
Phytoestrogen Coumestrol Selectively Inhibits Monoamine Oxidase-A and Amyloid β Self-Aggregation



Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Plant Source
2.3. HPLC–ESI-Q–TOF–MS Analysis
2.4. In Vitro hMAO Inhibitory Assay
2.5. Kinetic Assay for hMAO-A
2.6. Self Aβ25–35 Aggregation Assay
2.7. In Silico Docking Simulation
2.8. Prediction of Pharmacokinetic Parameters
2.9. Statistical Analysis
3. Results
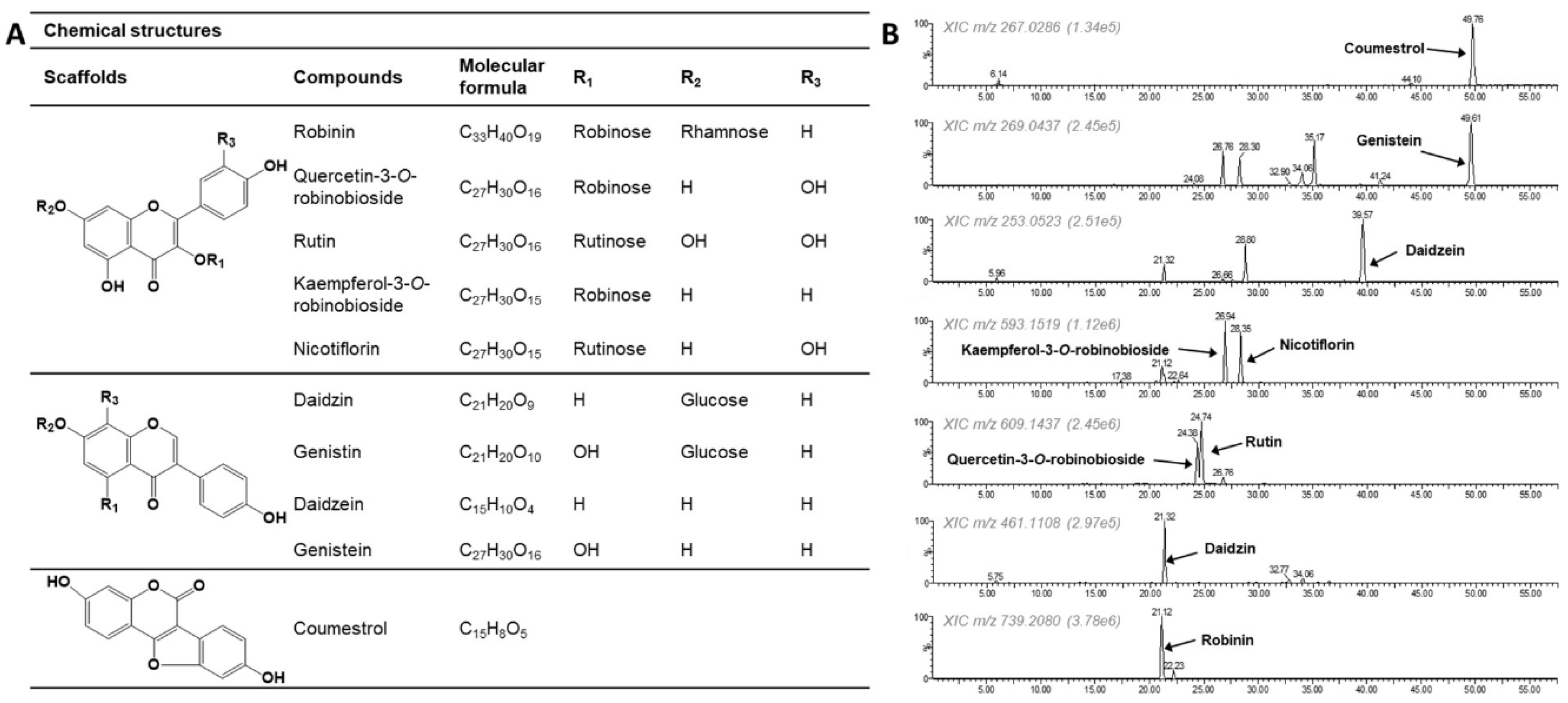
3.1. Extracted Ion Chromatogram (EIC) of Compounds Present in the PL-L-70E
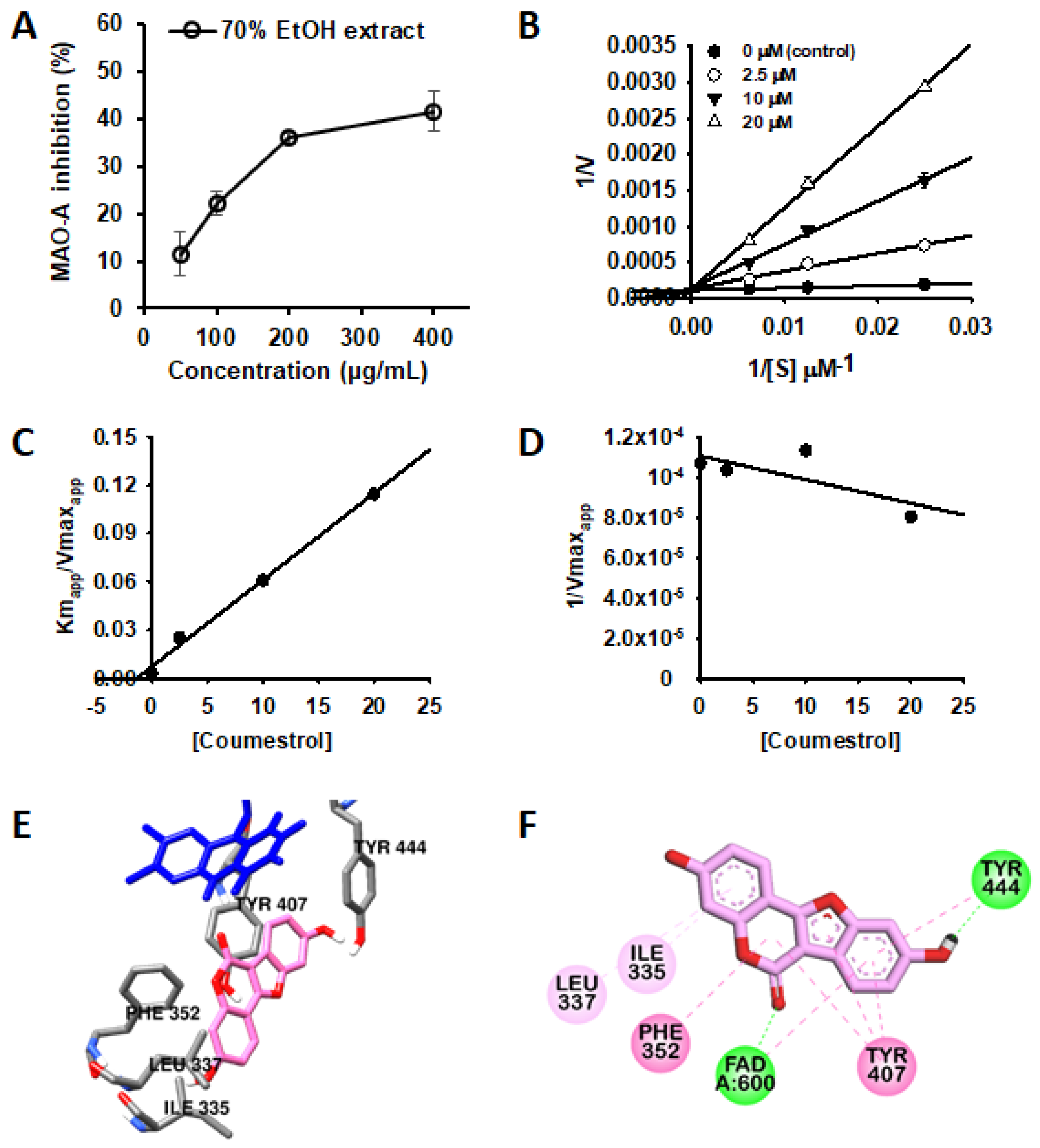
3.2. Inhibitory Activity of PL-L-70E and Its Constituents against Human MAO Isozymes
3.3. Competitive Inhibition of hMAO-A by Coumestrol
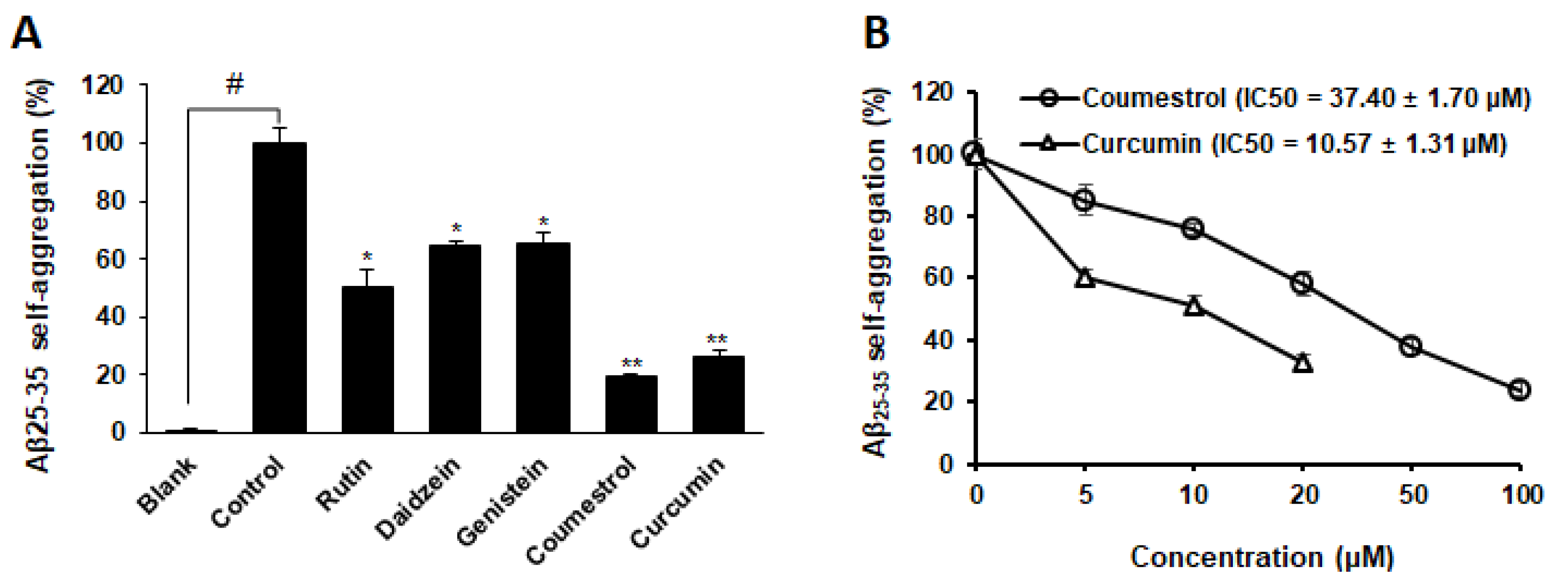
3.4. Inhibitory Activity of Phytoestrogens against Aβ25–35 Self-Aggregation
3.5. Pharmacokinetic Parameters of Coumestrol
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Santin, Y.; Resta, J.; Parini, A.; Mialet-Perez, J. Monoamine oxidases in age-associated diseases: New perspectives for old enzymes. Ageing Res. Rev. 2021, 66, 101256. [Google Scholar] [CrossRef]
- Manzoor, S.; Hoda, N. A comprehensive review of monoamine oxidase inhibitors as anti-Alzheimer’s disease agents: A review. Eur. J. Med. Chem. 2020, 206, 112787. [Google Scholar] [CrossRef]
- Suchting, R.; Tirumalaraju, V.; Gareeb, R.; Bockmann, T.; de Dios, C.; Aickareth, J.; Pinjari, O.; Soares, J.C.; Cowen, P.J.; Selvaraj, S. Revisiting monoamine oxidase inhibitors for the treatment of depressive disorders: A systematic review and network meta-analysis. J. Affect. Disord. 2021, 282, 1153–1160. [Google Scholar] [CrossRef]
- Schedin-Weiss, S.; Inoue, M.; Hromadkova, L.; Teranishi, Y.; Yamamoto, N.G.; Wiehager, B.; Bogdanovic, N.; Winblad, B.; Sandebring-Matton, A.; Frykman, S.; et al. Monoamine oxidase B is elevated in Alzheimer disease neurons, is associated with γ-secretase and regulates neuronal amyloid β-peptide levels. Alzheimers Res. Ther. 2017, 9, 57. [Google Scholar] [CrossRef]
