1. Introduction
Anthocyanins and other flavonoids extracted from plum, peach, raspberry, and other sources such as vegetables, grains, tea have been identified to exhibit bioactive property [
1,
2,
3,
4,
5]. This bioactive property has been demonstrated to provide beneficial effect on neuroprotection by reducing reactive oxygen species (ROS), increasing antioxidant enzyme activities [
6] and cardio-health promotion [
7]. However, literature reports that flavonoids, including anthocyanins, have poor bioavailability [
8,
9,
10,
11]. This low bioavailability leads to reduced health effect of anthocyanins when consumed in our daily life due to their low concentration in plasma or functional site in our body. Therefore, bioavailability is a key part for a bioactive compound to engender its beneficial effect.
Understanding factors that affect anthocyanins uptake into cells is an integral part of understanding their bioavailability. There is little research focusing on the uptake of anthocyanins into cells [
11,
12,
13] and even less detailing the factors affecting their uptake.
An investigation into the effect of glucose on anthocyanin absorption using animals provides some evidence for the role of glucose transporters in anthocyanin absorption [
14]. Reports on the involvement of glucose transporter pathway in anthocyanin absorption through intestinal cell have been documented elsewhere with conflicting conclusions [
15,
16].
Various anthocyanins with different structures are present in fruits and vegetables. Furthermore, anthocyanin stability is known to depend on pH [
17]. Therefore, apart from glucose, other factors such as pH and anthocyanin structure are worth further investigation. The aim of this study was to examine factors likely to affect the uptake into cells (Caco-2 cells) including anthocyanin structure, presence of glucose, and pH. In the present study, anthocyanin extract from strawberry and red grape was used because these fruits contain different structure of anthocyanins such as cyanidine-3 glucoside, pelargonidin-3 glucoside and malvidin-3 glucoside [
8] so the effect of anthocyanins structure can be revealed.
Caco-2 cells were chosen because they have been successfully used as an in vitro model to study the absorption or uptake of drugs and flavonoids [
12,
18,
19]. In culture Caco-2 cells are subject to spontaneous enterocytic differentiation and polarization and can be used as a model system for the small intestinal epithelium [
20,
21].
2. Materials and Methods
2.1. Chemical and Reagents
Acetonitrile, methanol, formic acid, hydrochloric acid, water (HPLC grade), MK-571, Amberlite XAD7-HP, anthocyanin standards (pelargonidin-3 glucoside, pelargonidin-3-rhamnosylglucoside, cyanidin-3-glucoside, delphinidin-3- glucoside, malvidin-3-glucoside, petunidin-3-glucoside, peonidin-3-glucoside), 4’,6-diamidino-2-phenylindole (DAPI) were purchased from Sigma-Aldrich, Gillingham, UK. Caco-2 cells were purchased from European Collection of Authenticated Cell Cultures (ECACC), Salisbury, UK. Concentrate of Strawberry was purchased from Cobell, Exeter, UK and red grape concentrate from Homebrew shop, Aldershot, UK. The other reagents were from Lonza, Basle, Switzerland.
2.2. Extraction
Strawberry or red grape concentrate (40 g or 50 g, respectively) was mixed into 100 mL of 1% aqueous formic acid followed by removing the solid by vacuum filtration with Whatman no 4 filter paper. The filtrate was extracted with ethyl acetate then loaded onto an amberlite XAD7-HP column 60 cm long, 2 cm diameter which had been washed with water followed by methanol to remove monomers trapped on the resin. Acidified water was subsequently passed through the resin to remove glucose and other polar compounds until no glucose was detected in the washing water. Finally, the anthocyanins were eluted with acidified methanol, followed by removal of solvent with a rotary evaporator at about 40 °C. This solution of anthocyanin extract was then stored at −80 °C for later experiments of cell viability and uptake.
2.3. Cell Cultures
Caco-2 cells were seeded at a density of 5 × 105 cells in a Petri dish of 10 cm diameter with a volume of 10 mL, containing DMEM supplemented with 10% FBS, 1% NEAA and 1% penicillin and streptomycin. The cells were grown in an incubator with 95% oxygen, 5% carbon dioxide, 95% humidity at 37 °C. Caco-2 cells were used between passages 65–76. The medium was changed every 2–3 days. On day 14, the cells were ready for the uptake experiment. Prior to the study, the medium were removed, and the cells were rinsed with PBS.
