

Fluctuations in Intestinal Microbiota Following Ingestion of Natto Powder Containing Bacillus subtilis var. natto SONOMONO Spores: Considerations Using a Large-Scale Intestinal Microflora Database

 , , , ,
, , , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. Study Design
2.3. Test Product
2.4. Stool Sample Collection
2.5. DNA Extraction
2.6. DNA Sequencing
2.7. 16S rRNA Data Analysis
2.8. Statistical Analysis
2.9. Large Japanese Gut Microbiota Database
3. Results
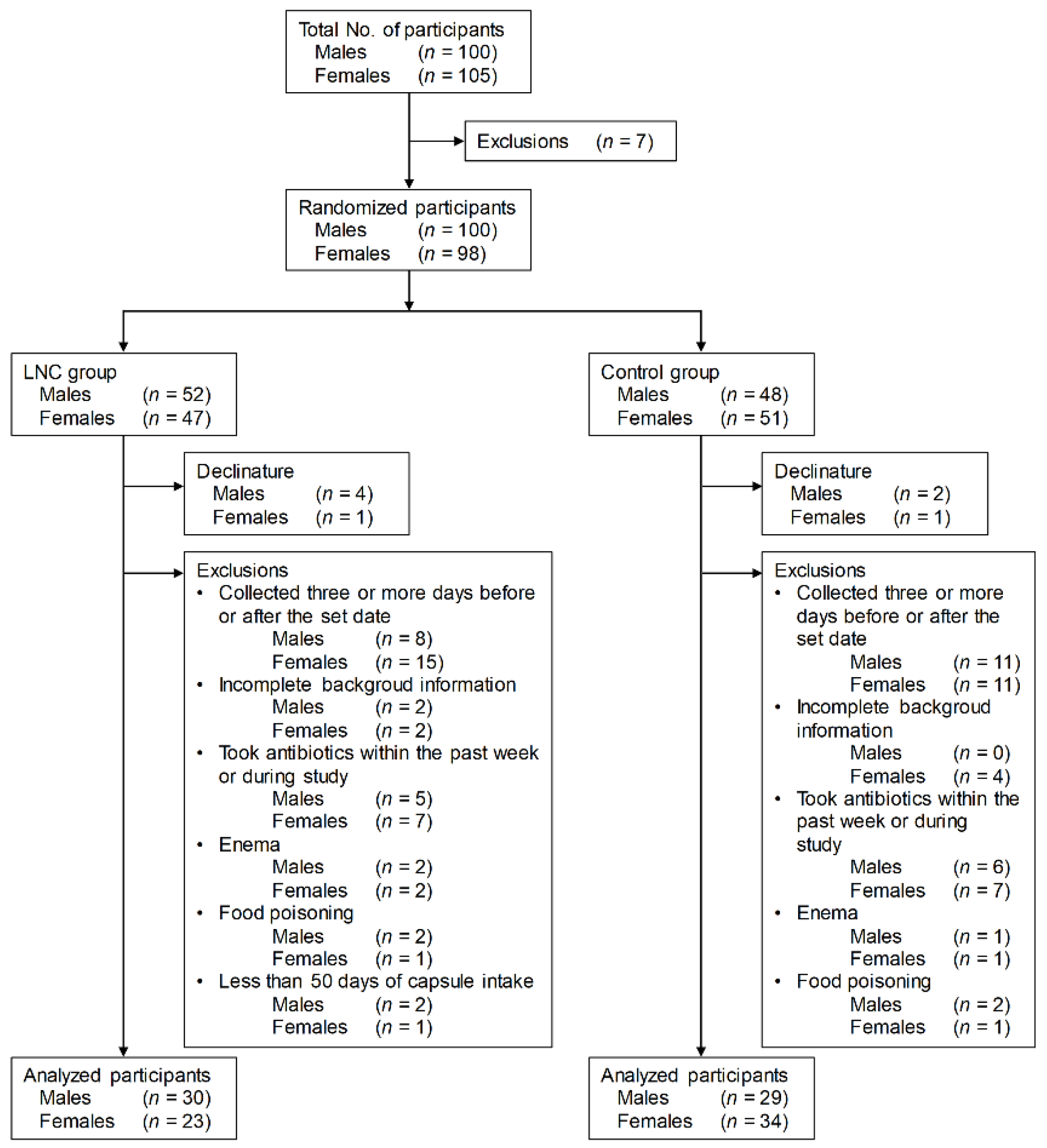
3.1. Selection of Analysis Population
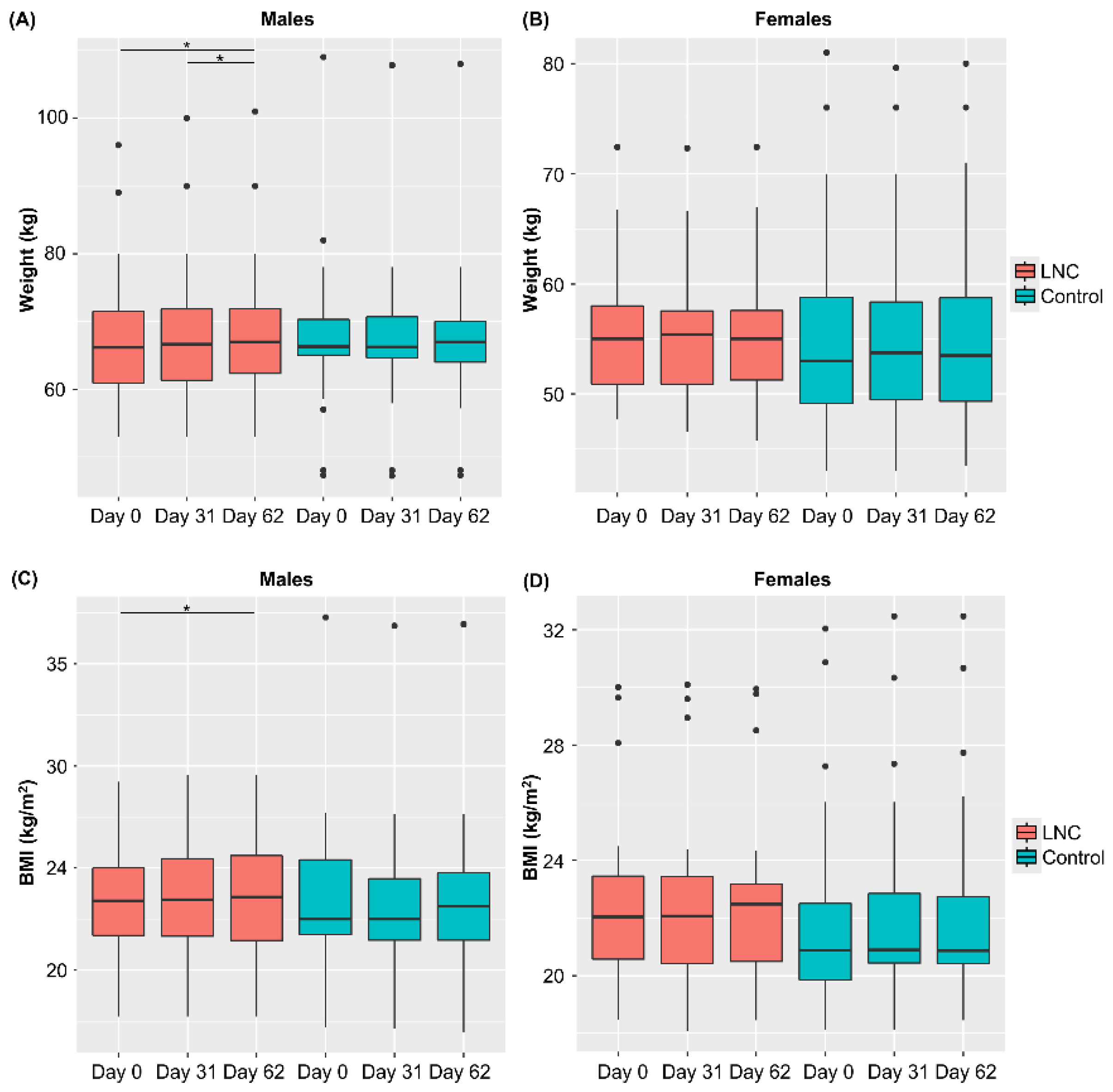
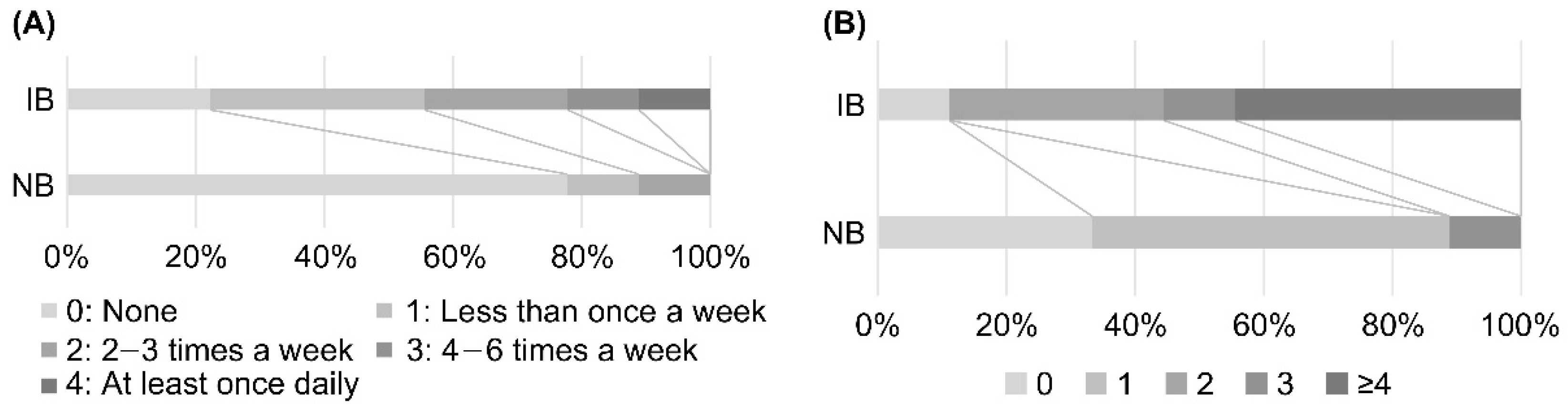
3.2. Variations in Weight, BMI, and Defecation Status
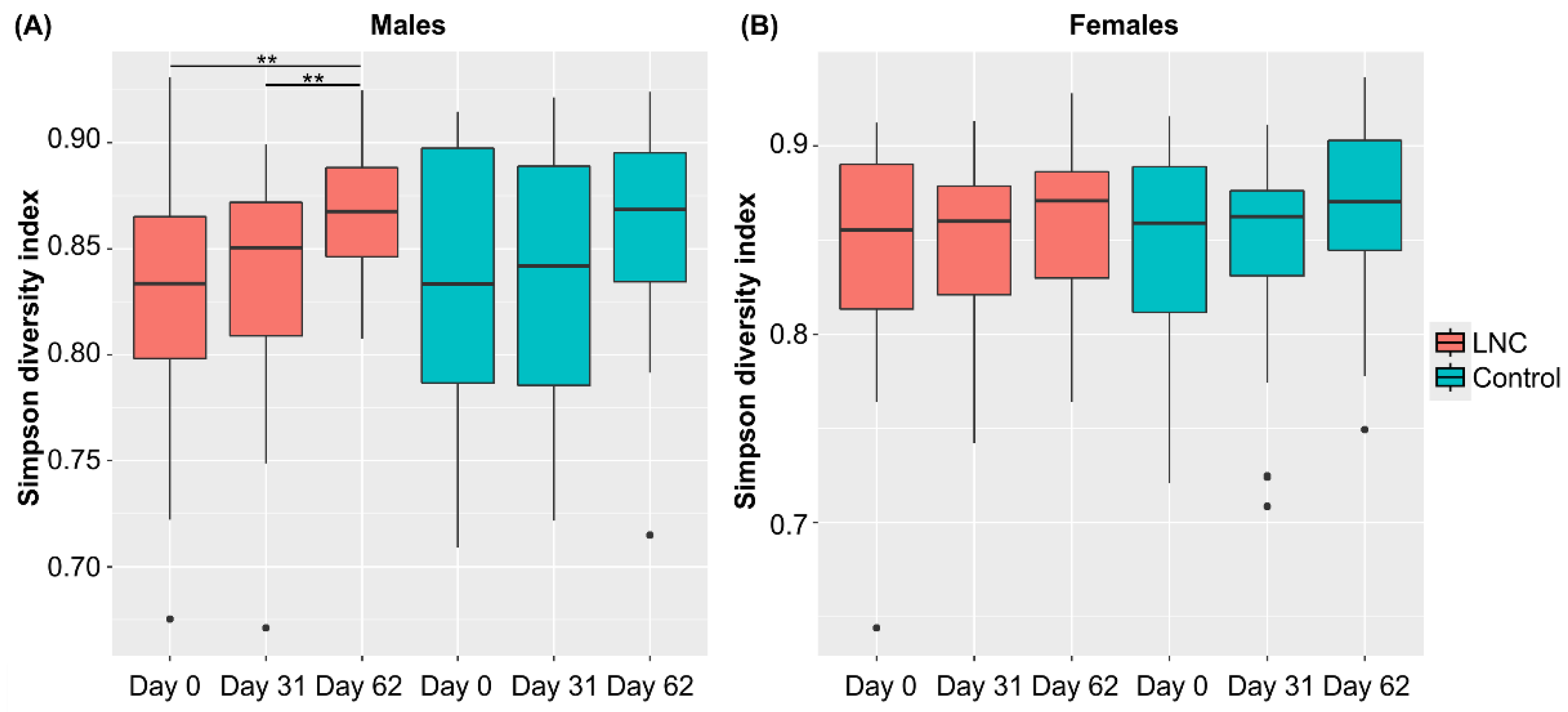
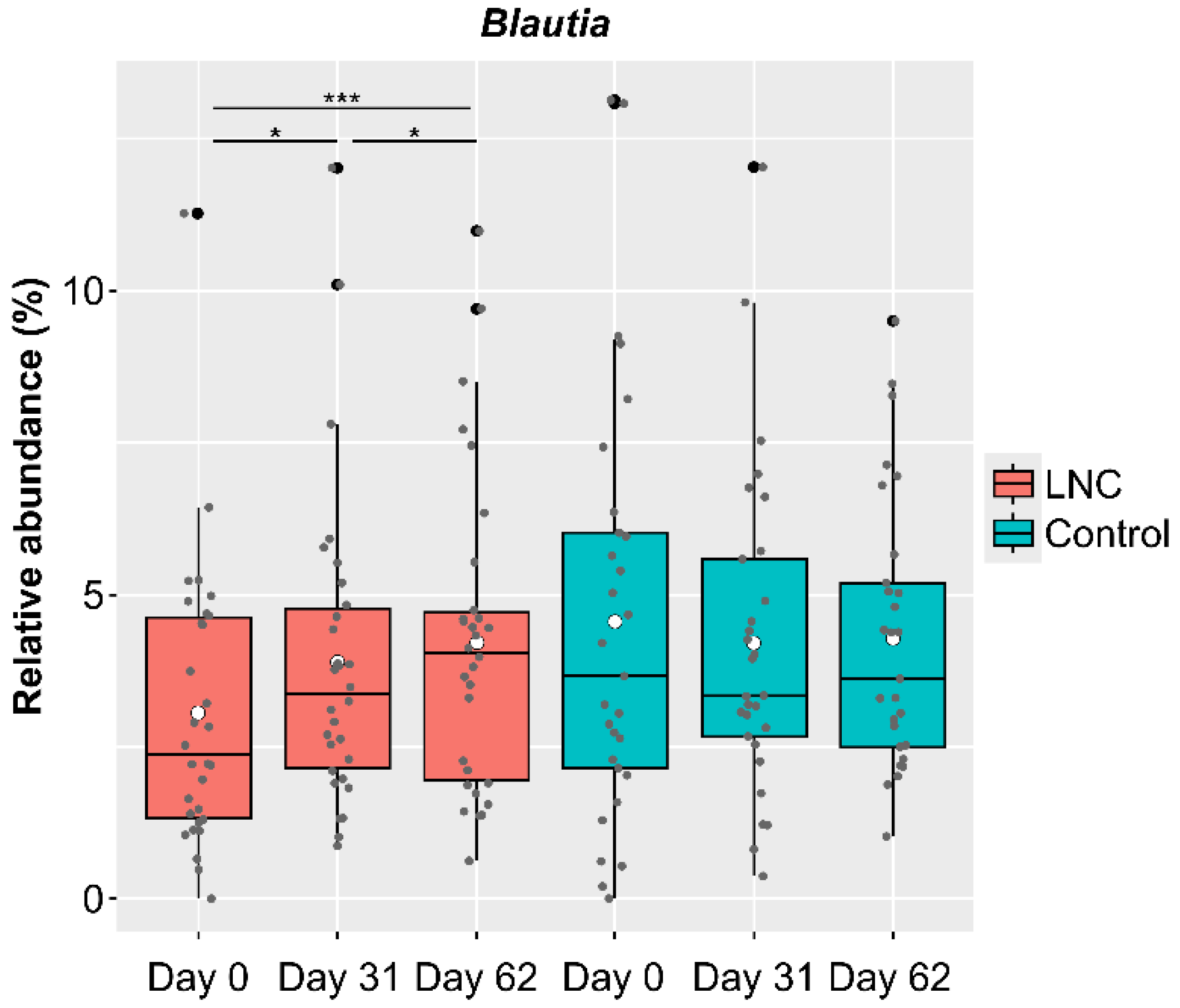
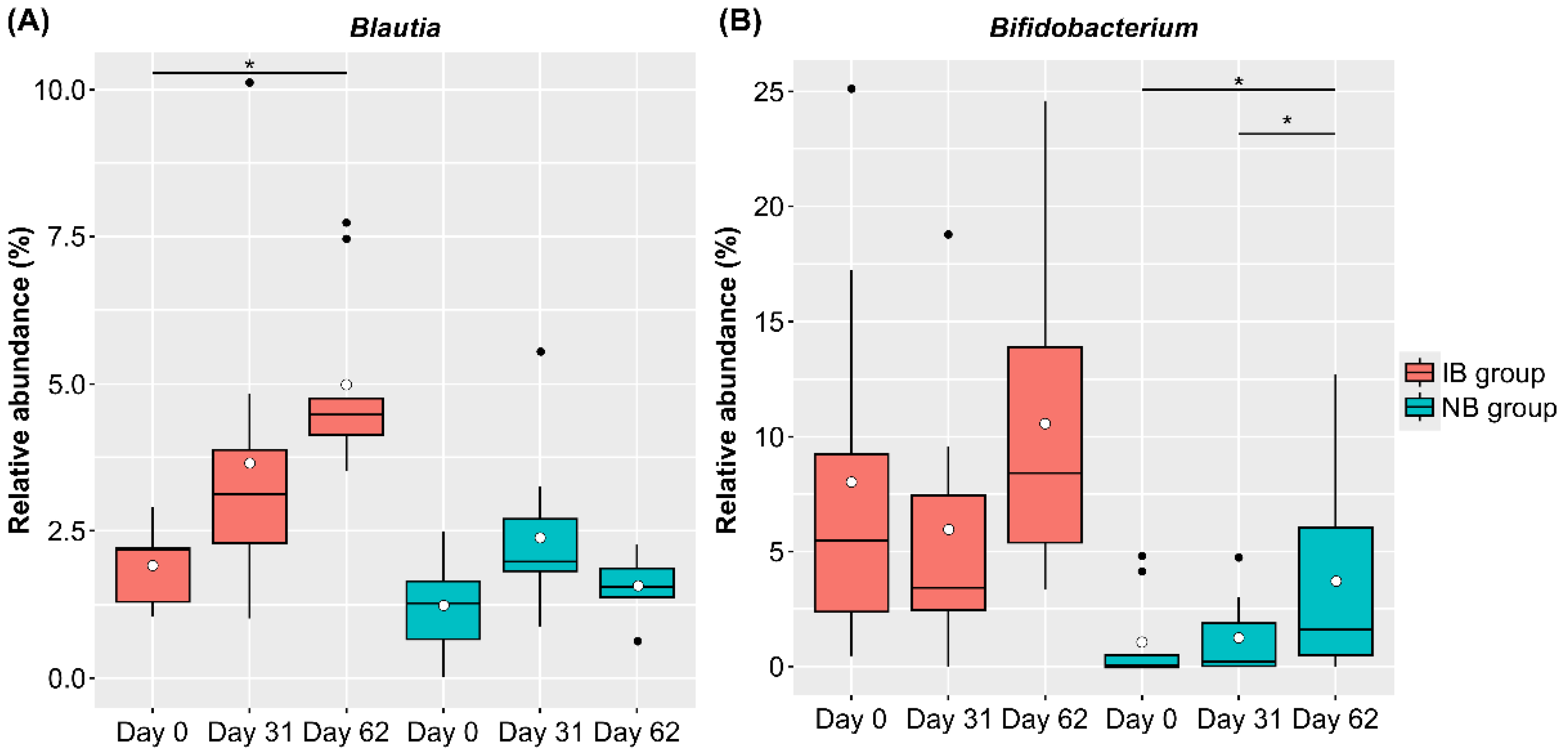
3.3. Changes in Intestinal Microbiota
3.4. Characteristics of Male Participants with a Significant Increase in Blautia Abundance in the LNC Group
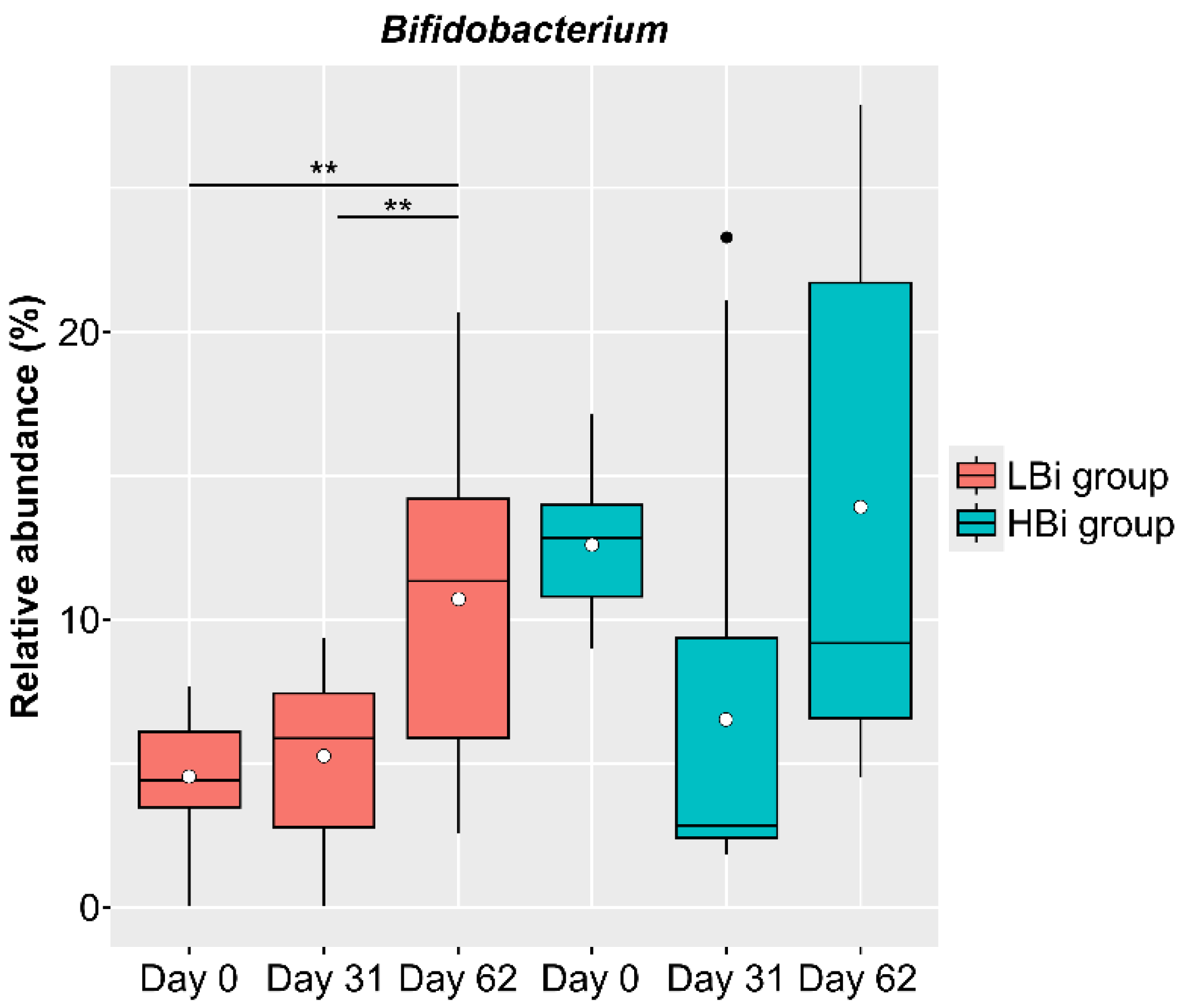
3.5. Baseline-Specific Analysis of Bifidobacterium in Females in the LNC Group
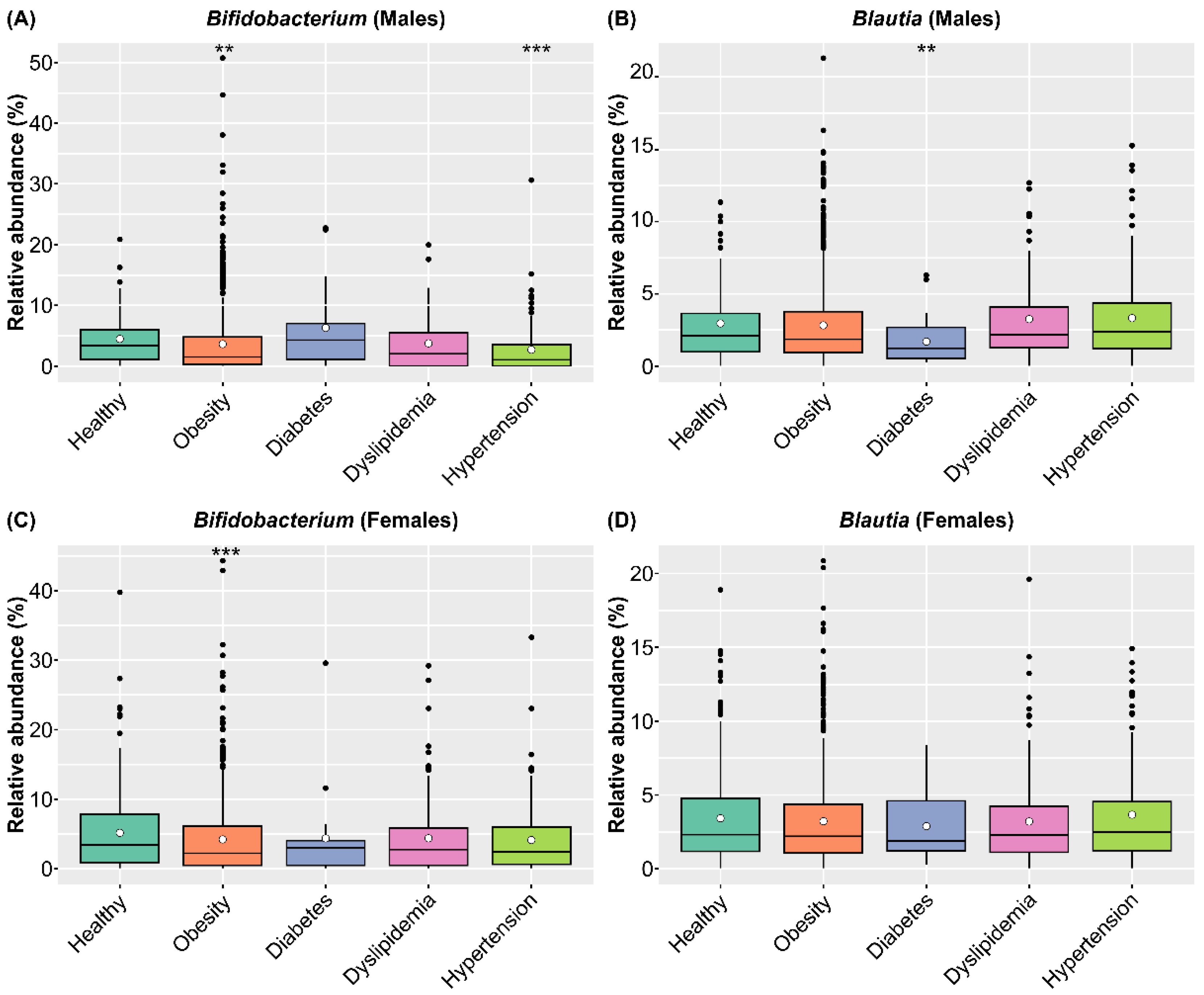
3.6. Effects of Ingesting SONOMONO NATTO POWDER CAPSULESTM Based on Analysis of the SymMAD Database
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Moore, W.E.; Holdeman, L.V. Human Fecal Flora: The Normal Flora of 20 Japanese-Hawaiians. Appl. Microbiol. 1974, 27, 961–979. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, H.; Sakamoto, M.; Benno, Y. Phylogenetic Analysis of the Human Gut Microbiota Using 16S RDNA Clone Libraries and Strictly Anaerobic Culture-Based Methods. Microbiol. Immunol. 2002, 46, 535–548. [Google Scholar] [CrossRef] [PubMed]
- Rajilić-Stojanović, M.; de Vos, W.M. The First 1000 Cultured Species of the Human Gastrointestinal Microbiota. FEMS Microbiol. Rev. 2014, 38, 996–1047. [Google Scholar] [CrossRef] [PubMed]
- Duan, R.; Zhu, S.; Wang, B.; Duan, L. Alterations of Gut Microbiota in Patients With Irritable Bowel Syndrome Based on 16S RRNA-Targeted Sequencing: A Systematic Review. Clin. Transl. Gastroenterol. 2019, 10, e00012. [Google Scholar] [CrossRef]
- Petrov, V.A.; Saltykova, I.V.; Zhukova, I.A.; Alifirova, V.M.; Zhukova, N.G.; Dorofeeva, Y.B.; Tyakht, A.V.; Kovarsky, B.A.; Alekseev, D.G.; Kostryukova, E.S.; et al. Analysis of Gut Microbiota in Patients with Parkinson’s Disease. Bull. Exp. Biol. Med. 2017, 162, 734–737. [Google Scholar] [CrossRef]
