
Anti-Colorectal Cancer Effects of Inonotus hispidus (Bull.: Fr.) P. Karst. Spore Powder through Regulation of Gut Microbiota-Mediated JAK/STAT Signaling

Abstract
:1. Introduction
2. Materials and Methods
2.1. IHS Component Analysis
2.1.1. Nutrients
2.1.2. Minerals and Heavy Metals
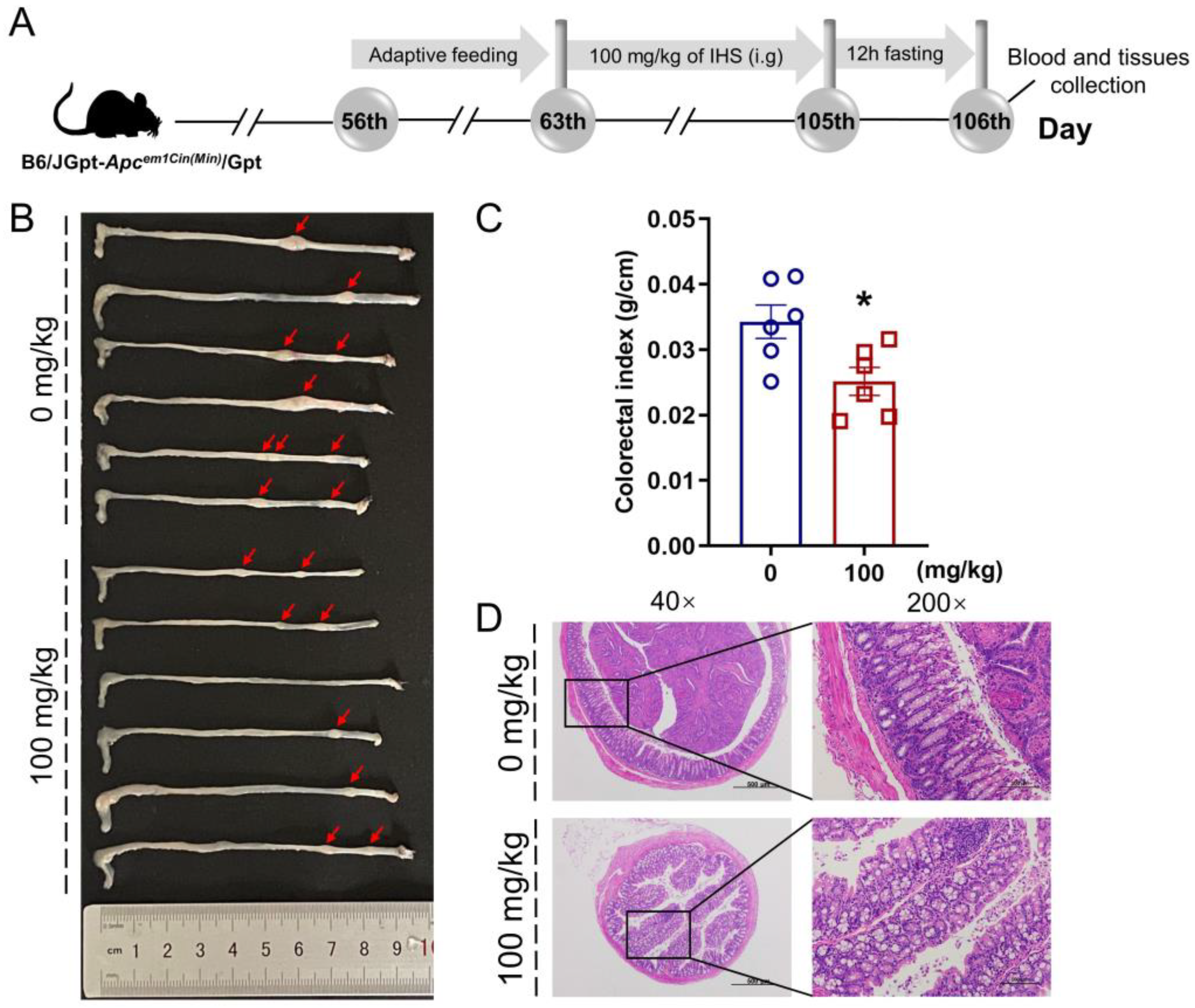
