

Exploring Active Ingredients, Beneficial Effects, and Potential Mechanism of Allium tenuissimum L. Flower for Treating T2DM Mice Based on Network Pharmacology and Gut Microbiota


Abstract
:
1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. A. tenuissimum Flower Extract Preparation
2.3. Isolation, Characterization, and Quantitation
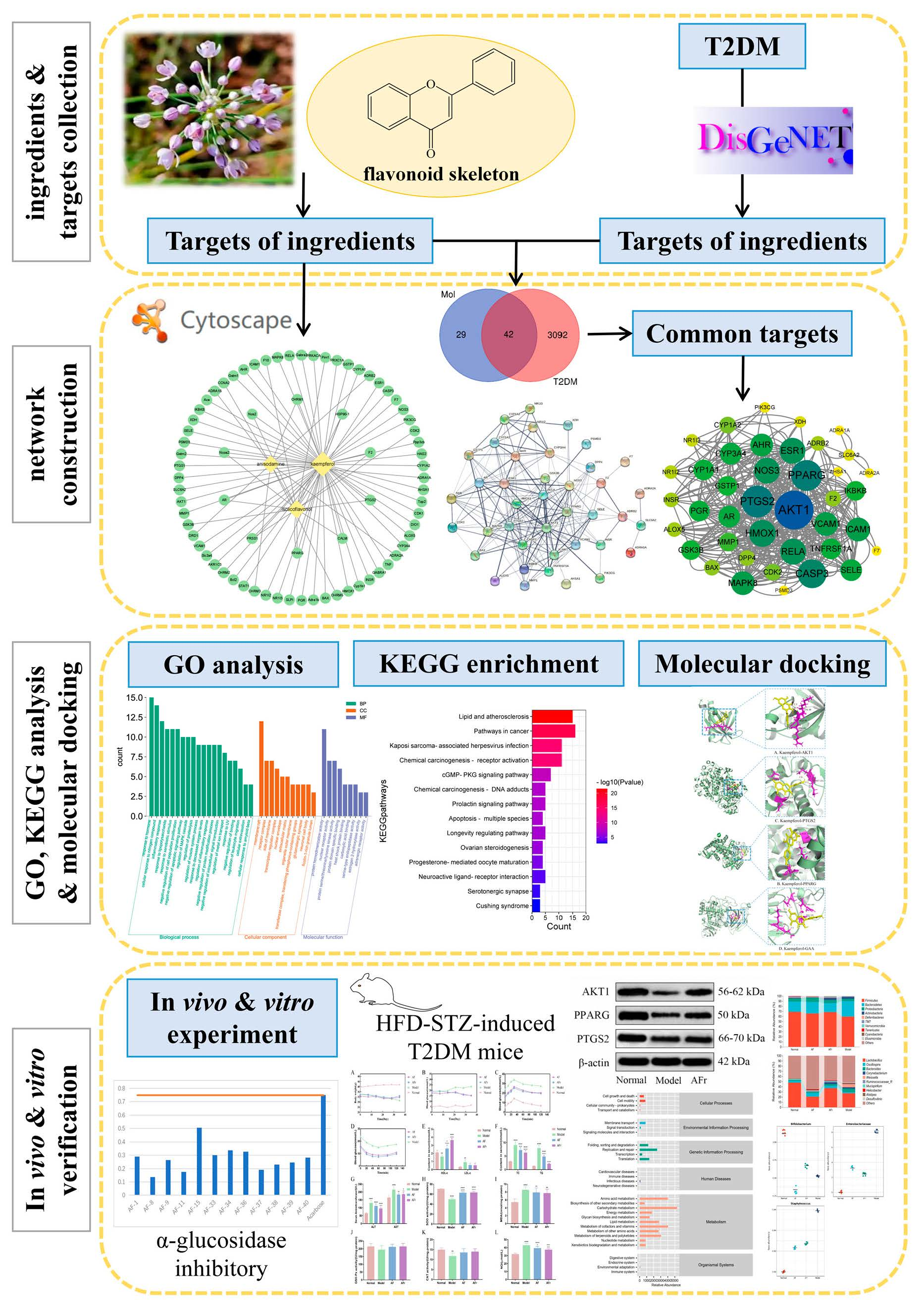
2.4. Network Pharmacology Predictive Analysis
2.4.1. Collecting and Analyzing Targets of Ingredients and T2DM
2.4.2. Protein−Protein Interaction Network Construction and Key Target Analysis
2.4.3. GO and KEGG Enrichment Analysis
2.4.4. Ingredient-Target Protein Molecular Docking
2.5. In Vitro α−Glucosidase Inhibitory Activity Assay
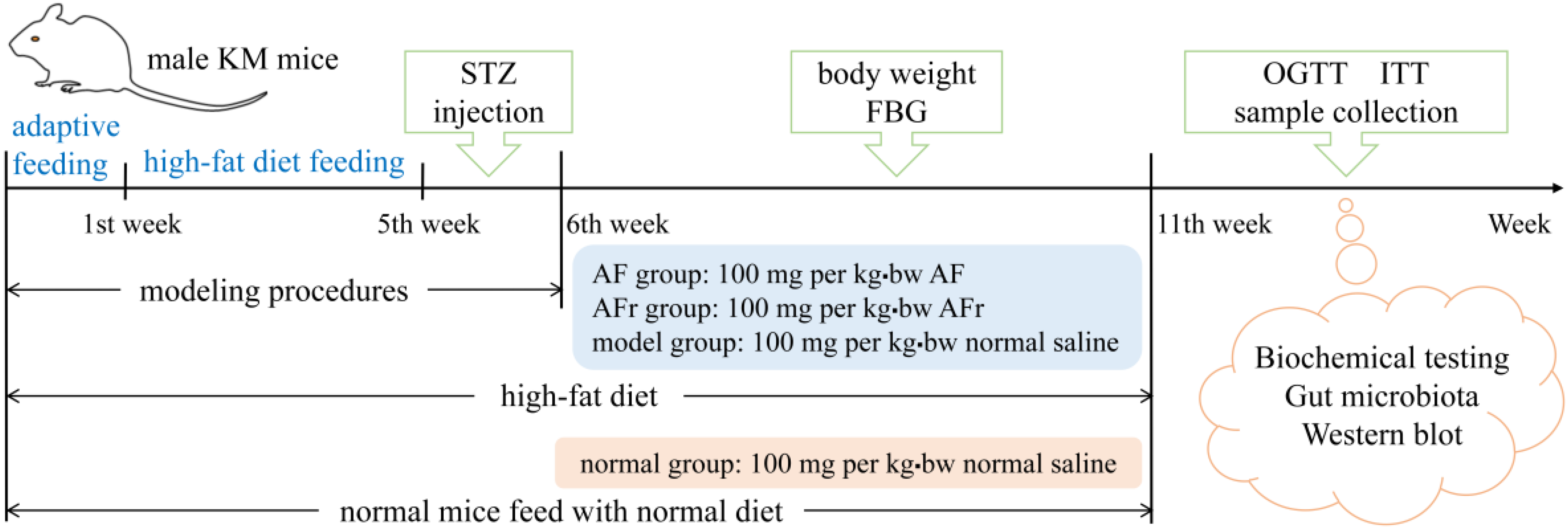
2.6. In Vivo Animal Experiments
2.7. Western Blot Analysis
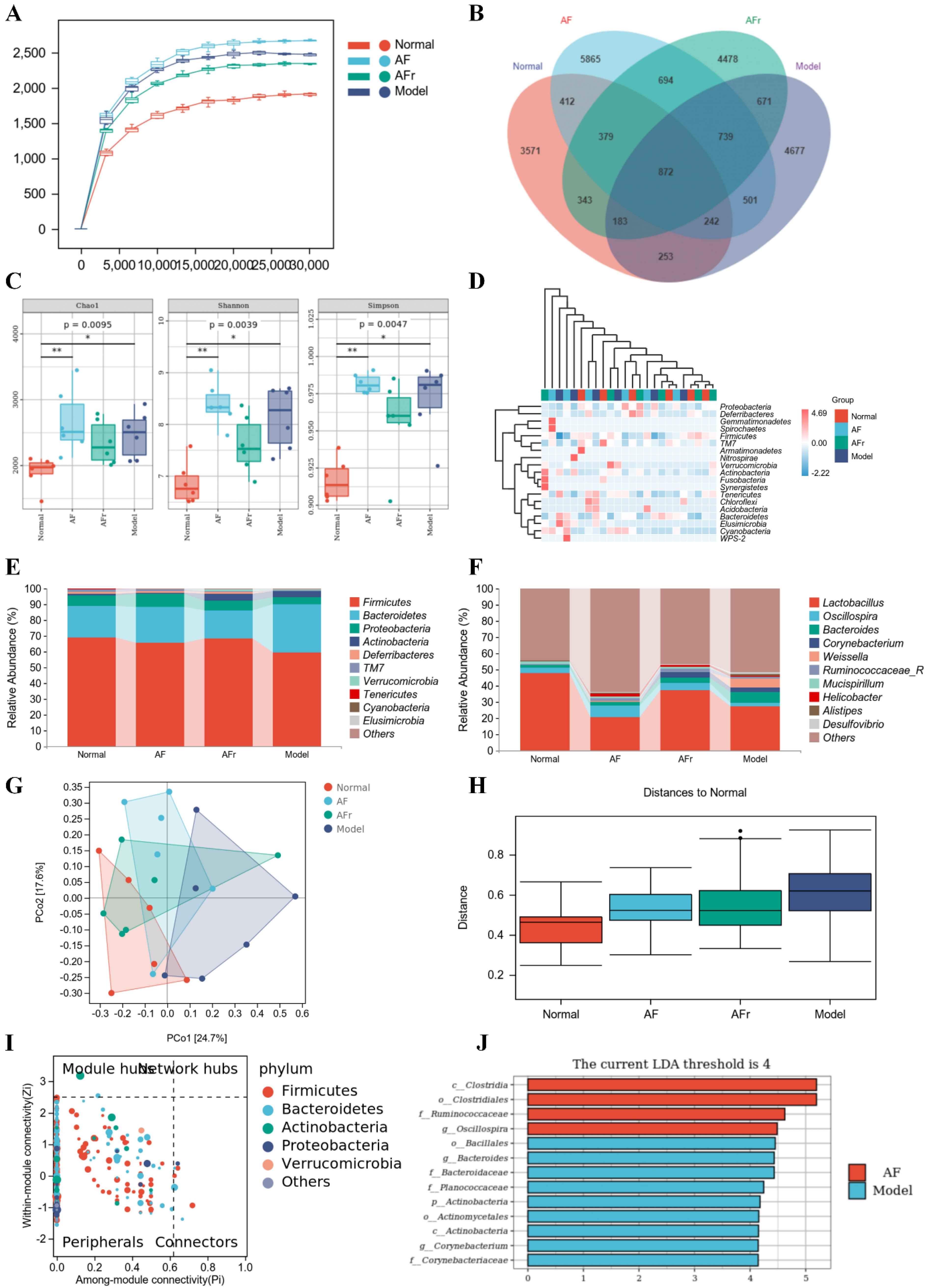
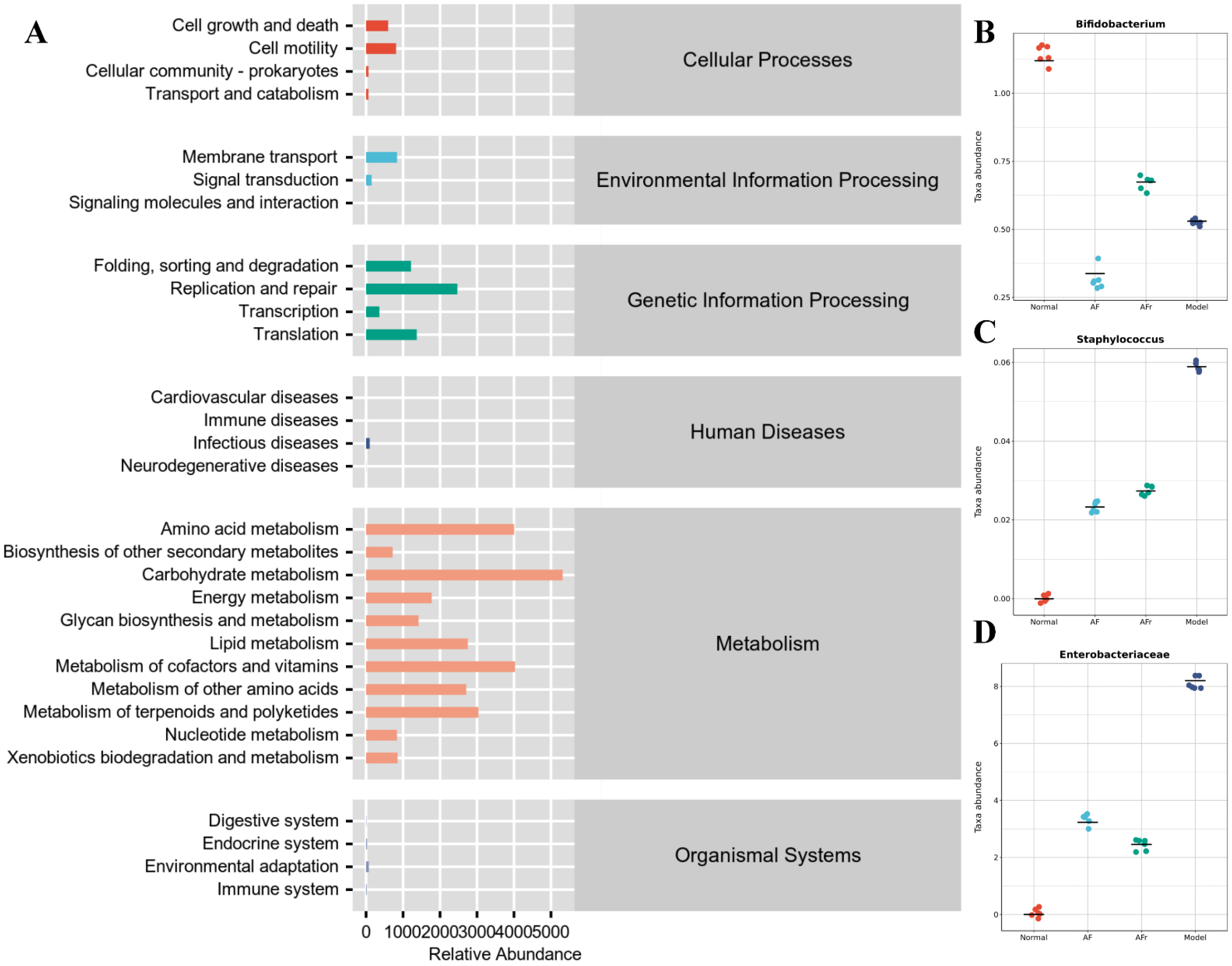
2.8. Gut Microbiota Analysis
2.9. Statistical Analysis
3. Results and Discussion
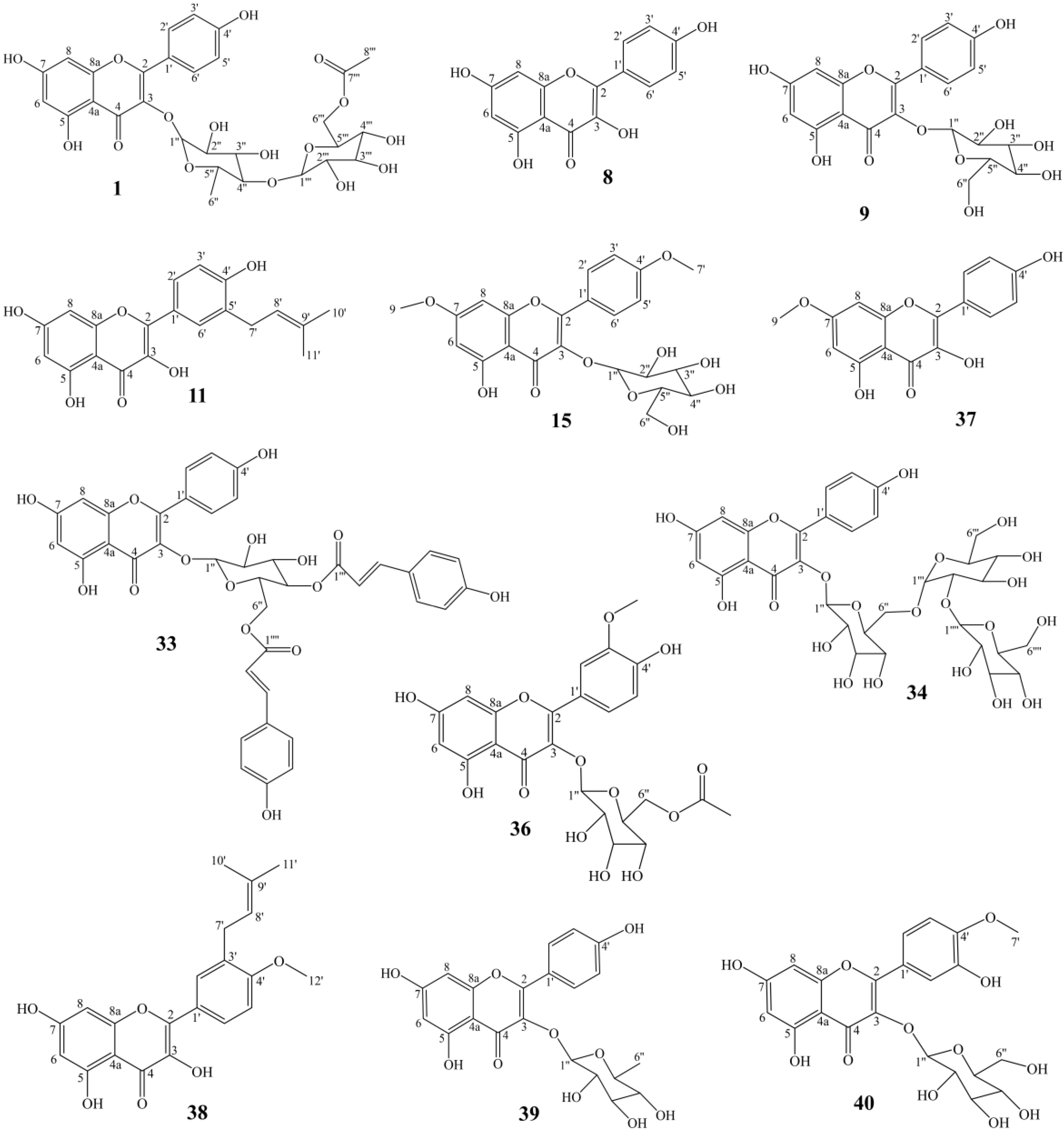
3.1. Identification and Quantitation of Compounds in A. tenuissimum Flower
3.2. Network Pharmacology Analysis
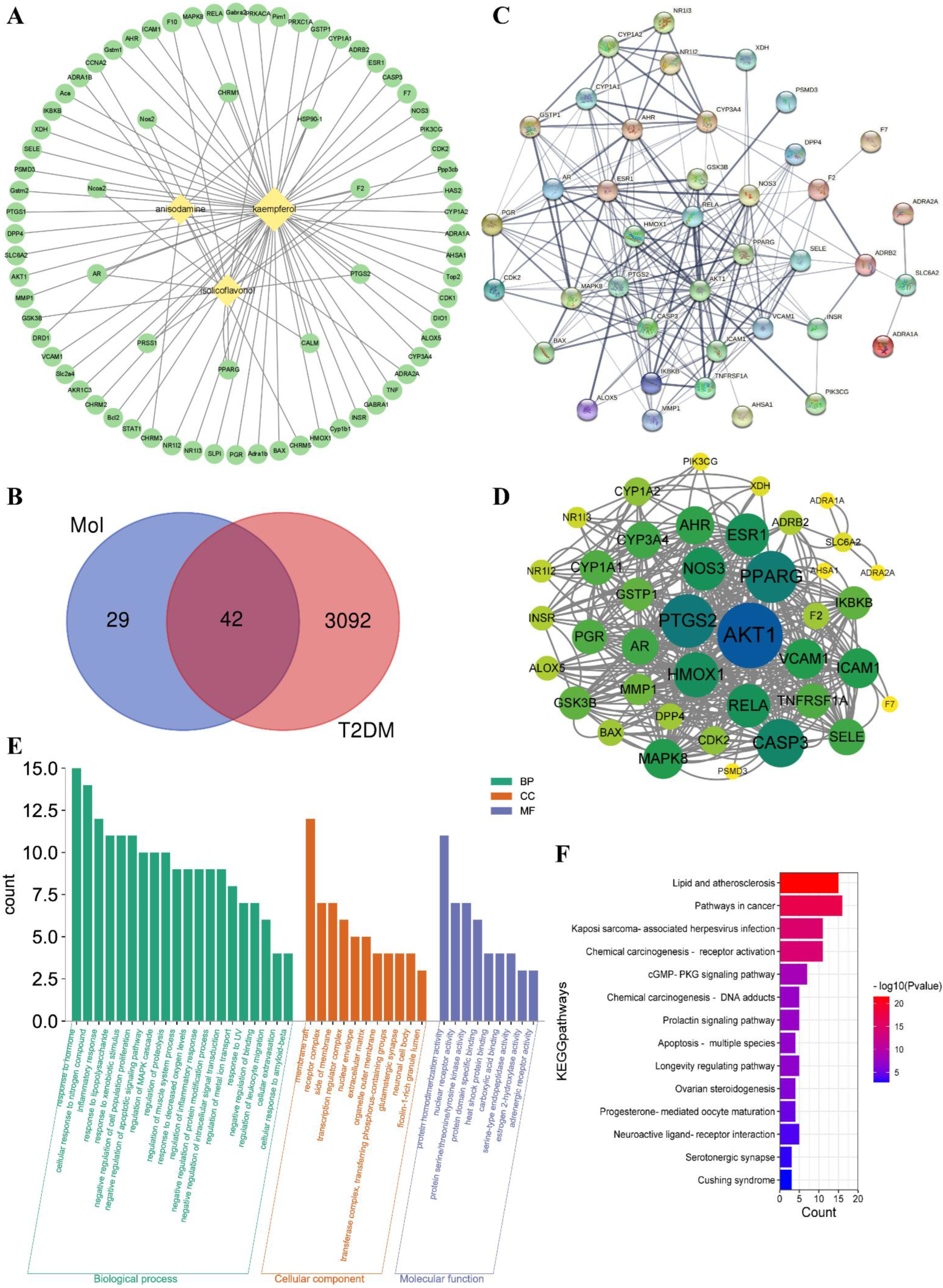
3.2.1. Potential Targets Analysis of Active Ingredients and T2DM
3.2.2. PPI Network Analysis
3.2.3. GO Analysis and KEGG Pathway Enrichment
3.2.4. Molecular Docking
3.3. α−Glucosidase Inhibitory Activity of Flavonoids
3.4. In Vivo Experiments Results
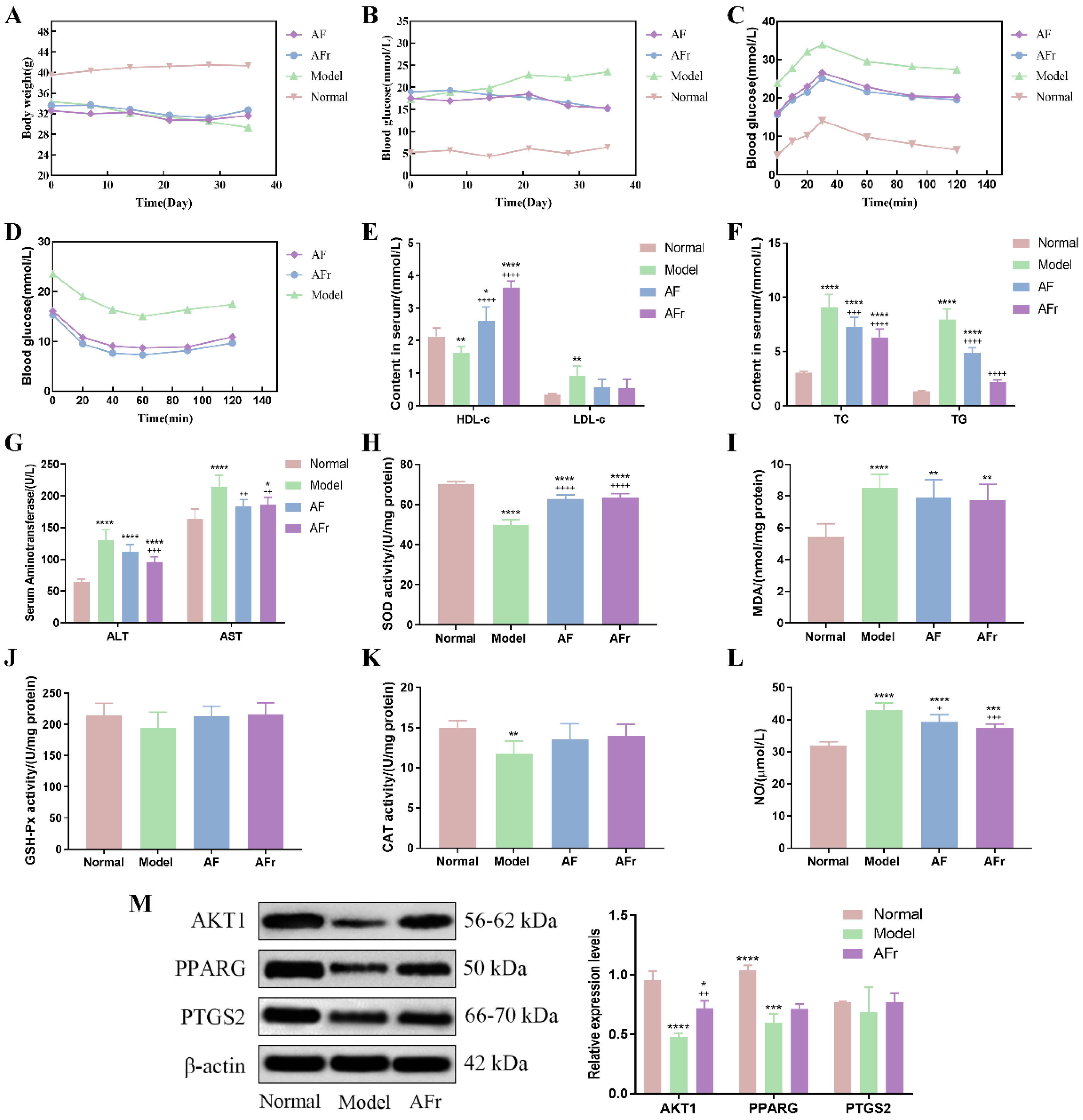
3.4.1. Effect on Body Weight, FBG, OGTT, and ITT of Diabetic Mice
3.4.2. Effect on Serum Biomarkers of Diabetic Mice
3.4.3. Western Blot Analysis
3.4.4. Gut Microbiota Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, Y.; Cao, Y.; Chen, J.; Qin, H.; Yang, L. A new possible mechanism by which punicalagin protects against liver injury induced by type 2 diabetes mellitus: Upregulation of autophagy via the Akt/FoxO3a signaling pathway. J. Agric. Food Chem. 2019, 67, 13948–13959. [Google Scholar] [CrossRef] [PubMed]
- Ma, G.; Chai, X.; Hou, G.; Zhao, F.; Meng, Q. Phytochemistry, bioactivities and future prospects of mulberry leaves: A review. Food Chem. 2022, 372, 131335. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Zhang, Y.; Peng, Y.; Li, X. The water extract of Radix scutellariae, its total flavonoids and baicalin inhibited CYP7A1 expression, improved bile acid, and glycolipid metabolism in T2DM mice. J. Ethnopharmacol. 2022, 293, 115238. [Google Scholar] [CrossRef] [PubMed]
- Duru, K.C.; Mukhlynina, E.A.; Moroz, G.A.; Gette, I.F.; Danilova, I.G.; Kovaleva, E.G. Anti-diabetic effect of isoflavone rich kudzu root extract in experimentally induced diabetic rats. J. Funct. Foods 2020, 68, 103922. [Google Scholar] [CrossRef]
- Kumar, A.; Sreedharan, S.; Kashyap, A.K.; Singh, P.; Ramchiary, N. A review on bioactive phytochemicals and ethnopharmacological potential of purslane (Portulaca oleracea L.). Heliyon 2022, 8, e08669. [Google Scholar] [CrossRef]
