
Toxicity Investigations of (R)-3-Hydroxybutyrate Glycerides In Vitro and in Male and Female Rats
 ,
,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Test Substance
2.2. Bacterial Reverse Mutation Test (Ames Test)
2.3. In Vitro Micronucleus Assay
2.4. 14-Day Range Finding/Palatability Study in Rats
2.5. 90-Day Repeat Dose Study in Rats
2.6. Statistical Methods
3. Results
3.1. Mutagenicity
3.2. Micronucleus Study
3.3. 14-Day Range Finding/Palatability Study in Rats
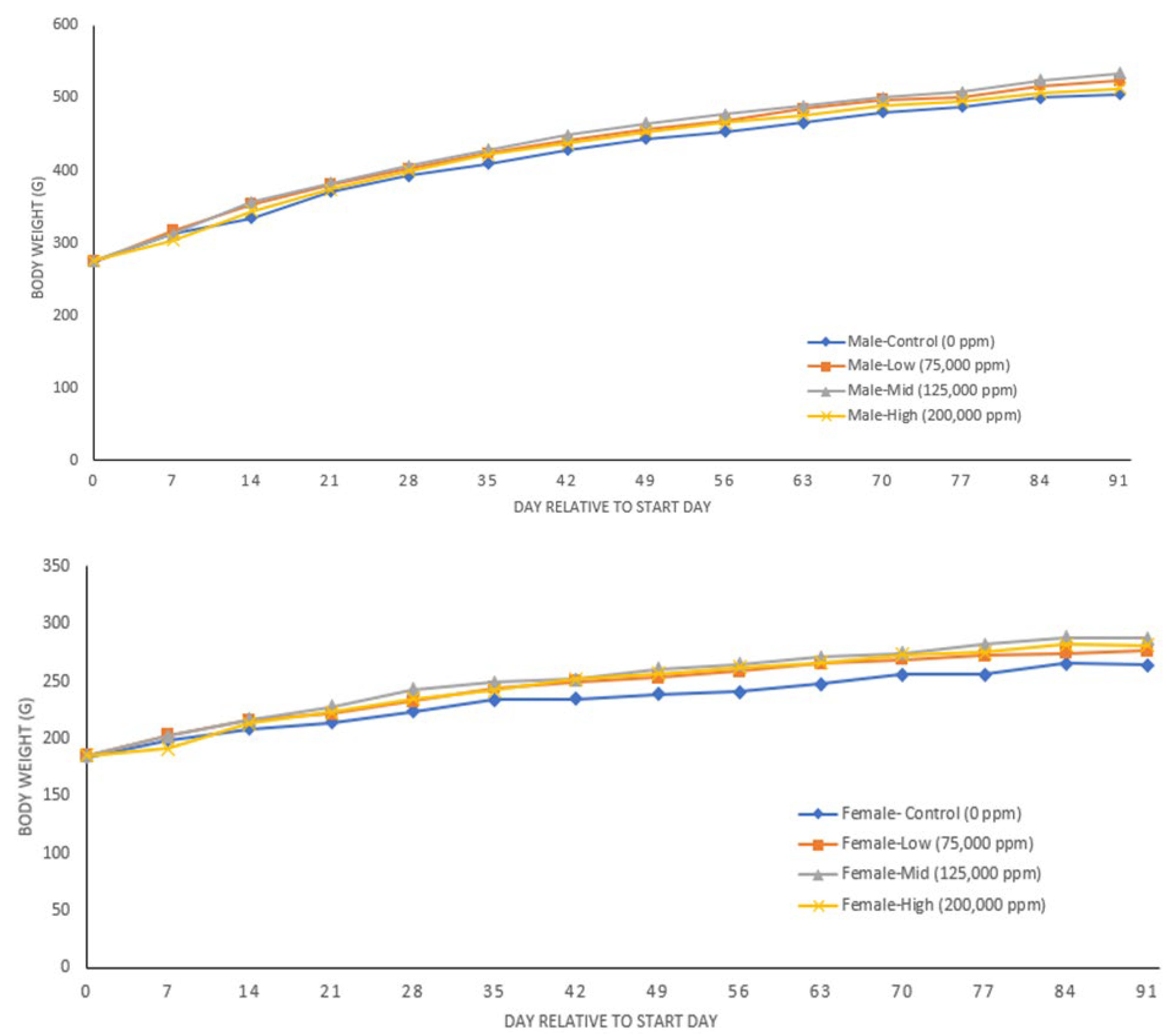
3.4. 90-Day Study
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Broom, G.M.; Shaw, I.C.; Rucklidge, J.J. The ketogenic diet as a potential treatment and prevention strategy for Alzheimer’s disease. Nutrition 2019, 60, 118–121. [Google Scholar] [CrossRef]
- LaFountain, R.A.; Miller, V.J.; Barnhart, E.C.; Hyde, P.N.; Crabtree, C.D.; McSwiney, F.T.; Beeler, M.K.; Buga, A.; Sapper, T.N.; Short, J.A.; et al. Extended Ketogenic Diet and Physical Training Intervention in Military Personnel. Mil. Med. 2019, 184, e538–e547. [Google Scholar] [CrossRef]
- Ma, S.; Suzuki, K. Keto-Adaptation and Endurance Exercise Capacity, Fatigue Recovery, and Exercise-Induced Muscle and Organ Damage Prevention: A Narrative Review. Sports 2019, 7, 40. [Google Scholar] [CrossRef] [PubMed]
- Martin-McGill, K.J.; Lambert, B.; Whiteley, V.J.; Wood, S.; Neal, E.G.; Simpson, Z.R.; Schoeler, N.E.; on behalf of the Ketogenic Dietitians Research Network (KDRN). Understanding the core principles of a ‘modified ketogenic diet’: A UK and Ireland perspective. J. Hum. Nutr. Diet. 2019, 32, 385–390. [Google Scholar] [CrossRef] [PubMed]
- Mohorko, N.; Černelič-Bizjak, M.; Poklar-Vatovec, T.; Grom, G.; Kenig, S.; Petelin, A.; Jenko-Pražnikar, Z. Weight loss, improved physical performance, cognitive function, eating behavior, and metabolic profile in a 12-week ketogenic diet in obese adults. Nutr. Res. 2018, 62, 64–77. [Google Scholar] [CrossRef] [PubMed]
- Muscogiuri, G.; Barrea, L.; Laudisio, D.; Pugliese, G.; Salzano, C.; Savastano, S.; Colao, A. The management of very low-calorie ketogenic diet in obesity outpatient clinic: A practical guide. J. Transl. Med. 2019, 17, 356. [Google Scholar] [CrossRef] [PubMed]
- Roehl, K.; Falco-Walter, J.; Ouyang, B.; Balabanov, A. Modified ketogenic diets in adults with refractory epilepsy: Efficacious improvements in seizure frequency, seizure severity, and quality of life. Epilepsy Behav. 2019, 93, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Stafstrom, C.E.; Rho, J.M. The Ketogenic Diet as a Treatment Paradigm for Diverse Neurological Disorders. Front. Pharmacol. 2012, 3, 59. [Google Scholar] [CrossRef] [PubMed]
- Westman, E.C.; Tondt, J.; Maguire, E.; Yancy, W.S., Jr. Implementing a low-carbohydrate, ketogenic diet to manage type 2 diabetes mellitus. Expert Rev. Endocrinol. Metab. 2018, 13, 263–272. [Google Scholar] [CrossRef] [PubMed]
- Włodarek, D. Role of Ketogenic Diets in Neurodegenerative Diseases (Alzheimer’s Disease and Parkinson’s Disease). Nutrients 2019, 11, 169. [Google Scholar] [CrossRef] [PubMed]
- Kolb, H.; Kempf, K.; Röhling, M.; Martin, S. Insulin: Too much of a good thing is bad. BMC Med. 2020, 18, 224. [Google Scholar] [CrossRef] [PubMed]
- Kolb, H.; Stumvoll, M.; Kramer, W.; Kempf, K.; Martin, S. Insulin translates unfavourable lifestyle into obesity. BMC Med. 2018, 16, 232. [Google Scholar] [CrossRef]
- Laffel, L. Ketone bodies: A review of physiology, pathophysiology and application of monitoring to diabetes. Diabetes/Metab. Res. Rev. 1999, 15, 412–426. [Google Scholar] [CrossRef]
