

The Effect of the Restrictive Ketogenic Diet on the Body Composition, Haematological and Biochemical Parameters, Oxidative Stress and Advanced Glycation End-Products in Young Wistar Rats with Diet-Induced Obesity



Abstract
1. Introduction
2. Materials and Methods
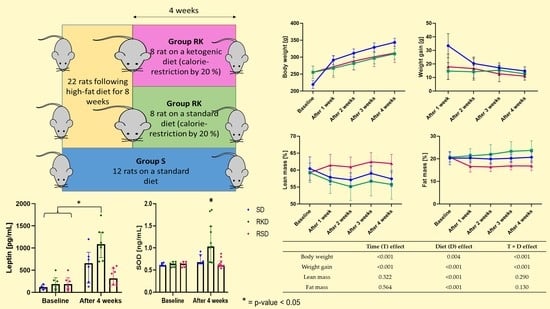
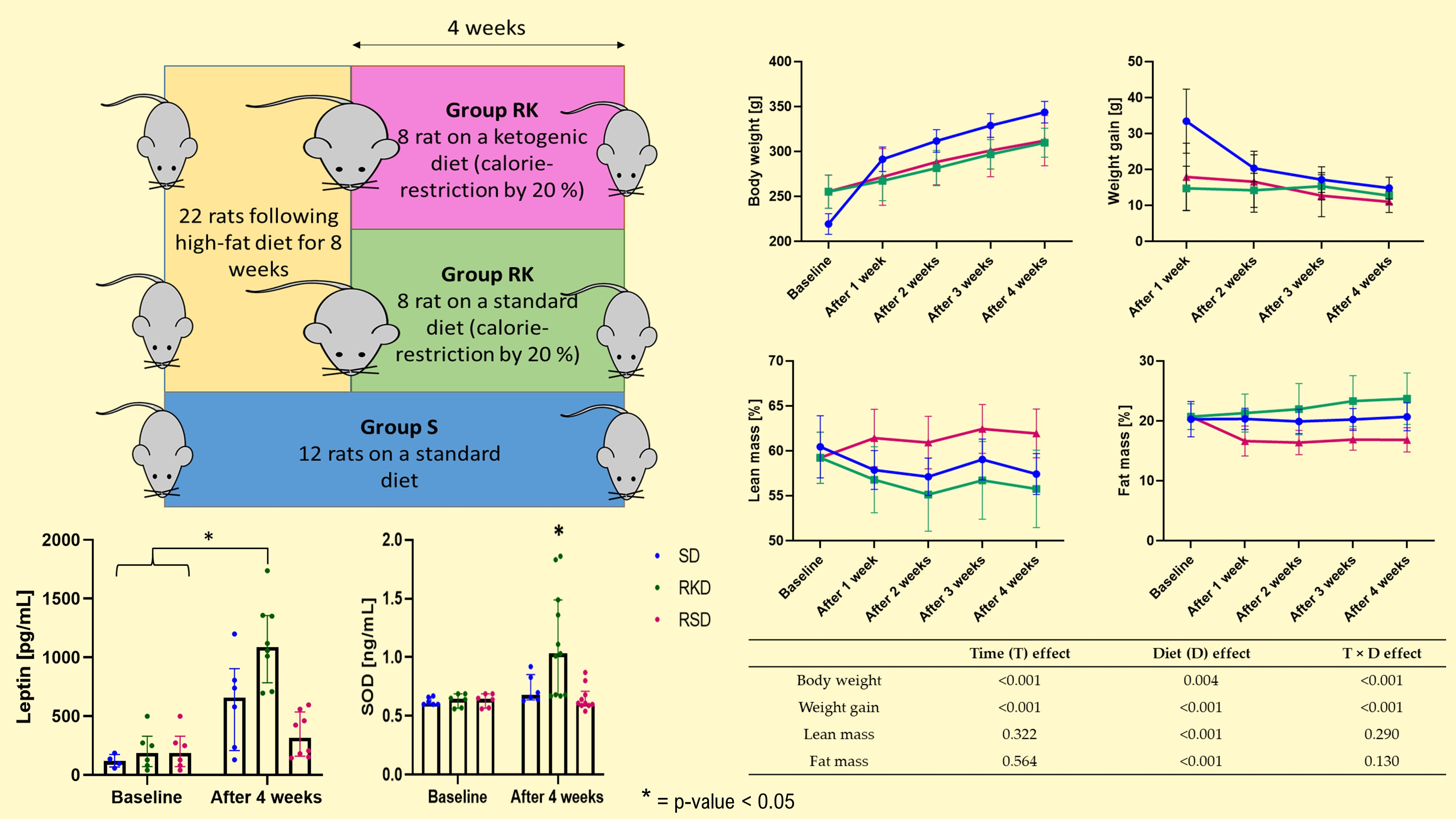
2.1. Animals and Diets
2.2. Sample Collection
2.3. Biochemical and Haematological Analyses
2.4. Oxidative Stress Markers
2.5. Statistical Analyses
3. Results
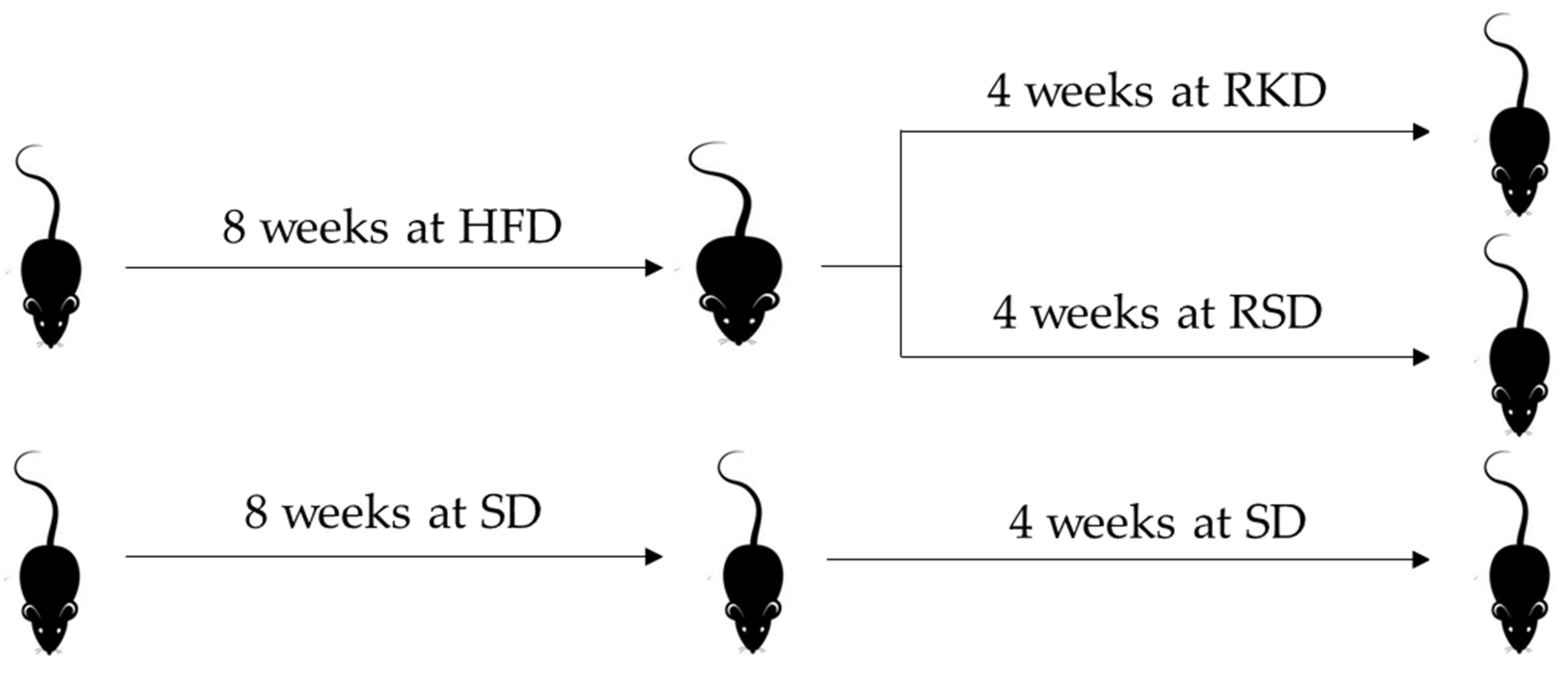
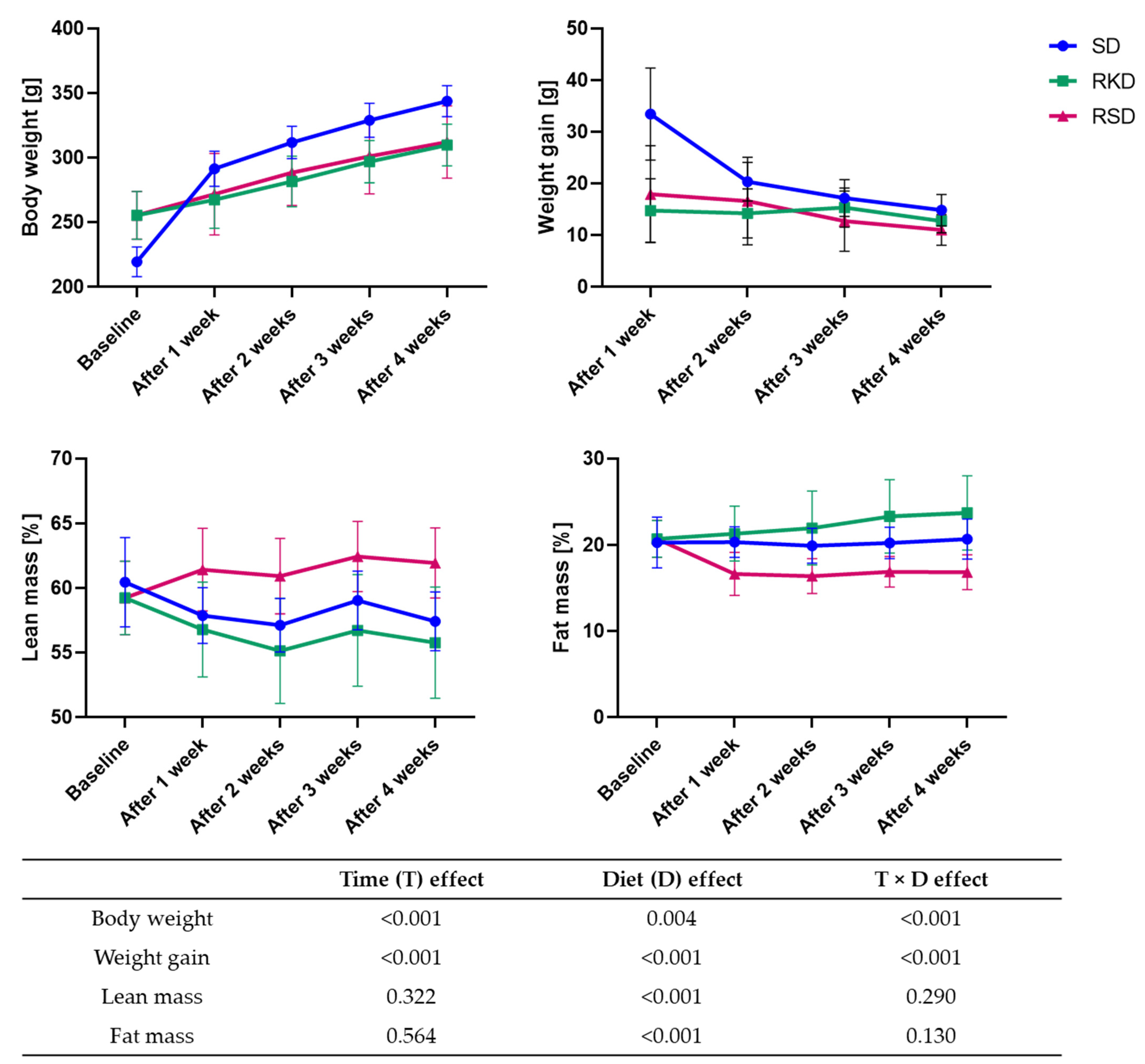
3.1. The Effect of the Ketogenic Diet on Body Weight and Composition
3.2. The Effect of the Ketogenic Diet on Blood Haematology
3.3. The Effect of KD on the Biochemical Parameters
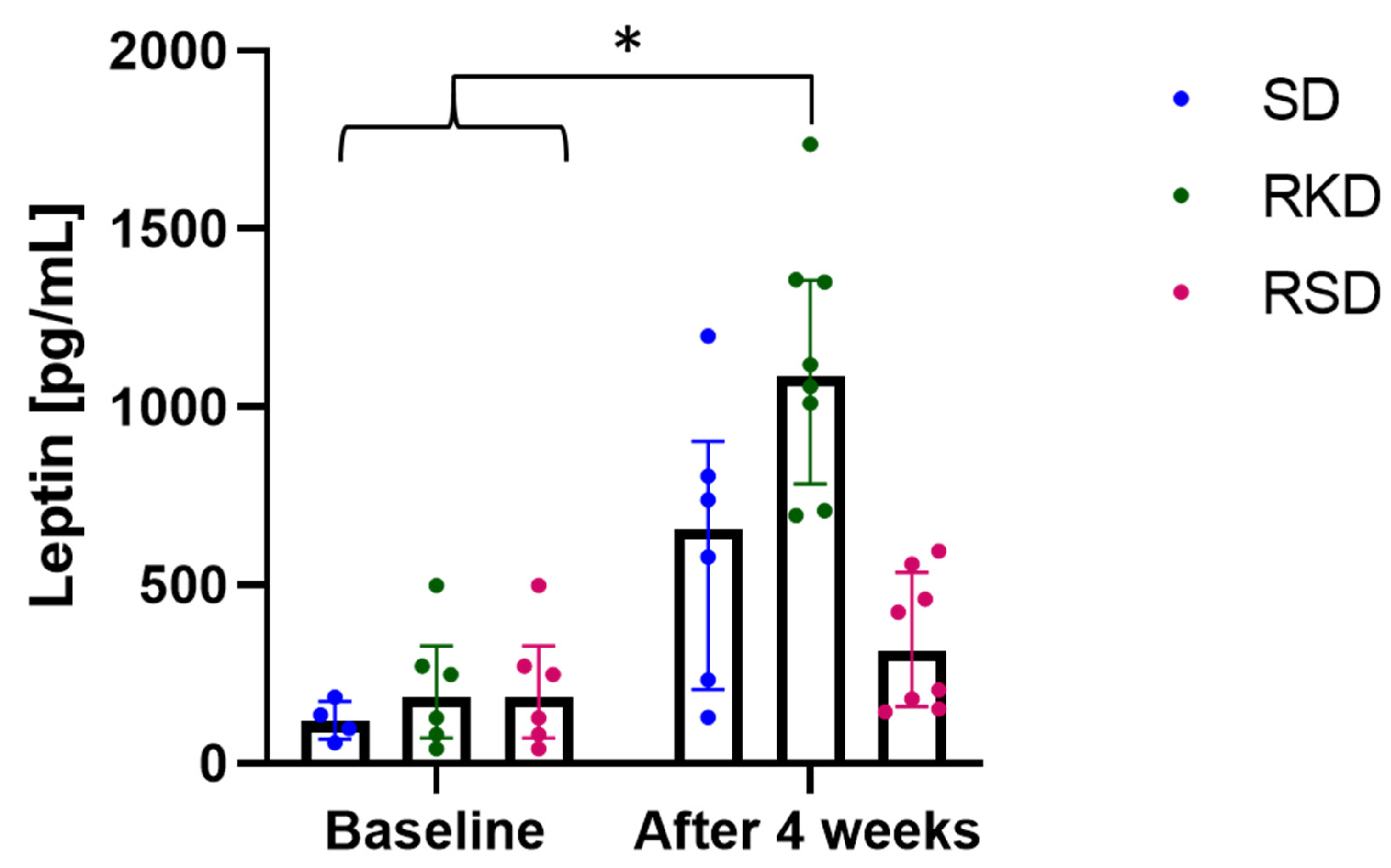
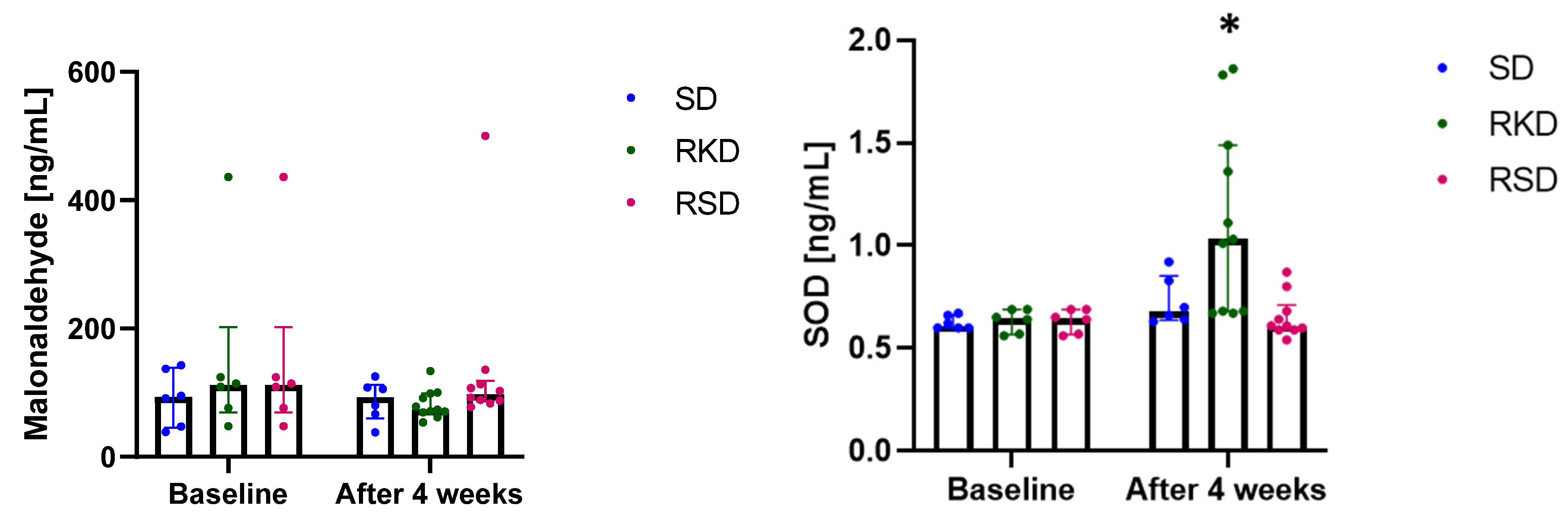
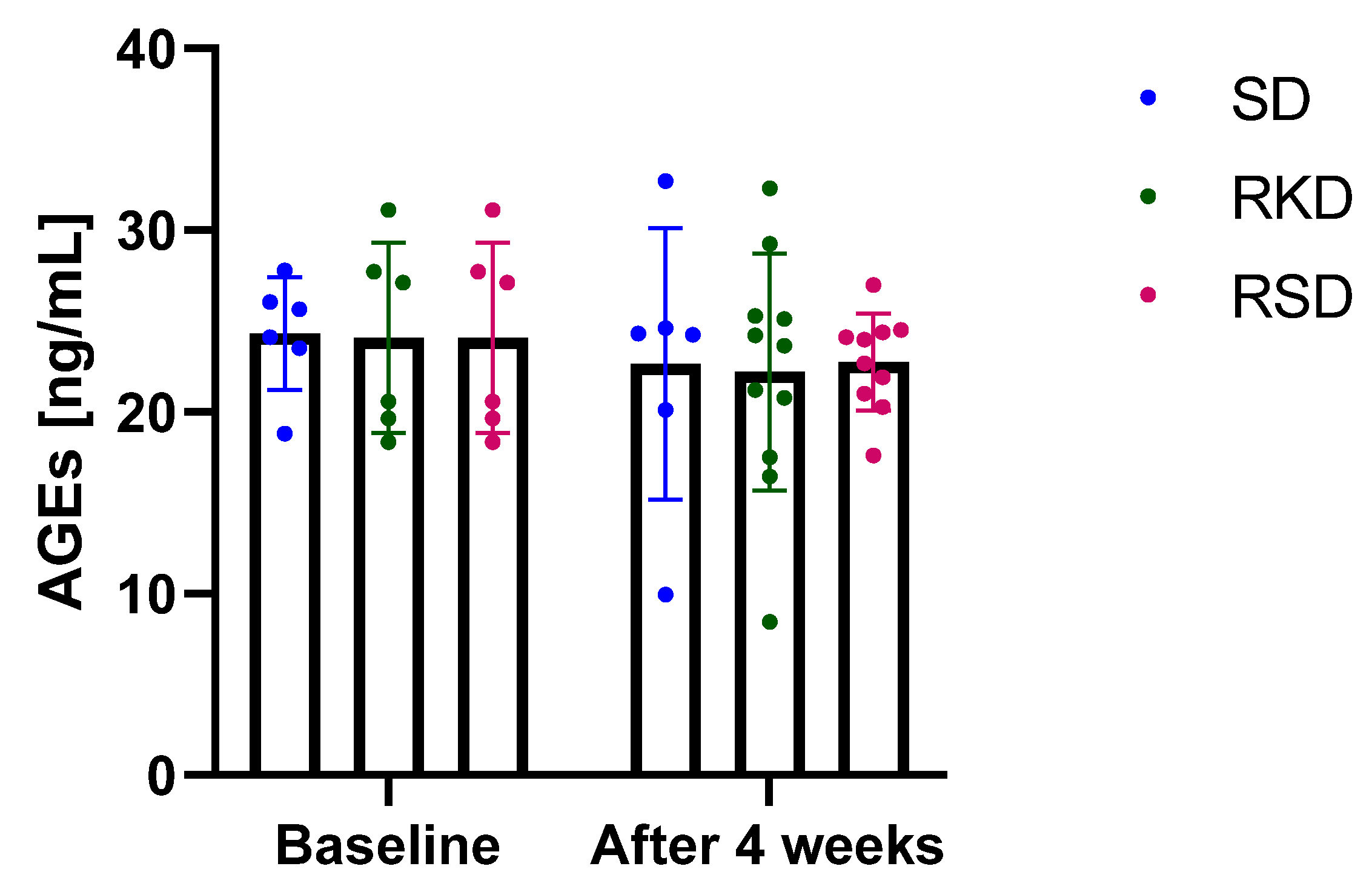
3.4. The Effect of the Ketogenic Diet on the Oxidative Status
3.5. Correlations between Body Composition and Biochemical Parameters
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Williams, E.P.; Mesidor, M.; Winters, K.; Dubbert, P.M.; Wyatt, S.B. Overweight and Obesity: Prevalence, Consequences, and Causes of a Growing Public Health Problem. Curr. Obes. Rep. 2015, 4, 363–370. [Google Scholar] [CrossRef] [PubMed]
- Graßmann, S.; Wirsching, J.; Eichelmann, F.; Aleksandrova, K. Association Between Peripheral Adipokines and Inflammation Markers: A Systematic Review and Meta-Analysis. Obesity 2017, 25, 1776–1785. [Google Scholar] [CrossRef] [PubMed]
- Marseglia, L.; Manti, S.; D’Angelo, G.; Nicotera, A.; Parisi, E.; di Rosa, G.; Gitto, E.; Arrigo, T. Oxidative Stress in Obesity: A Critical Component in Human Diseases. Int. J. Mol. Sci. 2014, 16, 378–400. [Google Scholar] [CrossRef] [PubMed]