- Xiao, J. Recent advances in dietary flavonoids for management of type 2 diabetes. Curr. Opin. Food Sci. 2022, 44, 100806. [Google Scholar] [CrossRef]
- Shi, X.Q.; Yue, S.J.; Tang, Y.P.; Chen, Y.Y.; Zhou, G.S.; Zhang, J.; Zhu, Z.H.; Liu, P.; Duan, J.A. A network pharmacology approach to investigate the blood enriching mechanism of Danggui buxue Decoction. J. Ethnopharmacol. 2019, 235, 227–242. [Google Scholar] [CrossRef]
- Zhou, W.; Dai, Y.; Meng, J.; Wang, P.; Wu, Y.; Dai, L.; Zhang, M.; Yang, X.; Xu, S.; Sui, F.; et al. Network pharmacology integrated with molecular docking reveals the common experiment-validated antipyretic mechanism of bitter-cold herbs. J. Ethnopharmacol. 2021, 274, 114042. [Google Scholar] [CrossRef]
- Zhou, C.; Liu, L.J.; Zhuang, J.; Wei, J.Y.; Zhang, T.T.; Gao, C.; Liu, C.; Li, H.Y.; Si, H.Z.; Sun, C.G. A systems biology-based approach to uncovering molecular mechanisms underlying effects of traditional chinese medicine qingdai in chronic myelogenous leukemia, involving integration of network pharmacology and molecular docking technology. Med. Sci. Monit. 2018, 24, 4305–4316. [Google Scholar] [CrossRef]
- Luo, Q.; Shi, X.; Ding, J.; Ma, Z.; Chen, X.; Leng, Y.; Zhang, X.; Liu, Y. Network pharmacology integrated molecular docking reveals the Antiosteosarcoma Mechanism of Biochanin A. Evid. Based Complement. Altern. Med. 2019, 2019, 1–10. [Google Scholar]
- Zhang, S.S.; Guo, S.; Zheng, Z.J.; Liu, S.J.; Hou, Y.F.; Ho, C.T.; Bai, N.S. Characterization of volatiles in Allium tenuissimum L. flower by headspace-gas chromatography-olfactometry-mass spectrometry, odor activity values, and the omission and recombination experiments. LWT-Food Sci. Technol. 2021, 151, 112144. [Google Scholar] [CrossRef]
- Li, M.; Liu, Y.; Zhang, S.; Guo, C.; Li-gang, Y. Study on chemical constituents and antibacterial activity of ethanol extract from Allium tenuissimum flowers. China Condiment 2020, 45, 35–38. (In Chinese) [Google Scholar]
- Li, M.; Ding, P.; Li, R.; Zhang, S.; Guo, C. Antioxidant properties and inhibitory effect on nitrosation of extracts from Allium tenuissimum flowers. Food Res. Dev. 2019, 45, 46–51. (In Chinese) [Google Scholar]
- Li, M.; Hou, J.; Liu, Y.; Wei, L.; Guo, C. Study on antioxidation of ethanol extract from Allium tenuissimum L. flowers in vitro and its inhibition on α-glucosidase. China Condiment 2022, 47, 30–34. (In Chinese) [Google Scholar]
- Kang, M.G.; Yi, S.H.; Lee, J.S. Production and characterization of a new α-glucosidase inhibitory peptide from Aspergillus oryzae N159-1. Mycobiology 2013, 41, 149–154. [Google Scholar] [CrossRef]
- Osonoi, T.; Saito, M.; Mochizuki, K.; Fukaya, N.; Muramatsu, T.; Inoue, S.; Fuchigami, M.; Goda, T. The α-glucosidase inhibitor miglitol decreases glucose fluctuations and inflammatory cytokine gene expression in peripheral leukocytes of Japanese patients with type 2 diabetes mellitus. Metabolism 2010, 59, 1816–1822. [Google Scholar] [CrossRef]