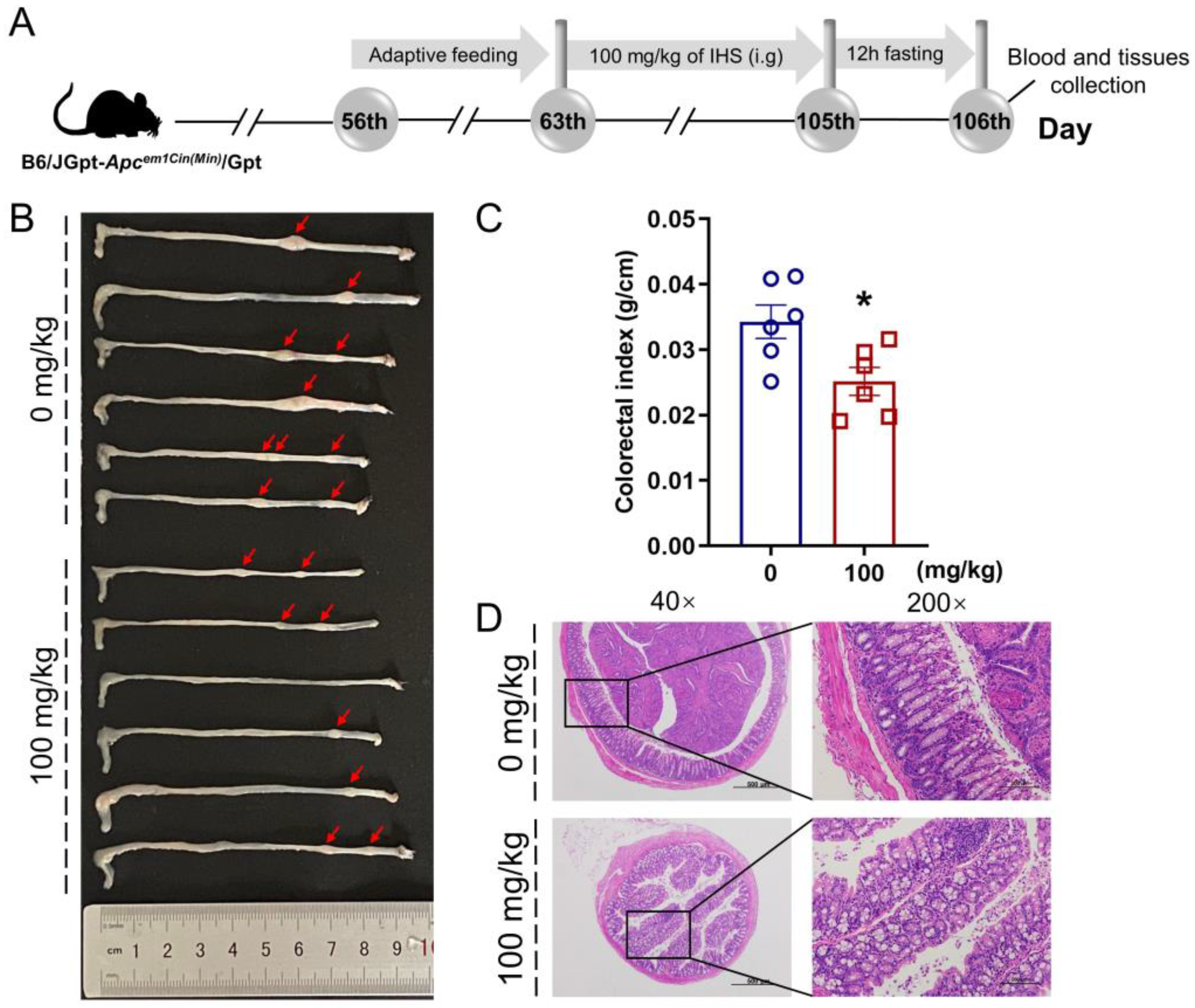
2.2. Animal Experimental
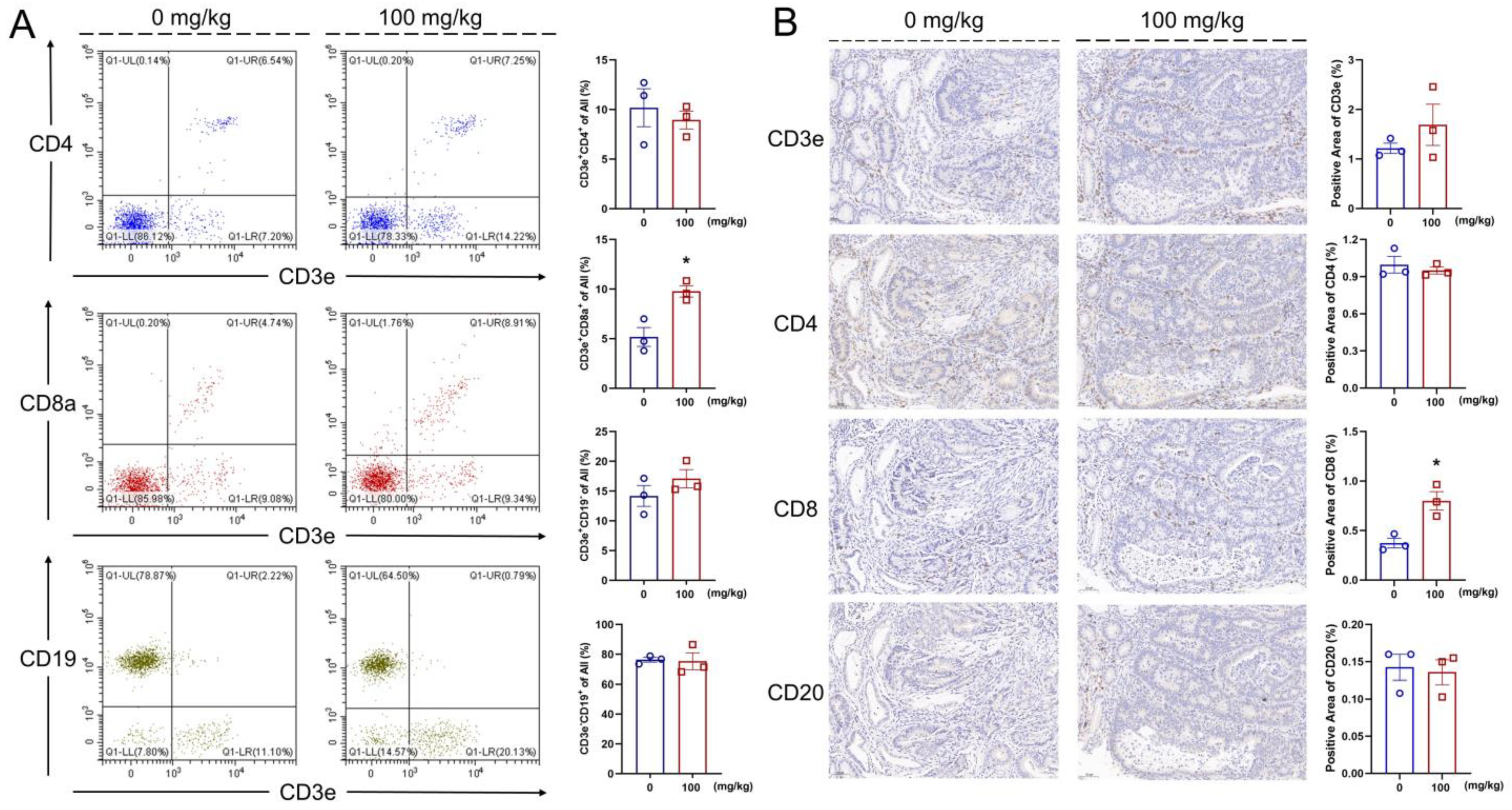