- Sohouli, M.H.; Sharifi-Zahabi, E.; Lari, A.; Fatahi, S.; Shidfar, F. The Impact of Low Advanced Glycation End Products Diet on Obesity and Related Hormones: A Systematic Review and Meta-Analysis. Sci. Rep. 2020, 10, 22194. [Google Scholar] [CrossRef] [PubMed]
- Uribarri, J.; Cai, W.; Woodward, M.; Tripp, E.; Goldberg, L.; Pyzik, R.; Yee, K.; Tansman, L.; Chen, X.; Mani, V.; et al. Elevated Serum Advanced Glycation Endproducts in Obese Indicate Risk for the Metabolic Syndrome: A Link Between Healthy and Unhealthy Obesity? J. Clin. Endocrinol. Metab. 2015, 100, 1957–1966. [Google Scholar] [CrossRef] [PubMed]
- Khalid, M.; Petroianu, G.; Adem, A. Advanced Glycation End Products and Diabetes Mellitus: Mechanisms and Perspectives. Biomolecules 2022, 12, 542. [Google Scholar] [CrossRef]
- Ribeiro, P.V.M.; Tavares, J.F.; Costa, M.A.C.; Mattar, J.B.; Alfenas, R.C.G. Effect of Reducing Dietary Advanced Glycation End Products on Obesity-Associated Complications: A Systematic Review. Nutr. Rev. 2019, 77, 725–734. [Google Scholar] [CrossRef]
- Ott, C.; Jacobs, K.; Haucke, E.; Navarrete Santos, A.; Grune, T.; Simm, A. Role of Advanced Glycation End Products in Cellular Signaling. Redox Biol. 2014, 2, 411–429. [Google Scholar] [CrossRef]
- Drabińska, N.; Wiczkowski, W.; Piskuła, M.K. Recent Advances in the Application of a Ketogenic Diet for Obesity Management. Trends Food Sci. Technol. 2021, 110, 28–38. [Google Scholar] [CrossRef]
- Dhamija, R.; Eckert, S.; Wirrell, E. Ketogenic Diet. Can. J. Neurol. Sci./J. Can. Des Sci. Neurol. 2013, 40, 158–167. [Google Scholar] [CrossRef]