- Silva, B.; Mantha, O.L.; Schor, J.; Pascual, A.; Plaçais, P.-Y.; Pavlowsky, A.; Preat, T. Glia fuel neurons with locally synthesized ketone bodies to sustain memory under starvation. Nat. Metab. 2022, 4, 213–224. [Google Scholar] [CrossRef]
- Weis, E.; Puchalska, P.; Nelson, A.B.; Taylor, J.; Moll, I.; Hasan, S.S.; Dewenter, M.; Hagenmüller, M.; Fleming, T.; Poschet, G.; et al. Ketone body oxidation increases cardiac endothelial cell proliferation. EMBO Mol. Med. 2022, 14, e14753. [Google Scholar] [CrossRef]
- Cameron, D.; Soto-Mota, A.; Willis, D.R.; Ellis, J.; Procter, N.E.K.; Greenwood, R.; Saunders, N.; Schulte, R.F.; Vassiliou, V.S.; Tyler, D.J.; et al. Evaluation of Acute Supplementation With the Ketone Ester (R)-3-Hydroxybutyl-(R)-3-Hydroxybutyrate (deltaG) in Healthy Volunteers by Cardiac and Skeletal Muscle 31P Magnetic Resonance Spectroscopy. Front. Physiol. 2022, 13, 793987. [Google Scholar] [CrossRef] [PubMed]
- Cuenoud, B.; Hartweg, M.; Godin, J.-P.; Croteau, E.; Maltais, M.; Castellano, C.-A.; Carpentier, A.C.; Cunnane, S.C. Metabolism of Exogenous D-Beta-Hydroxybutyrate, an Energy Substrate Avidly Consumed by the Heart and Kidney. Front. Nutr. 2020, 7, 13. [Google Scholar] [CrossRef] [PubMed]
- Bak, L.K.; Walls, A.B.; Schousboe, A.; Waagepetersen, H.S. Astrocytic glycogen metabolism in the healthy and diseased brain. J. Biol. Chem. 2018, 293, 7108–7116. [Google Scholar] [CrossRef] [PubMed]
- Bordone, M.; Salman, M.M.; Titus, H.E.; Amini, E.; Andersen, J.V.; Chakraborti, B.; Diuba, A.V.; Dubouskaya, T.G.; Ehrke, E.; De Freitas, A.E.; et al. The energetic brain—A review from students to students. J. Neurochem. 2019, 151, 139–165. [Google Scholar] [CrossRef] [PubMed]
- Courchesne-Loyer, A.; Croteau, E.; Castellano, C.-A.; St-Pierre, V.; Hennebelle, M.; Cunnane, S.C. Inverse relationship between brain glucose and ketone metabolism in adults during short-term moderate dietary ketosis: A dual tracer quantitative positron emission tomography study. J. Cereb. Blood Flow Metab. 2016, 37, 2485–2493. [Google Scholar] [CrossRef]
- Jensen, N.J.; Wodschow, H.Z.; Nilsson, M.; Rungby, J. Effects of Ketone Bodies on Brain Metabolism and Function in Neurodegenerative Diseases. Int. J. Mol. Sci. 2020, 21, 8767. [Google Scholar] [CrossRef] [PubMed]
- Oyarzabal, A.; Marin-Valencia, I. Synaptic energy metabolism and neuronal excitability, in sickness and health. J. Inherit. Metab. Dis. 2019, 42, 220–236. [Google Scholar] [CrossRef] [PubMed]
- Andreux, P.A.; Van Diemen, M.P.J.; Heezen, M.R.; Auwerx, J.; Rinsch, C.; Groeneveld, G.J.; Singh, A. Mitochondrial function is impaired in the skeletal muscle of pre-frail elderly. Sci. Rep. 2018, 8, 8548. [Google Scholar] [CrossRef] [PubMed]
- Bleeker, J.C.; Visser, G.; Clarke, K.; Ferdinandusse, S.; de Haan, F.H.; Houtkooper, R.H.; Ijlst, L.; Kok, I.L.; Langeveld, M.; van der Pol, W.L.; et al. Nutritional ketosis improves exercise metabolism in patients with very long-chain acyl-CoA dehydrogenase deficiency. J. Inherit. Metab. Dis. 2020, 43, 787–799. [Google Scholar] [CrossRef]
- Evans, M.; Cogan, K.E.; Egan, B. Metabolism of ketone bodies during exercise and training: Physiological basis for exogenous supplementation. J. Physiol. 2016, 595, 2857–2871. [Google Scholar] [CrossRef]
- Puchalska, P.; Crawford, P.A. Metabolic and Signaling Roles of Ketone Bodies in Health and Disease. Annu. Rev. Nutr. 2021, 41, 49–77. [Google Scholar] [CrossRef]
- Abdurrachim, D.; Woo, C.C.; Teo, X.Q.; Chan, W.X.; Radda, G.K.; Lee, P.T.H. A new hyperpolarized 13C ketone body probe reveals an increase in acetoacetate utilization in the diabetic rat heart. Sci. Rep. 2019, 9, 5532. [Google Scholar] [CrossRef]