2.4. Cell Viability
Prior to the uptake experiment, cytotoxicity effect of anthocyanins onto Caco-2 cells at different concentrations, pH and length of exposure was examined. This is important to ensure that the uptake experiment is carried out at the similar level of cell viability. Therefore, any responses of the uptake experiment results from either difference in pH, glucose or anthocyanin structure. To verify the effect of anthocyanin cytotoxicity, first of all, the cell suspension was seeded onto the inner wells of 96 well plates at a density of 106 cells/mL with the volume of 50 µL, while medium not containing cells was added to the outer wells. The cells were then incubated for 24 h in an incubator at 37 °C, under 5% CO2, 95% O2 and 95% humidity. Following the incubation, the medium was removed from all wells in the plate, and subsequently 50 µL of anthocyanin solutions (extracts from red grape and strawberry) ranging from 0 to 50 µM in DPBS was added into the wells, and then incubated for either 1 or 2 h. Each anthocyanin concentration was added into 4 wells per plate (3 with and 1 without cells). After the incubation, DPBS was removed, and chilled methanol was added into the wells. The methanol was then removed and 100 µL of DAPI solution was added followed by incubation for 30 min at 37 °C prior to measurement in a microplate reader at excitation and emission wavelengths of 340 and 465 nm, respectively. The experiment was repeated triplicate.
2.5. Uptake Experiment
The effect of anthocyanin structure was examined using anthocyanin extract from strawberry at 10 and 20 µM while the effect of glucose was performed at 20 µM red grape anthocyanin extract in the presence of 1 mM glucose. Additionally, 10 µM strawberry anthocyanins at pH 6.5 and 7 was used to examine the effect of pH on anthocyanin uptake.
Briefly anthocyanin stock solution was diluted to a final concentration of 10 or 20 µM in DPBS. The anthocyanins (10 mL) were immediately added to the cells, and subsequently incubated at 37 °C for 1h. Following the incubation, DPBS containing anthocyanins was removed and the cells were washed with PBS three times. The anthocyanins taken up by the cells were extracted with 6% formic acid and scraped off for collection. The extract along with the scraped cells were then sonicated, followed by centrifugation at 4 °C, 1700× g for 10 min. The supernatant was filtered and dried under nitrogen. The dry anthocyanins were redissolved in 200 µL of aqueous formic acid containing 10% acidified methanol, and then filtered into the insert prior to HPLC analysis.
2.6. HPLC Analysis
HPLC was used to identify and quantify anthocyanins present in juices and cellular extract following the uptake experiment. Anthocyanin standards and their standard curves were used to identify and quantify different anthocyanin structure present in juices and cellular extracts by determining their area under curve. The analysis was performed using an Agilent 1200 series HPLC with quaternary pump and diode array UV-visible detector. Sample (50 µL) was injected onto a reverse phase column of Nova-pak@C18 4 µm, with dimensions of 4.6 × 250 mm (Waters) equipped with a guard column. The mobile phases were 1% aqueous formic acid as solvent A and acetonitrile as solvent B. The flow rate was 1 mL/min with a column temperature of 35 °C. Compounds were eluted with a gradient of 100% solvent A at 0 min, changing to 81% A and 19% B at 19 min, 68% A and 32% B at 45 min, then 0% A and 100% B at 52 min. Solvent B was maintained at 100% for 5 min, and returned to 100% solvent A and 0% solvent B at 60 min. Wavelengths of 280 and 520 nm were chosen to detect the anthocyanins.
2.7. LC-MS Analysis
Several peaks detected in HPLC analysis either in juice or cellular extracts were not able to be identified due to the lack of anthocyanin standards; therefore, LC-MS was applied to identify the compounds. The identification of peaks using LC-MS particularly was conducted to main peaks present in juice (such as in red grape anthocyanins) or new peaks present in cellular extract which might be present as metabolites. Analysis by LC-MS was carried out with the same column and mobile phase as in the HPLC analysis. The LC-MS system consisted of an Agilent 1100 HPLC and a Bruker MicrotofQ II high resolution time of flight (TOF) mass spectrometer. The analysis was in the positive ion mode at 180 °C with electrospray ionisation. Nebulizer pressure at 1 bar N2, with flow rate of N2 gas 8 L/min were used. The collision voltage was 10 eV, collision RF 210 Vpp, capillary voltage −4000 V. Data were collected in high resolution mode.
2.8. Statistical Analysis
Data were analysed with the PASW statistic 17 package (SPSS Inc., Chicago, IL, USA). Analysis of variance, followed by LCD was used to examine the effect of anthocyanin concentration on cell viability and the effect of anthocyanin structure on uptake. Other data were analysed by Student’s t-test. Differences were considered as significant where p < 0.05.
