
Vitamin D Supplementation Does Not Influence SARS-CoV-2 Vaccine Efficacy or Immunogenicity: Sub-Studies Nested within the CORONAVIT Randomised Controlled Trial

 , , , , , , ,
, , , , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design
2.2. Participants
2.3. Randomisation
2.4. Intervention
2.5. Follow-Up Assessments
2.6. Laboratory Assays
2.6.1. 25(OH)D Testing
2.6.2. Anti-S Serology Testing
2.6.3. SARS-CoV-2 Neutralising Antibody
2.6.4. Whole Blood Stimulation Assay
2.6.5. Cytometric Bead Array
2.6.6. Peripheral Blood Mononuclear Cell (PBMC) Isolation
2.6.7. PBMC Stimulation Assay
2.6.8. Flow Cytometric Analysis
2.7. Outcomes
2.8. Statistical Methods
3. Results
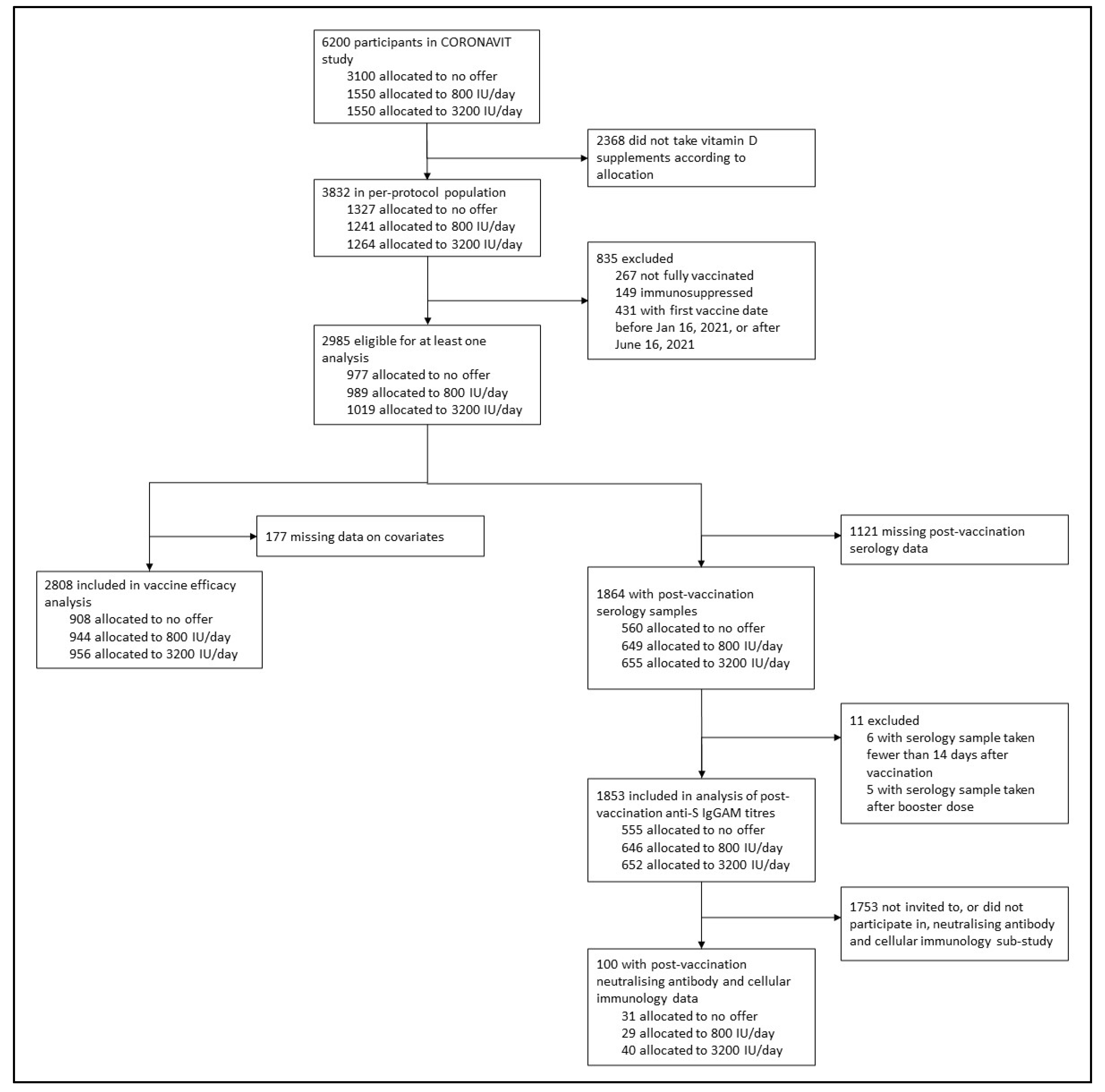
3.1. Participants
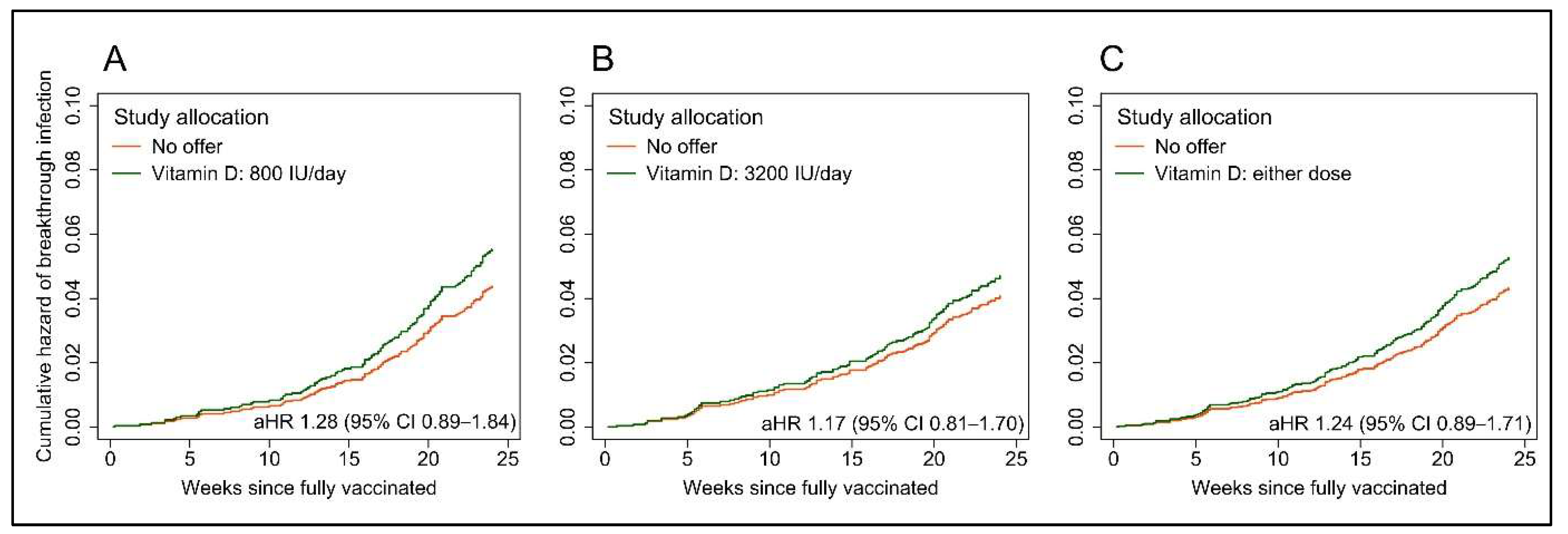
3.2. Breakthrough SARS-CoV-2 Infection
3.3. Immunological Outcomes
3.4. Sensitivity Analysis
3.5. Exploratory Responder Analysis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Feikin, D.R.; Higdon, M.M.; Abu-Raddad, L.J.; Andrews, N.; Araos, R.; Goldberg, Y.; Groome, M.J.; Huppert, A.; O’Brien, K.L.; Smith, P.G.; et al. Duration of effectiveness of vaccines against SARS-CoV-2 infection and COVID-19 disease: Results of a systematic review and meta-regression. Lancet 2022, 399, 924–944. [Google Scholar]
- Liang, Z.; Zhu, H.; Wang, X.; Jing, B.; Li, Z.; Xia, X.; Sun, H.; Yang, Y.; Zhang, W.; Shi, L.; et al. Adjuvants for Coronavirus Vaccines. Front. Immunol. 2020, 11, 589833. [Google Scholar]
- Ciabattini, A.; Nardini, C.; Santoro, F.; Garagnani, P.; Franceschi, C.; Medaglini, D. Vaccination in the elderly: The challenge of immune changes with aging. Semin. Immunol. 2018, 40, 83–94. [Google Scholar]
- De Maeyer, R.P.H.; Chambers, E.S. The impact of ageing on monocytes and macrophages. Immunol. Lett. 2020, 230, 1–10. [Google Scholar]
- Vukmanovic-Stejic, M.; Chambers, E.S.; Suárez-Fariñas, M.; Sandhu, D.; Fuentes-Duculan, J.; Patel, N.; Agius, E.; Lacy, K.E.; Turner, C.T.; Larbi, A.; et al. Enhancement of cutaneous immunity during aging by blocking p38 mitogen-activated protein (MAP) kinase-induced inflammation. J. Allergy Clin. Immunol. 2018, 142, 844–856. [Google Scholar]
