe (GRAS status) as therapeutics against bacterial adhesion and/or biofilms is reviewed.
- Application of machine learning-based computational tools is discussed as a perspective to improve HMO efficacy against bacteria and bacterial biofilms.
Abstract

1. Introduction
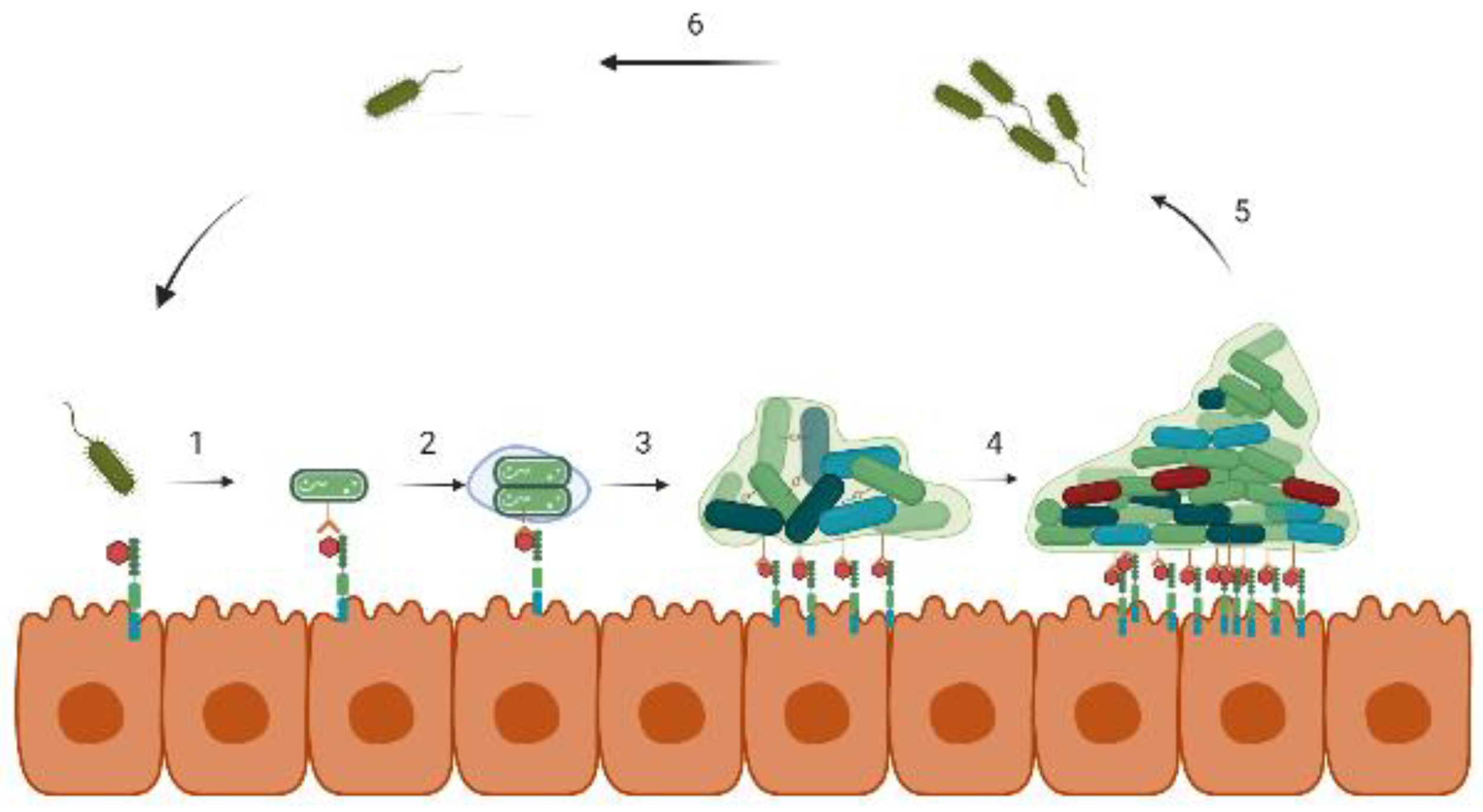
2. Biofilm Formation
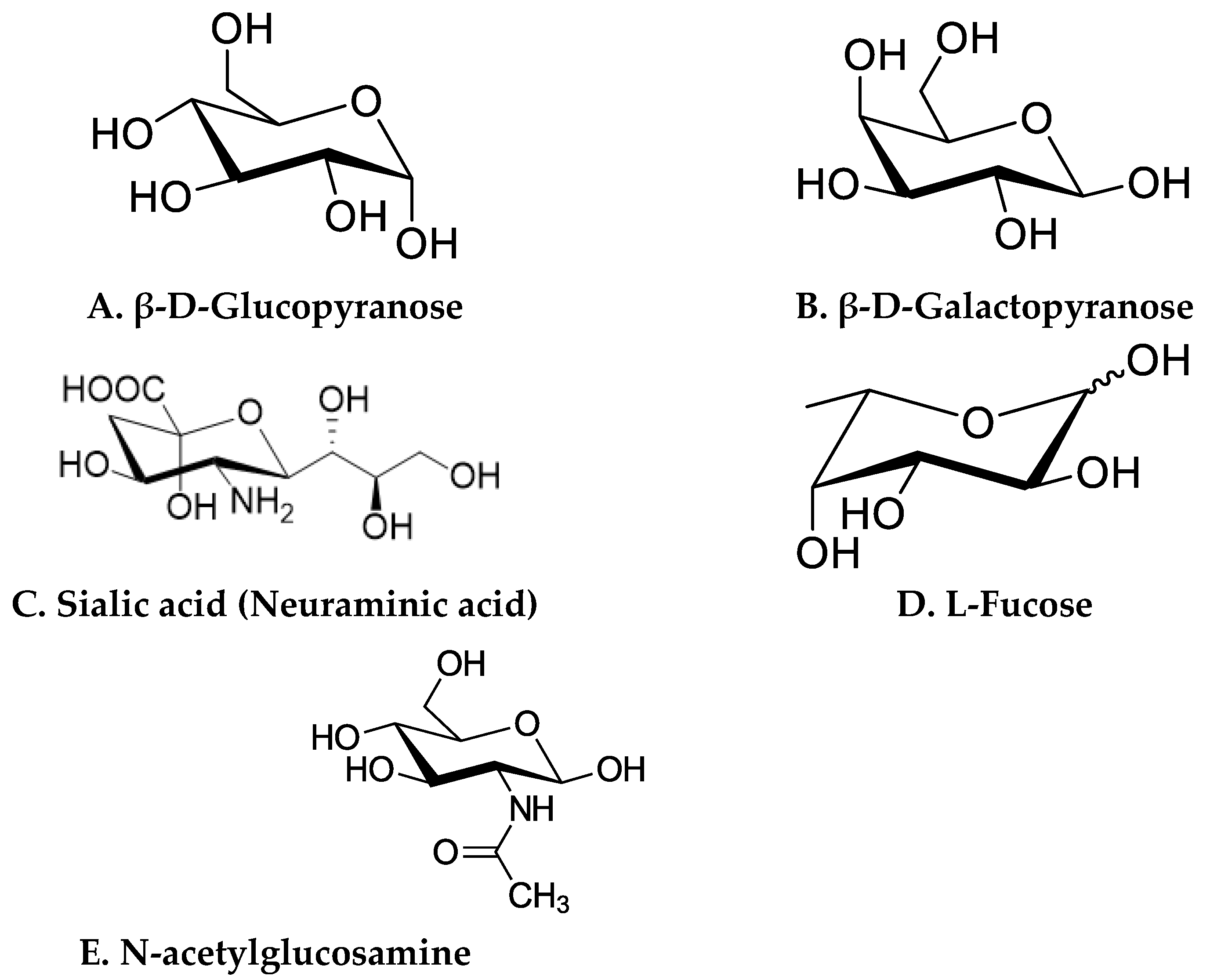
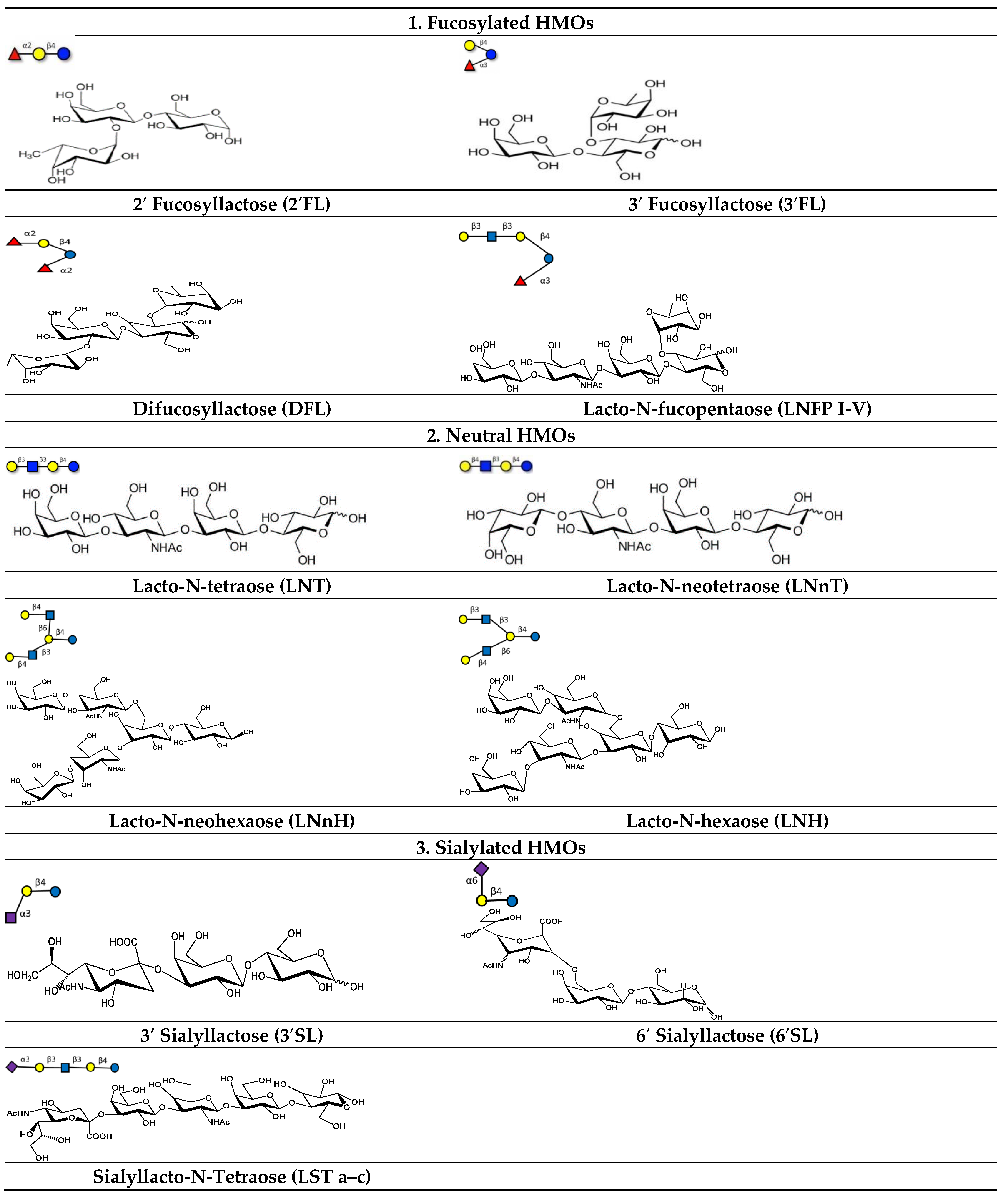
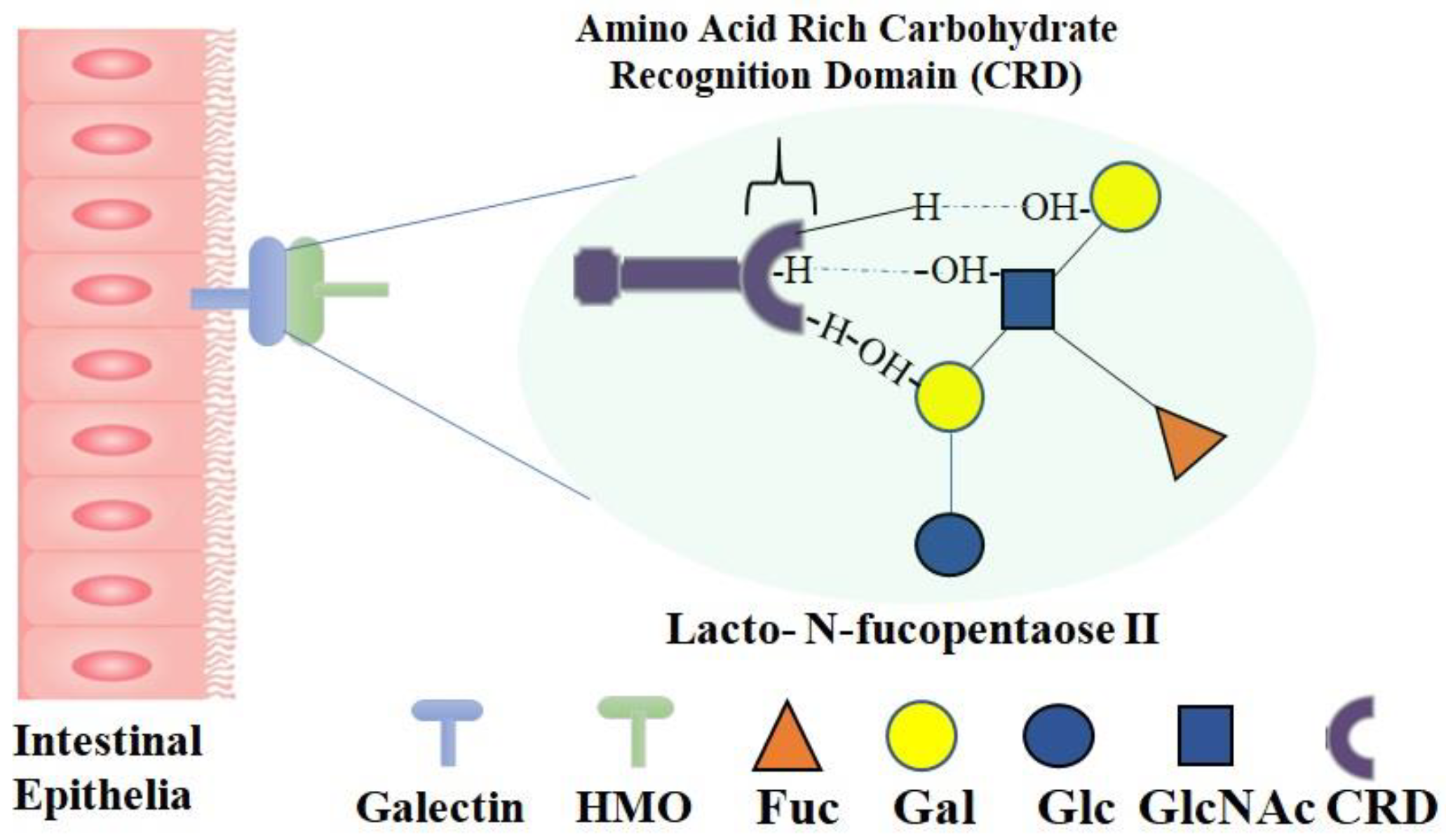
3. Different Classes of HMOs and Their Receptors
4. Antimicrobial Properties of Human Milk Oligosaccharides (HMOs)
4.1. HMOs Shape Gut Flora and Gut Immune Function
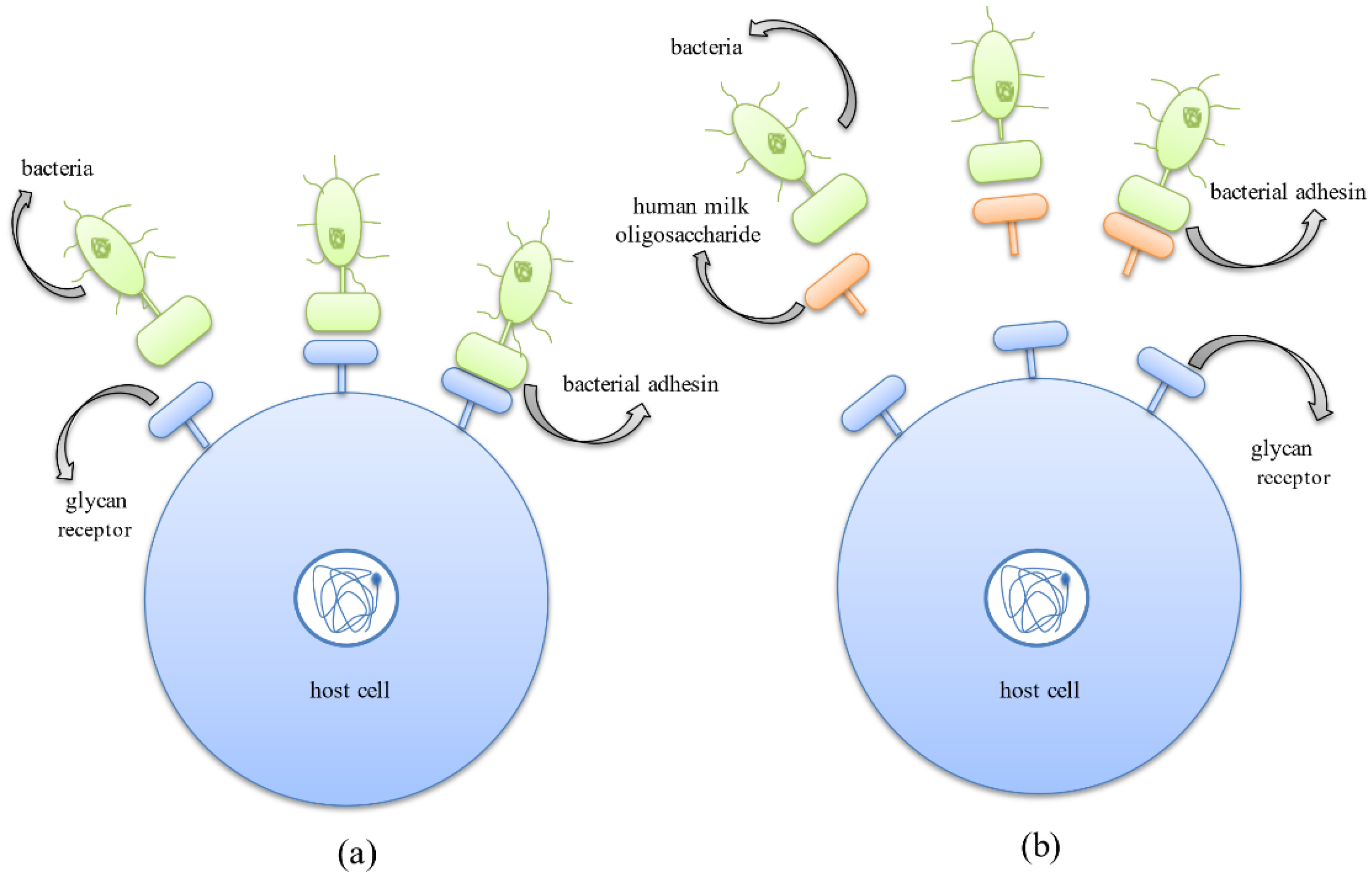
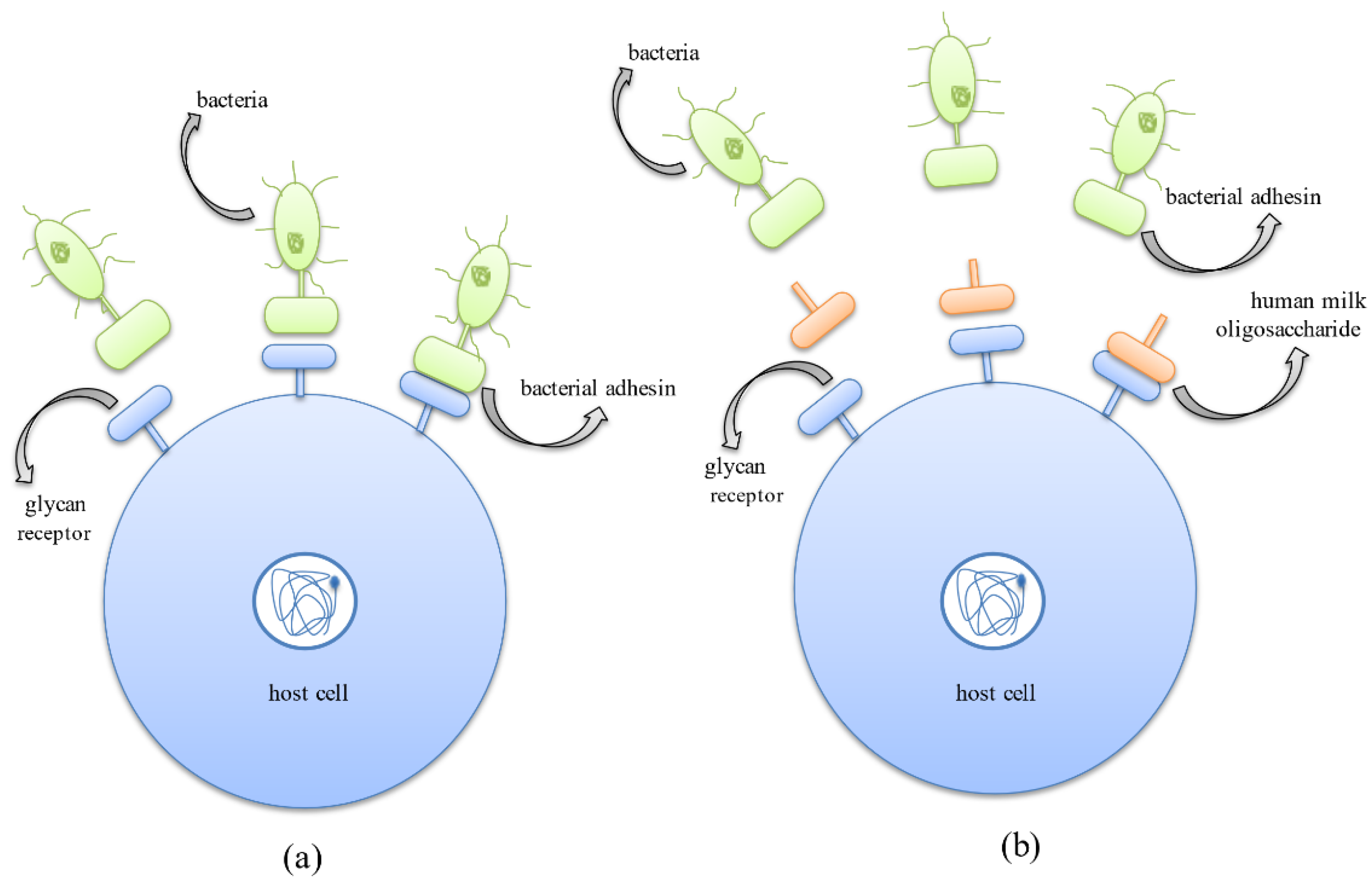
4.2. HMOs Prevent the Growth and Colonization of Pathogenic Bacteria
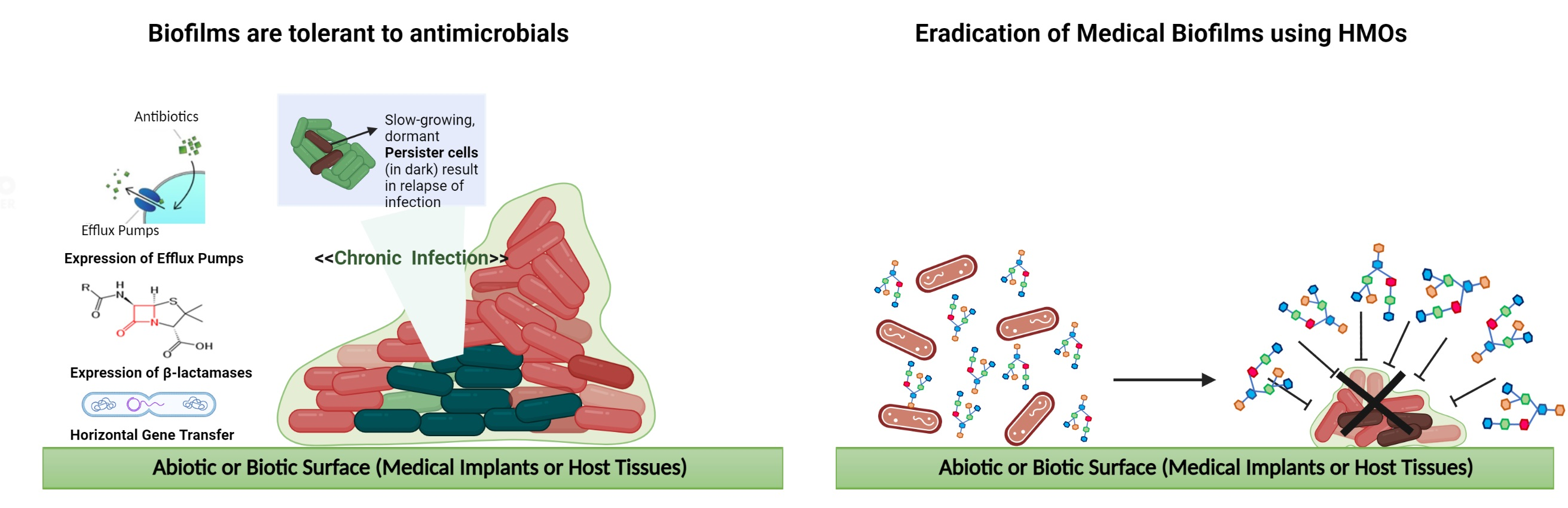
4.3. Antibiofilm Activity of HMOs
5. Conclusions and Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Bhowmik, A.; Malhotra, A.; Jana, S.; Chauhan, A. Biofilm Formation and Pathogenesis; Spinger: Berlin/Heidelberg, Germany, 2021; pp. 3–37. [Google Scholar]
- Singh, B.P.; Ghosh, S.; Chauhan, A. Development, dynamics and control of antimicrobial-resistant bacterial biofilms: A review. Environ. Chem. Lett. 2021, 19, 1983–1993. [Google Scholar] [CrossRef]
- Fuqua, C.; Filloux, A.; Ghigo, J.-M.; Visick, K.L. Biofilms 2018: A Diversity of Microbes and Mechanisms. J. Bacteriol. 2019, 201, e00118-19. [Google Scholar] [CrossRef] [PubMed]
- da Silva, R.A.G.; Afonina, I.; Kline, K.A. Eradicating biofilm infections: An update on current and prospective approaches. Curr. Opin. Microbiol. 2021, 63, 117–125. [Google Scholar] [PubMed]
- Flemming, H.C.; Wingender, J. The biofilm matrix. Nat. Rev. Microbiol. 2010, 8, 623–633. [Google Scholar] [CrossRef]
- Karygianni, L.; Ren, Z.; Koo, H.; Thurnheer, T. Biofilm Matrixome: Extracellular Components in Structured Microbial Communities. Trends Microbiol. 2020, 28, 668–681. [Google Scholar] [CrossRef] [PubMed]
- Canty, M.K.; Hansen, L.A.; Tobias, M.; Spencer, S.; Henry, T.; Luke-Marshall, N.R.; Campagnari, A.A.; Ehrensberger, M.T. Antibiotics Enhance Prevention and Eradication Efficacy of Cathodic-Voltage-Controlled Electrical Stimulation against Titanium-Associated Methicillin-Resistant Staphylococcus aureus and Pseudomonas aeruginosa Biofilms. mSphere 2019, 4, e00178-19. [Google Scholar] [CrossRef]
- Chauhan, A.; Bernardin, A.; Mussard, W.; Kriegel, I.; Estève, M.; Ghigo, J.-M.; Beloin, C.; Semetey, V. Preventing Biofilm Formation and Associated Occlusion by Biomimetic Glycocalyx like Polymer in Central Venous Catheters. J. Infect. Dis. 2014, 210, 1347–1356. [Google Scholar] [CrossRef]
- Chauhan, A.; Ghigo, J.-M.; Beloin, C. Study of in vivo catheter biofilm infections using pediatric central venous catheter implanted in rat. Nat. Protoc. 2016, 11, 525–541. [Google Scholar] [CrossRef]
- Sharma, D.; Misba, L.; Khan, A.U. Antibiotics versus biofilm: An emerging battleground in microbial communities. Antimicrob. Resist. Infect. Control 2019, 8, 76. [Google Scholar] [CrossRef]
- Lebeaux, D.; Chauhan, A.; Rendueles, O.; Beloin, C. From in vitro to in vivo Models of Bacterial Biofilm-Related Infections. Pathogens 2013, 2, 288–356. [Google Scholar] [CrossRef]
- Manciula, L.-G.; Jeican, I.I.; Tudoran, L.B.; Albu, S. Biofilms and inflammation in patients with chronic rhinosinusitis. Med. Pharm. Rep. 2020, 93, 374–383. [Google Scholar] [CrossRef] [PubMed]
- Dosler, S.; Hacioglu, M.; Yilmaz, F.N.; Oyardi, O. Biofilm modelling on the contact lenses and comparison of the in vitro activities of multipurpose lens solutions and antibiotics. PeerJ 2020, 8, e9419. [Google Scholar] [CrossRef] [PubMed]
- Koopaie, M.; Bordbar-Khiabani, A.; Kolahdooz, S.; Darbandsari, A.K.; Mozafari, M. Advanced surface treatment tech-niques counteract biofilm-associated infections on dental implants. Mater. Res. Express 2020, 7, 015417. [Google Scholar] [CrossRef]
- Diaconu, O.; Siriopol, I.; Poloșanu, L.I.; Grigoraș, I. Endotracheal Tube Biofilm and its Impact on the Pathogenesis of Ven-tilator-Associated Pneumonia. J. Crit. Care Med. 2018, 4, 50–55. [Google Scholar] [CrossRef]