- Frémont, M.; Coomans, D.; Massart, S.; De Meirleir, K. High-Throughput 16S RRNA Gene Sequencing Reveals Alterations of Intestinal Microbiota in Myalgic Encephalomyelitis/Chronic Fatigue Syndrome Patients. Anaerobe 2013, 22, 50–56. [Google Scholar] [CrossRef]
- Kim, Y.A.; Keogh, J.B.; Clifton, P.M. Probiotics, Prebiotics, Synbiotics and Insulin Sensitivity. Nutr. Res. Rev. 2018, 31, 35–51. [Google Scholar] [CrossRef]
- Markowiak, P.; Śliżewska, K. Effects of Probiotics, Prebiotics, and Synbiotics on Human Health. Nutrients 2017, 9, 1021. [Google Scholar] [CrossRef]
- Li, H.-Y.; Zhou, D.-D.; Gan, R.-Y.; Huang, S.-Y.; Zhao, C.-N.; Shang, A.; Xu, X.-Y.; Li, H.-B. Effects and Mechanisms of Probiotics, Prebiotics, Synbiotics, and Postbiotics on Metabolic Diseases Targeting Gut Microbiota: A Narrative Review. Nutrients 2021, 13, 3211. [Google Scholar] [CrossRef] [PubMed]
- Oki, K.; Toyama, M.; Banno, T.; Chonan, O.; Benno, Y.; Watanabe, K. Comprehensive Analysis of the Fecal Microbiota of Healthy Japanese Adults Reveals a New Bacterial Lineage Associated with a Phenotype Characterized by a High Frequency of Bowel Movements and a Lean Body Type. BMC Microbiol. 2016, 16, 284. [Google Scholar] [CrossRef] [PubMed]
- Nishijima, S.; Suda, W.; Oshima, K.; Kim, S.-W.; Hirose, Y.; Morita, H.; Hattori, M. The Gut Microbiome of Healthy Japanese and Its Microbial and Functional Uniqueness. DNA Res. 2016, 23, 125–133. [Google Scholar] [CrossRef] [PubMed]
- Cao, Z.-H.; Green-Johnson, J.M.; Buckley, N.D.; Lin, Q.-Y. Bioactivity of Soy-Based Fermented Foods: A Review. Biotechnol. Adv. 2019, 37, 223–238. [Google Scholar] [CrossRef] [PubMed]
- Hosoi, T. Probiotic Effects of Bacillus Subtilis (Natto). J. Brew. Soc. Japan. 2003, 98, 830–839. [Google Scholar] [CrossRef][Green Version]
- Terada, A.; Yamamoto, M.; Yoshimura, E. Effect of the Fermented Soybean Product “Natto” on the Composition and Metabolic Activity of the Human Fecal Flora. Jpn. J. Food Microbiol. 1999, 16, 221–230. [Google Scholar] [CrossRef][Green Version]
- Ishikawa, A.; Kishi, M.; Yamagami, K. Effect of Intake of Natto and Soybeans on Postprandial Blood Glucose Levels in Healthy Adults. SEIKATSU EISEI (J. Urban Living Health Assoc.) 2009, 53, 257–260. [Google Scholar] [CrossRef]