- Ułamek-Kozioł, M.; Czuczwar, S.J.; Januszewski, S.; Pluta, R. Ketogenic Diet and Epilepsy. Nutrients 2019, 11, 2510. [Google Scholar] [CrossRef] [PubMed]
- Fukao, T.; Lopaschuk, G.D.; Mitchell, G.A. Pathways and Control of Ketone Body Metabolism: On the Fringe of Lipid Biochemistry. Prostaglandins Leukot Essent Fat. Acids 2004, 70, 243–251. [Google Scholar] [CrossRef]
- Sumithran, P.; Proietto, J. Ketogenic Diets for Weight Loss: A Review of Their Principles, Safety and Efficacy. Obes. Res. Clin. Pract. 2008, 2, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Johnston, C.S.; Tjonn, S.L.; Swan, P.D.; White, A.; Hutchins, H.; Sears, B. Ketogenic Low-Carbohydrate Diets Have No Metabolic Advantage over Nonketogenic Low-Carbohydrate Diets. Am. J. Clin. Nutr. 2006, 83, 1055–1061. [Google Scholar] [CrossRef]
- Castro, A.I.; Gomez-Arbelaez, D.; Crujeiras, A.B.; Granero, R.; Aguera, Z.; Jimenez-Murcia, S.; Sajoux, I.; Lopez-Jaramillo, P.; Fernandez-Aranda, F.; Casanueva, F.F. Effect of a Very Low-Calorie Ketogenic Diet on Food and Alcohol Cravings, Physical and Sexual Activity, Sleep Disturbances, and Quality of Life in Obese Patients. Nutrients 2018, 10, 1348. [Google Scholar] [CrossRef] [PubMed]
- Schiavo, L.; Pilone, V.; Rossetti, G.; Barbarisi, A.; Cesaretti, M.; Iannelli, A. A 4-Week Preoperative Ketogenic Micronutrient-Enriched Diet Is Effective in Reducing Body Weight, Left Hepatic Lobe Volume, and Micronutrient Deficiencies in Patients Undergoing Bariatric Surgery: A Prospective Pilot Study. Obes. Surg. 2018, 28, 2215–2224. [Google Scholar] [CrossRef] [PubMed]
- de Luis, D.; Domingo, J.C.; Izaola, O.; Casanueva, F.F.; Bellido, D.; Sajoux, I. Effect of DHA Supplementation in a Very Low-Calorie Ketogenic Diet in the Treatment of Obesity: A Randomized Clinical Trial. Endocrine 2016, 54, 111–122. [Google Scholar] [CrossRef]