- Al-Zaid, N.S.; Dashti, H.M.; Mathew, T.C.; Juggi, J.S. Low carbohydrate ketogenic diet enhances cardiac tolerance to global ischaemia. Acta Cardiol. 2007, 62, 381–389. [Google Scholar] [CrossRef]
- Chu, Y.; Zhang, C.; Xie, M. Beta-Hydroxybutyrate, Friend or Foe for Stressed Hearts. Front. Aging 2021, 2, 681513. [Google Scholar] [CrossRef]
- Han, Y.-M.; Ramprasath, T.; Zou, M.-H. β-hydroxybutyrate and its metabolic effects on age-associated pathology. Exp. Mol. Med. 2020, 52, 548–555. [Google Scholar] [CrossRef]
- Dhillon, K.K.; Gupta, S. Biochemistry, Ketogenesis; [Updated 2022 February 10]; StatPearls: Treasure Island, FL, USA, 2022. [Google Scholar]
- Brahma, M.K.; Ha, C.; Pepin, M.E.; Mia, S.; Sun, Z.; Chatham, J.C.; Habegger, K.M.; Abel, E.D.; Paterson, A.J.; Young, M.E.; et al. Increased Glucose Availability Attenuates Myocardial Ketone Body Utilization. J. Am. Heart Assoc. 2020, 9, e013039. [Google Scholar] [CrossRef] [PubMed]
- Cahill, G.F., Jr. Fuel Metabolism in Starvation. Annu. Rev. Nutr. 2006, 26, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Jagadish, S.; Payne, E.T.; Wong-Kisiel, L.; Nickels, K.C.; Eckert, S.; Wirrell, E.C. The Ketogenic and Modified Atkins Diet Therapy for Children With Refractory Epilepsy of Genetic Etiology. Pediatr. Neurol. 2018, 94, 32–37. [Google Scholar] [CrossRef] [PubMed]
- Newman, J.C.; Verdin, E. Ketone bodies as signaling metabolites. Trends Endocrinol. Metab. 2013, 25, 42–52. [Google Scholar] [CrossRef] [PubMed]
- Poff, A.M.; Moss, S.; Soliven, M.; D’Agostino, D.P. Ketone Supplementation: Meeting the Needs of the Brain in an Energy Crisis. Front. Nutr. 2021, 8, 783659. [Google Scholar] [CrossRef]
- Poplawski, M.M.; Mastaitis, J.W.; Isoda, F.; Grosjean, F.; Zheng, F.; Mobbs, C.V. Reversal of Diabetic Nephropathy by a Ketogenic Diet. PLoS ONE 2011, 6, e18604. [Google Scholar] [CrossRef]
- Puchalska, P.; Crawford, P.A. Multi-dimensional Roles of Ketone Bodies in Fuel Metabolism, Signaling, and Therapeutics. Cell Metab. 2017, 25, 262–284. [Google Scholar] [CrossRef]
- Rojas-Morales, P.; Pedraza-Chaverri, J.; Tapia, E. Ketone bodies, stress response, and redox homeostasis. Redox Biol. 2019, 29, 101395. [Google Scholar] [CrossRef]
- Rusek, M.; Pluta, R.; Ułamek-Kozioł, M.; Czuczwar, S.J. Ketogenic Diet in Alzheimer’s Disease. Int. J. Mol. Sci. 2019, 20, 3892. [Google Scholar] [CrossRef]
- Waldman, H.S.; McAllister, M.J. Exogenous Ketones as Therapeutic Signaling Molecules in High-Stress Occupations: Implications for Mitigating Oxidative Stress and Mitochondrial Dysfunction in Future Research. Nutr. Metab. Insights 2020, 13, 1178638820979029. [Google Scholar] [CrossRef]
- Walsh, J.J.; Caldwell, H.G.; Neudorf, H.; Ainslie, P.N.; Little, J.P. Short-term ketone monoester supplementation improves cerebral blood flow and cognition in obesity: A randomized cross-over trial. J. Physiol. 2021, 599, 4763–4778. [Google Scholar] [CrossRef] [PubMed]
- Cox, P.J.; Kirk, T.; Ashmore, T.; Willerton, K.; Evans, R.; Smith, A.; Murray, A.J.; Stubbs, B.; West, J.; McLure, S.W.; et al. Nutritional Ketosis Alters Fuel Preference and Thereby Endurance Performance in Athletes. Cell Metab. 2016, 24, 256–268. [Google Scholar] [CrossRef] [PubMed]
- Harvey, K.L.; Holcomb, L.E.; Kolwicz, S.C., Jr. Ketogenic diets and exercise performance. Nutrients 2019, 11, 2296. [Google Scholar] [CrossRef] [PubMed]
- CIR. Safety Assessment of Glycerin as Used in Cosmetics. Available online: https://www.cir-safety.org/sites/default/files/glycer_122014_FR.pdf (accessed on 1 August 2022).
