
Regular Supplementation with Antioxidants Rescues Doxorubicin-Induced Bone Deformities and Mineralization Delay in Zebrafish


Abstract
1. Introduction
2. Materials and Methods
2.1. Housing Conditions
2.2. Micro Diet Preparation
2.3. Feeding Trial
2.4. Whole-Mount Double Staining and Evaluation of Skeletal Anomalies
2.5. Mineral Contents
2.6. Lipid Peroxidation (MDA) Analysis
2.7. RNA Extraction and qPCR
2.8. Histology
2.9. Statistical Analysis
3. Results
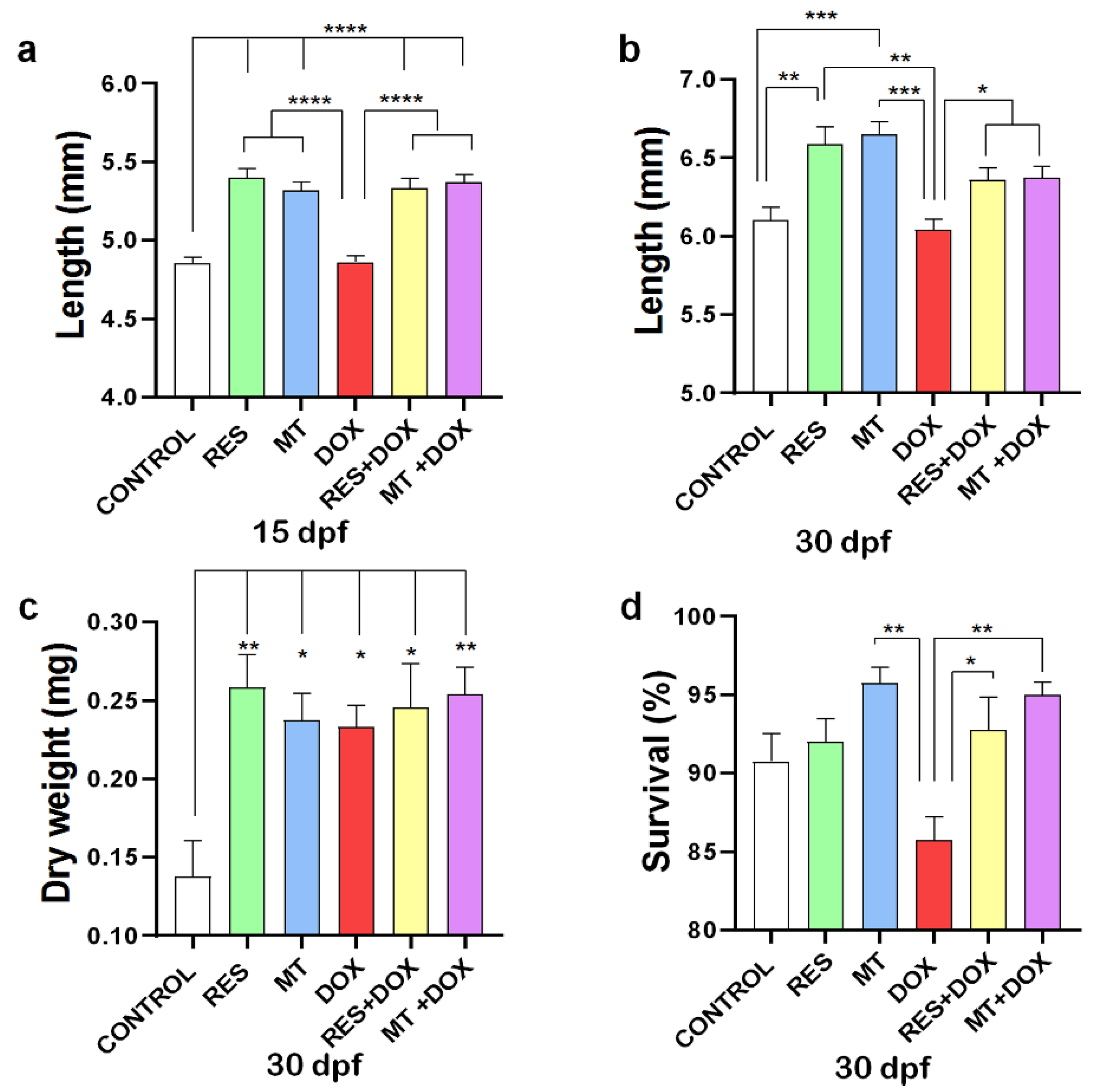
3.1. Fish Growth and Survival
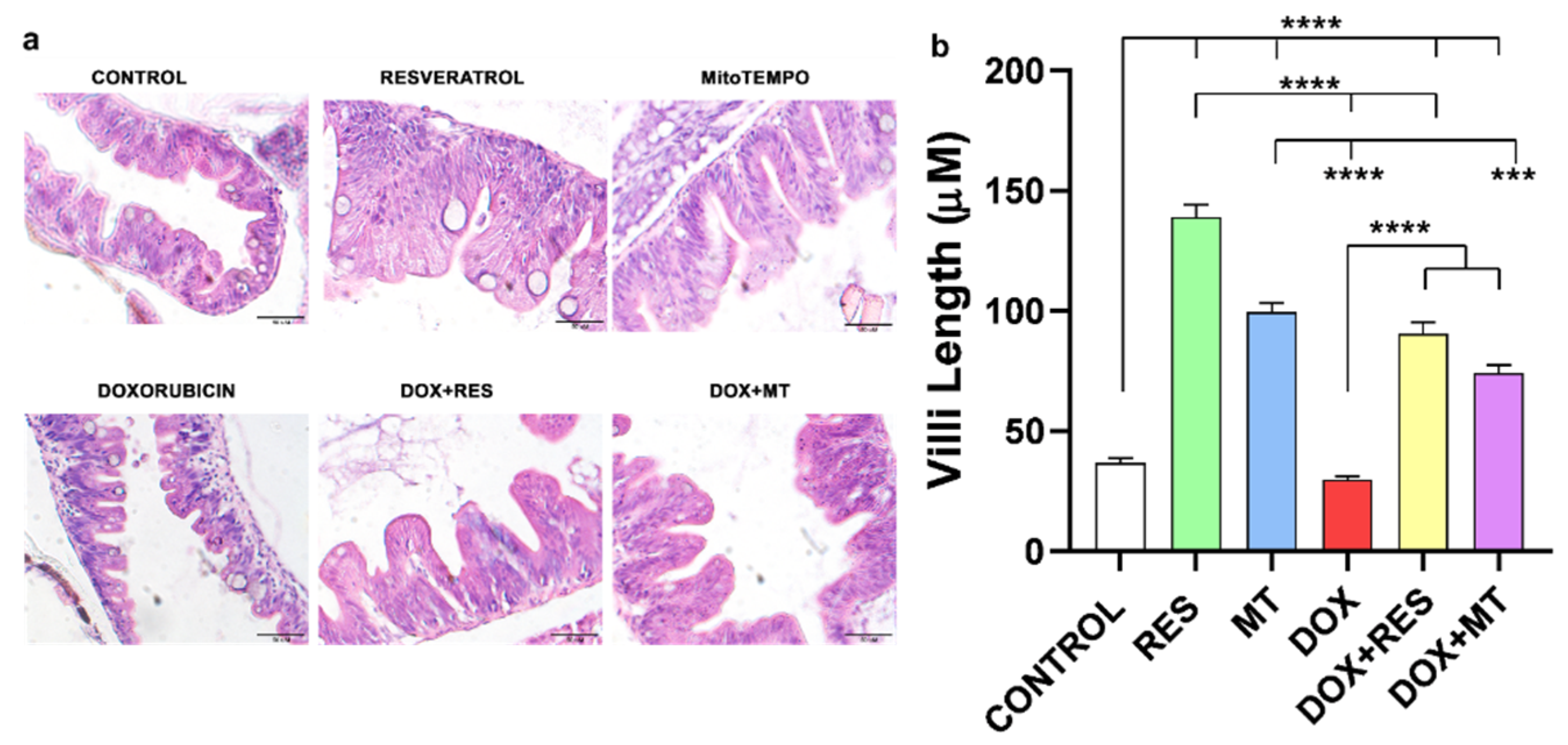
3.2. Intestinal Villi Morphology on Antioxidant and Pro-Oxidant Supplemented Groups
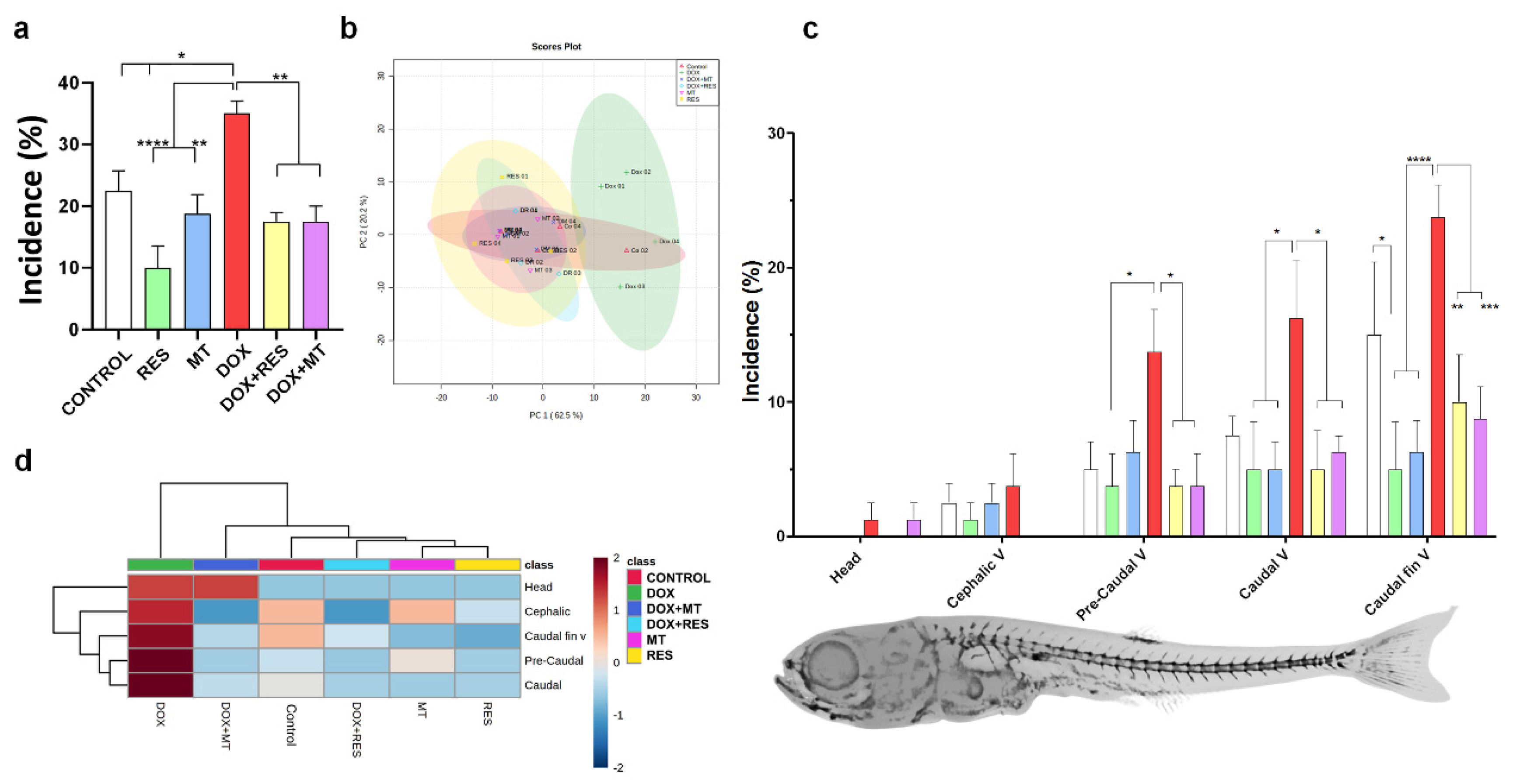
3.3. Antioxidants Prevent DOX-Induced Skeletal Deformities
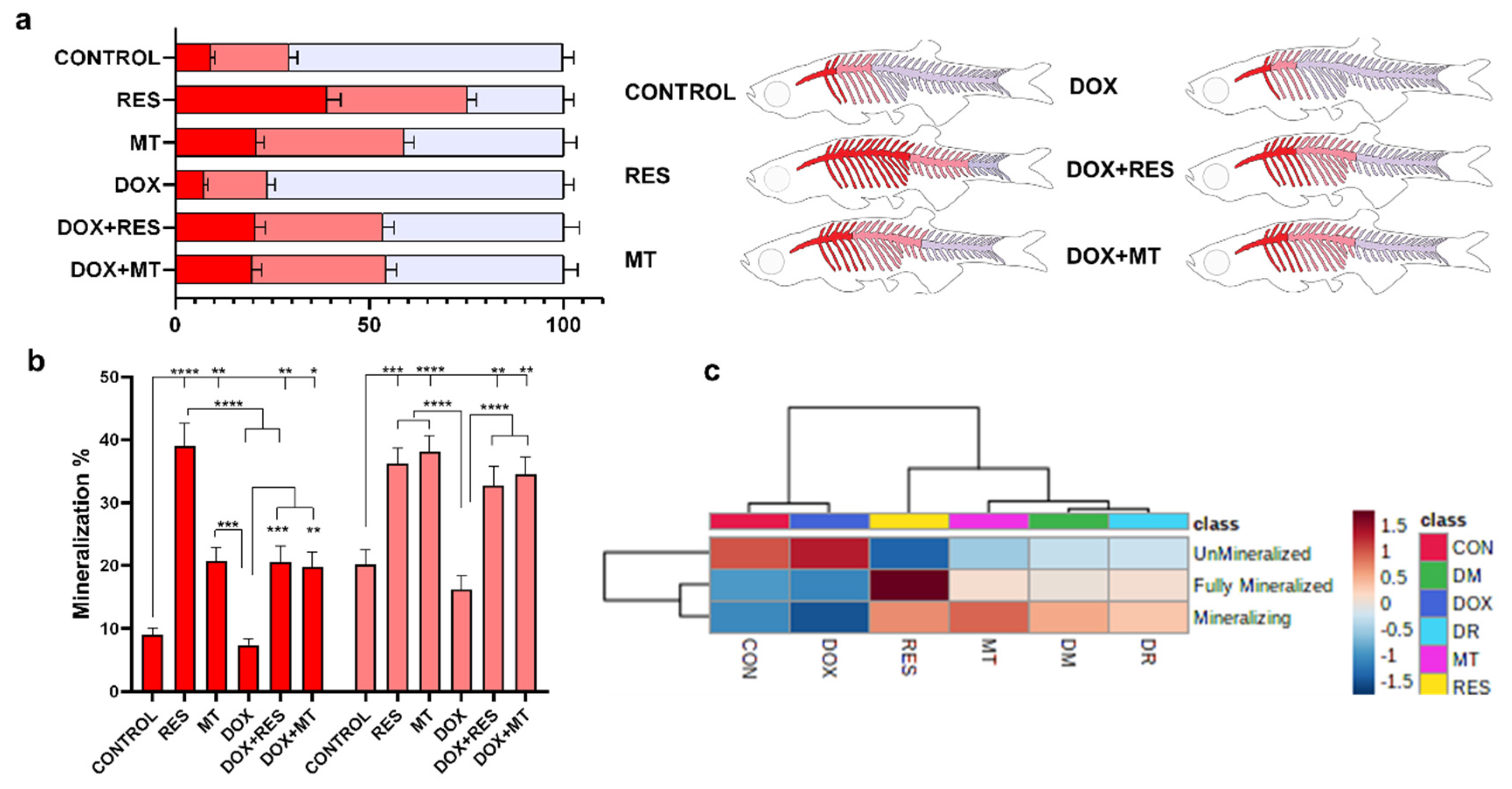
3.4. Antioxidants Improve Mineralization of the Axial Skeleton
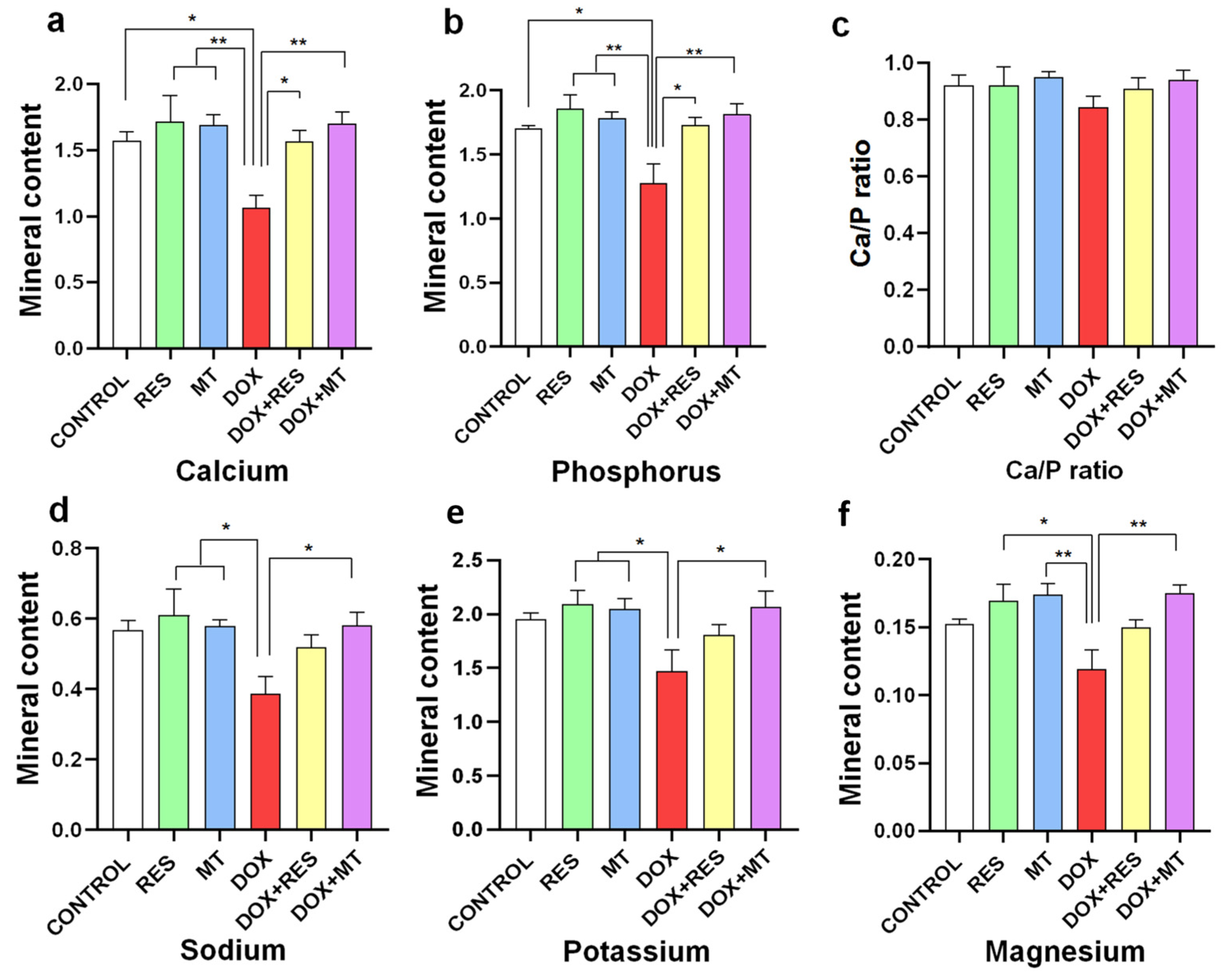
3.5. Doxorubicin Affects Mineral Content
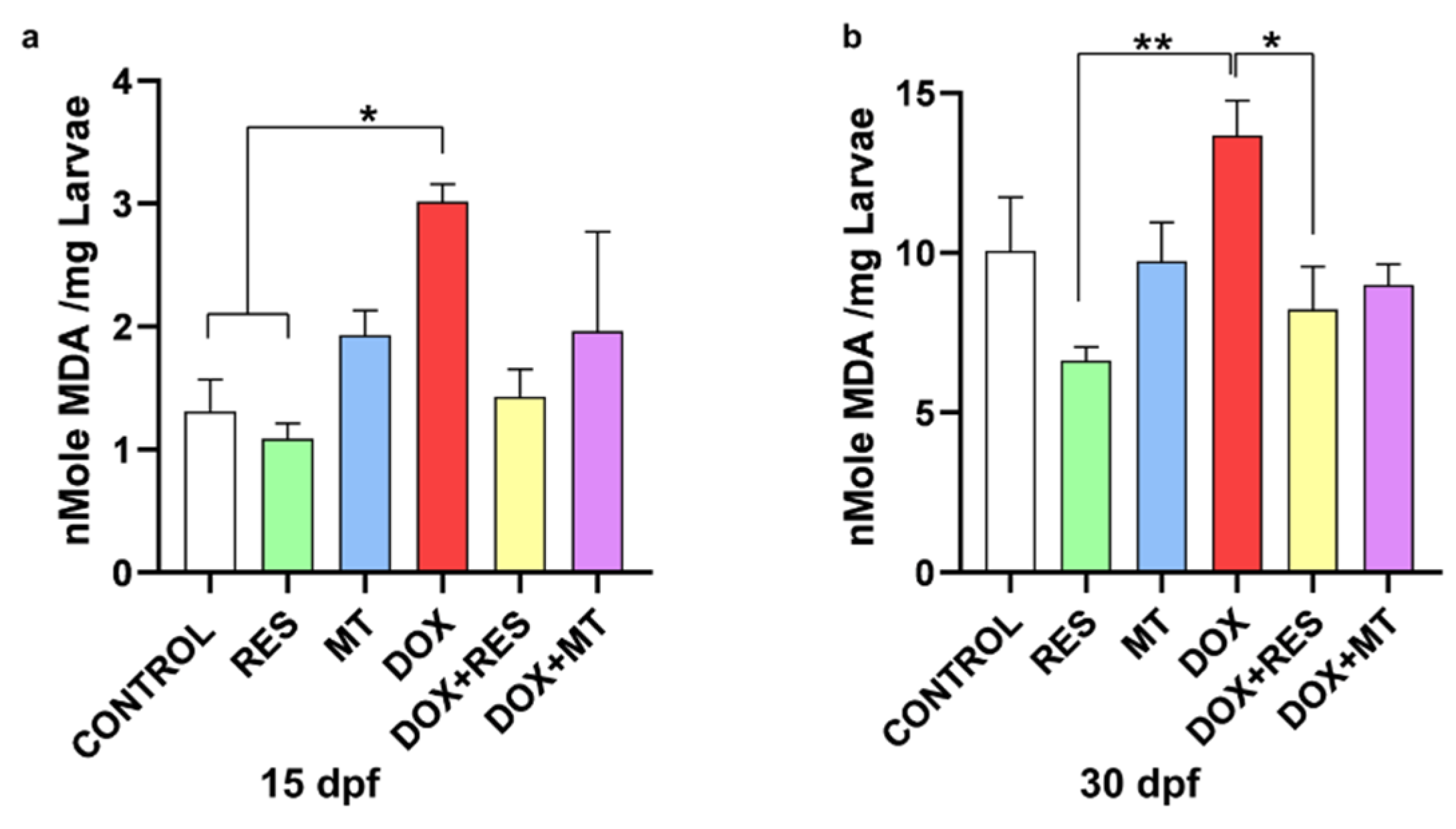
3.6. Antioxidants Reverse Doxorubicin-Induced Oxidative Stress
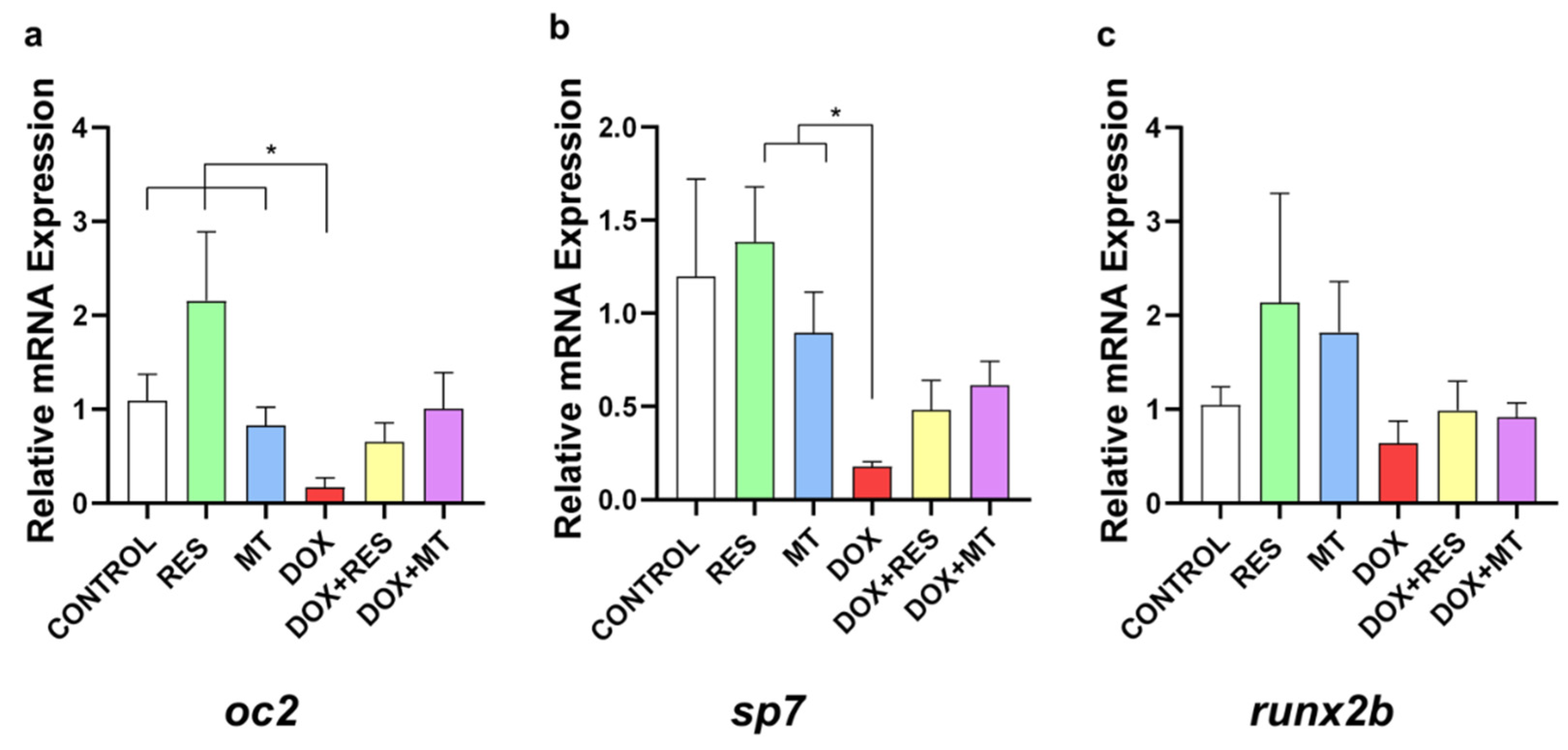
3.7. Doxorubicin-Induced Effects on Osteoblast Differentiation Markers
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mirza, F.; Canalis, E. Secondary osteoporosis: Pathophysiology and management. Eur. J. Endocrinol. 2015, 173, R131–R151. [Google Scholar] [CrossRef]
- Painter, S.E.; Kleerekoper, M.; Camacho, P.M. Secondary osteoporosis: A review of the recent evidence. Endocr. Pract. Off. J. Am. Coll. Endocrinol. Am. Assoc. Clin. Endocrinol. 2006, 12, 436–445. [Google Scholar] [CrossRef] [PubMed]
- US Department of Health and Human Services. Bone Health and Osteoporosis: A report of the Surgeon General; U.S. Department of Health and Human Services, Office of the Surgeon General: Washington, DC, USA, 2004; p. 436. [Google Scholar]
- Pfeilschifter, J.; Diel, I.J. Osteoporosis Due to Cancer Treatment: Pathogenesis and Management. J. Clin. Oncol. 2000, 18, 1570–1593. [Google Scholar] [CrossRef] [PubMed]
- Baek, K.H.; Oh, K.W.; Lee, W.Y.; Lee, S.S.; Kim, M.K.; Kwon, H.S.; Rhee, E.J.; Han, J.H.; Song, K.H.; Cha, B.Y.; et al. Association of oxidative stress with postmenopausal osteoporosis and the effects of hydrogen peroxide on osteoclast formation in human bone marrow cell cultures. Calcif. Tissue Int. 2010, 87, 226–235. [Google Scholar] [CrossRef] [PubMed]
- Maggio, D.; Barabani, M.; Pierandrei, M.; Polidori, M.C.; Catani, M.; Mecocci, P.; Senin, U.; Pacifici, R.; Cherubini, A. Marked decrease in plasma antioxidants in aged osteoporotic women: Results of a cross-sectional study. J. Clin. Endocrinol. Metab. 2003, 88, 1523–1527. [Google Scholar] [CrossRef] [PubMed]
- Yousefzadeh, G.; Larijani, B.; Mohammadirad, A.; Heshmat, R.; Dehghan, G.; Rahimi, R.; Abdollahi, M. Determination of oxidative stress status and concentration of TGF-β1 in the blood and saliva of osteoporotic subjects. Ann. N. Y. Acad. Sci. 2006, 1091, 142–150. [Google Scholar] [CrossRef]
- Östman, B.; Michaëlsson, K.; Helmersson, J.; Byberg, L.; Gedeborg, R.; Melhus, H.; Basu, S. Oxidative stress and bone mineral density in elderly men: Antioxidant activity of alpha-tocopherol. Free Radic. Biol. Med. 2009, 47, 668–673. [Google Scholar] [CrossRef]