- Jang, J.H.; Park, J.E.; Han, J.S. Scopoletin inhibits α-glucosidase in vitro and alleviates postprandial hyperglycemia in mice with diabetes. Eur. J. Pharmacol. 2018, 834, 152–156. [Google Scholar] [CrossRef]
- Hu, P.; Li, D.H.; Jia, C.C.; Liu, Q.; Wang, X.F.; Li, Z.L.; Hua, H.M. Bioactive constituents from Vitex negundo var. heterophylla and their antioxidant and α-glucosidase inhibitory activities. J. Funct. Foods 2017, 35, 236–244. [Google Scholar] [CrossRef]
- Zhang, Y.; Xiao, G.; Sun, L.; Wang, Y.; Wang, Y.; Wang, Y. A new flavan-3-ol lactone and other constituents from Euonymus alatus with inhibitory activities on a-glucosidase and differentiation of 3T3-L1 cells. Nat. Prod. Res. 2013, 27, 1513–1520. [Google Scholar] [CrossRef]
- Gou, S.H.; Liu, J.; He, M.; Qiang, Y.; Ni, J.M. Quantification and bio-assay of α-glucosidase inhibitors from the roots of Glycyrrhiza uralensis Fisch. Nat. Prod. Res. 2016, 30, 2130–2134. [Google Scholar] [CrossRef]
- Sun, J.; Zhang, F.; Yang, M.; Zhang, J.; Chen, L.; Zhan, R.; Li, L.; Chen, Y. Isolation of α-glucosidase inhibitors including a new flavonol glycoside from Dendrobium devonianum. Nat. Prod. Res. 2014, 28, 1900–1905. [Google Scholar] [CrossRef] [PubMed]
- Hong, D.F.; Hu, G.L.; Peng, X.R.; Wang, X.Y.; Wang, Y.B.; Al-Romaima, A.; Li, Z.R.; Qiu, M.H. Unusual ent-Kaurane Diterpenes from the coffea cultivar S288 coffee beans and molecular docking to α-glucosidase. J. Agric. Food Chem. 2022, 70, 615–625. [Google Scholar] [CrossRef] [PubMed]
- Xie, L.; Zhang, T.; Karrar, E.; Zheng, L.; Xie, D.; Jin, J.; Chang, M.; Wang, X.; Jin, Q. Insights into an α-glucosidase inhibitory profile of 4,4-dimethylsterols by multispectral techniques and molecular docking. J. Agric. Food Chem. 2021, 69, 15252–15260. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.S.; Zhang, N.N.; Guo, S.; Liu, S.J.; Hou, Y.F.; Li, S.; Ho, C.T.; Bai, N.S. Glycosides and flavonoids from the extract of Pueraria thomsonii Benth leaf alleviate type 2 diabetes in high-fat diet plus streptozotocin-induced mice by modulating the gut microbiota. Food Funct. 2022, 13, 3931–3945. [Google Scholar] [CrossRef]
- Cheng, N.; Chen, S.; Liu, X.; Zhao, H.; Cao, W. Impact of schisandrachinensis bee pollen on nonalcoholic fatty liver disease and gut microbiota in highfat diet induced obese mice. Nutrients 2019, 11, 346. [Google Scholar] [CrossRef]
- Han, L.; Song, J.; Yan, C.; Wang, C.; Wang, L.; Li, W.; Du, Y.; Li, Q.; Liang, T. Inhibitory activity and mechanism of calycosin and calycosin-7-O-β-D-glucoside on α-glucosidase: Spectroscopic and molecular docking analyses. Process Biochem. 2022, 118, 227–235. [Google Scholar] [CrossRef]
- Gu, Z.; Pan, M.; Liu, J.; Yang, M.; Zhang, W.; Mai, K. Molecular cloning of AKT1 and AKT2 and their divergent responses to insulin and glucose at transcriptional level in the liver of Japanese flounder Paralichthys olivaceus. Aquac. Rep. 2022, 23, 101066. [Google Scholar] [CrossRef]