- Kokubo, Y.; Hurukawa, Y.; Banno, M. Prospective Study of the Preventive Effect of Intensive Dietary Soy Intake on Atherosclerosis in an Urban General Population(Part 2). Soy Protein Res. Japan 2012, 15, 6–12. [Google Scholar]
- Hayashi, H.; Sakamoto, M.; Benno, Y. Evaluation of Three Different Forward Primers by Terminal Restriction Fragment Length Polymorphism Analysis for Determination of Fecal Bifidobacterium Spp. in Healthy Subjects. Microbiol. Immunol. 2004, 48, 1–6. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Holmes, S.P. Exact Sequence Variants Should Replace Operational Taxonomic Units in Marker-Gene Data Analysis. ISME J. 2017, 11, 2639–2643. [Google Scholar] [CrossRef] [PubMed]
- Švejstil, R.; Musilová, Š.; Rada, V. Raffinose-Series Oligosaccharides in Soybean Products. Sci. Agric. Bohem. 2015, 46, 73–77. [Google Scholar] [CrossRef]
- Benno, Y.; Endo, K.; Shiragami, N.; Sayama, K.; Mitsuokai, T. Effects of Raffinose Intake on Human Fecal Microflora. Bifidobact. Microflora 1987, 6, 59–63. [Google Scholar] [CrossRef][Green Version]
- Hayakawa, K.; Mizutani, J.; Wada, K.; Masai, T.; Yoshihara, I.; Mitsuoka, T. Effects of Soybean Oligosaccharides on Human Faecal Flora. Microb. Ecol. Health Dis. 1990, 3, 293–303. [Google Scholar] [CrossRef]
- Bouhnik, Y.; Raskine, L.; Simoneau, G.; Vicaut, E.; Neut, C.; Flourié, B.; Brouns, F.; Bornet, F.R. The Capacity of Nondigestible Carbohydrates to Stimulate Fecal Bifidobacteria in Healthy Humans: A Double-Blind, Randomized, Placebo-Controlled, Parallel-Group, Dose-Response Relation Study. Am. J. Clin. Nutr 2004, 80, 1658–1664. [Google Scholar] [CrossRef] [PubMed]
- Kanno, A.; Takamatsu, H.; Takano, N.; Akimoto, T. Change of Saccharides in Soybeans during Manufacturing of Natto. J. Jpn. Soc. Food Sci. Technol. 1982, 29, 105–110. [Google Scholar] [CrossRef]
- Ishikawa, C.; Kodama, S.; Takagaki, R.; Morimitsu, Y. Factors Affecting Manninotriose Concentration in Fermented Soybean Foods. J. Jpn. Soc. Food Sci. Technol. 2020, 67, 58–66. [Google Scholar] [CrossRef]
- Wada, K.; Mizutani, J.; Suzuki, H.; Hayakawa, K. Effect of Manninotriose on Human Fecal Microflora. JSNFS 1991, 44, 171–176. [Google Scholar] [CrossRef][Green Version]