- Sajoux, I.; Lorenzo, P.M.; Gomez-Arbelaez, D.; Zulet, M.A.; Abete, I.; Castro, A.I.; Baltar, J.; Portillo, M.P.; Tinahones, F.J.; Martinez, J.A.; et al. Effect of a Very-Low-Calorie Ketogenic Diet on Circulating Myokine Levels Compared with the Effect of Bariatric Surgery or a Low-Calorie Diet in Patients with Obesity. Nutrients 2019, 11, 2368. [Google Scholar] [CrossRef]
- Castaldo, G.; Monaco, L.; Castaldo, L.; Galdo, G.; Cereda, E. An Observational Study of Sequential Protein-Sparing, Very Low-Calorie Ketogenic Diet (Oloproteic Diet) and Hypocaloric Mediterranean-like Diet for the Treatment of Obesity. Int. J. Food Sci. Nutr. 2016, 67, 696–706. [Google Scholar] [CrossRef]
- Goday, A.; Bellido, D.; Sajoux, I.; Crujeiras, A.B.; Burguera, B.; García-Luna, P.P.; Oleaga, A.; Moreno, B.; Casanueva, F.F. Short-Term Safety, Tolerability and Efficacy of a Very Low-Calorie-Ketogenic Diet Interventional Weight Loss Program versus Hypocaloric Diet in Patients with Type 2 Diabetes Mellitus. Nutr. Diabetes 2016, 6, e230. [Google Scholar] [CrossRef]
- Lagiou, P.; Sandin, S.; Lof, M.; Trichopoulos, D.; Adami, H.-O.; Weiderpass, E. Low Carbohydrate-High Protein Diet and Incidence of Cardiovascular Diseases in Swedish Women: Prospective Cohort Study. BMJ Br. Med. J. 2012, 344, e4026. [Google Scholar] [CrossRef]
- Axen, K.V.; Axen, K. Very Low-Carbohydrate versus Isocaloric High-Carbohydrate Diet in Dietary Obese Rats. Obesity 2006, 14, 1344–1352. [Google Scholar] [CrossRef] [PubMed]
- Greco, T.; Glenn, T.C.; Hovda, D.A.; Prins, M.L. Ketogenic Diet Decreases Oxidative Stress and Improves Mitochondrial Respiratory Complex Activity. J. Cereb. Blood Flow Metab. 2016, 36, 1603–1613. [Google Scholar] [CrossRef] [PubMed]