- Staats, A.; Li, D.; Sullivan, A.C.; Stoodley, P. Biofilm formation in periprosthetic joint infections. Ann. Jt. 2021, 6, 43. [Google Scholar] [CrossRef]
- Gominet, M.; Compain, F.; Beloin, C.; Lebeaux, D. Central venous catheters and biofilms: Where do we stand in 2017? APMIS 2017, 125, 365–375. [Google Scholar] [CrossRef]
- He, E.; Serpelloni, S.; Alvear, P.; Rahimi, M.; Taraballi, F. Vascular Graft Infections: An Overview of Novel Treatments Using Nanoparticles and Nanofibers. Fibers 2022, 10, 12. [Google Scholar] [CrossRef]
- Bi, Y.; Xia, G.; Shi, C.; Wan, J.; Liu, L.; Chen, Y.; Wu, Y.; Zhang, W.; Zhou, M.; He, H.; et al. Therapeutic strategies against bacterial biofilms. Fundam. Res. 2021, 1, 193–212. [Google Scholar] [CrossRef]
- Asadpoor, M.; Ithakisiou, G.N.; van Putten JP, M.; Pieters, R.J.; Folkerts, G.; Braber, S. Antimicrobial Activities of Algi-nate and Chitosan Oligosaccharides Against Staphylococcus aureus and Group B Streptococcus. Front. Microbiol. 2021, 12, 700605. [Google Scholar] [CrossRef]
- Belorkar, S.A.; Gupta, A.K. Oligosaccharides: A boon from nature’s desk. AMB Express 2016, 6, 82. [Google Scholar] [CrossRef]
- Gibson, G.R.; Probert, H.M.; Loo, J.V.; Rastall, R.A.; Roberfroid, M.B. Dietary modulation of the human colonic microbi-ota: Updating the concept of prebiotics. Nutr. Res. Rev. 2004, 17, 259–275. [Google Scholar] [CrossRef] [PubMed]
- Roberfroid, M. Prebiotics: The Concept Revisited. J. Nutr. 2007, 137, 830S–837S. [Google Scholar] [CrossRef]
- Lordan, C.; Thapa, D.; Ross, R.P.; Cotter, P.D. Potential for enriching next-generation health-promoting gut bacteria through prebiotics and other dietary components. Gut Microbes 2020, 11, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Shoaf-Sweeney, K.D.; Hutkins, R.W. Adherence, anti-adherence, and oligosaccharides preventing pathogens from stick-ing to the host. Adv. Food Nutr. Res. 2009, 55, 101–161. [Google Scholar] [PubMed]
- Yuan, X.; Liu, J.; Li, R.; Zhou, J.; Wei, J.; Jiao, S.; Wang, Z.; Du, Y. Chitosan Oligosaccharides Coupling Inhibits Bacterial Biofilm-Related Antibiotic Resistance against Florfenicol. Molecules 2020, 25, 6043. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.; Tøndervik, A.; Sletta, H.; Klinkenberg, G.; Emanuel, C.; Onsøyen, E.; Myrvold, R.; Howe, R.A.; Walsh, T.R.; Hill, K.E.; et al. Overcoming Drug Resistance with Alginate Oligosaccharides Able To Potentiate the Action of Selected Antibiotics. Antimicrob. Agents Chemother. 2012, 56, 5134–5141. [Google Scholar] [CrossRef] [PubMed]
- Asadpoor, M.; Varasteh, S.; Pieters, R.J.; Folkerts, G.; Braber, S. Differential effects of oligosaccharides on the effec-tiveness of ampicillin against Escherichia coli in vitro. Pharma Nutr. 2021, 16, 100264. [Google Scholar]
- Mussatto, S.I.; Dragone, G.; Roberto, I.C. Ferulic and p-coumaric acids extraction by alkaline hydrolysis of brewer’s spent grain. Ind. Crops Prod. 2007, 25, 231–237. [Google Scholar] [CrossRef]
- Divyashri, G.; Sadanandan, B.; Chidambara Murthy, K.N.; Shetty, K.; Mamta, K. Neuroprotective Potential of Non-Digestible Oligosaccharides: An Overview of Experimental Evidence. Front. Pharmacol. 2021, 12, 712531. [Google Scholar] [CrossRef]
- Triantis, V.; Bode, L.; van Neerven, R.J. Immunological Effects of Human Milk Oligosaccharides. Front. Pediatr. 2018, 6, 190. [Google Scholar] [CrossRef]
- Walsh, C.; Lane, J.A.; van Sinderen, D.; Hickey, R.M. Human milk oligosaccharides: Shaping the infant gut microbiota and supporting health. J. Funct. Foods 2020, 72, 104074. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Li, L.-Q.; Liu, F.; Wu, J.-Y. Human milk oligosaccharides and infant gut microbiota: Molecular structures, utilization strategies and immune function. Carbohydr. Polym. 2021, 276, 118738. [Google Scholar] [CrossRef] [PubMed]
- Asadpoor, M.; Peeters, C.; Henricks, P.A.J.; Varasteh, S.; Pieters, R.J.; Folkerts, G.; Braber, S. Anti-Pathogenic Functions of Non-Digestible Oligosaccharides In Vitro. Nutrients 2020, 12, 1789. [Google Scholar] [CrossRef] [PubMed]
- Wiciński, M.; Sawicka, E.; Gębalski, J.; Kubiak, K.; Malinowski, B. Human Milk Oligosaccharides: Health Benefits, Potential Applications in Infant Formulas, and Pharmacology. Nutrients 2020, 12, 266. [Google Scholar] [CrossRef] [PubMed]
