
The Impact of Short-Chain Fatty Acids on Neonatal Regulatory T Cells


Abstract

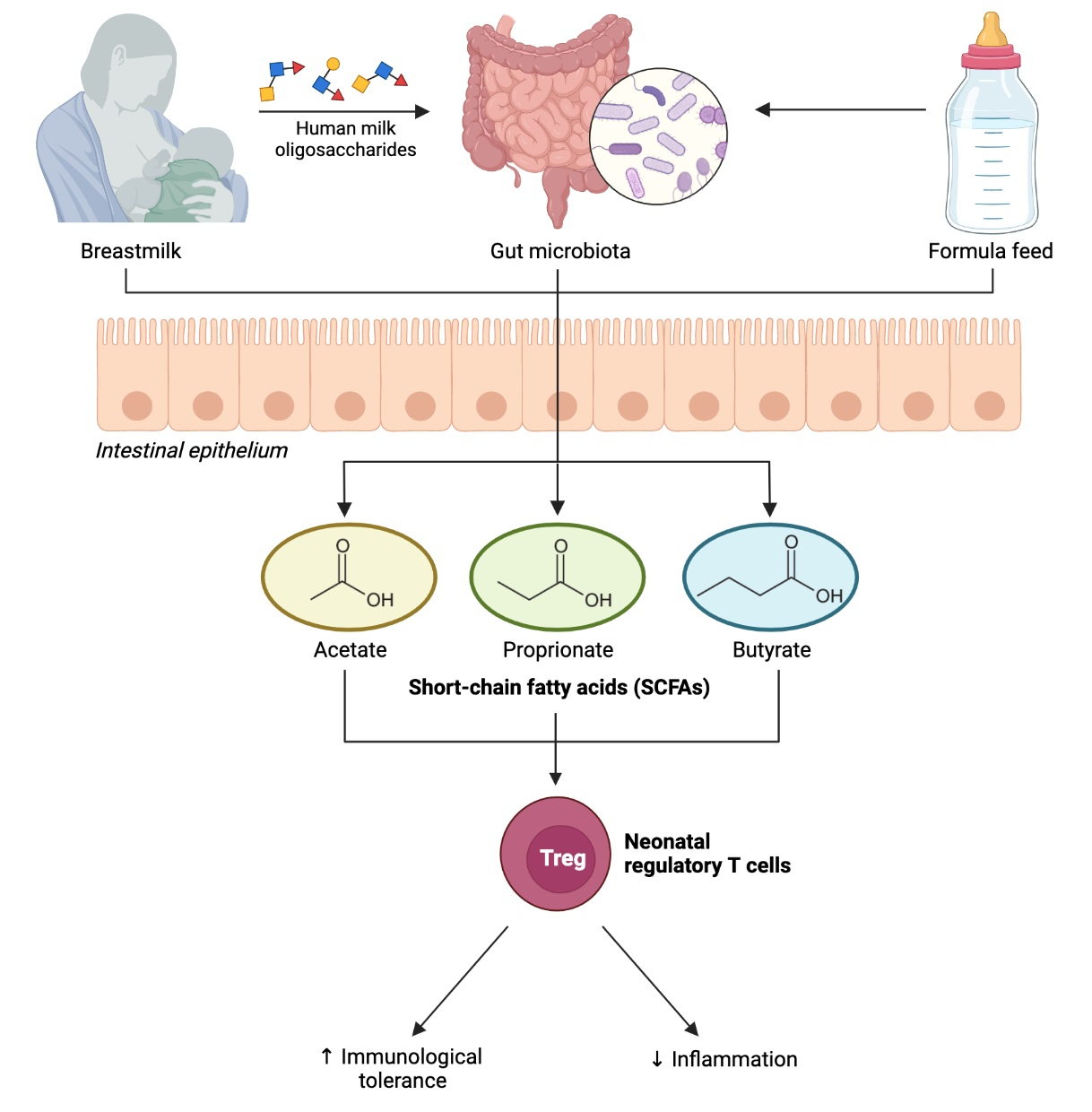{kind=link}
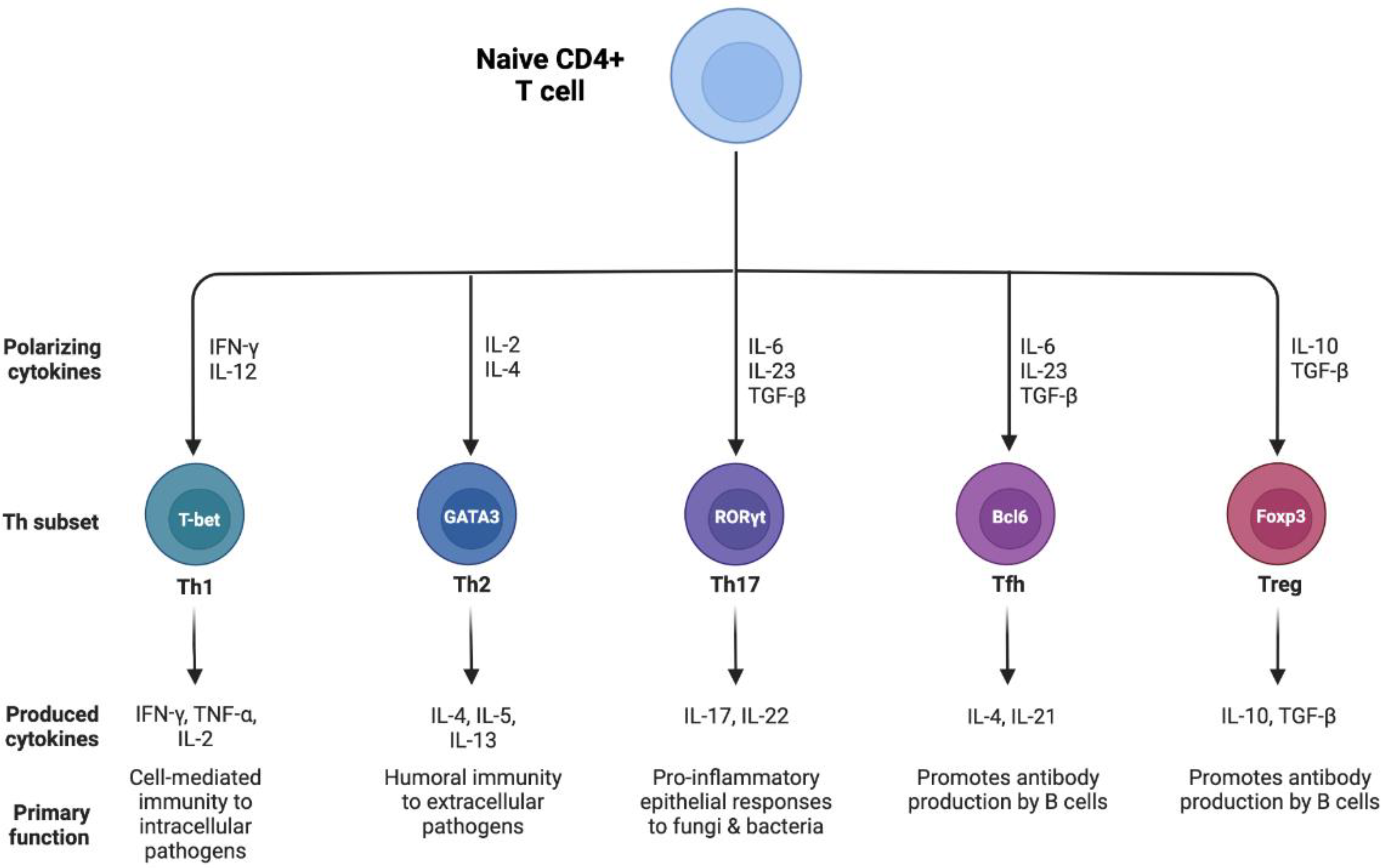{kind=link}
1. Introduction
2. T Cells in the Neonatal Period
3. Neonatal CD4+ T Cells
4. Neonatal Regulatory T Cells
5. Factors Influencing the Neonatal Regulatory T Cell Compartment
6. SCFA Levels in the Neonate
7. The Clinical Role of SCFAs in the Neonate
8. Future Directions of Research
- The description of how various types and concentrations of SCFAs affect neonatal Treg development and function compared to adults in culture and in vivo.