4. Discussion
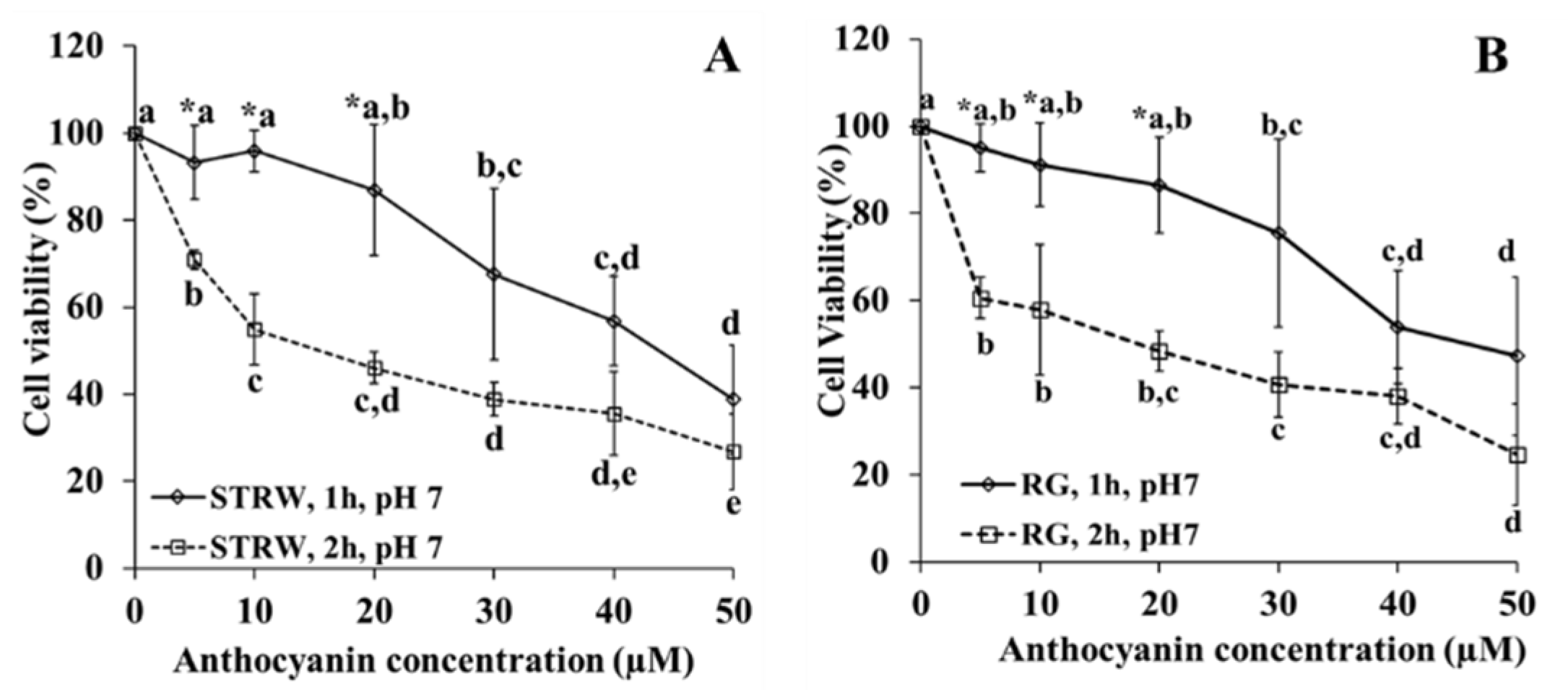
The anthocyanin addition onto Caco-2 cells may affect the cells viability due to the cytotoxicity effect. Therefore, it is pivotal to verify whether anthocyanins lead to a cytotoxicity effect. This information is also important to determine the anthocyanin concentration and the choice of pH for the anthocyanin uptake experiment. Ideally the chosen anthocyanin concentration is determined based on the highest anthocyanin concentration giving the highest cell viability.
The cytotoxicity experiment expressed in percentage of cell viability showed that anthocyanins at studied concentrations were toxic to Caco-2 cells. This finding was also demonstrated in another study on cytotoxicity of black carrot anthocyanin extract [
22]. The toxicity was dependent on anthocyanin concentration and length of exposure. Comparing the effect of length exposure of 1 h to 2 h it is obvious that restriction of cell exposure to 1 h allowed concentrations up to 20 µM to result in similar cell viability for both the strawberry and red grape anthocyanins extracts. This concentration was the highest concentration with the highest and similar cell viability. Consequently, for the experiment of structure effect, concentrations of 10 and 20 µM were applied. Meanwhile for the experiment of glucose effect and the uptake of anthocyanins from strawberry and red grape, the highest concentration of 20 µM was used.