- Cappetta, D.; De Angelis, A.; Sapio, L.; Prezioso, L.; Illiano, M.; Quaini, F.; Rossi, F.; Berrino, L.; Naviglio, S.; Urbanek, K. Oxidative stress and cellular response to doxorubicin: A common factor in the complex milieu of anthracycline cardiotoxicity. Oxid. Med. Cell. Longev. 2017, 2017, 1521020. [Google Scholar] [CrossRef]
- Olson, R.D.; Mushlin, P.S. Doxorubicin cardiotoxicity: Analysis of prevailing hypotheses. FASEB J. 1990, 4, 3076–3086. [Google Scholar] [CrossRef]
- De Beer, E.L.; Bottone, A.E.; Voest, E.E. Doxorubicin and mechanical performance of cardiac trabeculae after acute and chronic treatment: A review. Eur. J. Pharmacol. 2001, 415, 1–11. [Google Scholar] [CrossRef]
- Singal, P.; Li, T.; Kumar, D.; Danelisen, I.; Iliskovic, N. Adriamycin-induced heart failure: Mechanisms and modulation. Mol. Cell. Biochem. 2000, 207, 77–86. [Google Scholar] [CrossRef] [PubMed]
- Gianni, L.; Zweier, J.L.; Levy, A.; Myers, C.E. Characterization of the cycle of iron-mediated electron transfer from Adriamycin to molecular oxygen. J. Biol. Chem. 1985, 260, 6820–6826. [Google Scholar] [CrossRef] [PubMed]
- Sinha, B.K.; Politi, P.M. Anthracyclines. Cancer Chemother. Biol. Response Modif. 1990, 11, 45–57. [Google Scholar] [PubMed]
- Gutteridge, J.M.C.; Quinlan, G.J. Free radical damage to deoxyribose by anthracycline, aureolic acid and aminoquinone antitumour antibiotics: An essential requirement for iron, semiquinones and hydrogen peroxide. Biochem. Pharmacol. 1985, 34, 4099–4103. [Google Scholar] [CrossRef]
- Feinstein, E.; Canaani, E.; Weiner, L.M. Dependence of nucleic acid degradation on in situ free-radical production by adriamycin. Biochemistry 2002, 32, 13156–13161. [Google Scholar] [CrossRef]
- Huertas, J.R.; Battino, M.; Lenaz, G.; Mataix, F.J. Changes in mitochondrial and microsomal rat liver coenzyme Q9, and Q10 content induced by dietary fat and endogenous lipid peroxidation. FEBS Lett. 1991, 287, 89–92. [Google Scholar] [CrossRef]
- Huertas, J.R.; Battino, M.; Mataix, F.J.; Lenaz, G. Cytochrome oxidase induction after oxidative stress induced by adriamycin in liver of rats fed with dietary olive oil. Biochem. Biophys. Res. Commun. 1991, 181, 375–382. [Google Scholar] [CrossRef]
- Huertas, J.R.; Battino, M.; Barzanti, V.; Maranesi, M.; Parenti-Castelli, G.; Littarru, G.P.; Turchetto, E.; Mataix, F.J.; Lenaz, G. Mitochondrial and microsomal cholesterol mobilization after oxidative stress induced by adriamycin in rats fed with dietary olive and corn oil. Life Sci. 1992, 50, 2111–2118. [Google Scholar] [CrossRef]
- Mataix, J.; Mañas, M.; Quiles, J.; Battino, M.; Cassinello, M.; Lopez-Frias, M.; Huertas, J.R. Coenzyme Q content depends upon oxidative stress and dietary fat unsaturation. Mol. Aspects Med. 1997, 18, 129–135. [Google Scholar] [CrossRef]
- Quiles, J.L.; Ramirez-Tortosa, M.C.; Huertas, J.R.; Ibañez, S.; Gomez, J.A.; Battino, M.; Mataix, J. Olive oil supplemented with vitamin E affects mitochondrial coenzyme Q levels in liver of rats after an oxidative stress induced by adriamycin. BioFactors 1999, 9, 331–336. [Google Scholar] [CrossRef]