- The description of how the direct availability of SCFAs in breastmilk or formula milk, as well as SCFAs produced by HMO breakdown by the components of the microbiome, contribute to SCFA levels and composition in the neonate during the development.
- The dynamics of transition from HMOs to dietary fibre as fuel for SCFA production and the associated changes in microbiome composition during the weaning period and its implications on Treg function in the infant.
- The role of SCFAs in addressing inflammatory complications in preterm and term neonates, such as NEC, bronchopulmonary dysplasia or sepsis.
- The therapeutic potential of enteral or parenteral supplementation of SCFAs in neonates.
9. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Liu, L.; Johnson, H.L.; Cousens, S.; Perin, J.; Scott, S.; Lawn, J.E.; Rudan, I.; Campbell, H.; Cibulskis, R.; Li, M.; et al. Global, Regional, and National Causes of Child Mortality: An Updated Systematic Analysis for 2010 with Time Trends since 2000. Lancet 2012, 379, 2151–2161. [Google Scholar] [CrossRef]
- Moore, T.A.; Hanson, C.K.; Anderson-Berry, A. Colonization of the Gastrointestinal Tract in Neonates A Review. Child. Obes. Nutr. 2011, 3, 291–295. [Google Scholar] [CrossRef]
- Kalbermatter, C.; Fernandez Trigo, N.; Christensen, S.; Ganal-Vonarburg, S.C. Maternal Microbiota, Early Life Colonization and Breast Milk Drive Immune Development in the Newborn. Front. Immunol. 2021, 12, 1768. [Google Scholar] [CrossRef] [PubMed]
- Debock, I.; Flamand, V. Unbalanced Neonatal CD4+ T-Cell Immunity. Front. Immunol. 2014, 5, 393. [Google Scholar] [CrossRef] [PubMed]
- Basha, S.; Surendran, N.; Pichichero, M. Immune Responses in Neonates. Expert Rev. Clin. Immunol. 2014, 10, 1171. [Google Scholar] [CrossRef] [PubMed]
- Klein, L.; Kyewski, B.; Allen, P.M.; Hogquist, K.A. Positive and Negative Selection of the T Cell Repertoire: What Thymocytes See (and Don’t See). Nat. Rev. Immunol. 2014, 14, 377–391. [Google Scholar] [CrossRef]
- Fink, P.J. The Biology of Recent Thymic Emigrants. Annu. Rev. Immunol. 2013, 31, 31–50. [Google Scholar] [CrossRef]
- Semmes, E.C.; Chen, J.L.; Goswami, R.; Burt, T.D.; Permar, S.R.; Fouda, G.G. Understanding Early-Life Adaptive Immunity to Guide Interventions for Pediatric Health. Front. Immunol. 2021, 11, 595297. [Google Scholar] [CrossRef]
- Rechavi, E.; Lev, A.; Lee, Y.N.; Simon, A.J.; Yinon, Y.; Lipitz, S.; Amariglio, N.; Weisz, B.; Notarangelo, L.D.; Somech, R. Timely and Spatially Regulated Maturation of B and T Cell Repertoire during Human Fetal Development. Sci. Transl. Med. 2015, 7, 276ra25. [Google Scholar] [CrossRef]
- Thome, J.J.C.; Bickham, K.L.; Ohmura, Y.; Kubota, M.; Matsuoka, N.; Gordon, C.; Granot, T.; Griesemer, A.; Lerner, H.; Kato, T.; et al. Early Life Compartmentalization of Human T Cell Differentiation and Regulatory Function in Mucosal and Lymphoid Tissues. Nat. Med. 2016, 22, 72. [Google Scholar] [CrossRef]