2.3. Flow Cytometry
2.4. Hematoxylin and Eosin (H&E) Staining
2.5. Immunohistochemistry (IHC)
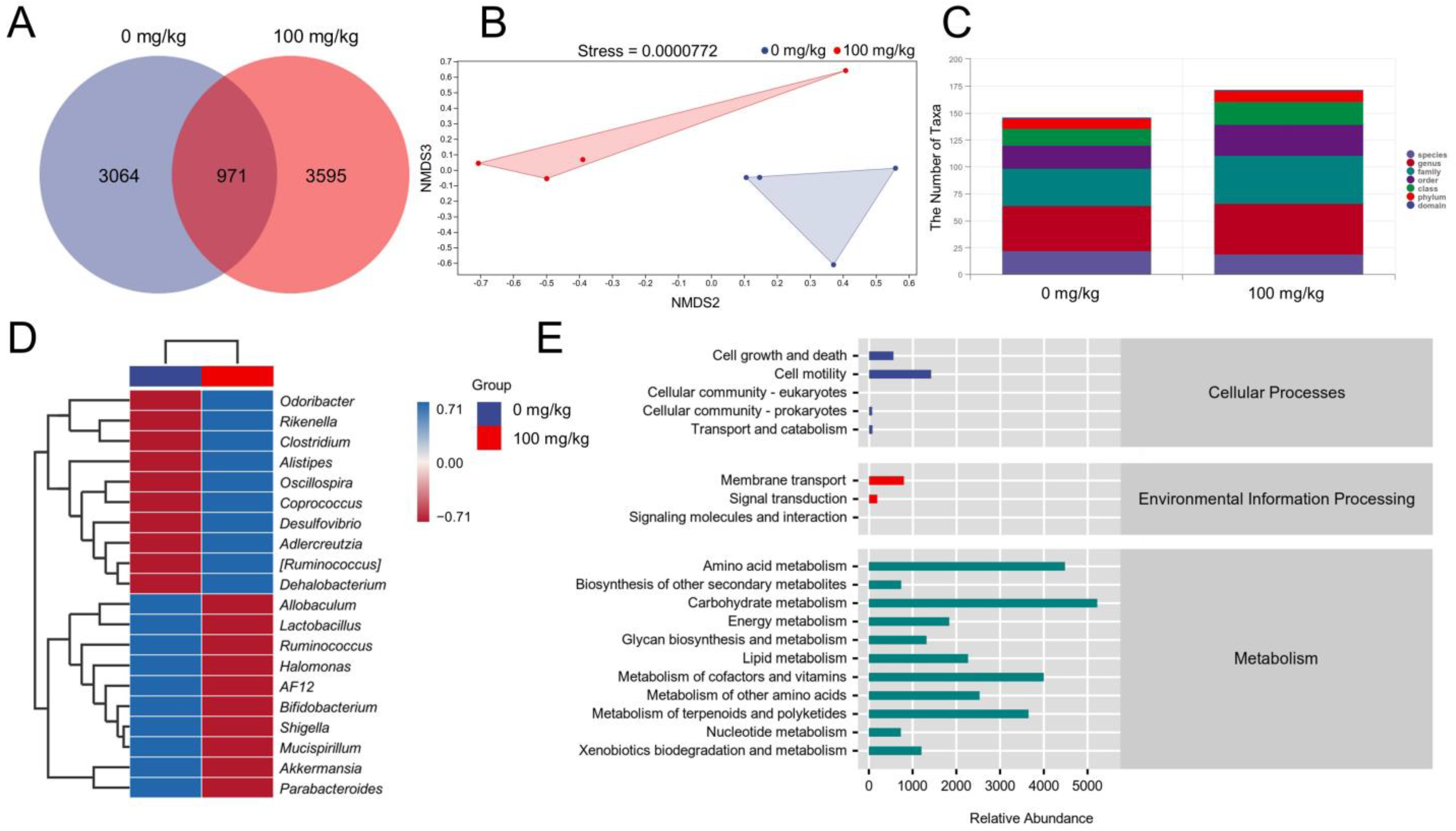
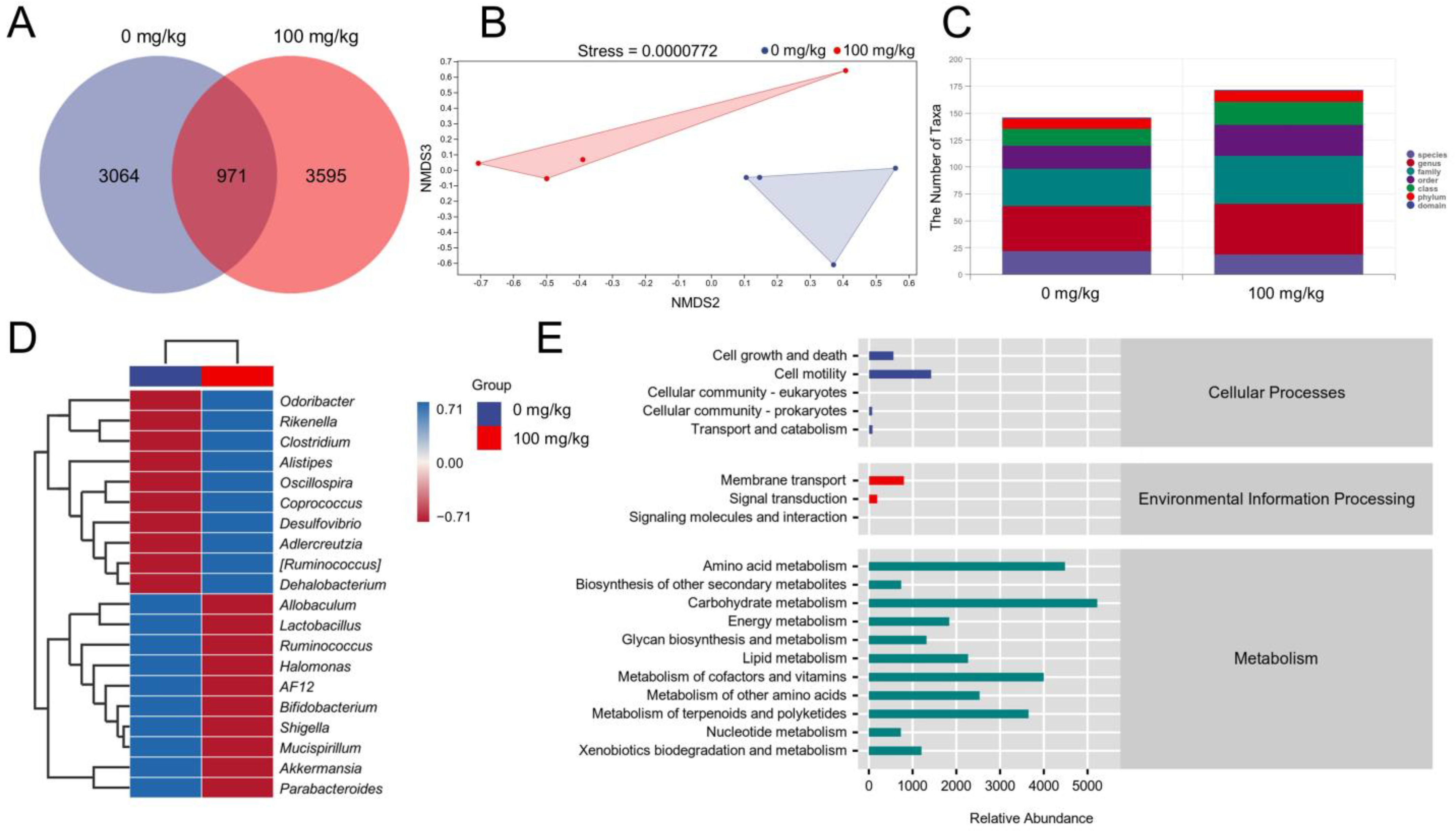
2.6. Gut Microbiota Analysis