- Quiles, J.L.; Ramírez-Tortosa, M.C.; Ibéñez, S.; Gonzélez, J.A.; Duthie, G.G.; Fiuertas, J.R.; Mataix, J. Vitamin E supplementation increases the stability and the in vivo antioxidant capacity of refined olive oil. Free Radic Res. 1999, 31, 129–135. [Google Scholar] [CrossRef]
- Van Leeuwen, B.L.; Kamps, W.A.; Hartel, R.M.; Veth, R.P.H.; Sluiter, W.J.; Hoekstra, H.J. Effect of single chemotherapeutic agents on the growing skeleton of the rat. Ann. Oncol. 2000, 11, 1121–1126. [Google Scholar] [CrossRef] [PubMed]
- Friedlaender, G.E.; Tross, R.B.; Doganis, A.C.; Kirkwood, J.M.; Baron, R. Effects of chemotherapeutic agents on bone. I. Short-term methotrexate and doxorubicin (adriamycin) treatment in a rat model. J. Bone Jt. Surg.-Ser. A 1984, 66, 602–607. [Google Scholar] [CrossRef]
- Lleras-Forero, L.; Winkler, C.; Schulte-Merker, S. Zebrafish and medaka as models for biomedical research of bone diseases. Dev. Biol. 2019, 457, 191–205. [Google Scholar] [CrossRef]
- Laizé, V.; Gavaia, P.J.; Cancela, M.L. Fish: A suitable system to model human bone disorders and discover drugs with osteogenic or osteotoxic activities. Drug Discov. Today Dis. Model. 2014, 13, 29–37. [Google Scholar] [CrossRef]
- Izquierdo, M.S.; Scolamacchia, M.; Betancor, M.; Roo, J.; Caballero, M.J.; Terova, G.; Witten, P.E. Effects of dietary DHA and α-tocopherol on bone development, early mineralisation and oxidative stress in Sparus aurata (Linnaeus, 1758) larvae. Br. J. Nutr. 2013, 109, 1796–1805. [Google Scholar] [CrossRef]
- Saleh, R.; Betancor, M.B.; Roo, J.; Benítez-Santana, T.; Zamorano, M.J.; Izquierdo, M. Biomarkers of bone development and oxidative stress in gilthead sea bream larvae fed microdiets with several levels of polar lipids and α-tocopherol. Aquac. Nutr. 2015, 21, 341–354. [Google Scholar] [CrossRef]
- Winston, G.W.; Di Giulio, R.T. Prooxidant and antioxidant mechanisms in aquatic organisms. Aquat. Toxicol. 1991, 19, 137–161. [Google Scholar] [CrossRef]
- Tou, J.C. Resveratrol supplementation affects bone acquisition and osteoporosis: Pre-clinical evidence toward translational diet therapy. Biochim. Biophys. Acta-Mol. Basis Dis. 2014, 1852, 1186–1194. [Google Scholar] [CrossRef]
- Cottart, C.-H.; Nivet-Antoine, V.; Beaudeux, J.-L. Review of recent data on the metabolism, biological effects, and toxicity of resveratrol in humans. Mol. Nutr. Food Res. 2014, 58, 7–21. [Google Scholar] [CrossRef] [PubMed]
- Luo, Q.; Liu, S.; Xie, L.; Yu, Y.; Zhou, L.; Feng, Y.; Cai, D. Resveratrol ameliorates glucocorticoid-induced bone damage in a zebrafish model. Front. Pharmacol. 2019, 10, 195. [Google Scholar] [CrossRef] [PubMed]
- Giordo, R.; Nasrallah, G.K.; Al-Jamal, O.; Paliogiannis, P.; Pintus, G. Resveratrol inhibits oxidative stress and prevents mitochondrial damage induced by zinc oxide nanoparticles in zebrafish (Danio rerio). Int. J. Mol. Sci. 2020, 21, 3838. [Google Scholar] [CrossRef] [PubMed]
- Ran, G.; Ying, L.; Li, L.; Yan, Q.; Yi, W.; Ying, C.; Wu, H.; Ye, X. Resveratrol ameliorates diet-induced dysregulation of lipid metabolism in zebrafish (Danio rerio). PLoS ONE 2017, 12, e0180865. [Google Scholar] [CrossRef] [PubMed]