- Chen, G.-F.; Xu, T.-H.; Yan, Y.; Zhou, Y.-R.; Jiang, Y.; Melcher, K.; Xu, H.E. Amyloid beta: Structure, biology and structure-based therapeutic development. Acta Pharmacol. Sin. 2017, 38, 1205–1235. [Google Scholar] [CrossRef]
- Wang, S.; Zhang, S.; Wang, S.; Gao, P.; Dai, L. A comprehensive review on Pueraria: Insights on its chemistry and medicinal value. Biomed. Pharmacother. 2020, 131, 110734. [Google Scholar] [CrossRef]
- Seong, S.H.; Roy, A.; Jung, H.A.; Jung, H.J.; Choi, J.S. Protein tyrosine phosphatase 1B and α-glucosidase inhibitory activities of Pueraria lobata root and its constituents. J. Ethnopharmacol. 2016, 194, 706–716. [Google Scholar] [CrossRef]
- Xiao, B.; Sun, Z.; Cao, F.; Wang, L.; Liao, Y.; Liu, X.; Pan, R.; Chang, Q. Brain pharmacokinetics and the pharmacological effects on striatal neurotransmitter levels of Pueraria lobata isoflavonoids in rat. Front. Pharmacol. 2017, 8, 599. [Google Scholar] [CrossRef]
- Zhang, X.; Xiong, J.; Liu, S.; Wang, L.; Huang, J.; Liu, L.; Yang, J.; Zhang, G.; Guo, K.; Zhang, Z.; et al. Puerarin protects dopaminergic neurons in parkinson’s disease models. Neuroscience 2014, 280, 88–98. [Google Scholar] [CrossRef]
- Yan, B.; Wang, D.-Y.; Xing, D.-M.; Ding, Y.; Wang, R.-F.; Lei, F.; Du, L.-J. The antidepressant effect of ethanol extract of radix puerariae in mice exposed to cerebral ischemia reperfusion. Pharmacol. Biochem. Behav. 2004, 78, 319–325. [Google Scholar] [CrossRef]
- Tsugeno, Y.; Hirashiki, I.; Ogata, F.; Ito, A. Regions of the molecule responsible for substrate specificity of monoamine oxidase A and B: A chimeric enzyme analysis. J. Biochem. 1995, 118, 974–980. [Google Scholar] [CrossRef]