- Zhang, L.; Han, L.; Ma, J.; Wu, T.; Wei, Y.; Zhao, L.; Tong, X. Exploring the synergistic and complementary effects of berberine and paeoniflorin in the treatment of type 2 diabetes mellitus by network pharmacology. Eur. J. Pharmacol. 2022, 919, 174769. [Google Scholar] [CrossRef]
- Verma, S.; Srivastava, N.; Banerjee, M. Genetic polymorphisms in TCF7L2 and PPARG genes and susceptibility to Type 2 diabetes mellitus. Meta Gene 2021, 28, 100864. [Google Scholar] [CrossRef]
- Lefterova, M.I.; Haakonsson, A.K.; Lazar, M.A.; Mandrup, S. PPARγ and the global map of adipogenesis and beyond. Trends Endocrinol. Metab. 2014, 25, 293–302. [Google Scholar] [CrossRef]
- Celi, F.S.; Shuldiner, A.R. The role of peroxisome proliferator-activated receptor gamma in diabetes and obesity. Curr. Diabetes Rep. 2002, 2, 179–185. [Google Scholar] [CrossRef]
- Horiki, M.; Ikegami, H.; Fujisawa, T.; Kawabata, Y.; Ono, M.; Nishino, M.; Shimamoto, K.; Ogihara, T. Association of Pro12Ala polymorphism of PPARγ gene with insulin resistance and related diseases. Diabetes Res. Clin. Pract. 2004, 66, S63–S67. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.; Lv, X.; Feng, Y.; Li, H.; Chen, C.; Lin, H.; Li, H.; Wang, C.; Chen, J.; Sun, J. Study on the effect of active components of Schisandra chinensis on liver injury and its mechanisms in mice based on network pharmacology. Eur. J. Pharmacol. 2021, 910, 174442. [Google Scholar] [CrossRef]
- El-Baz, A.M.; Shata, A.; Hassan, H.M.; El-Sokkary, M.M.A.; Khodir, A.E. The therapeutic role of Lactobacillus and montelukast in combination with metformin in diabetes mellitus complications through modulation of gut microbiota and suppression of oxidative stress. Int. Immunopharmacol. 2021, 96, 107757. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Zhang, B.; Hu, J.; Nie, S.; Xiong, T.; Xie, M. Intervention of five strains of Lactobacillus on obesity in mice induced by high-fat diet. J. Funct. Foods 2020, 72, 104078. [Google Scholar] [CrossRef]
- Wu, T.; Zhang, Y.; Li, W.; Zhao, Y.; Long, H.; Muhindo, E.M.; Liu, R.; Sui, W.; Li, Q.; Zhang, M. Lactobacillus rhamnosus LRa05 ameliorate hyperglycemia through a regulating glucagon-mediated signaling pathway and gut microbiota in type 2 diabetic mice. J. Agric. Food Chem. 2021, 69, 8797–8806. [Google Scholar] [CrossRef]
- Konikoff, T.; Gophna, U. Oscillospira: A central, enigmatic component of the human gut microbiota. Trends Microbiol. 2016, 24, 523–524. [Google Scholar] [CrossRef]
- Chaudhari, S.S.; Chauhan, H.C.; Sharma, K.K.; Patel, S.S.; Patel, A.C.; Mohapatra, S.K.; Srimali, M.D.; Chandel, B. Antibiotic susceptibility pattern of canine coagulase positive and coagulase negative Staphylococcus spp. in a hot and dry region of India. Top. Companion Anim. Med. 2022, 50, 100679. [Google Scholar] [CrossRef]