- Miura, S.; Saitoh, S.I.; Kokubun, T.; Owada, T.; Yamauchi, H.; Machii, H.; Takeishi, Y. Mitochondrial-targeted antioxidant maintains blood flow, mitochondrial function, and redox balance in old mice following prolonged limb ischemia. Int. J. Mol. Sci. 2017, 18, 1897. [Google Scholar] [CrossRef]
- Ni, R.; Cao, T.; Xiong, S.; Ma, J.; Fan, G.C.; Lacefield, J.C.; Lu, Y.; Tissier, S.L.; Peng, T. Therapeutic inhibition of mitochondrial reactive oxygen species with mito-TEMPO reduces diabetic cardiomyopathy. Free Radic. Biol. Med. 2016, 90, 12–23. [Google Scholar] [CrossRef]
- He, Q.; Harris, N.; Ren, J.; Han, X. Mitochondria-targeted antioxidant prevents cardiac dysfunction induced by tafazzin gene knockdown in cardiac myocytes. Oxid. Med. Cell. Longev. 2014, 2014, 654198. [Google Scholar] [CrossRef]
- Poudel, S.; Izquierdo, M.; Cancela, M.L.; Gavaia, P.J. Reversal of doxorubicin-induced bone loss and mineralization by supplementation of resveratrol and mitoTEMPO in the early development of Sparus Aurata. Nutr. 2022, 14, 1154. [Google Scholar] [CrossRef]
- Diogo, P.; Martins, G.; Quinzico, I.; Nogueira, R.; Gavaia, P.J.; Cabrita, E. Electric ultrafreezer (−150 °C) as an alternative for zebrafish sperm cryopreservation and storage. Fish Physiol. Biochem. 2018, 44, 1443–1455. [Google Scholar] [CrossRef]
- Martins, G.; Diogo, P.; Pinto, W.; Gavaia, P.J. Early transition to microdiets improves growth, reproductive performance and reduces skeletal anomalies in zebrafish (Danio rerio). Zebrafish 2019, 16, 300–307. [Google Scholar] [CrossRef]
- Poudel, S.; Izquierdo, M.; Cancela, M.L.; Gavaia, P.J. Reversal of Doxorubicin-induced bone loss by antioxidant supplement. Bone Rep. 2021, 14, 100984. [Google Scholar] [CrossRef]
- Peterman, E.M.; Sullivan, C.; Goody, M.F.; Rodriguez-Nunez, I.; Yoder, J.A.; Kim, C.H. Neutralization of mitochondrial superoxide by superoxide dismutase 2 promotes bacterial clearance and regulates phagocyte numbers in zebrafish. Infect. Immun. 2015, 83, 430–440. [Google Scholar] [CrossRef] [PubMed]
- Diogo, P.; Martins, G.; Gavaia, P.; Pinto, W.; Dias, J.; Cancela, L.; Martínez-Páramo, S. Assessment of nutritional supplementation in phospholipids on the reproductive performance of zebrafish, Danio rerio (Hamilton, 1822). J. Appl. Ichthyol. 2015, 31, 3–9. [Google Scholar] [CrossRef]
- Westerfield, M. The Zebrafish Book: A Guide for the Laboratory Use of Zebrafish Danio (“Brachydanio Rerio”); University of Oregon: Eugene, OR, USA, 2007. [Google Scholar]
- Martins, S.; Monteiro, J.F.; Vito, M.; Weintraub, D.; Almeida, J.; Certal, A.C. Toward an integrated zebrafish health management program supporting cancer and neuroscience research. Zebrafish 2016, 13 (Suppl. S1), S47–S55. [Google Scholar] [CrossRef] [PubMed]
- Gavaia, P.J.; Sarasquete, C.; Cancela, M.L. Detection of mineralized structures in early stages of development of marine Teleostei using a modified alcian blue-alizarin red double staining technique for bone and cartilage. Biotech. Histochem. 2000, 75, 79–84. [Google Scholar] [CrossRef] [PubMed]
- Walker, M.B.; Kimmel, C.B. A two-color acid-free cartilage and bone stain for zebrafish larvae. Biotech. Histochem. 2007, 82, 23–28. [Google Scholar] [CrossRef]