- Mannick, J.B.; Del Giudice, G.; Lattanzi, M.; Valiante, N.M.; Praestgaard, J.; Huang, B.; Lonetto, M.A.; Maecker, H.T.; Kovarik, J.; Carson, S.; et al. mTOR inhibition improves immune function in the elderly. Sci. Transl. Med. 2014, 6, 268ra179. [Google Scholar]
- Mannick, J.B.; Morris, M.; Hockey, H.U.; Roma, G.; Beibel, M.; Kulmatycki, K.; Watkins, M.; Shavlakadze, T.; Zhou, W.; Quinn, D.; et al. TORC1 inhibition enhances immune function and reduces infections in the elderly. Sci. Transl. Med. 2018, 10, eaaq1564. [Google Scholar]
- Bikle, D.D. Vitamin D Regulation of Immune Function. Curr. Osteoporos. Rep. 2022, 20, 186–193. [Google Scholar]
- Zhang, Y.; Leung, D.Y.; Richers, B.N.; Liu, Y.; Remigio, L.K.; Riches, D.W.; Goleva, E. Vitamin D inhibits monocyte/macrophage proinflammatory cytokine production by targeting MAPK phosphatase-1. J. Immunol. 2012, 188, 2127–2135. [Google Scholar]
- Lisse, T.S.; Hewison, M. Vitamin D: A new player in the world of mTOR signaling. Cell Cycle 2011, 10, 1888–1889. [Google Scholar]
- von Essen, M.R.; Kongsbak, M.; Schjerling, P.; Olgaard, K.; Odum, N.; Geisler, C. Vitamin D controls T cell antigen receptor signaling and activation of human T cells. Nat. Immunol. 2010, 11, 344–349. [Google Scholar]
- De Vita, F.; Lauretani, F.; Bauer, J.; Bautmans, I.; Shardell, M.; Cherubini, A.; Bondi, G.; Zuliani, G.; Bandinelli, S.; Pedrazzoni, M.; et al. Relationship between vitamin D and inflammatory markers in older individuals. Age 2014, 36, 9694. [Google Scholar]
- Laird, E.; McNulty, H.; Ward, M.; Hoey, L.; McSorley, E.; Wallace, J.M.W.; Carson, E.; Molloy, A.; Healy, M.; Casey, M.C.; et al. Vitamin D Deficiency Is Associated With Inflammation in Older Irish Adults. J. Clin. Endocrinol. Metab. 2014, 99, 1807–1815. [Google Scholar]
- Chambers, E.S.; Vukmanovic-Stejic, M.; Turner, C.T.; Shih, B.B.; Trahair, H.; Pollara, G.; Tsaliki, E.; Rustin, M.; Freeman, T.C.; Mabbott, A.N.; et al. Vitamin D3 replacement enhances antigen-specific immunity in older adults. Immunother. Adv. 2020, 1, ltaa008. [Google Scholar]