- Moro, E. Morphologie und bakteriologische Untersuchungen über die Darmbakterien des Säuglings: Die Bakterien-flora des normalen Frauenmilchstuhls. Jahrb. Kinderh. 1900, 61, 686–734. [Google Scholar]
- Bode, L. Human milk oligosaccharides: Every baby needs a sugar mama. Glycobiology 2012, 22, 1147–1162. [Google Scholar] [CrossRef]
- Craft, K.M.; Townsend, S.D. The Human Milk Glycome as a Defense Against Infectious Diseases: Rationale, Challenges, and Opportunities. ACS Infect. Dis. 2017, 4, 77–83. [Google Scholar] [CrossRef]
- Rousseaux, A.; Brosseau, C.; Le Gall, S.; Piloquet, H.; Barbarot, S.; Bodinier, M. Human Milk Oligosaccharides: Their Effects on the Host and Their Potential as Therapeutic Agents. Front. Immunol. 2021, 12, 680911. [Google Scholar] [CrossRef]
- Garrett, T.R.; Bhakoo, M.; Zhang, Z. Bacterial adhesion and biofilms on surfaces. Prog. Nat. Sci. 2008, 18, 1049–1056. [Google Scholar] [CrossRef]
- Herman-Bausier, P.; El-Kirat-Chatel, S.; Foster, T.J.; Geoghegan, J.A.; Dufrêne, Y.F. Staphylococcus aureus fibron-ectin-binding protein a mediates cell- cell adhesion through low-affinity homophilic bonds. MBio 2015, 6, 1–10. [Google Scholar] [CrossRef]
- O’toole, G.A.; Kolter, R. Flagellar and twitching motility are necessary for Pseudomonas aeruginosa biofilm development. Mol. Microbiol. 1998, 30, 295–304. [Google Scholar] [CrossRef] [PubMed]
- O’Connor, J.R.; Kuwada, N.J.; Huangyutitham, V.; Wiggins, P.A.; Harwood, C.S. Surface sensing and lateral subcellular localization of WspA, the receptor in a chemosensory-like system leading to c-di-GMP production. Mol. Microbiol. 2012, 86, 720–729. [Google Scholar] [CrossRef] [PubMed]
- de la Fuente-Núñez, C.; Reffuveille, F.; Fernández, L.; Hancock, R.E. Bacterial biofilm development as a multicellular adaptation: Antibiotic resistance and new therapeutic strategies. Curr. Opin. Microbiol. 2013, 16, 580–589. [Google Scholar] [CrossRef] [PubMed]
- Anderl, J.N.; Franklin, M.J.; Stewart, P.S. Role of Antibiotic Penetration Limitation in Klebsiella pneumoniae Biofilm Re-sistance to Ampicillin and Ciprofloxacin. ASM 2000, 44, 1818–1824. [Google Scholar]
- Wolfmeier, H.; Pletzer, D.; Mansour, S.C.; Hancock, R.E.W. New Perspectives in Biofilm Eradication. ACS Infect. Dis. 2017, 4, 93–106. [Google Scholar] [CrossRef] [PubMed]
- Guilhen, C.; Forestier, C.; Balestrino, D. Biofilm dispersal: Multiple elaborate strategies for dissemination of bacteria with unique properties. Mol. Microbiol. 2017, 105, 188–210. [Google Scholar] [CrossRef]
- Kuehl, R.; Al-Bataineh, S.; Gordon, O.; Luginbuehl, R.; Otto, M.; Textor, M.; Landmann, R. Furanone at subinhibitory con-centrations enhances Staphylococcal biofilm formation by luxS repression. Antimicrob. Agents Chemother. 2009, 53, 4159–4166. [Google Scholar] [CrossRef]
- Nelson, L.K.; D’Amours, G.H.; Sproule-Willoughby, K.M.; Morck, D.W.; Ceri, H. Pseudomonas aeruginosa las and rhl quorum-sensing systems are important for infection and inflammation in a rat prostatitis model. Microbiology 2009, 155, 2612–2619. [Google Scholar] [CrossRef]
- Morozov, V.; Hansman, G.; Hanisch, F.G.; Schroten, H.; Kunz, C. Human Milk Oligosaccharides as Promising Antivirals. Mol. Nutr. Food Res. 2018, 62, 1700679. [Google Scholar] [CrossRef]
- Zivkovic, A.M.; German, J.B.; Lebrilla, C.B.; Mills, D.A. Human milk glycobiome and its impact on the infant gastroin-testinal microbiota. Proc. Natl. Acad. Sci. USA 2011, 108, 4653–4658. [Google Scholar] [CrossRef]
- Tonon, K.M.; Miranda, A.; Abrão, A.C.F.; de Morais, M.B.; Morais, T.B. Validation and application of a method for the simultaneous absolute quantification of 16 neutral and acidic human milk oligosaccharides by graphitized carbon liquid chromatography—Electrospray ionization—Mass spectrometry. Food Chem. 2019, 274, 691–697. [Google Scholar] [CrossRef] [PubMed]
- Singh, K.S.; Singh, B.P.; Rokana, N.; Singh, N.; Kaur, J.; Singh, A.; Panwar, H. Bio-therapeutics from human milk: Pro-spects and perspectives. J. Appl. Microbiol. 2021, 131, 2669–2687. [Google Scholar] [CrossRef] [PubMed]
- Thurl, S.; Munzert, M.; Boehm, G.; Matthews, C.; Stahl, B. Systematic review of the concentrations of oligosaccharides in human milk. Nutr. Rev. 2017, 75, 920–933. [Google Scholar] [CrossRef] [PubMed]