- Bird, N.C.; Mabee, P.M. Developmental morphology of the axial skeleton of the zebrafish, Danio rerio (Ostariophysi: Cyprinidae). Dev. Dyn. 2003, 228, 337–357. [Google Scholar] [CrossRef]
- Dominguez, D.; Sehnine, Z.; Castro, P.; Zamorano, M.J.; Robaina, L.; Fontanillas, R.; Antony Jesu Prabhu, P.; Izquierdo, M. Dietary manganese levels for gilthead sea bream (Sparus aurata) fingerlings fed diets high in plant ingredients. Aquaculture 2020, 529, 735614. [Google Scholar] [CrossRef]
- Xu, H.; Li, C.; Zeng, Q.; Agrawal, I.; Zhu, X.; Gong, Z. Genome-wide identification of suitable zebrafish Danio rerio reference genes for normalization of gene expression data by RT-qPCR. J. Fish Biol. 2016, 88, 2095–2110. [Google Scholar] [CrossRef]
- Viegas, M.N.; Dias, J.; Cancela, M.L.; Laizé, V. Polyunsaturated fatty acids regulate cell proliferation, extracellular matrix mineralization and gene expression in a gilthead seabream skeletal cell line. J. Appl. Ichthyol. 2012, 28, 427–432. [Google Scholar] [CrossRef]
- Cardiff, R.D.; Miller, C.H.; Munn, R.J. Manual hematoxylin and eosin staining of mouse tissue sections. Cold Spring Harb. Protoc. 2014, 2014, pdb.prot073411. [Google Scholar] [CrossRef]
- Fischer, A.H.; Jacobson, K.A.; Rose, J.; Zeller, R. Hematoxylin and eosin staining of tissue and cell sections. Cold Spring Harb. Protoc. 2008, 2008, pdb.prot4986. [Google Scholar] [CrossRef] [PubMed]
- Baeverfjord, G.; Krogdahl, A. Development and regression of soybean meal induced enteritis in Atlantic salmon, Salmo salar L., distal intestine: A comparison with the intestines of fasted fish. J. Fish Dis. 1996, 19, 375–387. [Google Scholar] [CrossRef]
- Magalhães, R.; Guerreiro, I.; Santos, R.A.; Coutinho, F.; Couto, A.; Serra, C.R.; Olsen, R.E.; Peres, H.; Oliva-Teles, A. Oxidative status and intestinal health of gilthead sea bream (Sparus aurata) juveniles fed diets with different ARA/EPA/DHA ratios. Sci. Rep. 2020, 10, 13824. [Google Scholar] [CrossRef]
- Pang, Z.; Chong, J.; Li, S.; Xia, J. MetaboAnalystR 3.0: Toward an optimized workflow for global metabolomics. Metabolism 2020, 10, 186. [Google Scholar] [CrossRef]
- Pang, Z.; Chong, J.; Zhou, G.; De Lima Morais, D.A.; Chang, L.; Barrette, M.; Gauthier, C.; Jacques, P.É.; Li, S.; Xia, J. MetaboAnalyst 5.0: Narrowing the gap between raw spectra and functional insights. Nucleic Acids Res. 2021, 49, W388–W396. [Google Scholar] [CrossRef] [PubMed]
- Bijlsma, S.; Bobeldijk, I.; Verheij, E.R.; Ramaker, R.; Kochhar, S.; Macdonald, I.A.; Van Ommen, B.; Smilde, A.K. Large-scale human metabolomics studies: A strategy for data (pre-) processing and validation. Anal. Chem. 2006, 78, 567–574. [Google Scholar] [CrossRef]
- Hadji, P.; Ziller, M.; Maskow, C.; Albert, U.; Kalder, M. The influence of chemotherapy on bone mineral density, quantitative ultrasonometry and bone turnover in pre-menopausal women with breast cancer. Eur. J. Cancer 2009, 45, 3205–3212. [Google Scholar] [CrossRef]
- Mathis, K.M.; Sturgeon, K.M.; Winkels, R.M.; Wiskemann, J.; De Souza, M.J.; Schmitz, K.H. Bone resorption and bone metastasis risk. Med. Hypotheses 2018, 118, 36–41. [Google Scholar] [CrossRef]