- Olin, A.; Henckel, E.; Chen, Y.; Lakshmikanth, T.; Pou, C.; Mikes, J.; Gustafsson, A.; Bernhardsson, A.K.; Zhang, C.; Bohlin, K.; et al. Stereotypic Immune System Development in Newborn Children. Cell 2018, 174, 1277. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Zhu, J. CD4 T Helper Cell Subsets and Related Human Immunological Disorders. Int. J. Mol. Sci. 2020, 21, 8011. [Google Scholar] [CrossRef] [PubMed]
- Billingham, R.E.; Brent, L.; Medawar, P.B. Actively Acquired Tolerance of Foreign Cells. Nature 1953, 172, 603–606. [Google Scholar] [CrossRef] [PubMed]
- Nossal, G.J.V.; Pike, B.L. Functional Clonal Deletion in Immunological Tolerance to Major Histocompatibility Complex Antigens. Proc. Natl. Acad. Sci. USA 1981, 78, 3844. [Google Scholar] [CrossRef]
- Gammon, G.; Dunn, K.; Shastri, N.; Oki, A.; Wilbur, S.; Sercarz, E.E. Neonatal T-Cell Tolerance to Minimal Immunogenic Peptides Is Caused by Clonal Inactivation. Nature 1986, 319, 413–415. [Google Scholar] [CrossRef]
- Powell, T.J.; Streilein, J.W. Neonatal Tolerance Induction by Class II Alloantigens Activates IL-4-Secreting, Tolerogen-Responsive T Cells. J. Immunol. 1990, 144, 854–859. [Google Scholar]
- Abramowicz, D.; Vandervorst, P.; Bruyns, C.; Doutrelepont, J.-M.; Vandenabeele, P.; Goldman, M. Persistence of Anti-Donor Allohelper T Cells after Neonatal Induction of Allotolerance in Mice. Eur. J. Immunol. 1990, 20, 1647–1653. [Google Scholar] [CrossRef]
- Schurmans, S.; Heusser, C.H.; Qin, H.Y.; Merino, J.; Brighouse, G.; Lambert, P.H. In Vivo Effects of Anti-IL-4 Monoclonal Antibody on Neonatal Induction of Tolerance and on an Associated Autoimmune Syndrome. J. Immunol. 1990, 145, 2465–2473. [Google Scholar]
- Li, L.; Lee, H.H.; Bell, J.J.; Gregg, R.K.; Ellis, J.S.; Gessner, A.; Zaghouani, H. IL-4 Utilizes an Alternative Receptor to Drive Apoptosis of Th1 Cells and Skews Neonatal Immunity toward Th2. Immunity 2004, 20, 429–440. [Google Scholar] [CrossRef]
- Debock, I.; Delbauve, S.; Dubois, A.; Pétein, M.; Leo, O.; Goldman, M.; Flamand, V. Th17 Alloimmunity Prevents Neonatal Establishment of Lymphoid Chimerism in IL-4-Deprived Mice. Am. J. Transpl. 2012, 12, 81–89. [Google Scholar] [CrossRef]
- Hofstetter, H.H.; Kovalovsky, A.; Shive, C.L.; Lehmann, P.v.; Forsthuber, T.G. Neonatal Induction of Myelin-Specific Th1/Th17 Immunity Does Not Result in Experimental Autoimmune Encephalomyelitis and Can Protect against the Disease in Adulthood. J. Neuroimmunol. 2007, 187, 20–30. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Martinez, X.; Brandt, C.; Saddallah, F.; Tougne, C.; Barrios, C.; Wild, F.; Dougan, G.; Lambert, P.H.; Siegrist, C.A. DNA Immunization Circumvents Deficient Induction of T Helper Type 1 and Cytotoxic T Lymphocyte Responses in Neonates and during Early Life. Proc. Natl. Acad. Sci. USA 1997, 94, 8726–8731. [Google Scholar] [CrossRef] [PubMed]
- Brazolot Millan, C.L.; Weeratna, R.; Krieg, A.M.; Siegrist, C.A.; Davis, H.L. CpG DNA Can Induce Strong Th1 Humoral and Cell-Mediated Immune Responses against Hepatitis B Surface Antigen in Young Mice. Proc. Natl. Acad. Sci. USA 1998, 95, 15553–15558. [Google Scholar] [CrossRef] [PubMed]