- Calder, P.C.; Berger, M.M.; Gombart, A.F.; McComsey, G.A.; Martineau, A.R.; Eggersdorfer, M. Micronutrients to Support Vaccine Immunogenicity and Efficacy. Vaccines 2022, 10, 568. [Google Scholar]
- Chiu, S.-K.; Tsai, K.-W.; Wu, C.-C.; Zheng, C.-M.; Yang, C.-H.; Hu, W.-C.; Hou, Y.-C.; Lu, K.-C.; Chao, Y.-C. Putative Role of Vitamin D for COVID-19 Vaccination. Int. J. Mol. Sci. 2021, 22, 8988. [Google Scholar]
- Jolliffe, D.A.; Faustini, S.E.; Holt, H.; Perdek, N.; Maltby, S.; Talaei, M.; Greenig, M.; Vivaldi, G.; Tydeman, F.; Symons, J.; et al. Determinants of antibody responses to SARS-CoV-2 vaccines: Population-based longitudinal study (COVIDENCE UK). Vaccines (Basel), 2022; in press. [Google Scholar]
- Piec, I.; Cook, L.; Dervisevic, S.; Fraser, W.D.; Ruetten, S.; Berman, M.; English, E.; John, W. Age and vitamin D affect the magnitude of the antibody response to the first dose of the SARS-CoV-2 BNT162b2 vaccine. Curr. Res. Transl. Med. 2022, 70, 103344. [Google Scholar]
- Chillon, T.S.; Demircan, K.; Heller, R.A.; Hirschbil-Bremer, I.M.; Diegmann, J.; Bachmann, M.; Moghaddam, A.; Schomburg, L. Relationship between Vitamin D Status and Antibody Response to COVID-19 mRNA Vaccination in Healthy Adults. Biomedicines 2021, 9, 1714. [Google Scholar]
- Parthymou, A.; Habeos, E.E.; Habeos, G.I.; Deligakis, A.; Livieratos, E.; Marangos, M.; Chartoumpekis, D.V. Factors associated with anti-SARS-CoV-2 antibody titres 3 months post-vaccination with the second dose of BNT162b2 vaccine: A longitudinal observational cohort study in western Greece. BMJ Open 2022, 12, e057084. [Google Scholar]
- Jolliffe, D.; Holt, H.; Greenig, M.; Talaei, M.; Perdek, N.; Pfeffer, P.; Maltby, S.; Symons, J.; Barlow, N.; Normandale, A.; et al. Vitamin D Supplements for Prevention of Covid-19 or other Acute Respiratory Infections: A Phase 3 Randomized Controlled Trial (CORONAVIT). BMJ 2022, 378, e071230. [Google Scholar]
- Hypponen, E.; Power, C. Hypovitaminosis D in British adults at age 45 y: Nationwide cohort study of dietary and lifestyle predictors. Am. J. Clin. Nutr. 2007, 85, 860–868. [Google Scholar]