- Rana, T.; Chakrabarti, A.; Freeman, M.; Biswas, S. Doxorubicin-mediated bone loss in breast cancer bone metastases is driven by an interplay between oxidative stress and induction of TGFβ. PLoS ONE 2013, 8, e78043. [Google Scholar] [CrossRef]
- Yarmohammadi, F.; Rezaee, R.; Karimi, G. Natural compounds against doxorubicin-induced cardiotoxicity: A review on the involvement of Nrf2/ARE signaling pathway. Phyther. Res. 2021, 35, 1163–1175. [Google Scholar] [CrossRef]
- Pfeiffer, T.; Krause, U.; Thome, U.; Rajewski, A.; Skorzek, M.; Scheulen, M.E. Tissue toxicity of doxorubicin in first and second hyperthermic isolated limb perfusion—An experimental study in dogs. Eur. J. Surg. Oncol. 1997, 23, 439–444. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, M.A.; Ashour, O.M.; Ibrahim, Y.F.; EL-Bitar, H.I.; Gomaa, W.; Abdel-Rahim, S.R. Angiotensin-converting enzyme inhibition and angiotensin AT1-receptor antagonism equally improve doxorubicin-induced cardiotoxicity and nephrotoxicity. Pharmacol. Res. 2009, 60, 373–381. [Google Scholar] [CrossRef] [PubMed]
- Zakaria, Z.Z.; Benslimane, F.M.; Nasrallah, G.K.; Shurbaji, S.; Younes, N.N.; Mraiche, F.; Da’, S.I.; Yalcin, H.C. Using zebrafish for investigating the molecular mechanisms of drug-induced cardiotoxicity. BioMed Res. Int. 2018, 2018, 1642684. [Google Scholar] [CrossRef] [PubMed]
- Calienni, M.N.; Cagel, M.; Montanari, J.; Moretton, M.A.; Prieto, M.J.; Chiappetta, D.A.; del Alonso, S.V. Zebrafish (Danio rerio) model as an early stage screening tool to study the biodistribution and toxicity profile of doxorubicin-loaded mixed micelles. Toxicol. Appl. Pharmacol. 2018, 357, 106–114. [Google Scholar] [CrossRef]
- Chang, C.; Wu, S.L.; Zhao, X.D.; Zhao, C.T.; Li, Y.H. Developmental toxicity of doxorubicin hydrochloride in embryo-larval stages of zebrafish. Bio-Med. Mater. Eng. 2014, 24, 909–916. [Google Scholar] [CrossRef]
- Torno, C.; Staats, S.; de Pascual-Teresa, S.; Rimbach, G.; Schulz, C. Effects of resveratrol and genistein on growth, nutrient utilization and fatty acid composition of rainbow trout. Animal 2019, 13, 933–940. [Google Scholar] [CrossRef]
- Wilson, W.N.; Baumgarner, B.L.; Watanabe, W.O.; Alam, M.S.; Kinsey, S.T. Effects of resveratrol on growth and skeletal muscle physiology of juvenile southern flounder. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2015, 183, 27–35. [Google Scholar] [CrossRef]
- Yang, S.-G.; Park, H.-J.; Kim, J.-W.; Jung, J.-M.; Kim, M.-J.; Jegal, H.-G.; Kim, I.-S.; Kang, M.-J.; Wee, G.; Yang, H.-Y.; et al. Mito-TEMPO improves development competence by reducing superoxide in preimplantation porcine embryos. Sci. Rep. 2018, 8, 10130. [Google Scholar] [CrossRef]
- Valenzuela, C.A.; Escobar-Aguirre, S.; Zuloaga, R.; Vera-Tobar, T.; Mercado, L.; Björnsson, B.T.; Valdés, J.A.; Molina, A. Stocking density induces differential expression of immune-related genes in skeletal muscle and head kidney of fine flounder (Paralichthys adspersus). Vet. Immunol. Immunopathol. 2019, 210, 23–27. [Google Scholar] [CrossRef]
- Rigby, R.J.; Carr, J.; Orgel, K.; King, S.L.; Lund, P.K.; Dekaney, C.M. Intestinal bacteria are necessary for doxorubicin-induced intestinal damage but not for doxorubicin-induced apoptosis. Gut Microbe 2016, 7, 414–423. [Google Scholar] [CrossRef]