- Forsthuber, T.; Yip, H.C.; Lehmann, P.V. Induction of TH1 and TH2 Immunity in Neonatal Mice. Science 1996, 271, 1728–1730. [Google Scholar] [CrossRef] [PubMed]
- Prescott, S.L.; Macaubas, C.; Smallacombe, T.; Holt, B.J.; Sly, P.D.; Holt, P.G. Development of Allergen-Specific T-Cell Memory in Atopic and Normal Children. Lancet 1999, 353, 196–200. [Google Scholar] [CrossRef]
- Michaëlsson, J.; Mold, J.E.; McCune, J.M.; Nixon, D.F. Regulation of T Cell Responses in the Developing Human Fetus. J. Immunol. 2006, 176, 5741–5748. [Google Scholar] [CrossRef] [PubMed]
- Burt, T.D. Fetal Regulatory T Cells and Peripheral Immune Tolerance In utero: Implications for Development and Disease. Am. J. Reprod. Immunol. 2013, 69, 346–358. [Google Scholar] [CrossRef]
- Wood, H.; Acharjee, A.; Pearce, H.; Quraishi, M.N.; Powell, R.; Rossiter, A.; Beggs, A.; Ewer, A.; Moss, P.; Toldi, G. Breastfeeding Promotes Early Neonatal Regulatory T-Cell Expansion and Immune Tolerance of Non-Inherited Maternal Antigens. Allergy 2021, 76, 2447–2460. [Google Scholar] [CrossRef]
- Gao, Q.; Rouse, T.M.; Kazmerzak, K.; Field, E.H. CD4+CD25+ Cells Regulate CD8 Cell Anergy in Neonatal Tolerant Mice. Transplantation 1999, 68, 1891–1897. [Google Scholar] [CrossRef]
- Bettelli, E.; Carrier, Y.; Gao, W.; Korn, T.; Strom, T.B.; Oukka, M.; Weiner, H.L.; Kuchroo, V.K. Reciprocal Developmental Pathways for the Generation of Pathogenic Effector TH17 and Regulatory T Cells. Nature 2006, 441, 235–238. [Google Scholar] [CrossRef]
- Bronevetsky, Y.; Burt, T.D.; McCune, J.M. Lin28b Regulates Fetal Regulatory T Cell Differentiation through Modulation of TGF-β Signaling. J. Immunol. 2016, 197, 4344–4350. [Google Scholar] [CrossRef] [PubMed]
- Mold, J.E.; Michaëlsson, J.; Burt, T.D.; Muench, M.O.; Beckerman, K.P.; Busch, M.P.; Lee, T.H.; Nixon, D.F.; McCune, J.M. Maternal Alloantigens Promote the Development of Tolerogenic Fetal Regulatory T Cells In utero. Science 2008, 322, 1562–1565. [Google Scholar] [CrossRef]
- Ng, M.S.F.; Roth, T.L.; Mendoza, V.F.; Marson, A.; Burt, T.D. Helios Enhances the Preferential Differentiation of Human Fetal CD4 + Naïve T Cells into Regulatory T Cells. Sci. Immunol. 2019, 4, eaav5947. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Miyahara, Y.; Guo, Z.; Khattar, M.; Stepkowski, S.M.; Chen, W. “Default” Generation of Neonatal Regulatory T Cells. J. Immunol. 2010, 185, 71–78. [Google Scholar] [CrossRef] [PubMed]
- de Roock, S.; Hoeks, S.B.E.A.; Meurs, L.; Steur, A.; Hoekstra, M.O.; Prakken, B.J.; Boes, M.; de Kleer, I.M. Critical Role for Programmed Death 1 Signaling and Protein Kinase B in Augmented Regulatory T-Cell Induction in Cord Blood. J. Allergy Clin. Immunol. 2011, 128, 1369–1371. [Google Scholar] [CrossRef] [PubMed]
- Frese, S.A.; Hutton, A.A.; Contreras, L.N.; Shaw, C.A.; Palumbo, M.C.; Casaburi, G.; Xu, G.; Davis, J.C.C.; Lebrilla, C.B.; Henrick, B.M.; et al. Persistence of Supplemented Bifidobacterium Longum Subsp. Infantis EVC001 in Breastfed Infants. mSphere 2017, 2, e00501–e00517. [Google Scholar] [CrossRef]