- Hosoi, T.; Ametani, A.; Kiuchi, K.; Kaminogawa, S. Changes in Fecal Microflora Induced by Intubation of Mice with Bacillus Subtilis (Natto) Spores Are Dependent upon Dietary Components. Can. J. Microbiol. 1999, 45, 59–66. [Google Scholar] [CrossRef]
- Takemura, H.; Shioya, N.; Komori, Y.; Tho, Y. Effect of Fermented Soybeans Containing Bacillus Subtilis MC1 on Defecation, Fecal Properties and Fecal Microflora of Healthy Female Volunteers. SEIKATSU EISEI (J. Urban Living Health Assoc.) 2009, 53, 11–18. [Google Scholar] [CrossRef]
- Hatanaka, M.; Nakamura, Y.; Maathuis, A.; Venema, K.; Murota, I.; Yamamoto, N. Influence of Bacillus Subtilis C-3102 on Microbiota in a Dynamic in Vitro Model of the Gastrointestinal Tract Simulating Human Conditions. Benef. Microbes 2012, 3, 229–236. [Google Scholar] [CrossRef]
- Sadao, M.; Minoru, M.; Tsuneo, M. Effect of Oral Intake of Amazake Containing Sake Lees and Rice Koji on the Human Intestinal Microbiota of Amazake Using-The Randomized Placebo-Controlled Crossover Comparison Study. JPT 2020, 48, 1187–1193. [Google Scholar]
- Han, K.; Bose, S.; Wang, J.-H.; Kim, B.-S.; Kim, M.; Kim, E.; Kim, H. Contrasting Effects of Fresh and Fermented Kimchi Consumption on Gut Microbiota Composition and Gene Expression Related to Metabolic Syndrome in Obese Korean Women. Mol. Nutr. Food Res. 2015, 59, 1004–1008. [Google Scholar] [CrossRef]
- Liu, C.; Finegold, S.M.; Song, Y.; Lawson, P.A.Y. Reclassification of Clostridium Coccoides, Ruminococcus Hansenii, Ruminococcus Hydrogenotrophicus, Ruminococcus Luti, Ruminococcus Productus and Ruminococcus Schinkii as Blautia Coccoides Gen. Nov., Comb. Nov., Blautia Hansenii Comb. Nov., Blautia Hydrogenotrophica Comb. Nov., Blautia Luti Comb. Nov., Blautia Producta Comb. Nov., Blautia Schinkii Comb. Nov. and Description of Blautia Wexlerae Sp. Nov., Isolated from Human Faeces. Int. J. Syst. Evol. Microbiol. 2008, 58, 1896–1902. [Google Scholar] [CrossRef] [PubMed]
- Ezaki, T.; Li, N.; Kawamura, Y. The Anaerobic Gram-Positive Cocci. In The Prokaryotes: Volume 4: Bacteria: Firmicutes, Cyanobacteria; Dworkin, M., Falkow, S., Rosenberg, E., Schleifer, K.-H., Stackebrandt, E., Eds.; Springer: New York, NY, USA, 2006; pp. 795–808. ISBN 978-0-387-30744-2. [Google Scholar]