- Holt, H.; Relton, C.; Talaei, M.; Symons, J.; Davies, M.R.; Jolliffe, D.A.; Vivaldi, G.; Tydeman, F.; Williamson, A.; Pfeffer, P.E.; et al. Cohort Profile: Longitudinal population-based study of COVID-19 in UK adults (COVIDENCE UK). Int. J. Epidemiol. 2022; in press. [Google Scholar]
- Shea, R.L.; Berg, J.D. Self-administration of vitamin D supplements in the general public may be associated with high 25-hydroxyvitamin D concentrations. Ann. Clin. Biochem. 2017, 54, 355–361. [Google Scholar]
- Dawson-Hughes, B.; Heaney, R.P.; Holick, M.F.; Lips, P.; Meunier, P.J.; Vieth, R. Estimates of optimal vitamin D status. Osteoporos. Int. 2005, 16, 713–716. [Google Scholar]
- Vieth, R. What is the optimal vitamin D status for health? Prog. Biophys. Mol. Biol. 2006, 92, 26–32. [Google Scholar]
- Bischoff-Ferrari, H.A. The 25-hydroxyvitamin D threshold for better health. J. Steroid. Biochem. Mol. Biol. 2007, 103, 614–619. [Google Scholar]
- Cook, A.M.; Faustini, S.E.; Williams, L.J.; Cunningham, A.F.; Drayson, M.T.; Shields, A.M.; Kay, D.; Taylor, L.; Plant, T.; Huissoon, A.; et al. Validation of a combined ELISA to detect IgG, IgA and IgM antibody responses to SARS-CoV-2 in mild or moderate non-hospitalised patients. J. Immunol. Methods 2021, 494, 113046. [Google Scholar]
- Shields, A.; Faustini, S.; Kristunas, C.; Cook, A.; Backhouse, C.; Dunbar, L.; Ebanks, D.; Emmanuel, B.; Crouch, E.; Kröger, A.; et al. COVID-19: Seroprevalence and Vaccine Responses in UK Dental Care Professionals. J. Dent. Res. 2021, 100, 1220–1227. [Google Scholar]
- Vivaldi, G.; Jolliffe, D.A.; Faustini, S.; Holt, H.; Perdek, N.; Talaei, M.; Tydeman, F.; Chambers, E.S.; Cai, W.; Li, W.; et al. Correlation between post-vaccination titres of IgG, IgA and IgM anti-Spike antibodies and protection against breakthrough SARS-CoV-2 infection: A population-based longitudinal study (COVIDENCE UK). J. Infect. Dis. 2022. Available online: https://pubmed.ncbi.nlm.nih.gov/35906930/ (accessed on 17 August 2022).