- Özcan, E.; Sela, D.A. Inefficient Metabolism of the Human Milk Oligosaccharides Lacto-N-tetraose and Lac-to-N-neotetraose Shifts Bifidobacterium longum subsp. infantis Physiology. Front. Nutr. 2018, 5, 46. [Google Scholar] [PubMed]
- Wang, Y.; Zou, Y.; Wang, J.; Ma, H.; Zhang, B.; Wang, S. The protective effects of 2’-fucosyllactose against E. coli o157 infection are mediated by the regulation of gut microbiota and the inhibition of pathogen adhesion. Nutrients 2020, 12, 1284. [Google Scholar] [CrossRef]
- Csernák, O.; Rácz, B.; Alberti, Á.; Béni, S. Quantitative analysis of 3’- and 6’-sialyllactose in human milk samples by HPLC-MS/MS: A validated method for the comparison of two consecutive lactation periods in the same woman. J. Pharm. Biomed. Anal. 2020, 184, 113184. [Google Scholar] [CrossRef]
- Hirabayashi, J.; Hashidate, T.; Arata, Y.; Nishi, N.; Nakamura, T.; Hirashima, M.; Urashima, T.; Oka, T.; Futai, M.; Muller, W.E.; et al. Oligosaccharide specificity of galectins: A search by frontal affinity chromatography. Biochim. Biophys. Acta BBA Gen. Subj. 2002, 1572, 232–254. [Google Scholar] [CrossRef]
- Rapoport, E.M.; Kurmyshkina, O.V.; Bovin, N.V. Mammalian galectins: Structure, carbohydrate specificity, and functions. Biochemistry 2008, 73, 393–405. [Google Scholar] [CrossRef]
- López-Lucendo, M.F.; Solís, D.; André, S.; Hirabayashi, J.; Kasai, K.I.; Kaltner, H.; Gabius, H.J.; Romero, A. Growth-regulatory human galectin-1: Crystallographic characterisation of the structural changes induced by single-site mu-tations and their impact on the thermodynamics of ligand binding. J. Mol. Biol. 2004, 343, 957–970. [Google Scholar] [CrossRef]
- Wang, M.; Monaco, M.; Hauser, J.; Yan, J.; Dilger, R.; Donovan, S. Bovine Milk Oligosaccharides and Human Milk Oligosaccharides Modulate the Gut Microbiota Composition and Volatile Fatty Acid Concentrations in a Preclinical Neonatal Model. Microorganisms 2021, 9, 884. [Google Scholar] [CrossRef]
- Ten Bruggencate, S.J.M.; Bovee-Oudenhoven, I.M.; Feitsma, A.L.; Van Hoffen, E.; Schoterman, M.H. Functional role and mechanisms of sialyllactose and other sialylated milk oligosaccharides. Nutr. Rev. 2014, 72, 377–389. [Google Scholar] [CrossRef] [PubMed]
- Zenhom, M.; Hyder, A.; de Vrese, M.; Heller, K.J.; Roeder, T.; Rgen Schrezenmeir, J. Prebiotic Oligosaccharides Reduce Proinflammatory Cytokines in Intestinal Caco-2 Cells via Activation of PPARg and Peptidoglycan Recognition Protein 3 1-3. J. Nutr. Nutr. Immunol. 2011, 141, 971–977. [Google Scholar]
- Duncan, P.I.; Raymond, F.; Fuerholz, A.; Sprenger, N. Sialic Acid Utilisation and Synthesis in the Neonatal Rat Revisited. PLoS ONE 2009, 4, e8241. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.-T.; Nanda Nanthakumar, N.; Newburg, D.S. The Human Milk Oligosaccharide 2-Fucosyllactose Quenches Campylobacter jejuni-Induced Inflammation in Human Epithelial Cells HEp-2 and HT-29 and in Mouse Intestinal Mucosa 1-3. J. Nutr. 2016, 146, 1980–1990. [Google Scholar] [CrossRef] [PubMed]
- Bergstrom, K.S.B.; Xia, L. Mucin-type O-glycans and their roles in intestinal homeostasis. Glycobiology 2013, 23, 1026–1037. [Google Scholar] [CrossRef]
- You Wu, R.; Li, B.; Koike, Y.; Määttänen, P.; Miyake, H.; Cadete, M.; Johnson-Henry, K.C.; Botts, S.R.; Lee, C.; Abrahamsson, T.R.; et al. Human Milk Oligosaccharides Increase Mucin Expression in Experimental Necrotizing Enterocolitis. Mol. Nutr. Food Res. 2019, 63, 1800658. [Google Scholar]
- Wells, J.M.; Brummer, R.J.; Derrien, M.; MacDonald, T.T.; Troost, F.; Cani, P.D.; Theodorou, V.; Dekker, J.; Méheust, A.; De Vos, W.M.; et al. Homeostasis of the gut barrier and potential biomarkers. Am. J. Physiol. Liver Physiol. 2017, 312, G171–G193. [Google Scholar] [CrossRef]
- Šuligoj, T.; Vigsnæs, L.K.; Abbeele, P.V.D.; Apostolou, A.; Karalis, K.; Savva, G.M.; McConnell, B.; Juge, N. Effects of Human Milk Oligosaccharides on the Adult Gut Microbiota and Barrier Function. Nutrients 2020, 12, 2808. [Google Scholar] [CrossRef]
- Qiang, X.; YongLie, C.; QianBing, W. Health benefit application of functional oligosaccharides. Carbohydr. Polym. 2009, 77, 435–441. [Google Scholar] [CrossRef]