- Thorburn, A.N.; McKenzie, C.I.; Shen, S.; Stanley, D.; MacIa, L.; Mason, L.J.; Roberts, L.K.; Wong, C.H.Y.; Shim, R.; Robert, R.; et al. Evidence That Asthma Is a Developmental Origin Disease Influenced by Maternal Diet and Bacterial Metabolites. Nat. Commun. 2015, 6, 7320. [Google Scholar] [CrossRef]
- Gensollen, T.; Iyer, S.S.; Kasper, D.L.; Blumberg, R.S. How Colonization by Microbiota in Early Life Shapes the Immune System. Science 2016, 352, 539–544. [Google Scholar] [CrossRef]
- Sanidad, K.Z.; Zeng, M.Y. Neonatal Gut Microbiome and Immunity. Curr. Opin. Microbiol. 2020, 56, 30. [Google Scholar] [CrossRef]
- Lathrop, S.K.; Bloom, S.M.; Rao, S.M.; Nutsch, K.; Lio, C.W.; Santacruz, N.; Peterson, D.A.; Stappenbeck, T.S.; Hsieh, C.S. Peripheral Education of the Immune System by Colonic Commensal Microbiota. Nature 2011, 478, 250–254. [Google Scholar] [CrossRef]
- Arpaia, N.; Campbell, C.; Fan, X.; Dikiy, S.; van der Veeken, J.; Deroos, P.; Liu, H.; Cross, J.R.; Pfeffer, K.; Coffer, P.J.; et al. Metabolites Produced by Commensal Bacteria Promote Peripheral Regulatory T-Cell Generation. Nature 2013, 504, 451–455. [Google Scholar] [CrossRef] [PubMed]
- Rudd, B.D. Neonatal T Cells: A Reinterpretation. Annu. Rev. Immunol. 2020, 38, 229. [Google Scholar] [CrossRef] [PubMed]
- Underwood, M.A.; Mukhopadhyay, S.; Lakshminrusimha, S.; Bevins, C.L. Neonatal Intestinal Dysbiosis. J. Perinatol. 2020, 40, 1597–1608. [Google Scholar] [CrossRef]
- Dowds, C.M.; Blumberg, R.S.; Zeissig, S. Control of intestinal homeostasis through crosstalk between natural killer T cells and the intestinal microbiota. Clin. Immunol. 2015, 159, 128–133. [Google Scholar] [CrossRef]
- Park, J.; Kim, M.; Kang, S.G.; Jannasch, A.H.; Cooper, B.; Patterson, J.; Kim, C.H. Short-Chain Fatty Acids Induce Both Effector and Regulatory T Cells by Suppression of Histone Deacetylases and Regulation of the MTOR-S6K Pathway. Mucosal. Immunol. 2015, 8, 80–93. [Google Scholar] [CrossRef] [PubMed]
- Phillips-Farfán, B.; Gómez-Chávez, F.; Medina-Torres, E.A.; Vargas-Villavicencio, J.A.; Carvajal-Aguilera, K.; Camacho, L. Microbiota Signals during the Neonatal Period Forge Life-Long Immune Responses. Int. J. Mol. Sci. 2021, 22, 8162. [Google Scholar] [CrossRef]
- Kaisar, M.M.M.; Pelgrom, L.R.; van der Ham, A.J.; Yazdanbakhsh, M.; Everts, B. Butyrate Conditions Human Dendritic Cells to Prime Type 1 Regulatory T Cells via Both Histone Deacetylase Inhibition and G Protein-Coupled Receptor 109A Signaling. Front. Immunol. 2017, 8, 1429. [Google Scholar] [CrossRef]
- Bergman, E.N. Energy Contributions of Volatile Fatty Acids from the Gastrointestinal Tract in Various Species. Physiol. Rev. 1990, 70, 567–590. [Google Scholar] [CrossRef]
- Bridgman, S.L.; Azad, M.B.; Field, C.J.; Haqq, A.M.; Becker, A.B.; Mandhane, P.J.; Subbarao, P.; Turvey, S.E.; Sears, M.R.; Scott, J.A.; et al. Fecal Short-Chain Fatty Acid Variations by Breastfeeding Status in Infants at 4 Months: Differences in Relative versus Absolute Concentrations. Front. Nutr. 2017, 4, 11. [Google Scholar] [CrossRef]