- Reynolds, C.J.; Swadling, L.; Gibbons, J.M.; Pade, C.; Jensen, M.P.; Diniz, M.O.; Schmidt, N.M.; Butler, D.K.; Amin, O.E.; Bailey, S.N.; et al. Discordant neutralizing antibody and T cell responses in asymptomatic and mild SARS-CoV-2 infection. Sci. Immunol. 2020, 5, eabf3698. [Google Scholar]
- Grayling, M.J.; Wason, J.M. A web application for the design of multi-arm clinical trials. BMC Cancer 2020, 20, 80. [Google Scholar]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B 1995, 57, 289–300. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
| Overall (n = 2808) | No Offer (n = 908) | 800 IU/Day Offer (n = 944) | 3200 IU/Day Offer (n = 956) | |
|---|---|---|---|---|
| Age, years | ||||
| Median (IQR) | 61.9 (54.1–68.5) | 61.8 (53.6–68.1) | 61.4 (54.2–68.3) | 62.5 (54.1–69.1) |
| Range | 16.6–87.2 | 18.4–85.9 | 20.9–83.1 | 16.6–87.2 |
| Sex | ||||
| Female | 1848 (65.8%) | 579 (63.8%) | 634 (67.2%) | 635 (66.4%) |
| Male | 960 (34.2%) | 329 (36.2%) | 310 (32.8%) | 321 (33.6%) |
| Ethnicity | ||||
| White | 2706 (96.4%) | 885 (97.5%) | 910 (96.4%) | 911 (95.3%) |
| Asian/Asian British | 25 (0.9%) | 9 (1.0%) | 5 (0.5%) | 11 (1.2%) |
| Black/African/Caribbean/Black British | 14 (0.5%) | 2 (0.2%) | 6 (0.6%) | 6 (0.6%) |
| Mixed/Multiple/Other | 63 (2.2%) | 12 (1.3%) | 23 (2.4%) | 28 (2.9%) |
| Body mass index, kg/m² | ||||
| <25 | 1304 (46.5%) | 404 (44.5%) | 440 (46.8%) | 460 (48.1%) |
| 25–30 | 949 (33.8%) | 314 (34.6%) | 307 (32.6%) | 328 (34.3%) |
| >30 | 551 (19.7%) | 189 (20.8%) | 194 (20.6%) | 168 (17.6%) |
| Self-assessed general health | ||||
| Excellent | 617 (22.0%) | 203 (22.4%) | 196 (20.8%) | 218 (22.8%) |
| Very good | 1186 (42.3%) | 386 (42.5%) | 404 (42.8%) | 396 (41.5%) |
| Good | 721 (25.7%) | 213 (23.5%) | 257 (27.2%) | 251 (26.3%) |
| Fair | 222 (7.9%) | 86 (9.5%) | 67 (7.1%) | 69 (7.2%) |
| Poor | 61 (2.2%) | 20 (2.2%) | 20 (2.1%) | 21 (2.2%) |
| Medically diagnosed disease | ||||
| Hypertension | 577 (20.5%) | 191 (21.0%) | 190 (20.1%) | 196 (20.5%) |
| Diabetes | 135 (4.8%) | 50 (5.5%) | 35 (3.7%) | 50 (5.2%) |
| Heart disease | 113 (4.0%) | 41 (4.5%) | 39 (4.1%) | 33 (3.5%) |
| Asthma | 394 (14.0%) | 120 (13.2%) | 160 (16.9%) | 114 (11.9%) |
| COPD | 45 (1.6%) | 18 (2.0%) | 15 (1.6%) | 12 (1.3%) |
| Pre-vaccination SARS-CoV-2 infection (1) | 121 (4.3%) | 38 (4.2%) | 55 (5.8%) | 28 (2.9%) |
| Type of vaccine administered, primary course (2) | ||||
| 2 × ChAdOx1 | 1945 (69.3%) | 632 (69.6%) | 673 (71.3%) | 640 (66.9%) |
| 2 × BNT162b2 | 863 (30.7%) | 276 (30.4%) | 271 (28.7%) | 316 (33.1%) |
| Inter-dose interval, days | 77 (69–79) | 77 (68–79) | 77 (69–79) | 77 (69–79) |
| Mean 25(OH)D, nmol/L (SD) [range] (3) | 39.9 (14.5) [10.3–74.9] | -- (4) | 39.6 (14.7) [10.3–74.8] | 40.2 (14.4) [10.3–74.9] |
| <25.0 | 309 (11.0%) | -- (4) | 164 (17.4%) | 145 (15.2%) |
| 25.0 to <50.0 | 1104 (39.3%) | -- (4) | 543 (57.5%) | 561 (58.7%) |
| 50.0 to <75.0 | 482 (17.2%) | -- (4) | 235 (24.9%) | 247 (25.8%) |