- Katayama, T. Host-derived glycans serve as selected nutrients for the gut microbe: Human milk oligosaccharides and bifidobacteria. Biosci. Biotechnol. Biochem. 2016, 80, 621–632. [Google Scholar] [CrossRef]
- Sela, D.A.; Garrido, D.; Lerno, L.; Wu, S.; Tan, K.; Eom, H.J.; Joachimiak, A.; Lebrilla, C.B.; Mills, D.A. Bifidobacterium longum subsp. infantis ATCC 15697 α-fucosidases are active on fucosylated human milk oligosaccharides. Appl. Environ. Microbiol. 2012, 78, 795–803. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; An, H.J.; Garrido, D.; German, J.B.; Lebrilla, C.B.; Mills, D.A. Proteomic Analysis of Bifidobacterium longum subsp. infantis Reveals the Metabolic Insight on Consumption of Prebiotics and Host Glycans. PLoS ONE 2013, 8, e57535. [Google Scholar] [CrossRef] [PubMed]
- Kelly, C.J.; Zheng, L.; Campbell, E.L.; Saeedi, B.; Scholz, C.C.; Bayless, A.J.; Wilson, K.E.; Glover, L.E.; Kominsky, D.J.; Magnuson, A.; et al. Crosstalk between Microbiota-Derived Short-Chain Fatty Acids and Intestinal Epithelial HIF Augments Tissue Barrier Function. Cell Host Microbe 2015, 17, 662–671. [Google Scholar] [CrossRef] [PubMed]
- Thorburn, A.N.; Mckenzie, C.I.; Shen, S.; Stanley, D.; Macia, L.; Mason, L.J.; Roberts, L.K.; Wong CH, Y.; Shim, R.; Robert, R.; et al. Evidence that asthma is a developmental origin disease influenced by maternal diet and bacterial metabolites. Nat. Commun. 2015, 6, 7320. [Google Scholar] [CrossRef] [PubMed]
- Richards, J.L.; Yap, Y.A.; McLeod, K.H.; MacKay, C.R.; Marinõ, E. Dietary metabolites and the gut microbiota: An al-ternative approach to control inflammatory and autoimmune diseases. Clin. Transl. Immunol. 2016, 5, e82. [Google Scholar] [CrossRef]
- Yu, Z.T.; Chen, C.; Newburg, D.S. Utilization of major fucosylated and sialylated human milk oligosaccharides by isolat-ed human gut microbes. Glycobiology 2013, 23, 1281–1292. [Google Scholar] [CrossRef]
- Craft, K.M.; Thomas, H.C.; Townsend, S.D. Sialylated variants of lacto-: N -tetraose exhibit antimicrobial activity against Group B Streptococcus. Org. Biomol. Chem. 2019, 17, 1893–1900. [Google Scholar] [CrossRef]
- Mejia, M.E.; Ottinger, S.; Vrbanac, A.; Babu, P.; Zulk, J.J.; Moorshead, D.; Bode, L.; Nizet, V.; Patras, K.A. Human Milk Oligosaccharides Reduce Murine Group B Streptococcus Vaginal Colonization with Minimal Impact on the Vaginal Microbiota. mSphere 2022, 7, e00885-21. [Google Scholar] [CrossRef]
- Suzuki, R.; Wada, J.; Katayama, T.; Fushinobu, S.; Wakagi, T.; Shoun, H.; Sugimoto, H.; Tanaka, A.; Kumagai, H.; Ashida, H.; et al. Structural and Thermodynamic Analyses of Solute-binding Protein from Bifidobacterium longum Specific for Core 1 Disaccharide and Lacto-N-biose I. J. Biol. Chem. 2008, 283, 13165–13173. [Google Scholar] [CrossRef]
- Kunz, C.; Rudloff, S. Biological functions of oligosaccharides in human milk. Acta Paediatr. 1993, 82, 903–912. [Google Scholar] [CrossRef]
- Wada, J.; Ando, T.; Kiyohara, M.; Ashida, H.; Kitaoka, M.; Yamaguchi, M.; Kumagai, H.; Katayama, T.; Yamamoto, K. Bifidobacterium bifidum lacto-N-biosidase, a critical enzyme for the degradation of human milk oligosaccharides with a type 1 structure. Appl. Environ. Microbiol. 2008, 74, 3996–4004. [Google Scholar] [CrossRef] [PubMed]
- DZopf, S.R. Oligosaccharide anti-infective agents. Lancet 1996, 347, 1017–1021. [Google Scholar]
- Morrin, S.T.; Buck, R.H.; Farrow, M.; Hickey, R.M. Milk-derived anti-infectives and their potential to combat bacterial and viral infection. J. Funct. Foods 2021, 81, 104442. [Google Scholar] [CrossRef]
- Cravioto, A.; Tello, A.; Villafan, H.; Ruiz, J.; del Vedovo, S.; Neeser, J.-R. Inhibition of Localized Adhesion of Enteropatho-genic Escherichia coli to HEp-2 Cells by Immunoglobulin and Oligosaccharide Fractions of Human Colostrum and Breast Milk Materials and Methods. J. Infect. Dis. 1991, 163, 1247–1255. [Google Scholar] [CrossRef] [PubMed]
- Andersson, B.; Porras, O.; Hanson, L.; Lagergård, T.; Svanborg-Edén, C. Inhibition of Attachment of Streptococcus pneumoniae and Haemophilus influenzae by Human Milk and Receptor Oligosaccharides. J. Infect. Dis. 1986, 153, 232–237. [Google Scholar] [CrossRef]