- Lineweaver, H.; Burk, D. The determination of enzyme dissociation constants. J. Am. Chem. Soc. 1934, 56, 658–666. [Google Scholar] [CrossRef]
- Naldi, M.; Fiori, J.; Pistolozzi, M.; Drake, A.F.; Bertucci, C.; Wu, R.; Mlynarczyk, K.; Filipek, S.; De Simone, A.; Andrisano, V. Amyloid β-peptide 25–35 self-assembly and its inhibition: A model undecapeptide system to gain atomistic and secondary structure details of the Alzheimer’s disease process and treatment. ACS Chem. Neurosci. 2012, 3, 952–962. [Google Scholar] [CrossRef]
- Son, S.-Y.; Ma, J.; Kondou, Y.; Yoshimura, M.; Yamashita, E.; Tsukihara, T. Structure of human monoamine oxidase A at 2.2-Å; resolution: The control of opening the entry for substrates/inhibitors. Proc. Natl. Acad. Sci. USA 2008, 105, 5739–5744. [Google Scholar] [CrossRef]
- Goodsell, D.S.; Morris, G.M.; Olson, A.J. Automated docking of flexible ligands: Applications of autodock. J. Mol. Recognit. 1996, 9, 1–5. [Google Scholar] [CrossRef]
- Leung, T.K.C.; Lim, L.; Lai, J.C.K. Differential effects of metal ions on type A and type B monoamine oxidase activities in rat brain and liver mitochondria. Metab. Brain Dis. 1992, 7, 139–146. [Google Scholar] [CrossRef]
- Woo, J.C.G.; Wang, X.; Silverman, R.B. Monoamine oxidase-catalyzed amine oxidation in organic solvents. J. Org. Chem. 1995, 60, 6235–6236. [Google Scholar] [CrossRef]
- Egashira, T. Studies on monoamine oxidase. Xviii. Enzymic properties of placental monoamine oxidase. Jpn. J. Pharmacol. 1976, 26, 493–500. [Google Scholar] [CrossRef]
- Valley, M.P.; Zhou, W.; Hawkins, E.M.; Shultz, J.; Cali, J.J.; Worzella, T.; Bernad, L.; Good, T.; Good, D.; Riss, T.L.; et al. A bioluminescent assay for monoamine oxidase activity. Anal. Biochem. 2006, 359, 238–246. [Google Scholar] [CrossRef]
- Zarmouh, N.O.; Messeha, S.S.; Elshami, F.M.; Soliman, K.F.A. Evaluation of the isoflavone genistein as reversible human monoamine oxidase-A and -B inhibitor. Evid. Based Complement. Alternat. Med. 2016, 2016, 1423052. [Google Scholar] [CrossRef]