- Arsyad, A.; Idris, I.; Rasyid, A.A.; Usman, R.A.; Faradillah, K.R.; Latif, W.O.U.; Lubis, Z.I.; Aminuddin, A.; Yustisia, I.; Djabir, Y.Y. Long-Term Ketogenic Diet Induces Metabolic Acidosis, Anemia, and Oxidative Stress in Healthy Wistar Rats. J. Nutr. Metab. 2020, 2020, 3642035. [Google Scholar] [CrossRef] [PubMed]
- Burkitt, M.J. An Overlooked Danger of Ketogenic Diets: Making the Case That Ketone Bodies Induce Vascular Damage by the Same Mechanisms as Glucose. Nutrition 2020, 75–76, 110763. [Google Scholar] [CrossRef] [PubMed]
- Caton, S.J.; Yinglong, B.; Burget, L.; Spangler, L.J.; Tschöp, M.H.; Bidlingmaier, M. Low-Carbohydrate High-Fat Diets: Regulation of Energy Balance and Body Weight Regain in Rats. Obesity 2009, 17, 283–289. [Google Scholar] [CrossRef]
- Williams, S.; Basuald-Hammond, C.; Curtis, R.; Schuller, R. Growth Retardation in Children with Epilepsy on the Ketogenic Diet: A Retrospective Chart Review. J. Am. Diet. Assoc. 2002, 102, 405–407. [Google Scholar] [CrossRef]
- Hoffman, A.R. Treatment of the Adult Growth Hormone Deficiency Syndrome: Directions for Future Research. Growth Horm. IGF Res. 2005, 15, 48–52. [Google Scholar] [CrossRef]
- Ribeiro, L.C.; Chittó, A.L.; Müller, A.P.; Rocha, J.K.; Castro da Silva, M.; Quincozes-Santos, A.; Nardin, P.; Rotta, L.N.; Ziegler, D.R.; Gonçalves, C.-A.; et al. Ketogenic Diet-Fed Rats Have Increased Fat Mass and Phosphoenolpyruvate Carboxykinase Activity. Mol. Nutr. Food Res. 2008, 52, 1365–1371. [Google Scholar] [CrossRef]
- WHO. Obesity and Overweight: Fact Sheet. 2016. Available online: https://www.who.int/news-room/fact-sheets/detail/obesity-and-overweight (accessed on 9 October 2022).