- Stoewsand, G.S.; Dymsza, H.A. Synthetic sources of calories in the diets of rats and dogs. Proc. Seventh Int. Congr. Nutr. 1966, 4, 1082–1087. [Google Scholar]
- UK. SIDS Initial Assessment Report for SIAM 14: Glycerol, Sponsored by the United Kingdom for OECD SIDS Chemical Program. Available online: https://hpvchemicals.oecd.org/ui/handler.axd?id=4b0a2d87-3183-40d4-84f5-0e118c647b19 (accessed on 1 August 2022).
- Shivva, V.; Cox, P.J.; Clarke, K.; Veech, R.L.; Tucker, I.G.; Duffull, S.B. The Population Pharmacokinetics of d-β-hydroxybutyrate Following Administration of (R)-3-Hydroxybutyl (R)-3-Hydroxybutyrate. AAPS J. 2016, 18, 678–688. [Google Scholar] [CrossRef]
- Dedkova, E.N.; Blatter, L.A. Role of β-hydroxybutyrate, its polymer poly-β-hydroxybutyrate and inorganic polyphosphate in mammalian health and disease. Front. Physiol. 2014, 5, 260. [Google Scholar] [CrossRef]
- Clarke, K.; Tchabanenko, K.; Pawlosky, R.; Carter, E.; Knight, N.S.; Murray, A.J.; Cochlin, L.E.; King, M.T.; Wong, A.W.; Roberts, A.; et al. Oral 28-day and developmental toxicity studies of (R)-3-hydroxybutyl (R)-3-hydroxybutyrate. Regul. Toxicol. Pharmacol. 2012, 63, 196–208. [Google Scholar] [CrossRef]
- WHO. Chemical-Specific Adjustment Factors for Interspecies Differences and Human Variability: Guidance Document for Use of Data in Dose/Concentration-Response Assessment. Available online: https://apps.who.int/iris/bitstream/handle/10665/43294/9241546786_eng.pdf;sequence=1 (accessed on 15 September 2022).
- Borzelleca, J.F. Macronutrient substitutes: Safety evaluation. Regul. Toxicol. Pharmacol. 1992, 16, 253–264. [Google Scholar] [CrossRef]
- FDA. GRAS Notice (GRN) 515, D-Beta-Hydroxybutyrate Ester. Available online: https://www.cfsanappsexternal.fda.gov/scripts/fdcc/index.cfm?set=GRASNotices&id=515&sort=GRN_No&order=DESC&startrow=1&type=basic&search=515 (accessed on 15 September 2022).
- FDA. Gras Notice (GRN) No. 1032, D-β-Hydroxybutyrate. Available online: https://www.cfsanappsexternal.fda.gov/scripts/fdcc/?set=GRASNotices&id=1032&sort=GRN_No&order=DESC&startrow=1&type=basic&search=butyrate (accessed on 15 September 2022).
- Stefan, M.; Sharp, M.; Gheith, R.; Lowery, R.; Wilson, J. The Effects of Exogenous Beta-Hydroxybutyrate Supplementation on Metrics of Safety and Health. Int. J. Nutr. Food Sci. 2020, 9, 154–162. [Google Scholar] [CrossRef]


{kind=link}
| Concentration (µg/Plate) | TA98 | TA100 | TA1535 | TA1537 | WP2uvrA | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| −S9 | +S9 | −S9 | +S9 | −S9 | +S9 | −S9 | +S9 | −S9 | +S9 | |
| Experiment 1 | ||||||||||
| 0 a | 21 | 29 | 116 | 120 | 11 | 13 | 11 | 15 | 46 | 53 |
| 1.58 | 25 | 29 | 120 | 116 | 14 | 11 | 9 | 15 | 48 | 53 |
| 5 | 22 | 23 | 122 | 119 | 13 | 12 | 13 | 11 | 53 | 56 |
| 15.8 | 21 | 29 | 116 | 123 | 11 | 13 | 11 | 17 | 43 | 57 |
| 50 | 22 | 31 | 112 | 111 | 10 | 10 | 16 | 15 | 44 | 56 |
| 158 | 25 | 29 | 117 | 125 | 11 | 15 | 15 | 10 | 46 | 52 |
| 500 | 20 | 30 | 110 | 120 | 10 | 12 | 9 | 12 | 47 | 53 |
| 1580 | 22 | 29 | 119 | 122 | 12 | 8 | 9 | 16 | 54 | 50 |