- Angeloni, S.; Ridet, J.L.; Kusy, N.; Gao, H.; Crevoisier, F.; Guinchard, S.; Kochhar, S.; Sigrist, H.; Sprenger, N. Glycopro-filing with micro-arrays of glycoconjugates and lectins. Glycobiology 2005, 15, 31–41. [Google Scholar] [CrossRef]
- Coppa, G.V.; Zampini, L.; Galeazzi, T.; Facinelli, B.; Ferrante, L.; Capretti, R.; Orazio, G. Human Milk Oligosaccharides Inhibit the Adhesion to Caco-2 Cells of Diarrheal Pathogens: Escherichia coli, Vibrio cholerae, and Salmonella fyris. Pediatr. Res. 2006, 59, 377–382. [Google Scholar] [CrossRef]
- Craft, K.M.; Townsend, S.D. 1-Amino-2′-fucosyllactose inhibits biofilm formation by Streptococcus agalactiae. J. Antibiot. 2019, 72, 507–512. [Google Scholar] [CrossRef]
- Weichert, S.; Jennewein, S.; Hüfner, E.; Weiss, C.; Borkowski, J.; Putze, J.; Schroten, H. Bioengineered 2′-fucosyllactose and 3-fucosyllactose inhibit the adhesion of Pseudomonas aeruginosa and enteric pathogens to human intestinal and respiratory cell lines. Nutr. Res. 2013, 33, 831–838. [Google Scholar] [CrossRef]
- Lin, A.E.; Autran, C.A.; Szyszka, A.; Escajadillo, T.; Huang, M.; Godula, K.; Prudden, A.R.; Boons, G.-J.; Lewis, A.L.; Doran, K.S.; et al. Human milk oligosaccharides inhibit growth of group B Streptococcus. J. Biol. Chem. 2017, 292, 11243–11249. [Google Scholar] [CrossRef]
- Jantscher-Krenn, E.; Zherebtsov, M.; Nissan, C.; Goth, K.; Guner, Y.S.; Naidu, N.; Choudhury, B.; Grishin, A.V.; Ford, H.R.; Bode, L. The human milk oligosaccharide disialyllacto-N-tetraose prevents necrotising enterocolitis in neonatal rats. Gut 2011, 61, 1417–1425. [Google Scholar] [CrossRef] [PubMed]
- Firon, N.; Ofek, I.; Sharon, N. Interaction of Mannose-Containing Oligosaccharides With the Fimbrial Lectin of Escherichia Coli. Biochem. Biophys. Res. Commun. 1982, 105, 1426–1432. [Google Scholar] [CrossRef]
- Searle, L.E.J.; Cooley, W.A.; Jones, G.; Nunez, A.; Crudgington, B.; Weyer, U.; Dugdale, A.; Tzortzis, G.; Collins, J.W.; Woodward, M.J.; et al. Purified galactooligosaccharide, derived from a mixture produced by the enzymic activity of Bifidobacterium bifidum, reduces Salmonella enterica serovar Typhimurium adhesion and invasion in vitro and in vivo. J. Med. Microbiol. 2010, 59, 1428–1439. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Qiao, Y.; Peng, Q.; Gao, L.; Shi, B. Inhibitory effects of YCW and MOS from Saccharomyces cerevisiae on Escherichia coli and Salmonella pullorum adhesion to Caco-2 cells. Front. Biol. 2017, 12, 370–375. [Google Scholar] [CrossRef]
- Al-Ghazzewi, F.H.; Tester, R.F. Inhibition of the adhesion of Escherichia coli to human epithelial cells by carbohydrates. Bioact. Carbohydr. Diet. Fibre 2014, 4, 1–5. [Google Scholar] [CrossRef]
- Kim, J.K.; Shin, E.C.; Park, H.G. Fructooligosaccharides decreased the ability of probiotic Escherichia coli Nissle 1917 to adhere to co-cultures of human intestinal cell lines. J. Korean Soc. Appl. Biol. Chem. 2015, 58, 45–52. [Google Scholar] [CrossRef]
- Yue, H.; Han, Y.; Yin, B.; Cheng, C.; Liu, L. Comparison of the antipathogenic effect toward Staphylococcus aureus of N -linked and free oligosaccharides derived from human, bovine, and goat milk. J. Food Sci. 2020, 85, 2329–2339. [Google Scholar] [CrossRef]
- Wang, W.L.; Wang, W.; Du, Y.M.; Wu, H.; Yu, X.B.; Ye, K.P.; Li, C.B.; Jung, Y.S.; Qian, Y.J.; Voglmeir, J.; et al. Com-parison of anti-pathogenic activities of the human and bovine milk N-glycome: Fucosylation is a key factor. Food Chem. 2017, 235, 167–174. [Google Scholar] [CrossRef]
- Craft, K.M.; Thomas, H.C.; Townsend, S.D. Interrogation of Human Milk Oligosaccharide Fucosylation Patterns for An-timicrobial and Antibiofilm Trends in Group B Streptococcus. ACS Infect. Dis. 2018, 4, 1755–1765. [Google Scholar] [CrossRef]
- Marotta, M.; Ryan, J.T.; Hickey, R.M. The predominant milk oligosaccharide 6′-sialyllactose reduces the internalisation of Pseudomonas aeruginosa in human pneumocytes. J. Funct. Foods 2014, 6, 367–373. [Google Scholar] [CrossRef]
- Craft, K.M.; Townsend, S.D. Mother Knows Best: Deciphering the Antibacterial Properties of Human Milk Oligosaccha-rides. Acc. Chem. Res. 2019, 52, 760–768. [Google Scholar] [CrossRef] [PubMed]