- Simmering, R.; Taras, D.; Schwiertz, A.; Le Blay, G.; Gruhl, B.; Lawson, P.A.; Collins, M.D.; Blaut, M. Ruminococcus Luti Sp. Nov., Isolated from a Human Faecal Sample1 1The GenBank/EMBL/DDBJ Accession Number for the 16S RRNA Gene Sequence of Ruminococcus Luti Strain DSM 14534T Is AJ133124. Syst. Appl. Microbiol. 2002, 25, 189–193. [Google Scholar] [CrossRef]
- Noriega, B.S.; Sanchez-Gonzalez, M.A.; Salyakina, D.; Coffman, J. Understanding the Impact of Omega-3 Rich Diet on the Gut Microbiota. Case Rep. Med. 2016, 2016, e3089303. [Google Scholar] [CrossRef] [PubMed]
- Gurwara, S.; Dai, A.; Ajami, N.; El-Serag, H.B.; Graham, D.Y.; Jiao, L. 196 Caffeine Consumption and the Colonic Mucosa-Associated Gut Microbiota. Am. J. Gastroenterol. 2019, 114, S119. [Google Scholar] [CrossRef]
- Upadhyaya, B.; McCormack, L.; Fardin-Kia, A.R.; Juenemann, R.; Nichenametla, S.; Clapper, J.; Specker, B.; Dey, M. Impact of Dietary Resistant Starch Type 4 on Human Gut Microbiota and Immunometabolic Functions. Sci. Rep. 2016, 6, 28797. [Google Scholar] [CrossRef]
- Plichta, D.R.; Metagenomics of the Human Intestinal Tract (MetaHIT) Consortium; Juncker, A.S.; Bertalan, M.; Rettedal, E.; Gautier, L.; Varela, E.; Manichanh, C.; Fouqueray, C.; Levenez, F.; et al. Transcriptional Interactions Suggest Niche Segregation among Microorganisms in the Human Gut. Nat. Microbiol. 2016, 1, 16152. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.S.; Unno, T.; Kim, B.-Y.; Park, M.-S. Sex Differences in Gut Microbiota. World J. Men’s Health 2019, 38, 48–60. [Google Scholar] [CrossRef]
- Lim, H.J.; Shin, H.S. Antimicrobial and Immunomodulatory Effects of Bifidobacterium Strains: A Review. J. Microbiol. Biotechnol. 2020, 30, 1793–1800. [Google Scholar] [CrossRef]
- Natividad, J.M.M.; Hayes, C.L.; Motta, J.-P.; Jury, J.; Galipeau, H.J.; Philip, V.; Garcia-Rodenas, C.L.; Kiyama, H.; Bercik, P.; Verdu, E.F. Differential Induction of Antimicrobial REGIII by the Intestinal Microbiota and Bifidobacterium Breve NCC2950. Appl. Environ. Microbiol. 2013, 79, 7745–7754. [Google Scholar] [CrossRef]
- Moreno Muñoz, J.A.; Chenoll, E.; Casinos, B.; Bataller, E.; Ramón, D.; Genovés, S.; Montava, R.; Ribes, J.M.; Buesa, J.; Fàbrega, J.; et al. Novel Probiotic Bifidobacterium Longum Subsp. Infantis CECT 7210 Strain Active against Rotavirus Infections. Appl. Environ. Microbiol. 2011, 77, 8775–8783. [Google Scholar] [CrossRef]