- Corrêa-Oliveira, R.; Fachi, J.L.; Vieira, A.; Sato, F.T.; Vinolo, M.A.R. Regulation of Immune Cell Function by Short-Chain Fatty Acids. Clin. Transl. Immunol. 2016, 5, e73. [Google Scholar] [CrossRef]
- Furusawa, Y.; Obata, Y.; Fukuda, S.; Endo, T.A.; Nakato, G.; Takahashi, D.; Nakanishi, Y.; Uetake, C.; Kato, K.; Kato, T.; et al. Commensal Microbe-Derived Butyrate Induces the Differentiation of Colonic Regulatory T Cells. Nature 2013, 504, 446–450. [Google Scholar] [CrossRef] [PubMed]
- Smith, P.M.; Howitt, M.R.; Panikov, N.; Michaud, M.; Gallini, C.A.; Bohlooly-Y, M.; Glickman, J.N.; Garrett, W.S. The Microbial Metabolites, Short-Chain Fatty Acids, Regulate Colonic T Reg Cell Homeostasis. Science 2013, 341, 569–573. [Google Scholar] [CrossRef]
- Tan, J.; McKenzie, C.; Potamitis, M.; Thorburn, A.N.; Mackay, C.R.; Macia, L. The Role of Short-Chain Fatty Acids in Health and Disease. Adv. Immunol. 2014, 121, 91–119. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Thangaraju, M.; Prasad, P.D.; Martin, P.M.; Lambert, N.A.; Boettger, T.; Offermanns, S.; Ganapathy, V. Blockade of Dendritic Cell Development by Bacterial Fermentation Products Butyrate and Propionate through a Transporter (Slc5a8)-Dependent Inhibition of Histone Deacetylases. J. Biol. Chem. 2010, 285, 27601–27608. [Google Scholar] [CrossRef] [PubMed]
- Tao, R.; de Zoeten, E.F.; Özkaynak, E.; Chen, C.; Wang, L.; Porrett, P.M.; Li, B.; Turka, L.A.; Olson, E.N.; Greene, M.I.; et al. Deacetylase Inhibition Promotes the Generation and Function of Regulatory T Cells. Nat. Med. 2007, 13, 1299–1307. [Google Scholar] [CrossRef]
- Chang, P.V.; Hao, L.; Offermanns, S.; Medzhitov, R. The Microbial Metabolite Butyrate Regulates Intestinal Macrophage Function via Histone Deacetylase Inhibition. Proc. Natl. Acad. Sci. USA 2014, 111, 2247–2252. [Google Scholar] [CrossRef]
- Victora, C.G.; Bahl, R.; Barros, A.J.D.; França, G.V.A.; Horton, S.; Krasevec, J.; Murch, S.; Sankar, M.J.; Walker, N.; Rollins, N.C.; et al. Breastfeeding in the 21st Century: Epidemiology, Mechanisms, and Lifelong Effect. Lancet 2016, 387, 475–490. [Google Scholar] [CrossRef]
- Stinson, L.F.; Gay, M.C.L.; Koleva, P.T.; Eggesbø, M.; Johnson, C.C.; Wegienka, G.; du Toit, E.; Shimojo, N.; Munblit, D.; Campbell, D.E.; et al. Human Milk From Atopic Mothers Has Lower Levels of Short Chain Fatty Acids. Front. Immunol. 2020, 11, 1427. [Google Scholar] [CrossRef]
- Prentice, P.M.; Schoemaker, M.H.; Vervoort, J.; Hettinga, K.; Lambers, T.T.; van Tol, E.A.F.; Acerini, C.L.; Olga, L.; Petry, C.J.; Hughes, I.A.; et al. Human Milk Short-Chain Fatty Acid Composition Is Associated with Adiposity Outcomes in Infants. J. Nutr. 2019, 149, 716–722. [Google Scholar] [CrossRef]
- Smilowitz, J.T.; O’Sullivan, A.; Barile, D.; German, J.B.; Lönnerdal, B.; Slupsky, C.M. The Human Milk Metabolome Reveals Diverse Oligosaccharide Profiles. J. Nutr. 2013, 143, 1709–1718. [Google Scholar] [CrossRef]