In terms of pH effect on cytotoxicity, the pH from 6–7 showed no different toxicity at corresponding anthocyanin concentration. To minimize the effects of cytotoxicity, the concentration of 10 µM at pH 7 and 6.5 was chosen for the experiment of the effect of pH on the anthocyanin uptake. At this concentration and pH, the cells had the highest and similar viability.
Analyzing anthocyanins from strawberry and red grape juice using HPLC it was found that the main anthocyanins present in strawberry were cyanidin-3-glucoside, pelargonidin-3-glucoside and pelargonidin-3-rhamnosylglucoside. These anthocyanins constitute approximately 90% of anthocyanins present in strawberry juice. Meanwhile red grape contains delphinidin-3-glucoside, cyanidin-3-glucoside, petunidin-3-glucoside, peonidin-3-glucoside, and malvidin-3-glucoside identified as the main anthocyanin (39.76%). Two main peaks in red grape juice was not identifiable by HPLC due to the lack of the anthocyanin standards. Therefore, LC-MS was applied. The two peaks exhibited m/z values of 531/369 and 561/399. These compounds were also glycosides, as indicated by the loss of 162 Da following fragmentation in the mass spectrum. These two compounds had the molecular formula C
25H
23O
13 and C
26H
25O
14, respectively suggesting that the compounds were peonidin-3-glucoside pyruvic acid and malvidin-3-glucoside pyruvic acid, respectively, also known as vitisin A compounds. Vitisin A along with vitisin B have been commonly found in red wine as a fermentation product. Although vitisin B has been previously found in grape juice [
23]. In this study, we have also found vitisin A in red grape juice.
The uptake study using strawberry and red grape anthocyanin extracts demonstrated that anthocyanins are taken up by cells. Total uptake of red grape anthocyanins was smaller than that of strawberry (0.015% versus 0.073%) suggesting that uptake of anthocyanins in red grape are less favourable than those in strawberry. This finding may indicate the effect of anthocyanin structure on the extent of the uptake into cells. Several new peaks including those labelled as peaks 5, a, b, and c, corresponding to new compounds appeared in the cell extract. The literature includes conflicting reports about whether flavonoids, such as anthocyanins can be metabolised by Caco-2 cells, with metabolites found in one study [
24] but not in a second study [
25]. Anthocyanins are known to degrade rapidly at pH 7, so the possibility of the new peaks arising from chemical reaction during 1 h incubation was considered. In our case, peaks 5, a, b, and c were identified using LC-MS.
To assess degradation due to the pH, strawberry juice extract was incubated under the same conditions in the absence of the cells (
Figure 3II). The appearance of peak 5 within 1 h incubation in the absence of cells suggested that the compound corresponding to peak 5 was formed by chemical reaction such as oxidation, cleavage and ring opening or reaction with other compounds during the incubation and was not a metabolite formed by the Caco-2 cells. Anthocyanins can form additional compounds important for copigmentation with phenolic acids [
26] and can form pyranoanthocyanins [
27].
The fact that peaks a, b, and c were not detected in the absence of the cells does not necessarily mean that these compounds are metabolites formed by the cells; it is possible that these compounds were present in small quantities in the cell extract that were not detected prior to incubation. In this experiment, the cell extract was concentrated to a great extent to boost the sensitivity of detection due to the low concentration of the compounds of interest, while in the absence of the cells, concentration of the sample was not carried out owing to the relatively high concentration of the compounds already present in the solution. Consequently, small concentrations of a component formed in the absence of the cells might not be detected.
Peaks a, b and c had molecular masses of 442, 486 and 316 Da with formulae C19H38O11, C21H42O12 and C20H30NO2, respectively. These masses did not correspond to expected metabolites of anthocyanins formed by glucuronidation, sulfoconjugation or methylation, leading to a conclusion that these peaks were not metabolites. The presence of a nitrogen atom in the molecular formula of peak c was an indication that this was not an anthocyanin derivative but probably an amino acid or inorganic nitrate already present in the juice.
The experiment testing different concentrations of strawberry anthocyanins showed that the percentage increase in uptake of anthocyanins into the cells increased of anthocyanin concentration (
Table 1); demonstrating that the uptake was concentration-dependent and hence examining the effect of anthocyanin structure on the uptake should be estimated at the same concentration for each anthocyanin structure.