- Fukuda, S.; Toh, H.; Hase, K.; Oshima, K.; Nakanishi, Y.; Yoshimura, K.; Tobe, T.; Clarke, J.M.; Topping, D.L.; Suzuki, T.; et al. Bifidobacteria Can Protect from Enteropathogenic Infection through Production of Acetate. Nature 2011, 469, 543–547. [Google Scholar] [CrossRef] [PubMed]
- Fanning, S.; Hall, L.J.; Cronin, M.; Zomer, A.; MacSharry, J.; Goulding, D.; O’Connell Motherway, M.; Shanahan, F.; Nally, K.; Dougan, G.; et al. Bifidobacterial Surface-Exopolysaccharide Facilitates Commensal-Host Interaction through Immune Modulation and Pathogen Protection. Proc. Natl. Acad. Sci. USA 2012, 109, 2108–2113. [Google Scholar] [CrossRef]
- López, P.; González-Rodríguez, I.; Sánchez, B.; Gueimonde, M.; Margolles, A.; Suárez, A. Treg-Inducing Membrane Vesicles from Bifidobacterium Bifidum LMG13195 as Potential Adjuvants in Immunotherapy. Vaccine 2012, 30, 825–829. [Google Scholar] [CrossRef] [PubMed]
- López, P.; de Paz, B.; Rodríguez-Carrio, J.; Hevia, A.; Sánchez, B.; Margolles, A.; Suárez, A. Th17 Responses and Natural IgM Antibodies Are Related to Gut Microbiota Composition in Systemic Lupus Erythematosus Patients. Sci. Rep. 2016, 6, 24072. [Google Scholar] [CrossRef] [PubMed]
- Ozato, N.; Saito, S.; Yamaguchi, T.; Katashima, M.; Tokuda, I.; Sawada, K.; Katsuragi, Y.; Kakuta, M.; Imoto, S.; Ihara, K.; et al. Blautia Genus Associated with Visceral Fat Accumulation in Adults 20–76 Years of Age. NPJ Biofilms Microbiomes 2019, 5, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Liu, F.; Ling, Z.; Tong, X.; Xiang, C. Human Intestinal Lumen and Mucosa-Associated Microbiota in Patients with Colorectal Cancer. PLoS ONE 2012, 7, e39743. [Google Scholar] [CrossRef]
- Larsen, N.; Vogensen, F.K.; van den Berg, F.W.J.; Nielsen, D.S.; Andreasen, A.S.; Pedersen, B.K.; Al-Soud, W.A.; Sørensen, S.J.; Hansen, L.H.; Jakobsen, M. Gut Microbiota in Human Adults with Type 2 Diabetes Differs from Non-Diabetic Adults. PLoS ONE 2010, 5, e9085. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Males | Females | |||||
|---|---|---|---|---|---|---|
| LNC | Control | p Value | LNC | Control | p Value | |
| Number of samples | 30 | 29 | 23 | 34 | ||
| Age | 53.90 ± 14.91 | 49.52 ± 16.41 | 0.278 | 52.48 ± 15.58 | 53.76 ± 15.74 | 0.766 |
| Weight (kg) | 67.71 ± 9.64 | 67.93 ± 10.6 | 0.710 | 55.63 ± 6.14 | 55.11 ± 8.81 | 0.597 |
| BMI (kg/m2) | 23.18 ± 2.8 | 23.51 ± 3.58 | 0.964 | 22.59 ± 3.06 | 21.93 ± 3.19 | 0.229 |