- Ghose, A.K.; Viswanadhan, V.N.; Wendoloski, J.J. A knowledge-based approach in designing combinatorial or medicinal chemistry libraries for drug discovery. 1. A qualitative and quantitative characterization of known drug databases. J. Comb. Chem. 1999, 1, 55–68. [Google Scholar] [CrossRef]
- Wager, T.T.; Hou, X.; Verhoest, P.R.; Villalobos, A. Moving beyond rules: The development of a central nervous system multiparameter optimization (CNS MPO) approach to enable alignment of druglike properties. ACS Chem. Neurosci. 2010, 1, 435–449. [Google Scholar] [CrossRef]
- Lyman, R.L.; Bickoff, E.M.; Booth, A.N.; Livingston, A.L. Detection of coumestrol in leguminous plants. Arch. Biochem. Biophys. 1959, 80, 61–67. [Google Scholar] [CrossRef]
- Ha, J.; Kang, Y.-G.; Lee, T.; Kim, M.; Yoon, M.Y.; Lee, E.; Yang, X.; Kim, D.; Kim, Y.-J.; Lee, T.R.; et al. Comprehensive RNA sequencing and co-expression network analysis to complete the biosynthetic pathway of coumestrol, a phytoestrogen. Sci. Rep. 2019, 9, 1934. [Google Scholar] [CrossRef]
- Castro, C.C.; Pagnussat, A.S.; Orlandi, L.; Worm, P.; Moura, N.; Etgen, A.M.; Netto, A.C. Coumestrol has neuroprotective effects before and after global cerebral ischemia in female rats. Brain Res. 2012, 1474, 82–90. [Google Scholar] [CrossRef]
- Montero, G.; Arriagada, F.; Günther, G.; Bollo, S.; Mura, F.; Berríos, E.; Morales, J. Phytoestrogen coumestrol: Antioxidant capacity and its loading in albumin nanoparticles. Int. J. Pharm. 2019, 562, 86–95. [Google Scholar] [CrossRef]
- Zarmouh, N.O.; Eyunni, S.K.; Soliman, K.F.A. The benzopyrone Biochanin-A as a reversible, competitive, and selective monoamine oxidase B inhibitor. BMC Complement. Altern. Med. 2017, 17, 34. [Google Scholar] [CrossRef]