| ≥75.0 | 0 | -- (4) | 0 | 0 |
| Not determined | 913 (32.5%) | 908 (100.0%) | 2 (0.2%) | 3 (0.3%) |
| No Offer | 800 IU/Day Offer | 3200 IU/Day Offer | 800 IU/Day vs. No Offer | 3200 IU/Day vs. No Offer | |||
|---|---|---|---|---|---|---|---|
| Adjusted % difference (95% CI) * | P† | Adjusted % difference (95% CI) * | P† | ||||
| Sub-study 2 | |||||||
| Anti-S IgGAM ratio | 2.8 (1.9 to 3.9) [n = 555] | 2.9 (1.9 to 4.1) [n = 646] | 2.8 (1.9 to 4.1) [n = 652] | −20.6% (−84.9 to 316.6) | 0.781 | −64.4% (−92.9 to 78.5) | 0.204 |
| Sub-study 3 | |||||||
| Neutralising antibody titre | 186.9 (119.8 to 406.7) [n = 29] | 219.0 (107.0 to 333.0) [n = 29] | 241.2 (81.2 to 713.8) [n = 37] | 1.2% (−40.0 to 70.7) | 0.963 | −10.8% (−49.5 to 57.5) | 0.686 |
| S peptide-stimulated IFN-γ in whole blood supernatant, ng/mL | 0.011 (0.003 to 0.025) [n = 29] | 0.013 (0.005 to 0.046) [n = 28] | 0.013 (0.005 to 0.024) [n = 37] | 54.7% (−31.3 to 248.2) | 0.285 | 49.2% (−41.8 to 282.2) | 0.396 |
| S peptide-stimulated TNF in whole blood supernatant, ng/mL | 0.000 (0.000 to 0.003) [n = 29] | 0.000 (0.000 to 0.000) [n = 28] | 0.000 (0.000 to 0.027) [n = 39] | −18.9% (−72.5 to 139.0) | 0.700 | −68.4% (−90.0 to −0.5) | 0.049 |
| S peptide-stimulated IL-6 in whole blood supernatant, ng/mL | 0.136 (0.012 to 0.719) [n = 28] | 0.127 (0.000 to 1.246) [n = 27] | 0.080 (0.000 to 4.273) [n = 39] | −45.3% (−89.7 to 189.8) | 0.471 | −60.3% (−94.7 to 197.1) | 0.360 |
| S peptide-stimulated CXCL8 in whole blood supernatant, ng/mL | 0.790 (0.254 to 2.563) [n = 26] | 0.893 (0.295 to 2.577) [n = 27] | 1.416 (0.388 to 12.016) [n = 39] | 70.0% (−50.6 to 485.4) | 0.393 | −12.7% (−80.1 to 283.7) | 0.854 |
| LPS-stimulated IFN-γ in whole blood supernatant, ng/mL | 0.020 (0.004 to 0.067) [n = 29] | 0.027 (0.009 to 0.062) [n = 28] | 0.028 (0.006 to 0.109) [n = 37] | 44.9% (−52.7 to 343.4) | 0.509 | −14.9% (−67.8 to 125.0) | 0.739 |
| LPS-stimulated TNF in whole blood supernatant, ng/mL | 0.542 (0.465 to 0.919) [n = 29] | 0.597 (0.350 to 0.923) [n = 28] | 0.473 (0.302 to 0.836) [n = 40] | −57.8% (−85.1 to 19.2) | 0.101 | −17.1% (−51.5 to 41.7) | 0.484 |
| LPS-stimulated IL-6 in whole blood supernatant, ng/mL | 54.768 (32.362 to 73.147) [n = 29] | 43.202 (28.838 to 79.264) [n = 28] | 46.142 (32.146 to 91.936) [n = 40] | 10.4% (−26.3 to 65.5) | 0.625 | −15.3% (−45.4 to 31.3) | 0.449 |
| LPS-stimulated CXCL8 in whole blood supernatant, ng/mL | 4.231 (2.322 to 6.373) [n = 28] | 2.642 (1.540 to 4.213) [n = 28] | 2.732 (2.169 to 4.392) [n = 40] | −9.4% (−39.7 to 36.3) | 0.631 | −29.9% (−57.6 to 16.0) | 0.162 |
| Percentage of S peptide-stimulated CD3 + CD4+ cells positive for intracellular IFN-γ | 0.00% (0.00 to 0.02) [n = 29] | 0.00% (0.00 to 0.02) [n = 27] | 0.00% (0.00 to 0.02) [n = 37] | −31.7% (−59.6 to 15.4) | 0.151 | −17.5% (−60.1 to 70.9) | 0.598 |