| (A) Bacterial genera with significantly altered abundance in males in the LNC group | |||||||
| Genus | Relative Abundance (%) | p Value (Friedman Test) | p Value (Wilcoxon Signed Rank Test) | ||||
| Day 0 | Day 31 | Day 62 | Day 0 vs. 31 | Day 31 vs. 62 | Day 0 vs. 62 | ||
| Bacteroides | 5.63 ± 5.92 | 6.83 ± 6.48 | 4.57 ± 4.41 | 0.012 | 0.031 | 0.304 | 0.304 |
| Bifidobacterium | 6.17 ±8.20 | 3.65 ± 4.17 | 7.23 ± 5.81 | 0.000 | 0.510 | 0.000 | 0.000 |
| Blautia | 3.06 ± 2.32 | 3.90 ± 2.56 | 4.22 ± 2.60 | 0.001 | 0.029 | 0.047 | 0.000 |
| Collinsella | 2.34 ± 3.79 | 0.63 ± 1.01 | 3.19 ± 2.59 | 0.001 | 0.345 | 0.002 | 0.009 |
| Phocaeicola | 20.96 ± 16.41 | 22.95 ± 14.59 | 15.59 ± 12.53 | 0.012 | 0.016 | 0.103 | 0.331 |
| Unclassified | 10.65 ± 7.25 | 10.63 ± 4.43 | 12.92 ± 7.52 | 0.000 | 0.140 | 0.006 | 0.000 |
| (B) Changes in relative abundance of genera shown in (A) in males in the control group | |||||||
| Genus | Relative abundance (%) | p value (Friedman test) | p value (Wilcoxon signed rank test) | ||||
| Day 0 | Day 31 | Day 62 | Day 0 vs. 31 | Day 31 vs. 62 | Day 0 vs. 62 | ||
| Bacteroides | 5.73 ± 5.62 | 7.59 ± 5.69 | 5.12 ± 4.05 | 0.004 | 0.058 | 0.340 | 0.095 |
| Bifidobacterium | 8.51 ± 7.85 | 4.78 ± 3.54 | 9.45 ± 8.44 | 0.000 | 0.181 | 0.001 | 0.098 |
| Blautia | 4.56 ± 3.51 | 4.20 ± 2.65 | 4.28 ± 2.23 | 0.073 | - | - | - |
| Collinsella | 1.93 ± 2.75 | 0.34 ± 0.85 | 2.97 ± 2.17 | 0.000 | 0.119 | 0.000 | 0.004 |
| Phocaeicola | 24.52 ± 14.36 | 28.75 ± 13.87 | 21.08 ± 13.80 | 0.000 | 0.022 | 0.186 | 0.396 |
| Unclassified | 9.15 ± 4.48 | 10.79 ± 5.02 | 10.96 ± 5.54 | 0.000 | 0.067 | 0.067 | 0.003 |
| (C) Bacterial genera with significantly altered abundance in females in the LNC group | |||||||
| Genus | Relative abundance (%) | p value (Friedman test) | p value (Wilcoxon signed rank test) | ||||
| Day 0 | Day 31 | Day 62 | Day 0 vs. 31 | Day 31 vs. 62 | Day 0 vs. 62 | ||
| Bifidobacterium | 7.70 ± 4.68 | 5.77 ± 5.15 | 11.99 ± 7.14 | 0.001 | 0.448 | 0.000 | 0.003 |
| Faecalibacterium | 6.94 ± 4.67 | 5.30 ± 2.96 | 8.42 ± 6.27 | 0.002 | 0.710 | 0.011 | 0.066 |
| Parabacteroides | 3.05 ± 2.63 | 4.47 ± 3.36 | 2.37 ± 1.77 | 0.008 | 0.004 | 0.050 | 0.424 |
| Unclassified | 13.39 ± 8.98 | 15.84 ± 8.72 | 16.00 ± 9.38 | 0.001 | 0.018 | 0.254 | 0.001 |