- Wu, J.; Domellöf, M.; Zivkovic, A.M.; Larsson, G.; Öhman, A.; Nording, M.L. NMR-Based Metabolite Profiling of Human Milk: A Pilot Study of Methods for Investigating Compositional Changes during Lactation. Biochem. Biophys. Res. Commun. 2016, 469, 626–632. [Google Scholar] [CrossRef]
- Bode, L. The Functional Biology of Human Milk Oligosaccharides. Early Hum. Dev. 2015, 91, 619–622. [Google Scholar] [CrossRef] [PubMed]
- Laursen, M.F.; Sakanaka, M.; von Burg, N.; Mörbe, U.; Andersen, D.; Moll, J.M.; Pekmez, C.T.; Rivollier, A.; Michaelsen, K.F.; Mølgaard, C.; et al. Bifidobacterium Species Associated with Breastfeeding Produce Aromatic Lactic Acids in the Infant Gut. Nat. Microbiol. 2021, 6, 1367–1382. [Google Scholar] [CrossRef] [PubMed]
- Henrick, B.M.; Rodriguez, L.; Lakshmikanth, T.; Pou, C.; Henckel, E.; Arzoomand, A.; Olin, A.; Wang, J.; Mikes, J.; Tan, Z.; et al. Bifidobacteria-mediated immune system imprinting early in life. Cell 2021, 184, 3884–3898.e11. [Google Scholar] [CrossRef]
- Pokusaeva, K.; Fitzgerald, G.F.; van Sinderen, D. Carbohydrate Metabolism in Bifidobacteria. Genes Nutr. 2011, 6, 285–306. [Google Scholar] [CrossRef]
- Gray, L.E.K.; O’Hely, M.; Ranganathan, S.; Sly, P.D.; Vuillermin, P. The Maternal Diet, Gut Bacteria, and Bacterial Metabolites during Pregnancy Influence Offspring Asthma. Front. Immunol. 2017, 8, 365. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Du, W.; Xiao, S.; Zeng, B.; She, X.; Liu, D.; Du, H.; Li, L.; Li, F.; Ai, Q.; et al. Colonization of Fecal Microbiota from Patients with Neonatal Necrotizing Enterocolitis Exacerbates Intestinal Injury in Germfree Mice Subjected to Necrotizing Enterocolitis-Induction Protocol via Alterations in Butyrate and Regulatory T Cells. J. Transl. Med. 2021, 19, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.K.; Meng, Q.; Sadowitz, B.D.; Kollisch-Singule, M.; Yepuri, N.; Satalin, J.; Gatto, L.A.; Nieman, G.F.; Cooney, R.N.; Clark, D. Enteral Administration of Bacteria Fermented Formula in Newborn Piglets: A High Fidelity Model for Necrotizing Enterocolitis (NEC). PLoS ONE 2018, 13, e0201172. [Google Scholar] [CrossRef]
- Nafday, S.M.; Chen, W.; Peng, L.; Babyatsky, M.W.; Holzman, I.R.; Lin, J. Short-Chain Fatty Acids Induce Colonic Mucosal Injury in Rats with Various Postnatal Ages. Pediatr. Res. 2005, 57, 201–204. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chun, J.; Toldi, G. The Impact of Short-Chain Fatty Acids on Neonatal Regulatory T Cells. Nutrients 2022, 14, 3670. https://doi.org/10.3390/nu14183670
Chun J, Toldi G. The Impact of Short-Chain Fatty Acids on Neonatal Regulatory T Cells. Nutrients. 2022; 14(18):3670. https://doi.org/10.3390/nu14183670
Chicago/Turabian StyleChun, Jessica, and Gergely Toldi. 2022. "The Impact of Short-Chain Fatty Acids on Neonatal Regulatory T Cells" Nutrients 14, no. 18: 3670. https://doi.org/10.3390/nu14183670
APA StyleChun, J., & Toldi, G. (2022). The Impact of Short-Chain Fatty Acids on Neonatal Regulatory T Cells. Nutrients, 14(18), 3670. https://doi.org/10.3390/nu14183670