| Percentage of CD3-stimulated CD3 + CD4+ cells positive for intracellular IFN-γ | 0.04% (0.00 to 0.12) [n = 29] | 0.09% (0.01 to 0.23) [n = 26] | 0.12% (0.04 to 0.42) [n = 37] | 285.6% (26.6 to 1074.1) | 0.019 | 156.1% (−30.8 to 847.2) | 0.154 |
| Percentage of S peptide-stimulated CD3 + CD8+ cells positive for intracellular IFN-γ | 0.00% (0.00 to 0.04) [n = 29] | 0.00% (0.00 to 0.08) [n = 27] | 0.01% (0.00 to 0.06) [n = 37] | 10.8% (−48.4 to 137.8) | 0.788 | 50.2% (−41.5 to 285.6) | 0.389 |
| Percentage of CD3-stimulated CD3 + CD8+ cells positive for intracellular IFN-γ | 0.62% (0.18 to 2.15) [n = 29] | 0.93% (0.40 to 2.09) [n = 26] | 0.95% (0.25 to 1.96) [n = 37] | 70.3% (−35.4 to 349.0) | 0.276 | 106.1% (−29.5 to 501.9) | 0.181 |
| Percentage of S peptide-stimulated CD3 + CD4+ cells positive for intracellular IL-2 | 0.00% (0.00 to 0.02) [n = 29] | 0.01% (0.00 to 0.03) [n = 27] | 0.01% (0.00 to 0.02) [n = 37] | 23.7% (−18.6 to 88.0) | 0.312 | 36.4% (−12.9 to 113.7) | 0.170 |
| Percentage of CD3-stimulated CD3 + CD4+ cells positive for intracellular IL-2 | 0.12% (0.09 to 0.25) [n = 29] | 0.17% (0.06 to 0.32) [n = 26] | 0.18% (0.09 to 0.35) [n = 37] | 45.7% (−18.8 to 161.7) | 0.202 | 47.8% (−36.2 to 242.8) | 0.353 |
| Percentage of S peptide-stimulated CD3 + CD8+ cells positive for intracellular IL-2 | 0.00% (0.00 to 0.00) [n = 29] | 0.00% (0.00 to 0.00) [n = 27] | 0.00% (0.00 to 0.00) [n = 37] | 43.1% (−0.6 to 106.2) | 0.054 | 28.6% (−21.1 to 109.6) | 0.304 |
| Percentage of CD3-stimulated CD3 + CD8+ cells positive for intracellular IL-2 | 0.13% (0.05 to 0.26) [n = 29] | 0.20% (0.11 to 0.47) [n = 26] | 0.17% (0.08 to 0.31) [n = 37] | 56.0% (−19.6 to 202.7) | 0.184 | 78.9% (−23.3 to 317.4) | 0.173 |
| Percentage of S peptide-stimulated CD3 + CD4+ cells positive for intracellular TNF | 0.00% (0.00 to 0.00) [n = 29] | 0.00% (0.00 to 0.04) [n = 27] | 0.00% (0.00 to 0.02) [n = 37] | 22.6% (−36.0 to 135.2) | 0.532 | 63.0% (−28.8 to 273.1) | 0.241 |
| Percentage of CD3-stimulated CD3 + CD4+ cells positive for intracellular TNF | 0.37% (0.19 to 0.67) [n = 29] | 0.67% (0.37 to 1.08) [n = 26] | 0.74% (0.22 to 1.37) [n = 37] | 80.4% (−15.2 to 283.9) | 0.123 | 101.9% (−24.4 to 439.2) | 0.156 |
| Percentage of S peptide-stimulated CD3 + CD8+ cells positive for intracellular TNF | 0.02% (0.00 to 0.06) [n = 29] | 0.03% (0.00 to 0.11) [n = 27] | 0.00% (0.00 to 0.06) [n = 37] | −41.8% (−76.6 to 45.0) | 0.239 | −6.5% (−70.6 to 197.8) | 0.908 |
| Percentage of CD3-stimulated CD3 + CD8+ cells positive for intracellular TNF | 1.45% (0.72 to 3.84) [n = 29] | 2.57% (1.56 to 4.26) [n = 26] | 2.38% (0.82 to 4.40) [n = 37] | 129.4% (−11.5 to 495.1) | 0.086 | 176.1% (−10.3 to 749.4) | 0.075 |
| Percentage of CD3 + CD4+ cells with naive phenotype | 35.8% (21.7 to 48.7) [n = 29] | 31.9% (20.2 to 42.6) [n = 27] | 36.8% (23.6 to 43.1) [n = 37] | −0.2% (−23.5 to 30.2) | 0.988 | −8.6% (−32.0 to 22.8) | 0.541 |
| Percentage of CD3 + CD8+ cells with naive phenotype | 32.5% (22.9 to 42.6) [n = 29] | 24.4% (19.5 to 32.7) [n = 27] | 24.8% (19.6 to 36.3) [n = 37] | −14.0% (−32.1 to 8.9) | 0.204 | −14.5% (−34.4 to 11.5) | 0.241 |