- Kaczmarek, A.; Brinkman, B.M.; Heyndrickx, L.; Vandenabeele, P.; Krysko, D.V. Severity of doxorubicin-induced small intestinal mucositis is regulated by the TLR-2 and TLR-9 pathways. J. Pathol. 2012, 226, 598–608. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.H.; Zhao, K.L.; Chen, X.L.; Xu, J.X. Impact of weaning and an antioxidant blend on intestinal barrier function and antioxidant status in pigs. J. Anim. Sci. 2012, 90, 2581–2589. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, N.; Ahmad, R.; Alam, M.A.; Ahmad, F.J. Enhancement of oral bioavailability of doxorubicin through surface modified biodegradable polymeric nanoparticles. Chem. Cent. J. 2018, 12, 65. [Google Scholar] [CrossRef] [PubMed]
- Ighwela, K.A.; Bin Ahmad, A.; Abol-Munafi, A.; Terengganu, M. Water stability and nutrient leaching of different levels of maltose formulated fish pellets. Glob. Vet. 2013, 10, 638–642. [Google Scholar] [CrossRef]
- Watts, S.A.; Powell, M.; D’Abramo, L.R. Fundamental approaches to the study of zebrafish nutrition. ILAR J. 2012, 53, 144–160. [Google Scholar] [CrossRef]
- Prestinicola, L.; Boglione, C.; Makridis, P.; Spanò, A.; Rimatori, V.; Palamara, E.; Scardi, M.; Cataudella, S. Environmental conditioning of skeletal anomalies typology and frequency in gilthead seabream (Sparus aurata L., 1758) Juveniles. PLoS ONE 2013, 8, e55736. [Google Scholar] [CrossRef]
- Koumoundouros, G.; Gagliardi, F.; Divanach, P.; Boglione, C.; Cataudella, S.; Kentouri, M. Normal and abnormal osteological development of caudal fin in Sparus aurata L. fry. Aquaculture 1997, 149, 215–226. [Google Scholar] [CrossRef]
- Koumoundouros, G.; Gagliardi, F.; Divanach, P.; Stefanakis, S.; Kentouri, M. Osteological study of the origin and development of the abnormal caudal fin in gilthead sea bream (Sparus aurata) fry. In Proceedings of the Quality in Aquaculture. Aquaculture Europe ’95 Int. Conference, Trondheim, Norway, 9–12 August 1995; pp. 16–18. [Google Scholar]
- Fazenda, C.; Martins, G.; Gavaia, P.J.; Cancela, M.L.; Conceição, N. Generation of zebrafish Danio rerio (Hamilton, 1822) transgenic lines overexpressing a heat-shock mediated Gla-rich protein. J. Appl. Ichthyol. 2018, 34, 472–480. [Google Scholar] [CrossRef]
- Fernández, I.; Hontoria, F.; Ortiz-Delgado, J.B.; Kotzamanis, Y.; Estévez, A.; Zambonino-Infante, J.L.; Gisbert, E. Larval performance and skeletal deformities in farmed gilthead sea bream (Sparus aurata) fed with graded levels of Vitamin A enriched rotifers (Brachionus plicatilis). Aquaculture 2008, 283, 102–115. [Google Scholar] [CrossRef]
- Boglione, C.; Pulcini, D.; Scardi, M.; Palamara, E.; Russo, T.; Cataudella, S. Skeletal anomaly monitoring in rainbow trout (Oncorhynchus mykiss, Walbaum 1792) reared under different conditions. PLoS ONE 2014, 9, e96983. [Google Scholar] [CrossRef]
- Divanach, P.; Boglione, C.; Menu, B.; Koumoundouros, G.; Kentouri, M.; Cataudella, S. Abnormalities in finfish mariculture: An overview of the problem, causes and solutions. In Seabass and Seabream Culture: Problems and Prospects; Chatain, B., Saroglia, M., Sweetman, J., Lavens, P., Eds.; EAS International Workshop: Verona, Italy; pp. 45–66.