- Jeong, G.S.; Kang, M.-G.; Lee, J.Y.; Lee, S.R.; Park, D.; Cho, M.; Kim, H. Inhibition of butyrylcholinesterase and human monoamine oxidase-B by the coumarin glycyrol and liquiritigenin isolated from Glycyrrhiza uralensis. Molecules 2020, 25, 3896. [Google Scholar] [CrossRef]
- La Regina, G.; Silvestri, R.; Artico, M.; Lavecchia, A.; Novellino, E.; Befani, O.; Turini, P.; Agostinelli, E. New pyrrole inhibitors of monoamine oxidase: Synthesis, biological evaluation, and structural determinants of MAO-A and MAO-B selectivity. J. Med. Chem. 2007, 50, 922–931. [Google Scholar] [CrossRef]
- Geha, R.M.; Chen, K.; Wouters, J.; Ooms, F.; Shih, J.C. Analysis of conserved active site residues in monoamine oxidase A and B and their three-dimensional molecular modeling. J. Biol. Chem. 2002, 277, 17209–17216. [Google Scholar] [CrossRef]
- Liu, M.-H.; Tsuang, F.-Y.; Sheu, S.-Y.; Sun, J.-S.; Shih, C.-M. The protective effects of coumestrol against amyloid-beta peptide- and lipopolysaccharide-induced toxicity on mice astrocytes. Neurol. Res. 2011, 33, 663–672. [Google Scholar] [CrossRef] [PubMed]
- Koirala, P.; Seong, S.H.; Jung, H.A.; Choi, J.S. Comparative evaluation of the antioxidant and anti-Alzheimer’s disease potential of coumestrol and puerarol isolated from Pueraria lobata using molecular modeling studies. Molecules 2018, 23, 785. [Google Scholar] [CrossRef] [PubMed]
- Weinberg, R.P.; Koledova, V.V.; Shin, H.; Park, J.H.; Tan, Y.A.; Sinskey, A.J.; Sambanthamurthi, R.; Rha, C. Oil palm phenolics inhibit the in vitro aggregation of β-amyloid peptide into oligomeric complexes. Int. J. Alzheimers Dis. 2018, 2018, 7608038. [Google Scholar]
- Brenn, A.; Grube, M.; Peters, M.; Fischer, A.; Jedlitschky, G.; Kroemer, H.K.; Warzok, R.W.; Vogelgesang, S. Beta-amyloid downregulates MDR1-P-glycoprotein (Abcb1) expression at the blood-brain barrier in mice. Int. J. Alzheimers Dis. 2011, 2011, 690121. [Google Scholar] [CrossRef]