| Percentage of CD3 + CD4+ cells with central memory phenotype | 33.5% (26.6 to 40.6) [n = 29] | 39.6% (31.2 to 46.5) [n = 27] | 35.6% (30.7 to 44.9) [n = 37] | 14.4% (−6.8 to 40.4) | 0.194 | 24.3% (3.4 to 49.3) | 0.021 |
| Percentage of CD3 + CD8+ cells with central memory phenotype | 20.5% (13.9 to 28.8) [n = 29] | 24.7% (15.9 to 33.2) [n = 27] | 22.6% (11.6 to 28.1) [n = 37] | 10.1% (−20.8 to 53.0) | 0.559 | 35.5% (0.0 to 83.7) | 0.050 |
| Percentage of CD3 + CD4+ cells with effector memory phenotype | 11.7% (7.3 to 15.7) [n = 29] | 14.4% (9.8 to 17.3) [n = 27] | 13.0% (8.8 to 17.9) [n = 37] | 8.4% (−17.7 to 42.9) | 0.558 | 16.6% (−13.6 to 57.4) | 0.306 |
| Percentage of CD3 + CD8+ cells with effector memory phenotype | 8.2% (5.6 to 12.7) [n = 29] | 12.2% (9.5 to 15.7) [n = 27] | 11.4% (6.5 to 15.1) [n = 37] | 32.3% (−3.7 to 81.7) | 0.083 | 44.4% (3.3 to 102.0) | 0.033 |
| Percentage of CD3 + CD4+ cells with EMRA phenotype | 3.9% (2.4 to 7.9) [n = 29] | 3.6% (2.4 to 6.1) [n = 27] | 3.1% (1.7 to 5.2) [n = 37] | −35.6% (−59.0 to 1.2) | 0.056 | −20.5% (−53.2 to 35.1) | 0.388 |
| Percentage of CD3 + CD8+ cells with EMRA phenotype | 14.9% (10.3 to 29.7) [n = 29] | 18.5% (9.5 to 25.9) [n = 27] | 20.7% (14.0 to 28.7) [n = 37] | 15.6% (−16.0 to 59.2) | 0.366 | 0.2% (−33.9 to 52.0) | 0.992 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jolliffe, D.A.; Vivaldi, G.; Chambers, E.S.; Cai, W.; Li, W.; Faustini, S.E.; Gibbons, J.M.; Pade, C.; Coussens, A.K.; Richter, A.G.; et al. Vitamin D Supplementation Does Not Influence SARS-CoV-2 Vaccine Efficacy or Immunogenicity: Sub-Studies Nested within the CORONAVIT Randomised Controlled Trial. Nutrients 2022, 14, 3821. https://doi.org/10.3390/nu14183821
Jolliffe DA, Vivaldi G, Chambers ES, Cai W, Li W, Faustini SE, Gibbons JM, Pade C, Coussens AK, Richter AG, et al. Vitamin D Supplementation Does Not Influence SARS-CoV-2 Vaccine Efficacy or Immunogenicity: Sub-Studies Nested within the CORONAVIT Randomised Controlled Trial. Nutrients. 2022; 14(18):3821. https://doi.org/10.3390/nu14183821
Chicago/Turabian StyleJolliffe, David A., Giulia Vivaldi, Emma S. Chambers, Weigang Cai, Wenhao Li, Sian E. Faustini, Joseph M. Gibbons, Corinna Pade, Anna K. Coussens, Alex G. Richter, and et al. 2022. "Vitamin D Supplementation Does Not Influence SARS-CoV-2 Vaccine Efficacy or Immunogenicity: Sub-Studies Nested within the CORONAVIT Randomised Controlled Trial" Nutrients 14, no. 18: 3821. https://doi.org/10.3390/nu14183821
APA StyleJolliffe, D. A., Vivaldi, G., Chambers, E. S., Cai, W., Li, W., Faustini, S. E., Gibbons, J. M., Pade, C., Coussens, A. K., Richter, A. G., McKnight, Á., & Martineau, A. R. (2022). Vitamin D Supplementation Does Not Influence SARS-CoV-2 Vaccine Efficacy or Immunogenicity: Sub-Studies Nested within the CORONAVIT Randomised Controlled Trial. Nutrients, 14(18), 3821. https://doi.org/10.3390/nu14183821