| 5000 | 24 | 30 | 112 | 109 | 13 | 14 | 13 | 10 | 55 | 50 |
| Positive control | 194 b | 159 c | 519 d | 746 c | 547 d | 492 e | 247 f | 254 e | 612 g | 180 e |
| Historical negative control (−S9, range) | 15–35 | 72–129 | 6–20 | 5–17 | 30–66 | |||||
| Historical negative control (+S9, range) | 18–32 | 78–123 | 8–18 | 8–20 | 32–68 | |||||
| Experiment 2 | ||||||||||
| 0 a | 21 | 27 | 119 | 120 | 12 | 12 | 9 | 10 | 46 | 54 |
| 1.58 | 20 | 24 | 109 | 118 | 13 | 12 | 8 | 8 | 50 | 53 |
| 5 | 18 | 25 | 111 | 117 | 13 | 15 | 8 | 11 | 45 | 59 |
| 15.8 | 21 | 23 | 114 | 120 | 10 | 9 | 9 | 12 | 43 | 62 |
| 50 | 25 | 25 | 105 | 102 | 10 | 9 | 10 | 11 | 44 | 52 |
| 158 | 20 | 25 | 111 | 113 | 13 | 11 | 8 | 12 | 42 | 59 |
| 500 | 21 | 25 | 113 | 107 | 17 | 11 | 10 | 13 | 48 | 50 |
| 1580 | 22 | 21 | 102 | 119 | 11 | 11 | 9 | 13 | 49 | 53 |
| 5000 | 24 | 21 | 110 | 118 | 11 | 9 | 10 | 10 | 47 | 53 |
| Positive control | 176 b | 168 c | 510 d | 622 c | 521 d | 391 e | 452 f | 248 e | 702 g | 178 e |
| Historical negative control (−S9, range) | 17–38 | 80–138 | 6–22 | 6–16 | 26–59 | |||||
| Historical negative control (+S9, range) | 17–38 | 82–134 | 6–17 | 6–14 | 32–62 | |||||
| Concentration (µg/mL) | % A/N | Nuclei: Beads | % Relative Survival | % MN | p-Value | RICC |
|---|---|---|---|---|---|---|
| Four hour incubation without S9 | ||||||
| 0 (water) | 1.8 | 9.0 | 100 | 0.5 | NR | 100.0 |
| 55 | 2.2 | 8.5 | 94.0 | 0.5 | 0.9398 | 101.7 |
| 111 | 1.6 | 8.8 | 97.9 | 0.6 | 0.8308 | 103.9 |
| 223 | 2.1 | 9.1 | 100.8 | 0.4 | 0.9986 | 95.8 |
| 445 | 2.0 | 8.5 | 94.2 | 0.7 | 0.1550 | 98.1 |
| 890 | 1.6 | 9.0 | 99.9 | 0.6 | 0.6813 | 95.5 |
| 1780 | 1.8 | 8.7 | 95.9 | 0.5 | 0.9270 | 93.7 |
| Trend | 0.4361 | |||||
| VIN | 15.6 | 5.1 | 56.5 | 14.4 * | <0.0001 | 98.1 |
| Four hour incubation with S9 | ||||||
| 0 (water) | 1.7 | 12.6 | 100 | 0.5 | NR | 100 |
| 55 | 1.3 | 12.8 | 101.5 | 0.4 | 0.9995 | 96.7 |
| 111 | 1.4 | 13.5 | 107.0 | 0.5 | 0.9120 | 82.2 |
| 223 | 1.4 | 15.1 | 120.1 | 0.4 | 0.9995 | 92.9 |
| 445 | 1.3 | 14.1 | 112.0 | 0.6 | 0.7759 | 95.1 |
| 890 | 1.1 | 13.2 | 105.0 | 0.6 | 0.6593 | 93.9 |
| 1780 | 1.5 | 15.1 | 120.2 | 0.6 | 0.4419 | 82.2 |
| Trend | 0.0553 | |||||
| CP | 7.5 | 8.1 | 64.2 | 4.9 * | 0.0005 | 39.7 |
| Twenty four hour incubation without S9 | ||||||
| 0 (water) | 1.8 | 11.7 | 100.0 | 0.6 | NR | 100 |
| 55 | 1.6 | 11.6 | 98.9 | 0.5 | 0.9999 | 101.3 |
| 111 | 1.8 | 11.8 | 101.0 | 0.4 | 1.0000 | 107.8 |
| 223 | 1.6 | 12.7 | 108.3 | 0.4 | 1.0000 | 102.5 |
| 445 | 1.5 | 11.3 | 96.3 | 0.4 | 1.0000 | 81.9 |
| 890 | 1.5 | 11.3 | 96.3 | 0.6 | 0.9866 | 100.1 |
| 1780 | 1.4 | 13.2 | 112.4 | 0.3 | 1.0000 | 109.5 |
| Trend | 0.0902 | |||||
| VIN | 11.8 | 7.2 | 61.2 | 12.0 * | <0.0001 | 59.9 |
| Parameter | Control | 75,000 ppm | 125,000 ppm | 200,000 ppm | Historical Control |
|---|---|---|---|---|---|
| Males | |||||
| AST (U/L) | 79.5 ± 22.3 | 80.5 ± 14.8 | 77.6 ± 18.9 | 72.3 ± 10.4 | 63–175 |
| ALT (U/L) | 33.1 ± 7.7 | 35.1 ± 8.0 | 33.2 ± 7.7 | 24.1 ± 3.1 ** | 19–48 |
| ALKP (U/L) | 52.2 ± 10.1 | 48.0 ± 6.7 | 49.6 ± 8.9 | 47.4 ± 10.3 | 36–141 |