- Fu, W.J.; Stromberg, A.J.; Viele, K.; Carroll, R.J.; Wu, G. Statistics and Bioinformatics in Nutritional Sciences: Analysis of Complex Data in the Era of Systems Biology. J. Nutr. Biochem. 2010, 21, 561–572. [Google Scholar] [CrossRef]
- Kinzig, K.P.; Scott, K.A.; Hyun, J.; Bi, S.; Moran, T.H. Altered Hypothalamic Signaling and Responses to Food Deprivation in Rats Fed a Low-Carbohydrate Diet. Obes. Res. 2005, 13, 1672–1682. [Google Scholar] [CrossRef] [PubMed]
- Sahagun, E.; Bachman, B.B.; Kinzig, K.P. Sex-Specific Effects of Ketogenic Diet after Pre-Exposure to a High-Fat, High-Sugar Diet in Rats. Nutr. Metab. Cardiovasc. Dis. 2021, 31, 961–971. [Google Scholar] [CrossRef] [PubMed]
- Vernon, G.; Baranova, A.; Younossi, Z.M. Systematic Review: The Epidemiology and Natural History of Non-Alcoholic Fatty Liver Disease and Non-Alcoholic Steatohepatitis in Adults. Aliment Pharmacol. Ther. 2011, 34, 274–285. [Google Scholar] [CrossRef] [PubMed]
- Kinzig, K.P.; Honors, M.A.; Hargrave, S.L.; Davenport, B.M.; Strader, A.D.; Wendt, D. Sensitivity to the Anorectic Effects of Leptin Is Retained in Rats Maintained on a Ketogenic Diet despite Increased Adiposity. Neuroendocrinology 2010, 92, 100–111. [Google Scholar] [CrossRef]
- Schwartz, M.W.; Woods, S.C.; Porte, D.; Seeley, R.J.; Baskin, D.G. Central Nervous System Control of Food Intake. Nature 2000, 404, 661–671. [Google Scholar] [CrossRef]
- Fam, B.C.; Morris, M.J.; Hansen, M.J.; Kebede, M.; Andrikopoulos, S.; Proietto, J.; Thorburn, A.W. Modulation of Central Leptin Sensitivity and Energy Balance in a Rat Model of Diet-Induced Obesity. Diabetes Obes. Metab. 2007, 9, 840–852. [Google Scholar] [CrossRef]
- Banks, W.A.; Coon, A.B.; Robinson, S.M.; Moinuddin, A.; Shultz, J.M.; Nakaoke, R.; Morley, J.E. Triglycerides Induce Leptin Resistance at the Blood-Brain Barrier. Diabetes 2004, 53, 1253–1260. [Google Scholar] [CrossRef]
- Choi, H.R.; Kim, J.; Lim, H.; Park, Y.K. Two-Week Exclusive Supplementation of Modified Ketogenic Nutrition Drink Reserves Lean Body Mass and Improves Blood Lipid Profile in Obese Adults: A Randomized Clinical Trial. Nutrients 2018, 10, 1895. [Google Scholar] [CrossRef]
- Kenig, S.; Petelin, A.; Poklar Vatovec, T.; Mohorko, N.; Jenko-Pražnikar, Z. Assessment of Micronutrients in a 12-Wk Ketogenic Diet in Obese Adults. Nutrition 2019, 67–68, 2–8. [Google Scholar] [CrossRef]
- Badman, M.K.; Kennedy, A.R.; Adams, A.C.; Pissios, P.; Maratos-Flier, E. A Very Low Carbohydrate Ketogenic Diet Improves Glucose Tolerance in Ob/Ob Mice Independently of Weight Loss. Am. J. Physiol. Endocrinol. Metab. 2009, 297, E1197–E1204. [Google Scholar] [CrossRef]
- Thio, L.L.; Erbayat-Altay, E.; Rensing, N.; Yamada, K.A. Leptin Contributes to Slower Weight Gain in Juvenile Rodents on a Ketogenic Diet. Pediatr. Res. 2006, 60, 413–417. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Liu, K.; Pei, L.; Li, S.; Zhao, J.; Zhang, K.; Zong, C.; Zhao, L.; Fang, H.; Wu, J.; et al. Atherogenic Index of Plasma Predicts Outcomes in Acute Ischemic Stroke. Front Neurol 2021, 12, 1728. [Google Scholar] [CrossRef] [PubMed]
- Mazidi, M.; Katsiki, N.; Mikhailidis, D.P.; Banach, M. Association of Ideal Cardiovascular Health Metrics with Serum Uric Acid, Inflammation and Atherogenic Index of Plasma: A Population-Based Survey. Atherosclerosis 2019, 284, 44–49. [Google Scholar] [CrossRef] [PubMed]
- Garbow, J.R.; Doherty, J.M.; Schugar, R.C.; Travers, S.; Weber, M.L.; Wentz, A.E.; Ezenwajiaku, N.; Cotter, D.G.; Brunt, E.M.; Crawford, P.A.; et al. Hepatic Steatosis, Inflammation, and ER Stress in Mice Maintained Long Term on a Very Low-Carbohydrate Ketogenic Diet. Am. J. Physiol. Gastrointest. Liver Physiol. 2011, 300, 956–967. [Google Scholar] [CrossRef] [PubMed]
- Jornayvaz, F.R.; Jurczak, M.J.; Lee, H.-Y.; Birkenfeld, A.L.; Frederick, D.W.; Zhang, D.; Zhang, X.-M.; Samuel, V.T.; Shulman, G.I. A High-Fat, Ketogenic Diet Causes Hepatic Insulin Resistance in Mice, despite Increasing Energy Expenditure and Preventing Weight Gain. Am. J. Physiol. Gastrointest. Liver Physiol. 2010, 299, 808–815. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, M.; Tuccinardi, D.; Ernesti, I.; Basciani, S.; Mariani, S.; Genco, A.; Manfrini, S.; Lubrano, C.; Gnessi, L. Scientific Evidence Underlying Contraindications to the Ketogenic Diet: An Update. Obes. Rev. 2020, 21, e13053. [Google Scholar] [CrossRef] [PubMed]