- Ackerman, D.L.; Doster, R.S.; Weitkamp, J.-H.; Aronoff, D.; Gaddy, J.A.; Townsend, S.D. Human Milk Oligosaccharides Exhibit Antimicrobial and Anti-Biofilm Properties Against Group B Streptococcus. ACS Infect Dis. 2017, 3, 595–605. [Google Scholar] [CrossRef] [PubMed]
- Ackerman, D.L.; Craft, K.M.; Doster, R.S.; Weitkamp, J.H.; Aronoff, D.M.; Gaddy, J.A.; Townsend, S.D. Antimicrobial and Antibiofilm Activity of Human Milk Oligosaccharides against Streptococcus agalactiae, Staphylococcus aureus, and Acineto-bacter baumannii. ACS Infect. Dis. 2018, 4, 315–324. [Google Scholar] [CrossRef]
- Spicer, S.K.; Moore, R.E.; Lu, J.; Guevara, M.A.; Marshall, D.R.; Manning, S.D. Antibiofilm Activity of Human Milk Oligosaccharides against Multidrug Resistant and Susceptible Isolates of Acinetobacter baumannii. ACS Infect. Dis. 2021, 7, 3254–3263. [Google Scholar] [CrossRef] [PubMed]
- Craft, K.M.; Gaddy, J.A.; Townsend, S.D. Human Milk Oligosaccharides (HMOs) Sensitize Group B Streptococcus to Clindamycin, Erythromycin, Gentamicin, and Minocycline on a Strain Specific Basis. ACS Chem. Biol. 2018, 13, 2020–2026. [Google Scholar] [CrossRef]
- Moore, R.E.; Craft, K.M.; Xu, L.L.; Chambers, S.A.; Nguyen, J.M.; Marion, K.C.; Gaddy, J.A.; Townsend, S.D. Leveraging Stereoelectronic Effects in Biofilm Eradication: Synthetic β-Amino Human Milk Oligosaccharides Impede Microbial Adhesion As Observed by Scanning Electron Microscopy. J. Org. Chem. 2021, 85, 16128–16135. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Palacios, G.M.; Cervantes, L.E.; Ramos, P.; Chavez-Munguia, B.; Newburg, D.S. Campylobacter jejuni binds intesti-nal H(O) antigen (Fucα1, 2Galβ1, 4GlcNAc), and fucosyl oligosaccharides of human milk inhibit its binding and infection. J. Biol. Chem. 2003, 278, 14112–14120. [Google Scholar] [CrossRef]
- Jarzynka, S.; Spott, R.; Tchatchiashvili, T.; Ueberschaar, N.; Martinet, M.G.; Strom, K.; Kryczka, T.; Wesołowska, A.; Pletz, M.W.; Olędzka, G.; et al. Human Milk Oligosaccharides Exhibit Biofilm Eradication Activity Against Matured Biofilms Formed by Different Pathogen Species. Front. Microbiol. 2022, 12, 794441. [Google Scholar] [CrossRef]
- Morrow, A.L.; Ruiz-Palacios, G.M.; Altaye, M.; Jiang, X.; Guerrero, M.L.; Meinzen-Derr, J.K.; Farkas, T.; Chaturvedi, P.; Pickering, L.K.; Newburg, D.S. Human milk oligosaccharides are associated with protection against diarrhea in breast-fed infants. J. Pediatr. 2004, 145, 297–303. [Google Scholar] [CrossRef]
- El-Hawiet, A.; Kitova, E.N.; Klassen, J.S. Recognition of human milk oligosaccharides by bacterial exotoxins. Glycobiology 2015, 25, 845–854. [Google Scholar] [CrossRef]
- Martín-Sosa, S.; Martín, M.-J.; Hueso, P. Nutrient Metabolism The Sialylated Fraction of Milk Oligosaccharides Is Partially Responsible for Binding to Enterotoxigenic and Uropathogenic Escherichia coli Human Strains. J. Nutr. 2002, 132, 3067–3072. [Google Scholar] [CrossRef] [PubMed]
- Murphy, K.; Park, A.J.; Hao, Y.; Brewer, D.; Lam, J.S.; Khursigaraa, C.M. Influence of O polysaccharides on biofilm de-velopment and outer membrane vesicle biogenesis in Pseudomonas aeruginosa PAO1. J. Bacteriol. 2014, 196, 1306–1317. [Google Scholar] [CrossRef] [PubMed]
- Arciola, C.R.; Campoccia, D.; Ravaioli, S.; Montanaro, L. Polysaccharide intercellular adhesin in biofilm: Structural and regulatory aspects. Front. Cell. Infect. Microbiol. 2015, 5, 7. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, S.; Chauhan, A. Chapter 11—Anti-influenza agents. In Viral Infections and Antiviral Therapies; Dhara, A.K., Nayak, A.K., Eds.; Academic Press: Cambridge, MA, USA, 2023; pp. 211–239. [Google Scholar]
- Koo, H.; Allan, R.N.; Howlin, R.P.; Stoodley, P.; Hall-Stoodley, L. Targeting microbial biofilms: Current and prospective therapeutic strategies. Nat. Rev. Microbiol. 2017, 15, 740–755. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Biofilm Infection | Associated Pathogens | References | |
|---|---|---|---|
| Tissue-associated infections | Chronic rhinosinusitis | Corynebacterium tuberculostearicum, Haemophilus influenzae, Lactobacillus sakei, P. aeruginosa, S. aureus, Streptococcus pneumoniae | [11,12] |
| Periodontitis | Fusobacterium nucleatum, Porphyromonus gingivalis, Tannerella forsythia, Treponema denticola, | [1,11] | |