| BILI (mg/dL) | 0.068 ± 0.019 | 0.065 ± 0.012 | 0.058 ± 0.018 | 0.053 ± 0.0016 | 0.04–0.2 |
| BUN (mg/dL) | 19.9 ± 3.4 | 18.4 ± 1.7 | 18.8 ± 2.9 | 17.9 ± 3.3 | 10.7–20.0 |
| CREA (mg/dL) | 0.249 ± 0.039 | 0.224 ± 0.033 | 0.263 ± 0.050 | 0.233 ± 0.056 | 0.3–0.5 |
| CHOL (mg/dL) | 70.9 ± 11.3 | 72.8 ± 15.1 | 65.1 ± 15.5 | 64.7 ± 7.5 | 37–95 |
| LDL (mmol/L) | 0.375 ± 0.073 | 0.342 ± 0.126 | 0.313 ± 0.120 | 0.268 ± 0.039 | ND |
| HDL (mmol/L) | 1.221 ± 0.258 | 1.291 ± 0.298 | 1.145 ± 0.302 | 1.108 ± 0.146 | ND |
| TRIG (mg/dL) | 63.7 ± 18.4 | 99.8 ± 28.4 ** | 85.8 ± 25.2 | 101.2 ± 35.7 ** | 27–160 |
| SDH (U/L) | 15.29 ± 4.31 | 20.50 ± 8.84 | 18.70 ± 5.78 | 14.94 ± 5.19 | ND |
| GLUC (mg/dL) | 230.6 ± 48.0 | 237.3 ± 50.8 | 259.7 ± 47.7 | 227.7 ± 49.5 | 106–184 |
| TP (g/dL) | 5.46 ± 0.26 | 5.56 ± 0.35 | 5.73 ± 0.29 | 5.62 ± 0.15 | 5.6–7.6 |
| ALB (g/dL) | 3.90 ± 0.16 | 3.96 ± 0.23 | 4.05 ± 0.18 | 3.94 ± 0.17 | 3.6–4.7 |
| GLOB (g/dL) | 1.56 ± 0.19 | 1.60 ± 0.23 | 1.68 ± 0.23 | 1.68 ± 0.14 | 1.8–2.5 |
| Ca (mg/dL) | 12.02 ± 0.66 | 11.92 ± 0.72 | 12.52 ± 0.78 | 11.78 ± 0.32 | 9.1–11.9 |
| P (mg/dL) | 9.24 ± 0.69 | 9.27 ± 1.02 | 10.04 ± 0.80 | 8.81 ± 0.52 | 3.64–8.4 |
| Na (mmol/L) | 142.2 ± 2.7 | 141.0 ± 1.9 | 141.2 ± 2.1 | 142.0 ± 3.2 | 137–147 |
| K (mmol/L) | 7.16 ± 0.90 | 7.80 ± 0.97 | 8.31 ± 1.24 * | 6.79 ± 0.82 | 3.88–6.11 |
| Cl (mmol/L) | 103.62 ± 1.70 | 103.52 ± 2.11 | 103.20 ± 1.84 | 103.25 ± 1.59 | 98–106 |
| TSH (ng/mL) | 3.245 ± 0.212 | 2.827 ± 0.293 * | 3.930 ± 0.444 ** | 3.590 ± 0.595 | ND |
| T3 (ng/mL) | 1.365 ± 0.052 | 1.471 ± 0.097 * | 1.684 ± 0.122 *** | 1.768 ± 0.085 *** | ND |
| T4 (ng/mL) | 62.818 ± 5.247 | 57.874 ± 5.593 | 51.336 ± 4.332 *** | 53.748 ± 3.091 *** | ND |
| Females | |||||
| AST (U/L) | 108.8 ± 17.0 | 86.4 ± 26.6 * | 88.7 ± 18.3 | 69.0 ± 11.6 *** | 64–222 |
| ALT (U/L) | 31.4 ± 8.3 | 23.5 ± 7.3 * | 24.5 ± 4.7 | 20.1 ± 4.1 *** | 14–64 |
| ALKP (U/L) | 30.4 ± 10.2 | 19.6 ± 2.3 ** | 22.4 ± 6.4 * | 18.5 ± 3.9 *** | 18–62 |
| BILI (mg/dL) | 0.067 ± 0.020 | 0.070 ± 0.018 | 0.057 ± 0.016 | 0.062 ± 0.018 | 0.07–0.2 |
| BUN (mg/dL) | 23.8 ± 3.9 | 23.0 ± 4.1 | 22.2 ± 4.7 | 21.2 ± 4.5 | 11.7–25.0 |
| CREA (mg/dL) | 0.313 ± 0.050 | 0.289 ± 0.056 | 0.268 ± 0.046 | 0.237 ± 0.035 ** | 0.3–0.6 |
| CHOL (mg/dL) | 46.5 ± 12.0 | 61.7 ± 11.0 * | 63.7 ± 12.0 ** | 68.7 ± 9.9 *** | 23–97 |
| LDL (mmol/L) | 0.120 ± 0.036 | 0.144 ± 0.044 | 0.146 ± 0.040 | 0.160 ± 0.043 | ND |
| HDL (mmol/L) | 1.023 ± 0.266 | 1.358 ± 0.217 * | 1.374 ± 0.255 ** | 1.477 ± 0.261 *** | ND |
| TRIG (mg/dL) | 40.9 ± 11.9 | 50.3 ± 9.8 | 51.2 ± 10.4 | 55.6 ± 16.5 | 16–175 |
| SDH (U/L) | 13.75 ± 6.31 | 15.79 ± 7.21 | 11.36 ± 5.10 | 11.60 ± 3.76 | ND |
| GLUC (mg/dL) | 174.0 ± 50.0 | 175.5 ± 51.9 | 190.4 ± 63.8 | 195.2 ± 28.0 | 89–163 |
| TP (g/dL) | 5.24 ± 0.43 | 5.78 ± 0.26 * | 5.59 ± 0.33 | 5.73 ± 0.63 * | 5.7–8.3 |
| ALB (g/dL) | 4.10 ± 0.42 | 4.57 ± 0.17 | 4.32 ± 0.34 | 4.59 ± 0.67 | 3.7–5.8 |
| GLOB (g/dL) | 1.14 ± 0.18 | 1.21 ± 0.17 | 1.27 ± 0.09 | 1.14 ± 0.16 | 1.6–2.3 |