- Klein-Nulend, J.; Bakker, A.D.; Bacabac, R.G.; Vatsa, A.; Weinbaum, S. Mechanosensation and transduction in osteocytes. Bone 2013, 54, 182–190. [Google Scholar] [CrossRef] [PubMed]
- Bensimon-Brito, A.; Cardeira, J.; Cancela, M.L.; Huysseune, A.; Witten, P.E. Distinct patterns of notochord mineralization in zebrafish coincide with the localization of Osteocalcin isoform 1 during early vertebral centra formation. BMC Dev. Biol. 2012, 12, 28. [Google Scholar] [CrossRef]
- Lall, S.P.; Kaushik, S.J. Nutrition and Metabolism of Minerals in Fish. Animals 2021, 11, 2711. [Google Scholar] [CrossRef]
- Berillis, P.; Panagiotopoulos, N.; Boursiaki, V.; Karapanagiotidis, I.T.; Mente, E. Vertebrae length and ultra-structure measurements of collagen fibrils and mineral content in the vertebrae of lordotic gilthead seabreams (Sparus aurata). Micron 2015, 75, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Lall, S.P. The Minerals. In Fish Nutrition, 3rd ed.; Halver, J.E., Hardy, R.W., Eds.; Academic Press: Cambridge, MA, USA, 2003; pp. 259–308. [Google Scholar] [CrossRef]
- Lall, S.P.; Lewis-McCrea, L.M. Role of nutrients in skeletal metabolism and pathology in fish—An overview. Aquaculture 2007, 267, 3–19. [Google Scholar] [CrossRef]
- Sun, M.; Wu, X.; Yu, Y.; Wang, L.; Xie, D.; Zhang, Z.; Chen, L.; Lu, A.; Zhang, G.; Li, F. Disorders of calcium and phosphorus metabolism and the proteomics/metabolomics-based research. Front. Cell Dev. Biol. 2020, 8, 576110. [Google Scholar] [CrossRef]
- Carvalho, F.S.; Burgeiro, A.; Garcia, R.; Moreno, A.J.; Carvalho, R.A.; Oliveira, P.J. Doxorubicin-induced cardiotoxicity: From bioenergetic failure and cell death to cardiomyopathy. Med. Res. Rev. 2014, 34, 106–135. [Google Scholar] [CrossRef]
- Filho, D.W.; Giulivi, C.; Boveris, A. Antioxidant defences in marine fish—I. Teleosts. Comp. Biochem. Physiol. Part C Pharmacol. Toxicol. Endocrinol. 1993, 106, 409–413. [Google Scholar] [CrossRef]
- Betancor, M.B.; Nordrum, S.; Atalah, E.; Caballero, M.J.; Benítez-Santana, T.; Roo, J.; Robaina, L.; Izquierdo, M. Potential of three new krill products for seabream larval production. Aquac. Res. 2012, 43, 395–406. [Google Scholar] [CrossRef]
- Saleh, R.; Betancor, M.B.; Roo, J.; Montero, D.; Zamorano, M.J.; Izquierdo, M. Selenium levels in early weaning diets for gilthead seabream larvae. Aquaculture 2014, 426–427, 256–263. [Google Scholar] [CrossRef]
- Gavaia, P.J.; Simes, D.C.; Ortiz-Delgado, J.B.; Viegas, C.S.B.; Pinto, J.P.; Kelsh, R.N.; Sarasquete, M.C.; Cancela, M.L. Osteocalcin and matrix Gla protein in zebrafish (Danio rerio) and Senegal sole (Solea senegalensis): Comparative gene and protein expression during larval development through adulthood. Gene Expr. Patterns 2006, 6, 637–652. [Google Scholar] [CrossRef]
- Carvalho, F.R.; Fernandes, A.R.; Cancela, M.L.; Gavaia, P.J. Improved regeneration and de novo bone formation in a diabetic zebrafish model treated with paricalcitol and cinacalcet. Wound Repair Regen. 2017, 25, 432–442. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Felber, K.; Elks, P.; Croucher, P.; Roehl, H.H. Tracking gene expression during zebrafish osteoblast differentiation. Dev. Dyn. 2009, 238, 459–466. [Google Scholar] [CrossRef] [PubMed]
- Boskey, A.L.; Gadaleta, S.; Gundberg, C.; Doty, S.B.; Ducy, P.; Karsenty, G. Fourier transform infrared microspectroscopic analysis of bones of osteocalcin-deficient mice provides insight into the function of osteocalcin. Bone 1998, 23, 187–196. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence | GenBank (Accession No.) | |
|---|---|---|---|
| oc2/bglapl | Fw: | CCAACTCCGCATCAGACTCCGCATCA | NM_001291889 |
| Rev: | AGCAACACTCCGCTTCAGCAGCACAT | ||
| sp7 | Fw: | GCTAAGTCCAGGGCAGGCTCAG | NM_212863 |
| Rev: | CAATGGCGTGAAATCAGGAGTGTAAC | ||
| runx2b | Fw: | TCAGGAATGCCTCAGGGGTTATG | NM_212862 |
| Rev: | CTTGCGGTGGGTTTGTGAATACT | ||
| eef1a1l1 | Fw: | TTGAGAAGAAAATCGGTGGTGCTG | NM_131263 |
| Rev: | GGAACGGTGTGATTGAGGGAAATTC | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Poudel, S.; Martins, G.; Cancela, M.L.; Gavaia, P.J. Regular Supplementation with Antioxidants Rescues Doxorubicin-Induced Bone Deformities and Mineralization Delay in Zebrafish. Nutrients 2022, 14, 4959. https://doi.org/10.3390/nu14234959
Poudel S, Martins G, Cancela ML, Gavaia PJ. Regular Supplementation with Antioxidants Rescues Doxorubicin-Induced Bone Deformities and Mineralization Delay in Zebrafish. Nutrients. 2022; 14(23):4959. https://doi.org/10.3390/nu14234959
Chicago/Turabian StylePoudel, Sunil, Gil Martins, M. Leonor Cancela, and Paulo J. Gavaia. 2022. "Regular Supplementation with Antioxidants Rescues Doxorubicin-Induced Bone Deformities and Mineralization Delay in Zebrafish" Nutrients 14, no. 23: 4959. https://doi.org/10.3390/nu14234959
APA StylePoudel, S., Martins, G., Cancela, M. L., & Gavaia, P. J. (2022). Regular Supplementation with Antioxidants Rescues Doxorubicin-Induced Bone Deformities and Mineralization Delay in Zebrafish. Nutrients, 14(23), 4959. https://doi.org/10.3390/nu14234959