- Kilonzo-Nthenge, A.; Liu, S.; Hashem, F.; Millner, P.; Githua, S. Prevalence of Enterobacteriaceae on fresh produce and food safety practices in small-acreage farms in Tennessee, USA. J. Consum. Prot. Food Saf. 2018, 13, 279–287. [Google Scholar] [CrossRef]
- Palmer, J.D.; Mortzfeld, B.M.; Piattelli, E.; Silby, M.W.; McCormick, B.A.; Bucci, V. Microcin H47: A Class IIb microcin with potent activity against multidrug resistant Enterobacteriaceae. ACS Infect. Dis. 2020, 6, 672–679. [Google Scholar] [CrossRef]
- Derrien, M.; Turroni, F.; Ventura, M.; van Sinderen, D. Insights into endogenous Bifidobacterium species in the human gut microbiota during adulthood. Trends Microbiol. 2022, 30, 940–947. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Li, T.; Chen, G.; Chen, D.; Peng, Y.; Hu, B.; Sun, Y.; Zeng, X. Commensal relationship of three Bifidobacterial Species leads to increase of Bifidobacterium in Vitro Fermentation of sialylated immunoglobulin G by human gut microbiota. J. Agric. Food Chem. 2020, 68, 9110–9119. [Google Scholar] [CrossRef] [PubMed]
- Reagan-Shaw, S.; Nihal, M.; Ahmad, N. Dose translation from animal to human studies revisited. FASEB J. 2008, 22, 659–661. [Google Scholar] [CrossRef] [PubMed]
- Li, M.X.; Xie, J.; Bai, X.; Du, Z.Z. Anti-aging potential, anti-tyrosinase and antibacterial activities of extracts and compounds isolated from Rosa chinensis cv. ‘JinBian’. Ind. Crops Prod. 2021, 159, 113059. [Google Scholar] [CrossRef]
- Zhang, W.Z.; Li, X.L.; Shi, L.G.; Wang, J.L.; Zhao, M.; Zhao, D.F.; Zhang, S.J. Sesquiterpene lactones from Ixeris sonchifolia (Bge.) Hance II. J. Asian Nat. Prod. Res. 2008, 10, 1087–1091. [Google Scholar] [CrossRef] [PubMed]
- Nagashima, F.; Suzuki, M.; Asakawa, Y. seco-cuparane-type sesquiterpenoid from the Japanese liverwort Jungermannia infusca. Phytochemistry 2001, 56, 807–810. [Google Scholar] [CrossRef]
- Parmar, V.S.; Bisht, K.S.; Malhotra, A.; Jha, A.; Errington, W.; Howarth, O.W.; Tyagi, O.D.; Stein, P.C.; Jensen, S.; Boll, P.M.; et al. A benzoic acid ester from uvaria narum. Phytochemistry 1995, 38, 951–955. [Google Scholar] [CrossRef]
- Erb, B.; Borschberg, H.J.; Arigoni, D. The structure of laricinolic acid and its biomimetic transformation into officinalic acid. J. Chem. Soc. Perkin Trans. 1 2000, 31, 2307–2309. [Google Scholar] [CrossRef]
- Samoylenko, V.; Rahman, M.M.; Tekwani, B.L.; Tripathi, L.M.; Wang, Y.H.; Khan, S.I.; Khan, I.A.; Miller, L.S.; Joshi, V.C.; Muhammad, I. Banisteriopsis caapi, a unique combination of MAO inhibitory and antioxidative constituents for the activities relevant to neurodegenerative disorders and Parkinson’s disease. J. Ethnopharmacol. 2010, 127, 357–367. [Google Scholar] [CrossRef]
- Paridhavi, M.; Agrawal, S.S. Isolation and characterization of flowers of Rosa damascena. Asian J. Chem. 2007, 19, 2751–2756. [Google Scholar]