| (D) Changes in relative abundance of genera shown in (C) in females in the control group | |||||||
| Genus | Relative abundance (%) | p value (Friedman test) | p value (Wilcoxon signed rank test) | ||||
| Day 0 | Day 31 | Day 62 | Day 0 vs. 31 | Day 31 vs. 62 | Day 0 vs. 62 | ||
| Bifidobacterium | 9.77 ± 7.32 | 6.54 ± 4.76 | 9.94 ± 6.14 | 0.020 | 0.224 | 0.019 | 0.265 |
| Faecalibacterium | 5.79 ± 4.74 | 4.84 ± 4.00 | 5.40 ± 3.92 | 0.152 | - | - | - |
| Parabacteroides | 3.53 ± 4.89 | 4.21 ± 4.73 | 3.63 ± 5.14 | 0.282 | - | - | - |
| Unclassified | 10.83 ± 7.90 | 11.03 ± 7.65 | 13.91 ± 10.91 | 0.000 | 0.301 | 0.001 | 0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kono, K.; Murakami, Y.; Ebara, A.; Okuma, K.; Tokuno, H.; Odachi, A.; Ogasawara, K.; Hidaka, E.; Mori, T.; Satoh, K.; et al. Fluctuations in Intestinal Microbiota Following Ingestion of Natto Powder Containing Bacillus subtilis var. natto SONOMONO Spores: Considerations Using a Large-Scale Intestinal Microflora Database. Nutrients 2022, 14, 3839. https://doi.org/10.3390/nu14183839
Kono K, Murakami Y, Ebara A, Okuma K, Tokuno H, Odachi A, Ogasawara K, Hidaka E, Mori T, Satoh K, et al. Fluctuations in Intestinal Microbiota Following Ingestion of Natto Powder Containing Bacillus subtilis var. natto SONOMONO Spores: Considerations Using a Large-Scale Intestinal Microflora Database. Nutrients. 2022; 14(18):3839. https://doi.org/10.3390/nu14183839
Chicago/Turabian StyleKono, Kanako, Yasufumi Murakami, Aya Ebara, Kana Okuma, Hidetaka Tokuno, Ayano Odachi, Kazuya Ogasawara, Emi Hidaka, Teruaki Mori, Kazuko Satoh, and et al. 2022. "Fluctuations in Intestinal Microbiota Following Ingestion of Natto Powder Containing Bacillus subtilis var. natto SONOMONO Spores: Considerations Using a Large-Scale Intestinal Microflora Database" Nutrients 14, no. 18: 3839. https://doi.org/10.3390/nu14183839
APA StyleKono, K., Murakami, Y., Ebara, A., Okuma, K., Tokuno, H., Odachi, A., Ogasawara, K., Hidaka, E., Mori, T., Satoh, K., Kimoto, S., Masuyama, H., Takeda, M., & Managi, S. (2022). Fluctuations in Intestinal Microbiota Following Ingestion of Natto Powder Containing Bacillus subtilis var. natto SONOMONO Spores: Considerations Using a Large-Scale Intestinal Microflora Database. Nutrients, 14(18), 3839. https://doi.org/10.3390/nu14183839