- Hartz, A.M.S.; Zhong, Y.; Wolf, A.; LeVine, H.; Miller, D.S.; Bauer, B. Aβ40 reduces P-glycoprotein at the blood–brain barrier through the ubiquitin–proteasome pathway. J. Neurosci. 2016, 36, 1930–1941. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Compounds | RT (min) | Measured Mass | Molecular Formula | Error (ppm) | Fragment Ions (m/z) a |
|---|---|---|---|---|---|
| Robinin | 21.12 | 739.2080 [M-H]− | C33H40O19 | −8.0 | 593.1519 |
| Quercetin-3-O-robinobioside | 24.38 | 609.1437 [M-H]− | C27H30O16 | −3.1 | 301.0340, 300.0283 |
| Rutin | 24.74 | 609.1437 [M-H]− | C27H30O16 | −3.1 | 301.0340, 300.0283 |
| Kaempferol-3-O-robinobioside | 26.94 | 593.1519 [M-H]− | C27H30O15 | 2.2 | 285.0408, 284.0315, 269.0437 |
| Nicotiflorin | 28.35 | 593.1519 [M-H]− | C27H30O15 | 2.2 | 285.0408 |
| Daidzin | 21.32 | 461.1108 [M + HCOOH-H]− | C21H20O9 | 5.2 | 415.1031 [M-H]−, 253.0523 |
| Genistin | 28.30 | 477.1048 [M + HCOOH-H]− | C22H22O12 | 3.1 | 431.0973 [M-H]−, 269.0474 |
| Daidzein | 39.57 | 253.0523 [M-H]− | C15H10O4 | 8.7 | 225.0543, 224.0486, 209.0623, 197.0588, 91.0185 |
| Genistein | 49.61 | 269.0474 [M-H]− | C15H10O5 | −4.8 | 241.0490, 225.0543, 224.0486, 201.0550, 159.0453, 133.0300 |
| Coumestrol | 49.76 | 267.0286 [M-H]− | C15H8O5 | −2.6 | 167.0480 |
| Compounds | hMAO-A | hMAO-B | SI a |
|---|---|---|---|
| IC50 (Mean ± SD, µM) | |||
| Robinin | >400 | >400 | - |
| Quercetin-3-O-robinobioside | >400 | >400 | - |
| Rutin | 387.12 ± 4.63 | >400 | - |
| Kaempferol-3-O-robinobioside | >400 | >400 | - |
| Nicotiflorine | >400 | >400 | - |
| Daidzin | >400 | >400 | - |
| Genistin | >400 | >400 | - |
| Daidzein | 304.05 ± 4.72 | 356.86 ± 1.05 | 0.85 |
| Genistein | 4.77 ± 0.51 | 3.42 ± 0.39 | 1.39 |
| Coumestrol | 1.99 ± 0.68 | 77.79 ± 2.10 | 0.02 |
| Selegiline hydrochloride b | 12.57 ± 0.51 | 0.38 ± 0.001 | 33.08 |