- Crosby, L.; Davis, B.; Joshi, S.; Jardine, M.; Paul, J.; Neola, M.; Barnard, N.D. Ketogenic Diets and Chronic Disease: Weighing the Benefits Against the Risks. Front. Nutr. 2021, 8, 702802. [Google Scholar] [CrossRef] [PubMed]
- Bruci, A.; Tuccinardi, D.; Tozzi, R.; Balena, A.; Santucci, S.; Frontani, R.; Mariani, S.; Basciani, S.; Spera, G.; Gnessi, L.; et al. Very Low-Calorie Ketogenic Diet: A Safe and Effective Tool for Weight Loss in Patients with Obesity and Mild Kidney Failure. Nutrients 2020, 12, 333. [Google Scholar] [CrossRef]
- Jia, P.; Huang, B.; You, Y.; Su, H.; Gao, L. Ketogenic Diet Aggravates Kidney Dysfunction by Exacerbating Metabolic Disorders and Inhibiting Autophagy in Spontaneously Hypertensive Rats. Biochem. Biophys. Res. Commun. 2021, 573, 13–18. [Google Scholar] [CrossRef]
- Rejeki, P.S.; Lutvyani, A.; Tjempakasari, A. Long-Term Ketogenic Diet Alters Kidney Function through Increasing Serum Creatinine Levels in Mice. J. Hunan Univ. Nat. Sci. 2021, 48, 113–120. [Google Scholar]
- Willi, S.M.; Oexmann, M.J.; Wright, N.M.; Collop, N.A.; Key Jr, L.L. The Effects of a High-Protein, Low-Fat, Ketogenic Diet on Adolescents With Morbid Obesity: Body Composition, Blood Chemistries, and Sleep Abnormalities. Pediatrics 1998, 101, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Nazarewicz, R.R.; Ziolkowski, W.; Vaccaro, P.S.; Ghafourifar, P. Effect of Short-Term Ketogenic Diet on Redox Status of Human Blood. Rejuvenation Res. 2007, 10, 435–440. [Google Scholar] [CrossRef] [PubMed]
- Kose, E.; Guzel, O.; Arslan, N. Analysis of Hematological Parameters in Patients Treated with Ketogenic Diet Due to Drug-Resistant Epilepsy. Neurol. Sci. 2018, 39, 85–89. [Google Scholar] [CrossRef] [PubMed]
- Finaud, J.; Lac, G.; Filaire, E. Oxidative Stress. Sport Med. 2006, 36, 327–358. [Google Scholar] [CrossRef] [PubMed]
- Rhyu, H.; Cho, S.-Y.; Roh, H.-T. The Effects of Ketogenic Diet on Oxidative Stress and Antioxidative Capacity Markers of Taekwondo Athletes. J. Exerc. Rehabil. 2014, 10, 362–366. [Google Scholar] [CrossRef]
- Valenzano, A.; Polito, R.; Trimigno, V.; di Palma, A.; Moscatelli, F.; Corso, G.; Sessa, F.; Salerno, M.; Montana, A.; di Nunno, N.; et al. Effects of Very Low Calorie Ketogenic Diet on the Orexinergic System, Visceral Adipose Tissue, and ROS Production. Antioxidants 2019, 8, 643. [Google Scholar] [CrossRef] [PubMed]
- Ho, E.; Karimi Galougahi, K.; Liu, C.-C.; Bhindi, R.; Figtree, G.A. Biological Markers of Oxidative Stress: Applications to Cardiovascular Research and Practice. Redox Biol. 2013, 1, 483–491. [Google Scholar] [CrossRef]
- Kanikarla-Marie, P.; Jain, S.K. Hyperketonemia (Acetoacetate) Upregulates NADPH Oxidase 4 and Elevates Oxidative Stress, ICAM-1, and Monocyte Adhesivity in Endothelial Cells. Cell. Physiol. Biochem. 2015, 35, 364–373. [Google Scholar] [CrossRef]
- Kaburagi, T.; Kanaki, K.; Otsuka, Y.; Hino, R. Low-Carbohydrate Diet Inhibits Different Advanced Glycation End Products in Kidney Depending on Lipid Composition but Causes Adverse Morphological Changes in a Non-Obese Model Mice. Nutrients 2019, 11, 2801. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Baseline | After 4 Weeks | ||||
|---|---|---|---|---|---|
| SD | HFD | SD | RKD | RSD | |
| Red blood cell system | |||||
| RBC [106/µL] | 7.68 ± 0.28 b* | 7.36 ± 0.24 b | 8.41 ± 0.35 a | 8.37 ± 0.40 a | 8.23 ± 0.50 a |
| HGB [g/dL] | 14.52 ± 0.57 a | 13.18 ± 0.65 c | 14.18 ± 0.33 a,b | 14.13 ± 0.83 a,b | 13.71 ± 0.81 b,c |
| HCT [%] | 42.09 ± 1.15 a,b | 40.59 ± 1.58 a,b | 41.66 ± 1.25 a,b | 42.11 ± 1.70 a | 40.29 ± 2.18 b |
| MCV [fl] | 55.00 (54.25; 55.00) a | 55.00 (54.25; 55.00) a | 49.50 (49.00; 50.75) b | 50.00 (49.75; 51.00) a,b | 49.50 (48.00; 50.00) b |
| MCH [pg] | 18.83 ± 1.00 a | 17.15 ± 2.03 b | 16.87 ± 0.58 b | 16.91 ± 0.57 b | 16.71 ± 0.28 b |
| MCHC [g/dL] | 33.95 (33.60; 34.68) a | 32.35 (29.30; 33.90) a | 34.00 (33.73; 34.20) a | 33.50 (33.30; 33.80) a | 33.90 (33.75; 34.33) a |
| RDWc [%] | 17.93 ± 0.34 b | 18.10 ± 0.49 a,b | 18.45 ± 0.31 a,b | 18.64 ± 0.41 a | 18.26 ± 0.36 a,b |
| White blood cell system | |||||