- Da Liu, C.; Chen, J.; Wang, J.H. A novel kaempferol triglycoside from flower buds of Panax quinquefolium. Chem. Nat. Compd. 2009, 45, 808–810. [Google Scholar] [CrossRef]
- Rezende, F.M.; Ferreira, M.J.P.; Clausen, M.H.; Rossi, M.; Furlan, C.M. Acylated flavonoid glycosides are the main pigments that determine the flower colour of the brazilian native tree Tibouchina pulchra (Cham.) Cogn. Molecules 2019, 24, 718. [Google Scholar] [CrossRef] [PubMed]
- Tewari, A.; Bhakuni, R.S. Terpenoid and lipid constituents from Artemisia annua. Indian J. Chem.-Sect. B Org. Med. Chem. 2003, 42, 1782–1785. [Google Scholar] [CrossRef]
- Wang, C.Z.; Yu, D.Q. Lignan and acetylenic glycosides from Aster auriculatus. Phytochemistry 1998, 48, 711–717. [Google Scholar] [CrossRef]
- Hyun, S.K.; Jung, H.A.; Min, B.S.; Jung, J.H.; Choi, J.S. Isolation of phenolics, nucleosides, saccharides and an alkaloid from the root of Aralia cordata. Nat. Prod. Sci. 2010, 16, 20–25. [Google Scholar]
- Nycz, J.E.; Malecki, G.; Morag, M.; Nowak, G.; Ponikiewski, L.; Kusz, J.; Switlicka, A. Arbutin: Isolation, X-ray structure and computional studies. J. Mol. Struct. 2010, 980, 13–17. [Google Scholar] [CrossRef]
- Wei-li, Y.; Jun, T.; Liu-sheng, D. Chemical Constituents of Gymnotheca involucrata Pei. China J. Chinese Mater. Medica 2001, 26, 43–44. [Google Scholar]
- Zhou, Y.Z.; Ma, H.Y.; Chen, H.; Qiao, L.; Yao, Y.; Cao, J.Q.; Pei, Y.H. New acetylenic glucosides from Carthamus tinctorius. Chem. Pharm. Bull. 2006, 54, 1455–1456. [Google Scholar] [CrossRef]
- Ding, Z.; Liu, Y.; Ruan, J.; Yang, S.; Yu, H.; Chen, M.; Zhang, Y.; Wang, T. Bioactive constituents from the whole plants of Gentianella acuta (Michx.) Hulten. Molecules 2017, 22, 1309. [Google Scholar] [CrossRef]
- Chun-xia, X.; Yan, X.; Cai-xia, D.; Chun-mao, Y.; Lie-jun, H.; Wei, C.; Xiao-jiang, H. Chemical Constituents from Water-soluble Parts of Baphicacanthus cusia. J. Chinese Med. Mater. 2020, 43, 596–601. [Google Scholar]
- Abou-Hussein, D.R.; Badr, J.M.; Youssef, D.T.A. Dragmacidoside: A new nucleoside from the red sea sponge dragmacidon coccinea. Nat. Prod. Res. 2014, 28, 1134–1141. [Google Scholar] [CrossRef] [PubMed]
- Mori, T.; Yamashita, T.; Furihata, K.; Nagai, K.; Suzuki, K.I.; Hayakawa, Y.; Shin-ya, K. Burkholone, a new cytotoxic antibiotic against IGF-I dependent cells from Burkholderia sp. J. Antibiot. (Tokyo) 2007, 60, 713–716. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Zhang, W.; Ji, Y.P.; Zhao, Y.; Wang, C.G.; Hu, J.F. Gynostemosides A-E, megastigmane glycosides from Gynostemma pentaphyllum. Phytochemistry 2010, 71, 693–700. [Google Scholar] [CrossRef]
- Rovirosa, J.; Sepulveda, M.; Quezada, E.; San-Martin, A. Isoepitaondiol, a diterpenoid of Stypopodium flabelliforme and the insecticidal activity of stypotriol, epitaondiol and derivatives. Phytochemistry 1992, 31, 2679–2681. [Google Scholar] [CrossRef]
- Zang, E.H.; Chen, Z.W.; Zhang, C.H.; Li, M.H. Chemical constituents of Physochlaina physaloides (L.) G. Don (Solanaceae). Biochem. Syst. Ecol. 2021, 98, 104332. [Google Scholar] [CrossRef]