| Compounds | Inhibition Mode | Inhibition Constant (Ki, µM) | Binding Energy (kcal/mol) a | H-Bond Interaction Residues b | Other Interaction Residues b |
|---|---|---|---|---|---|
| Coumestrol | Competitive | 1.32 | −9.36 | FAD, Tyr444 | FAD and Phe352 (π–π T shaped), Tyr407 and Tyr444 (π–π stacking), Ile335, and Leu337 (π–alkyl) |
| Harmine c | ND | ND | −8.43 | ND | Tyr407 (π-π stacking, π-Alkyl), FAD (Van der Waals), Cys323 (π –sulfur), Ile335 (π–σ, π–alkyl), Tyr444, Ile180, and Leu337 (π–alkyl) |
| Model Name | Predicted Values |
|---|---|
| Log Po/w a | 2.94 |
| BBB penetration b | 0.76 |
| HIA c | 93.51 |
| P-glycoprotein | Non-inhibition |
| Carcino-rat/mouse | Negative |
| CNS MPO score d | 5.24 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Seong, S.H.; Kim, B.-R.; Cho, M.L.; Kim, T.-S.; Im, S.; Han, S.; Jeong, J.-W.; Jung, H.A.; Choi, J.S. Phytoestrogen Coumestrol Selectively Inhibits Monoamine Oxidase-A and Amyloid β Self-Aggregation. Nutrients 2022, 14, 3822. https://doi.org/10.3390/nu14183822
Seong SH, Kim B-R, Cho ML, Kim T-S, Im S, Han S, Jeong J-W, Jung HA, Choi JS. Phytoestrogen Coumestrol Selectively Inhibits Monoamine Oxidase-A and Amyloid β Self-Aggregation. Nutrients. 2022; 14(18):3822. https://doi.org/10.3390/nu14183822
Chicago/Turabian StyleSeong, Su Hui, Bo-Ram Kim, Myoung Lae Cho, Tae-Su Kim, Sua Im, Seahee Han, Jin-Woo Jeong, Hyun Ah Jung, and Jae Sue Choi. 2022. "Phytoestrogen Coumestrol Selectively Inhibits Monoamine Oxidase-A and Amyloid β Self-Aggregation" Nutrients 14, no. 18: 3822. https://doi.org/10.3390/nu14183822
APA StyleSeong, S. H., Kim, B.-R., Cho, M. L., Kim, T.-S., Im, S., Han, S., Jeong, J.-W., Jung, H. A., & Choi, J. S. (2022). Phytoestrogen Coumestrol Selectively Inhibits Monoamine Oxidase-A and Amyloid β Self-Aggregation. Nutrients, 14(18), 3822. https://doi.org/10.3390/nu14183822