| Pharyngitis: | Group A Streptococcus (GAS), H. influenzae, S. aureus | ||
| Otitis media | GAS, H. influenzae, P. aeruginosa, S. pneumoniae | ||
| Infective endocarditis | Lactobacillus lactis, S. aureus | ||
| Cystic fibrosis | Burkholderia cenocepacia, H. influenza, P. aeruginosa, S. aureus | ||
| Colorectal cancer and ulcerative colitis | Bacteriodes fragilis, Enterobacteriaceae, E. coli Fusobacterium spp., Shigella spp. | ||
| Vaginosis | Bacteroides, Gardnerella vaginalis, Mycoplasma | ||
| Urinary tract infections | E. coli, Enterobacter spp., Klebsiella pneumonia, Proteus spp., Staphylococcus saprophyticus | ||
| Prostatitis | E. coli | ||
| Osteomyelitis | E. coli, H. influenzae, Streptococcus agalactiae, S. aureus, Streptococcus pyogenes | ||
| Wound infections | P. aeruginosa, S. aureus | ||
| Device-associated Infections | Contact lenses | P. aeruginosa, S. aureus | [13] |
| Dental implants | Aggregatibacter actinomycetemcomitans, Eikenella corrodens, P. gingivalis, | [14] | |
| Endotracheal tubes | Acinetobacter baumannii, Enterobacter spp., Enterococcus faecalis, K. pneumoniae, P. aeruginosa, S. aureus | [15] | |
| Prosthetic joints | P. aeruginosa, S. aureus, S. epidermidis | [16] | |
| Vascular catheters | K. pneumonia, P. aeruginosa, Coagulase-negative Staphylococci, S. aureus | [17] | |
| Vascular grafts | P. aeruginosa, S. aureus | [18] |
| HMOs | Pathogens | Antibacterial Properties | References |
|---|---|---|---|
| 2′-Fucosyllactose (2′-FL) | C. jejuni, Enteropathogenic E. coli, P. aeruginosa, S. enterica serovar fyris, S. dysenteriae, | Interferes with binding of specific receptors on epithelial cells and hence prevents infection development | [90,108,110,111] |
| A. baumanii | Represses biofilm formation | [105] | |
| 3-Fucosyllactose (3′-FL) | C. jejuni, E. coli, P. aeruginosa | Prevents adhesion of bacteria to receptors | [88,90,108] |
| Group B Streptococcus | 10% decrease in biofilm formation | [90] | |
| A. baumanii | Represses biofilm formation | [105] | |
| 3′-sialyllactose (3′-SL) | E. coli, H. pylori, S. fyris, V. cholerae | Inhibits bacterial adhesion | [87,88,112] |
| GBS | 10% decrease in biofilm biomass | [102] | |
| 6′-sialyllactose (6′-SL) | E. coli, S. fyris, V. cholerae | Interferes with bacterial adhesion | [87] |
| GBS | 9% decrease in biofilm biomass | [102] | |
| 1-amino-2′-FL | GBS 590 | 37% decrease in biofilm production | [89] |
| GBS 2 | 46% decrease in biofilm production | ||
| Lacto-N-neotetraose (LNnT) | GBS | 13% decrease in biofilm biomass | [102] |
| A. baumanii | Represses biofilm formation | [103] | |
| Difucosyllactose (DFL) | A. baumanii | Represses biofilm formation | |
| Lacto-N-fucopentaose I (LNFP I) | A. baumanii | Represses biofilm formation | |
| Lacto-N-fucopentaose II (LNFP II) | A. baumanii | Represses biofilm formation | |
| Lacto-N-fucopentaose III (LNFP III) | A. baumanii | Represses biofilm formation | |
| Lacto-N-triose II (LNT II) | A. baumanii | Represses biofilm formation |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bhowmik, A.; Chunhavacharatorn, P.; Bhargav, S.; Malhotra, A.; Sendrayakannan, A.; Kharkar, P.S.; Nirmal, N.P.; Chauhan, A. Human Milk Oligosaccharides as Potential Antibiofilm Agents: A Review. Nutrients 2022, 14, 5112. https://doi.org/10.3390/nu14235112
Bhowmik A, Chunhavacharatorn P, Bhargav S, Malhotra A, Sendrayakannan A, Kharkar PS, Nirmal NP, Chauhan A. Human Milk Oligosaccharides as Potential Antibiofilm Agents: A Review. Nutrients. 2022; 14(23):5112. https://doi.org/10.3390/nu14235112
Chicago/Turabian StyleBhowmik, Ankurita, Phatchada Chunhavacharatorn, Sharanya Bhargav, Akshit Malhotra, Akalya Sendrayakannan, Prashant S. Kharkar, Nilesh Prakash Nirmal, and Ashwini Chauhan. 2022. "Human Milk Oligosaccharides as Potential Antibiofilm Agents: A Review" Nutrients 14, no. 23: 5112. https://doi.org/10.3390/nu14235112
APA StyleBhowmik, A., Chunhavacharatorn, P., Bhargav, S., Malhotra, A., Sendrayakannan, A., Kharkar, P. S., Nirmal, N. P., & Chauhan, A. (2022). Human Milk Oligosaccharides as Potential Antibiofilm Agents: A Review. Nutrients, 14(23), 5112. https://doi.org/10.3390/nu14235112