- Sugimoto, S.; Yamano, Y.; Desoukey, S.Y.; Katakawa, K.; Wanas, A.S.; Otsuka, H.; Matsunami, K. Isolation of Sesquiterpene-Amino Acid Conjugates, Onopornoids A-D, and a Flavonoid Glucoside from Onopordum alexandrinum. J. Nat. Prod. 2019, 82, 1471–1477. [Google Scholar] [CrossRef]
- Costa, M.; Fernández, R.; Pérez, M.; Thorsteinsdottir, M. Two new spongian diterpene analogues isolated from the marine sponge Acanthodendrilla sp. Nat. Prod. Res. 2020, 34, 1053–1060. [Google Scholar] [CrossRef]
- León, E.I.; Martín, A.; Peréz-Martín, I.; Quintanal, L.M.; Suárez, E. Hydrogen atom transfer experiments provide chemical evidence for the conformational differences between C-and O-disaccharides. European J. Org. Chem. 2010, 2010, 5248–5262. [Google Scholar] [CrossRef]
- Kitajima, J.; Komori, T.; Kawasaki, T.; Schulten, H. rolf Basic steroid saponins from aerial parts of Fritillaria thunbergii. Phytochemistry 1982, 21, 187–192. [Google Scholar] [CrossRef]
- Liu, H.; Liu, S.; Guo, L.; Zhang, Y.; Cui, L.; Ding, G. New furanones from the plant endophytic fungus Pestalotiopsis besseyi. Molecules 2012, 17, 14015–14021. [Google Scholar] [CrossRef]
- El-Gamal, A.A.; Al-Massarani, S.M.; Shaala, L.A.; Alahdald, A.M.; Al-Said, M.S.; Ashour, A.E.; Kumar, A.; Abdel-Kader, M.S.; Abdel-Mageed, W.M.; Youssef, D.T.A. Cytotoxic compounds from the Saudi red sea sponge Xestospongia testudinaria. Mar. Drugs 2016, 14, 82. [Google Scholar] [CrossRef] [PubMed]
- Sun, F.; Zhang, L.; Tian, J.K.; Cheng, J.Y.; Xiao, P.G. Studies on Chemical Constituents of Clematis terniflora. Chin. Pharm. J. 2007, 42, 102–103. [Google Scholar]
- Huang, X.A.; Yang, R.Z. A new hydroquinone diglucoside from Lysimachia fordiana. Chem. Nat. Compd. 2004, 40, 457–459. [Google Scholar] [CrossRef]
- Gao, Q.; Cheng, Y. A New Lactam from Pinellia ternata. Nat. Prod. Res. Dev. 2015, 27, 1693–1696. [Google Scholar]
- Yang, C.P.; Shie, P.H.; Huang, G.J.; Chien, S.C.; Kuo, Y.H. New anti-inflammatory flavonol glycosides from Lindera akoensis hayata. Molecules 2019, 24, 563. [Google Scholar] [CrossRef]
- Lin, Q.G.; Jie, O.Y.; De Yun, K.; Shan, D.S. Studies on Chemical Condtituents of Primula maximowiczii Regel II. Chin. Pharm. J. 2008, 43, 1300–1304. [Google Scholar]
- Zhang, Z.; Wang, D.; Zhao, Y.; Gao, H.; Hu, Y.H.; Hu, J.F. Fructose-derived carbohydrates from Alisma orientalis. Nat. Prod. Res. 2009, 23, 1013–1020. [Google Scholar] [CrossRef]
- Gudej, J.; Nazaruk, J. Flavonol glycosides from the flowers of Bellis perennis. Fitoterapia 2001, 72, 839–840. [Google Scholar] [CrossRef]
- Johnst, P.I.M.; Barrientos, R.E.; Simirgiotis, M.J.; Palacios, J. Characterization of Polyphenol Compounds from Endothelium-Dependent Vascular Relaxation Effect in Rat Aorta. Molecules 2020, 25, 3105. [Google Scholar]