| Ca (mg/dL) | 11.20 ± 0.54 | 11.98 ± 0.67 | 12.55 ± 1.25 ** | 12.65 ± 1.06 ** | 9.5–12.1 |
| P (mg/dL) | 9.12 ± 1.38 | 9.13 ± 1.00 | 9.35 ± 1.42 | 9.23 ± 1.13 | 4.53–9.51 |
| Na (mmol/L) | 138.1 ± 3.7 | 140.9 ± 1.9 | 142.0 ± 3.0 * | 139.8 ± 2.7 | 135–146 |
| K (mmol/L) | 7.67 ± 1.70 | 7.33 ± 1.27 | 6.93 ± 0.91 | 7.40 ± 1.00 | 3.37–5.11 |
| Cl (mmol/L) | 104.72 ± 2.74 | 105.67 ± 2.25 | 107.06 ± 2.58 | 104.98 ± 1.81 | 97–106 |
| TSH (ng/mL) | 3.316 ± 0.424 | 2.955 ± 0.191 | 3.956 ± 0.500 ** | 3.564 ± 0.463 | ND |
| T3 (ng/mL) | 1.579 ± 0.139 | 1.710 ± 0.299 | 1.827 ± 0.069 ** | 1.844 ± 0.177 ** | ND |
| T4 (ng/mL) | 59.631 ± 8.321 | 57.190 ± 9.115 | 57.918 ± 6.676 | 53.628 ± 5.312 | ND |
| Parameter | Control | 75,000 ppm | 125,000 ppm | 200,000 ppm |
|---|---|---|---|---|
| Males | ||||
| Terminal body weight (g) | 483.6 ± 31.3 | 500.3 ± 32.3 | 510.2 ± 35.7 | 496.3 ± 24.7 |
| Adrenals (g) | 0.078 ± 0.011 | 0.075 ± 0.013 | 0.075 ± 0.011 | 0.076 ± 0.018 |
| Adrenals/TBW | 0.163 ± 0.027 | 0.149 ± 0.021 | 0.146 ± 0.017 | 0.152 ± 0.032 |
| Brain (g) | 2.194 ± 0.073 | 2.188 ± 0.083 | 2.159 ± 0.263 | 2.178 ± 0.115 |
| Brain/TBW | 4.551 ± 0.276 | 4.385 ± 0.259 | 4.235 ± 0.490 | 4.397 ± 0.300 |
| Epididymides (g) | 1.546 ± 0.109 | 1.640 ± 0.125 | 1.666 ± 0.146 | 1.578 ± 0.198 |
| Epididymides/TBW | 3.204 ± 0.258 | 3.286 ± 0.272 | 3.283 ± 0.408 | 3.180 ± 0.366 |
| Heart (g) | 1.323 ± 0.099 | 1.335 ± 0.074 | 1.398 ± 0.066 | 1.280 ± 0.076 |
| Heart/TBW | 2.736 ± 0.125 | 2.673 ± 0.140 | 2.750 ± 0.209 | 2.581 ± 0.124 |
| Kidneys (g) | 3.403 ± 0.304 | 3.418 ± 0.308 | 3.560 ± 0.214 | 3.460 ± 0.229 |
| Kidneys/TBW | 7.052 ± 0.659 | 6.827 ± 0.320 | 7.005 ± 0.599 | 6.972 ± 0.315 |
| Liver (g) | 12.803 ± 1.164 | 13.572 ± 1.488 | 14.248 ± 0.960 | 13.590 ± 0.673 |
| Liver/TBW | 26.441 ± 1.044 | 27.094 ± 1.962 | 27.964 ± 1.530 | 27.415 ± 1.412 |
| Pituitary (g) | 0.016 ± 0.006 | 0.016 ± 0.004 | 0.017 ± 0.006 | 0.021 ± 0.004 |
| Pituitary/TBW | 0.003 ± 0.001 | 0.003 ± 0.001 | 0.003 ± 0.001 | 0.004 ± 0.001 |
| Prostate + SV + CG (g) | 3.580 ± 0.412 | 3.329 ± 0.665 | 3.525 ± 0.524 | 3.242 ± 0.337 |
| Prostate + SV + CG/TBW | 0.007 ± 0.001 | 0.007 ± 0.001 | 0.007 ± 0.001 | 0.007 ± 0.001 |
| Spleen (g) | 0.897 ± 0.154 | 0.940 ± 0.142 | 1.021 ± 0.114 | 0.937 ± 0.102 |
| Spleen/TBW | 1.853 ± 0.275 | 1.885 ± 0.310 | 2.003 ± 0.198 | 1.892 ± 0.227 |
| Testes (g) | 3.882 ± 0.192 | 3.856 ± 0.193 | 3.905 ± 0.297 | 3.761 ± 0.168 |
| Testes/TBW | 8.044 ± 0.437 | 7.726 ± 0.467 | 7.705 ± 0.947 | 7.586 ± 0.342 |
| Thymus (g) | 0.324 ± 0.073 | 0.333 ± 0.093 | 0.315 ± 0.060 | 0.353 ± 0.061 |
| Thymus/TBW | 0.667 ± 0.134 | 0.668 ± 0.188 | 0.616 ± 0.108 | 0.709 ± 0.109 |
| Thyroid + PT | 0.040 ± 0.010 | 0.043 ± 0.009 | 0.043 ± 0.009 | 0.045 ± 0.013 |
| Thyroid + PT/TBW | 0.824 ± 0.236 | 0.855 ± 0.182 | 0.843 ± 0.169 | 0.914 ± 0.268 |
| Thyroid + PT/TBrW | 0.0180 ± 0.004 | 0.0195 ± 0.004 | 0.020 ± 0.004 | 0.021 ± 0.006 |
| Females | ||||
| Terminal body weight (g) | 249.6 ± 20.4 | 263.7 ± 27.7 | 270.9 ± 19.2 | 267.5 ± 20.5 |
| Adrenals (g) | 0.100 ± 0.010 | 0.093 ± 0.008 | 0.094 ± 0.011 | 0.101 ± 0.012 |