Amongst the anthocyanins present in the strawberry extract, cyanidin-3 glucoside was the most taken up by cells at the same initial amount of anthocyanin (10 µg/10 mL). The addition of rhamnosyl–glucoside to pelargonidin seems to favourably increase the uptake compared to the addition of glucoside. This finding demonstrates that anthocyanins structure affects the extent of anthocyanin uptake. Our finding is in line with that of another study highlighting the importance of anthocyanin structure in the uptake [
11] which also found that cyanidin-3-glucoside was more taken up by cells than pelargonidin-3-glucoside. The fact that cyanidin-3-glucoside which is more polar than pelargonidin-3-glucoside was taken up more highly suggests that the uptake might not occur through the cell membrane by passive diffusion, as the cell membrane is hydrophobic in nature; the uptake might take place through facilitative transporter. The higher uptake of the more polar compound was also demonstrated in the higher uptake of total anthocyanins from strawberry than from red grape. The former predominantly contained pelargonidin-3 glucoside and malvidin-3 glucoside, respectively. These findings from this in vitro study is in agreement with our human intervention study where more anthocyanins excretion from strawberry was found in urine than from red grape [
8].
The presence of 1mM glucose decreased the uptake of total red grape anthocyanins by 62%. The decrease in uptake in the presence of glucose indicates potential competition between glucose and anthocyanins for uptake into cells through the same transporter channels as previously described [
28]. Our data indicate the possibility that the uptake of anthocyanins occur through apical membrane cells involving the glucose transporter pathway. This finding is in line with previous work demonstrating that absorption of anthocyanins through Caco-2 cells involves facilitative glucose transporters2 (GLUT2) [
15] and also aligns with a human intervention study [
29]. This concept is akin to the suggested uptake mechanism for flavonol glycosides [
30,
31]. This suggests the possibility of competition with other flavonoids in the uptake of anthocyanins when other flavonoids are present, particularly when the glucose transporter is saturated with these compounds.
The uptake competition amongst anthocyanins may be indicated from the different level of anthocyanin sensitivity to the glucose effect. In the absence of glucose, the more polar anthocyanins were more favourably taken up by cells. In other words, the high sensitivity of malvidin-3 glucoside to glucose compared to cyanidin and delphinidin-3 glucoside may relate to its lower uptake due to the less polar nature of malvidin.
The study investigating the impact of pH shows that pH of milieu affects the extent of anthocyanin uptake. The increased uptake with decreasing pH may be linked to the anthocyanins stability and indicate the involvement of pH dependent transport mechanisms involving a proton co-transporter. Observation of anthocyanins during a 1 h incubation indicated that stability of cyanidin-3 glucoside, pelargonidin-3 glucoside and pelargonidin-3 rhamnocylglucoside at pH 7 was 63.1, 67.7 and 72.3%, respectively and it increased to 88.7, 93.7 and 91.8% at lower pH of 6.5.
The small intestine consists of 3 sections namely the duodenum, jejunum and ileum in which the pH has been shown to progressively increase from about 5.5–6 in the duodenum to about 7–7.5 in the terminal ileum [
32,
33]. Therefore, anthocyanin absorption through the apical membrane could be much better in the upper part of the intestinal tract including the duodenum and jejunum than in the ileum.
The acidic pH studied in this experiment was closer to the environment of the jejunum. Considering the length of the duodenum which is only 5% of the length of the small intestine, it is reasonable to speculate that the jejunum, which is much longer than the duodenum (almost 50% of small intestinal length), is a more favourable site for anthocyanin uptake. Using our data for total anthocyanin at pH 6.5, uptake reached 479 pg/cm
2 using 10 µM concentration of anthocyanins; the uptake of anthocyanins through the jejunum with surface area of 184 m
2 with length of 280 cm [
32] might reach as high as 0.88 mg in 1 h. In vivo trials have shown that the maximum concentration of anthocyanins in plasma is reached after about 1 h post ingestion, so uptake in 1 h is relevant. Absorption of 0.88 mg pelargonidin glucoside would lead to a blood concentration of 400 nM, or 727 nM in plasma, assuming a volume of 5 litres of blood. In an intervention study, the main metabolite, pelargonidin-O-glucuronide, reached a peak plasma concentration of 274 +/−24 nM after 1.1 +/−0.4 h when strawberries containing 222 µmol of pelargonidin-3-O-glucoside were ingested [
34], so the calculation leads to concentrations of the right order of magnitude.










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}