| WBC [103/µL] | 6.58 ± 0.47 a | 6.64 ± 1.41 a | 6.32 ± 1.02 a,b | 5.43 ± 0.67 b,c | 4.65 ± 0.56 c |
| LYM [103/µL] | 5.18 ± 0.72 a,b | 5.70 ± 1.24 a | 5.08 ± 1.04 a,b | 4.48 ± 0.79 b,c | 3.83 ± 0.38 c |
| MID [103/µL] | 0.57 (0.52; 0.61) a | 0.20 (0.13; 0.31) a,b | 0.43 (0.18; 0.47) a | 0.16 (0.11; 0.19) b | 0.20 (0.12; 0.27) b |
| GRA [103/µL] | 0.74 (0.72; 0.76) a | 0.62 (0.61; 0.63) a | 0.85 (0.83; 0.89) a | 0.79 (0.62; 0.81) a | 0.64 (0.50; 0.70) a |
| LYM% [%] | 82.08 (80.22; 82.48) a | 87.45 (83.25; 89.48) a | 77.90 (75.48; 82.80) a | 82.10 (79.88; 85.20) a | 85.05 (81.65; 93.00) a |
| MID% [%] | 8.39 (7.90; 9.12) a | 3.20 (2.23; 3.88) b | 7.50 (2.88; 8.38) a,b | 3.10 (2.00; 4.25) b | 4.61 (2.78; 5.30) a,b |
| GRA% [%] | 11.29 (10.85; 11.75) a | 9.55 (8.20; 12.85) a | 15.25 (13.58; 15.88) a | 13.85 (12.60; 14.93) a | 12.80 (11.13; 14.20) a |
| Platelet system | |||||
| PLT [103/µL] | 613.5 ± 114.62 b | 769.33 ± 121.95 a | 687.17 ± 68.48 a,b | 677.75 ± 98.4 a,b | 724.25 ± 76.21 a |
| PCT [%] | 0.51 ± 0.10 b | 0.63 ± 0.12 a | 0.59 ± 0.06 a,b | 0.54 ± 0.09 a,b | 0.60 ± 0.07 a,b |
| MPV [fl] | 8.05 (7.80; 8.53) a,b | 8.20 (7.98; 8.35) a,b | 8.55 (8.50; 8.68) a | 7.90 (7.80; 8.03) b | 7.95 (7.78; 8.40) a,b |
| PDWc [%] | 34.48 ± 1.59 b | 34.85 ± 0.43 b | 36.75 ± 0.62 a | 34.99 ± 0.46 b | 35.11 ± 1.10 b |
| Baseline | After 4 Weeks | ||||
|---|---|---|---|---|---|
| SD | HFD | SD | RKD | RSD | |
| Lipid profile | |||||
| Cholesterol [mmol/l] | 1.56 ± 0.07 a,b * | 1.67 ± 0.22 a | 1.55 ± 0.16 a,b | 1.57 ± 0.16 a | 1.43 ± 0.13 b |
| Tg [mmol/l] | 1.01 ± 0.21 b | 1.34 ± 0.18 a | 0.95 ± 0.27 b,c | 1.16 ± 0.36 a,b | 0.70 ± 0.24 c |
| HDL [mmol/l] | 0.61 ± 0.03 b | 0.51 ± 0.03 c | 0.64 ± 0.04 a,b | 0.66 ± 0.05 a | 0.55 ± 0.07 c |
| LDL [mmol/l] | 0.95 ± 0.05 b | 1.17 ± 0.21 a | 0.91 ± 0.14 b | 0.91 ± 0.16 b | 0.87 ± 0.12 b |
| Glucose [mmol/l] | 7.38 ± 1.06 b | 7.28 ± 0.71 b | 9.18 ± 0.42 b | 12.73 ± 2.52 a | 11.89 ± 2.16 a |
| AIP | 0.21 (0.18–0.28) b,c | 0.42 (0.36–0.46) a | 0.15 (0.14–0.23) b,c | 0.23 (0.15–0.32) b | 0.08 (0.01–0.20) c |
| Liver parameters | |||||
| AST [U/l] | 67.33 ± 3.71 b | 70.9 ± 3.81 a,b | 68.37 ± 7.37 b | 76.3 ± 8.70 a | 67.26 ± 7.44 b |
| ALT [U/l] | 33.65 (31.55; 37.10) a,b | 46.55 (42.88; 50.00) a | 17.30 (15.65; 18.20) b | 37.70 (33.90; 41.40) a | 15.65 (13.48; 16.68) b |
| ALP [U/l] | 122.65 ± 14.96 b | 139.57 ± 29.03 b | 72.13 ± 11.15 c | 173.07 ± 27.83 a | 87.67 ± 19.04 c |
| GGT [U/l] | <0.5 | <0.5 | <0.5 | <0.5 | <0.5 |
| Bilirubin [µmol/L] | <2.9 | <2.9 | <2.9 | <2.9 | <2.9 |
| Kidney parameters | |||||
| Uric Acid [µmol/L] | 31.00 (30.00; 37.25) a | 23.00 (22.25; 23.75) b | 37.00 (33.75; 40.25) a | 25.00 (22.50; 27.00) b | 35.50 (32.25; 50.55) a |
| Urea [mmol/l] | 4.50 ± 0.65 a | 3.06 ± 0.32 c | 3.66 ± 0.76 b | 1.77 ± 0.29 d | 2.98 ± 0.41 c |
| Creatinine [µmol/L] | 18.82 ± 2.07 c | 18.92 ± 1.30 c | 22.42 ± 1.14 b | 23.34 ± 2.10 b | 25.68 ± 1.29 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Drabińska, N.; Juśkiewicz, J.; Wiczkowski, W. The Effect of the Restrictive Ketogenic Diet on the Body Composition, Haematological and Biochemical Parameters, Oxidative Stress and Advanced Glycation End-Products in Young Wistar Rats with Diet-Induced Obesity. Nutrients 2022, 14, 4805. https://doi.org/10.3390/nu14224805
Drabińska N, Juśkiewicz J, Wiczkowski W. The Effect of the Restrictive Ketogenic Diet on the Body Composition, Haematological and Biochemical Parameters, Oxidative Stress and Advanced Glycation End-Products in Young Wistar Rats with Diet-Induced Obesity. Nutrients. 2022; 14(22):4805. https://doi.org/10.3390/nu14224805
Chicago/Turabian StyleDrabińska, Natalia, Jerzy Juśkiewicz, and Wiesław Wiczkowski. 2022. "The Effect of the Restrictive Ketogenic Diet on the Body Composition, Haematological and Biochemical Parameters, Oxidative Stress and Advanced Glycation End-Products in Young Wistar Rats with Diet-Induced Obesity" Nutrients 14, no. 22: 4805. https://doi.org/10.3390/nu14224805
APA StyleDrabińska, N., Juśkiewicz, J., & Wiczkowski, W. (2022). The Effect of the Restrictive Ketogenic Diet on the Body Composition, Haematological and Biochemical Parameters, Oxidative Stress and Advanced Glycation End-Products in Young Wistar Rats with Diet-Induced Obesity. Nutrients, 14(22), 4805. https://doi.org/10.3390/nu14224805