| Adrenals/TBW | 0.401 ± 0.050 | 0.356 ± 0.042 | 0.351 ± 0.048 | 0.381 ± 0.061 |
| Brain (g) | 1.996 ± 0.112 | 1.975 ± 0.065 | 2.001 ± 0.109 | 2.032 ± 0.101 |
| Brain/TBW | 8.026 ± 0.537 | 7.571 ± 0.900 | 7.414 ± 0.589 | 7.625 ± 0.533 |
| Heart (g) | 0.873 ± 0.090 | 0.875 ± 0.075 | 0.889 ± 0.059 | 0.919 ± 0.085 |
| Heart/TBW | 3.503 ± 0.286 | 3.330 ± 0.221 | 3.285 ± 0.141 | 3.441 ± 0.252 |
| Kidneys (g) | 1.827 ± 0.148 | 1.925 ± 0.190 | 2.017 ± 0.177 | 2.269 ± 0.302 *** |
| Kidneys/TBW | 7.347 ± 0.662 | 7.334 ± 0.717 | 7.451 ± 0.486 | 8.482 ± 0.866 ** |
| Kidneys/TBrW | 0.915 ± 0.050 | 0.975 ± 0.096 | 1.009 ± 0.084 | 1.114 ± 0.111 *** |
| Liver (g) | 7.037 ± 0.903 | 7.519 ± 0.818 | 7.614 ± 0.610 | 7.961 ± 0.870 |
| Liver/TBW | 28.132 ± 2.089 | 28.534 ± 1.538 | 28.153 ± 1.989 | 29.794 ± 2.744 |
| Ovaries (g) | 0.148 ± 0.030 | 0.137 ± 0.016 | 0.134 ± 0.020 | 0.135 ± 0.023 |
| Ovaries/TBW | 0.595 ± 0.128 | 0.521 ± 0.055 | 0.500 ± 0.093 | 0.502 ± 0.066 |
| Pituitary (g) | 0.020 ± 0.006 | 0.018 ± 0.003 | 0.018 ± 0.004 | 0.020 ± 0.005 |
| Pituitary/TBW | 0.008 ± 0.002 | 0.007 ± 0.001 | 0.007 ± 0.002 | 0.007 ± 0.002 |
| Spleen (g) | 0.560 ± 0.075 | 0.574 ± 0.091 | 0.656 ± 0.118 | 0.597 ± 0.067 |
| Spleen/TBW | 2.245 ± 0.252 | 2.172 ± 0.198 | 2.42 ± 0.381 | 2.231 ± 0.187 |
| Thymus (g) | 0.249 ± 0.075 | 0.218 ± 0.044 | 0.237 ± 0.071 | 0.229 ± 0.078 |
| Thymus/TBW | 0.987 ± 0.241 | 0.827 ± 0.136 | 0.878 ± 0.252 | 0.853 ± 0.264 |
| Thyroid+ PT | 0.048 ± 0.014 | 0.033 ± 0.008 ** | 0.036 ± 0.010 * | 0.033 ± 0.007 ** |
| Thyroid + PT/TBW | 1.946 ± 0.597 | 1.245 ± 0.260 ** | 1.349 ± 0.424 ** | 1.225 ± 0.286 ** |
| Thyroid + PT/TBrW | 0.024 ± 0.007 | 0.017 ± 0.004 ** | 0.018 ± 0.005 * | 0.016 ± 0.003 ** |
| Uterus (g) | 0.757 ± 0.363 | 0.632 ± 0.232 | 0.648 ± 0.285 | 0.647 ± 0.254 |
| Uterus/TBW | 2.988 ± 1.235 | 2.408 ± 0.874 | 2.403 ± 1.066 | 2.397 ± 0.833 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dolan, L.C.; Karikachery, A.R.; Thipe, V.C.; Arceneaux, B.G.; Katti, K.K.; Katti, K.V.; Chesne, A.M. Toxicity Investigations of (R)-3-Hydroxybutyrate Glycerides In Vitro and in Male and Female Rats. Nutrients 2022, 14, 4426. https://doi.org/10.3390/nu14204426
Dolan LC, Karikachery AR, Thipe VC, Arceneaux BG, Katti KK, Katti KV, Chesne AM. Toxicity Investigations of (R)-3-Hydroxybutyrate Glycerides In Vitro and in Male and Female Rats. Nutrients. 2022; 14(20):4426. https://doi.org/10.3390/nu14204426
Chicago/Turabian StyleDolan, Laurie C., Alice Raphael Karikachery, Velaphi C. Thipe, Benjamin G. Arceneaux, Kavita K. Katti, Kattesh V. Katti, and Alton M. Chesne. 2022. "Toxicity Investigations of (R)-3-Hydroxybutyrate Glycerides In Vitro and in Male and Female Rats" Nutrients 14, no. 20: 4426. https://doi.org/10.3390/nu14204426
APA StyleDolan, L. C., Karikachery, A. R., Thipe, V. C., Arceneaux, B. G., Katti, K. K., Katti, K. V., & Chesne, A. M. (2022). Toxicity Investigations of (R)-3-Hydroxybutyrate Glycerides In Vitro and in Male and Female Rats. Nutrients, 14(20), 4426. https://doi.org/10.3390/nu14204426