- Pailee, P.; Sangpetsiripan, S.; Mahidol, C.; Ruchirawat, S.; Prachyawarakorn, V. Cytotoxic and cancer chemopreventive properties of prenylated stilbenoids from Macaranga siamensis. Tetrahedron 2015, 71, 5562–5571. [Google Scholar] [CrossRef]
- Duan, Y.; Hu, Y.; Yang, W.; Xiong, Y.; Du, C.; Yuan, C.; Hao, X.; Gu, W. Study on chemical constituents and α-glucosidase inhibitory activity of Cyclocarya paliurus in Guizhou province. Nat. Prod. Res. Dev. 2019, 31, 940–945. [Google Scholar]
- Kavtaradze, N.; Alaniya, M.; Masullo, M.; Cerulli, A.; Piacente, S. New Flavone Glycosides from Astragalus tanae Endemic to Georgia. Chem. Nat. Compd. 2020, 56, 70–74. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Calibration Curves 1 | R2 | LOD 2 (μg/mL) | LOQ 3 (μg/mL) | Content in AFr (μg/g) | Content in AF (μg/g) |
|---|---|---|---|---|---|---|
| AF−1 | y = 118.4x + 1441.9 | 0.9995 | 0.15 | 0.40 | 211.1 | 105.1 |
| AF−8 | y = 83.8x − 148.3 | 0.9999 | 0.05 | 0.25 | 284.1 | 59.2 |
| AF−9 | y = 185.1x − 177.3 | 0.9994 | 0.25 | 1.50 | 99.4 | 10.5 |
| AF−11 | y = 120.3x − 369.5 | 0.9997 | 0.50 | 1.25 | 188.8 | 26.8 |
| AF−15 | y = 279.6x − 226.0 | 0.9997 | 0.20 | 1.30 | 15.7 | 8.7 |
| AF−33 | y = 104.5x − 241.8 | 0.9998 | 0.15 | 0.70 | 196.3 | 54.7 |
| AF−34 | y = 352.5x − 292.2 | 0.9992 | 0.10 | 1.55 | 26.2 | 21.4 |
| AF−36 | y = 77.6x − 46.5 | 0.9997 | 0.45 | 2.75 | 143.7 | 37.3 |
| AF−37 | y = 57.2x + 121.7 | 0.9998 | 0.15 | 1.70 | 15.1 | 7.6 |
| AF−38 | y = 33.3x + 579.1 | 0.9998 | 0.20 | 1.50 | 171.6 | 21.1 |
| AF−39 | y = 304.6x + 280.8 | 1.0000 | 0.10 | 1.20 | 19.6 | 8.9 |
| AF−40 | y = 243.0x + 294.2 | 0.9995 | 0.05 | 0.50 | 57.9 | 40.7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, S.-S.; Hou, Y.-F.; Liu, S.-J.; Guo, S.; Ho, C.-T.; Bai, N.-S. Exploring Active Ingredients, Beneficial Effects, and Potential Mechanism of Allium tenuissimum L. Flower for Treating T2DM Mice Based on Network Pharmacology and Gut Microbiota. Nutrients 2022, 14, 3980. https://doi.org/10.3390/nu14193980
Zhang S-S, Hou Y-F, Liu S-J, Guo S, Ho C-T, Bai N-S. Exploring Active Ingredients, Beneficial Effects, and Potential Mechanism of Allium tenuissimum L. Flower for Treating T2DM Mice Based on Network Pharmacology and Gut Microbiota. Nutrients. 2022; 14(19):3980. https://doi.org/10.3390/nu14193980
Chicago/Turabian StyleZhang, Shan-Shan, Yu-Fei Hou, Shao-Jing Liu, Sen Guo, Chi-Tang Ho, and Nai-Sheng Bai. 2022. "Exploring Active Ingredients, Beneficial Effects, and Potential Mechanism of Allium tenuissimum L. Flower for Treating T2DM Mice Based on Network Pharmacology and Gut Microbiota" Nutrients 14, no. 19: 3980. https://doi.org/10.3390/nu14193980
APA StyleZhang, S.-S., Hou, Y.-F., Liu, S.-J., Guo, S., Ho, C.-T., & Bai, N.-S. (2022). Exploring Active Ingredients, Beneficial Effects, and Potential Mechanism of Allium tenuissimum L. Flower for Treating T2DM Mice Based on Network Pharmacology and Gut Microbiota. Nutrients, 14(19), 3980. https://doi.org/10.3390/nu14193980