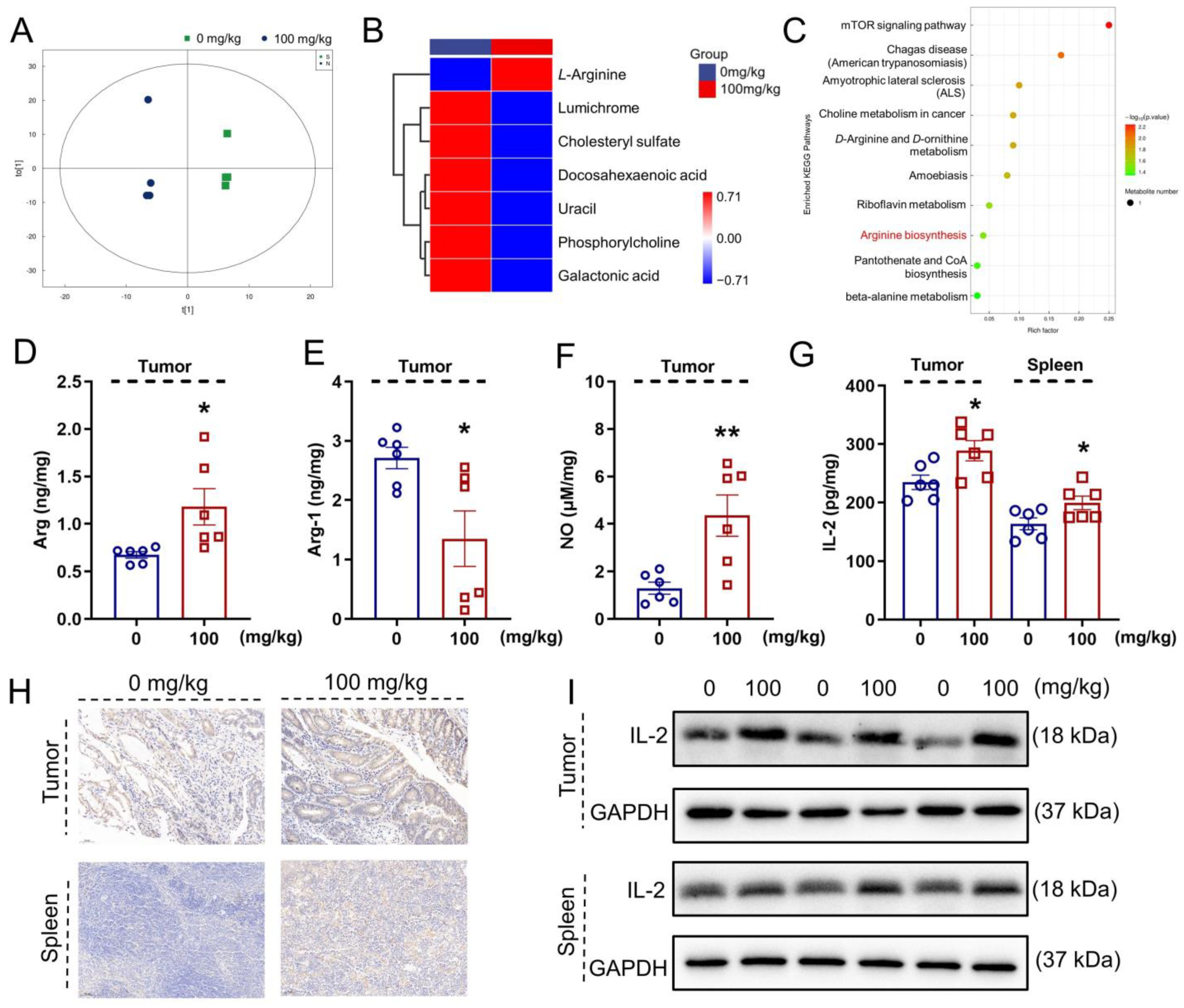
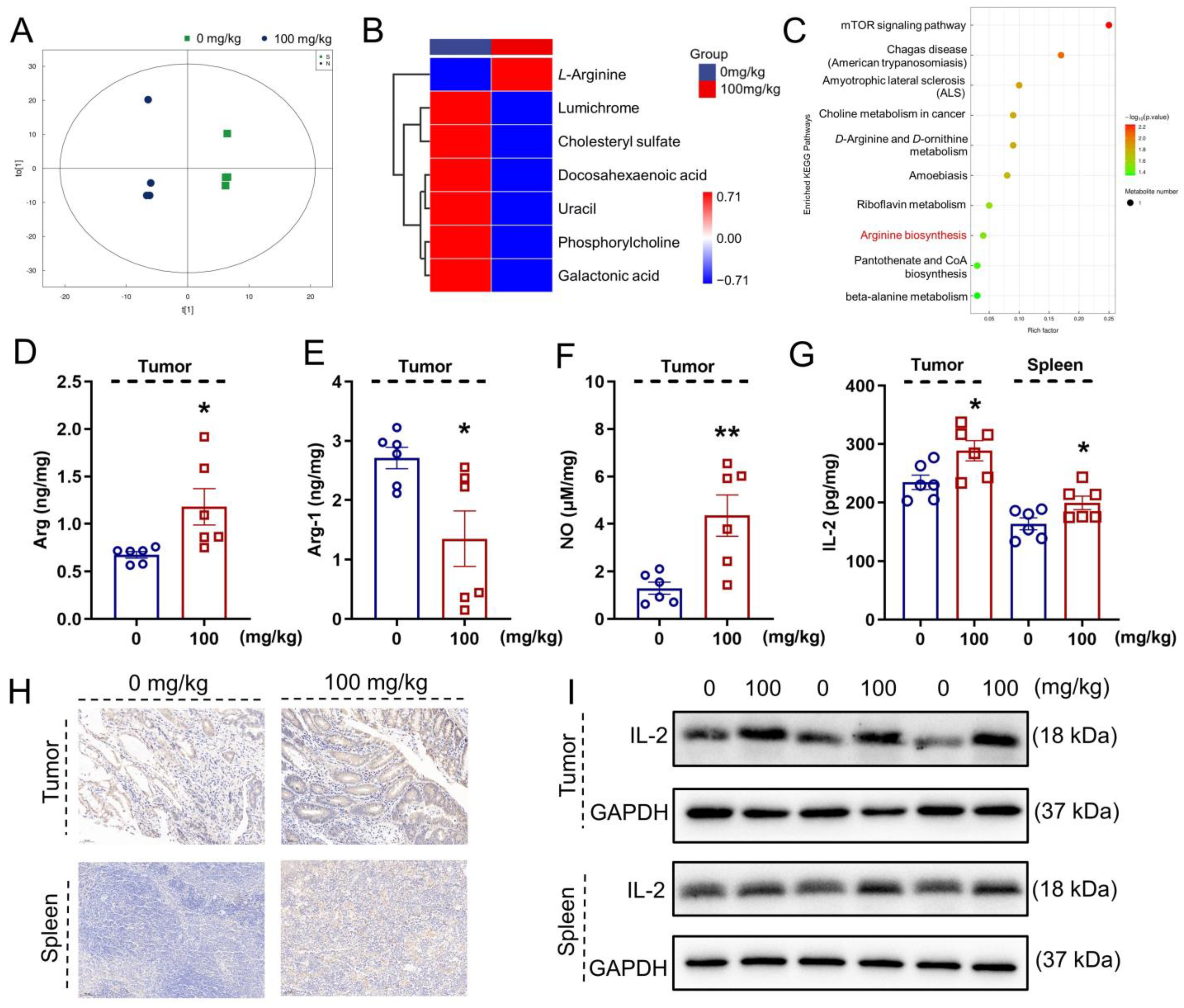
2.7. Metabolomics Analysis
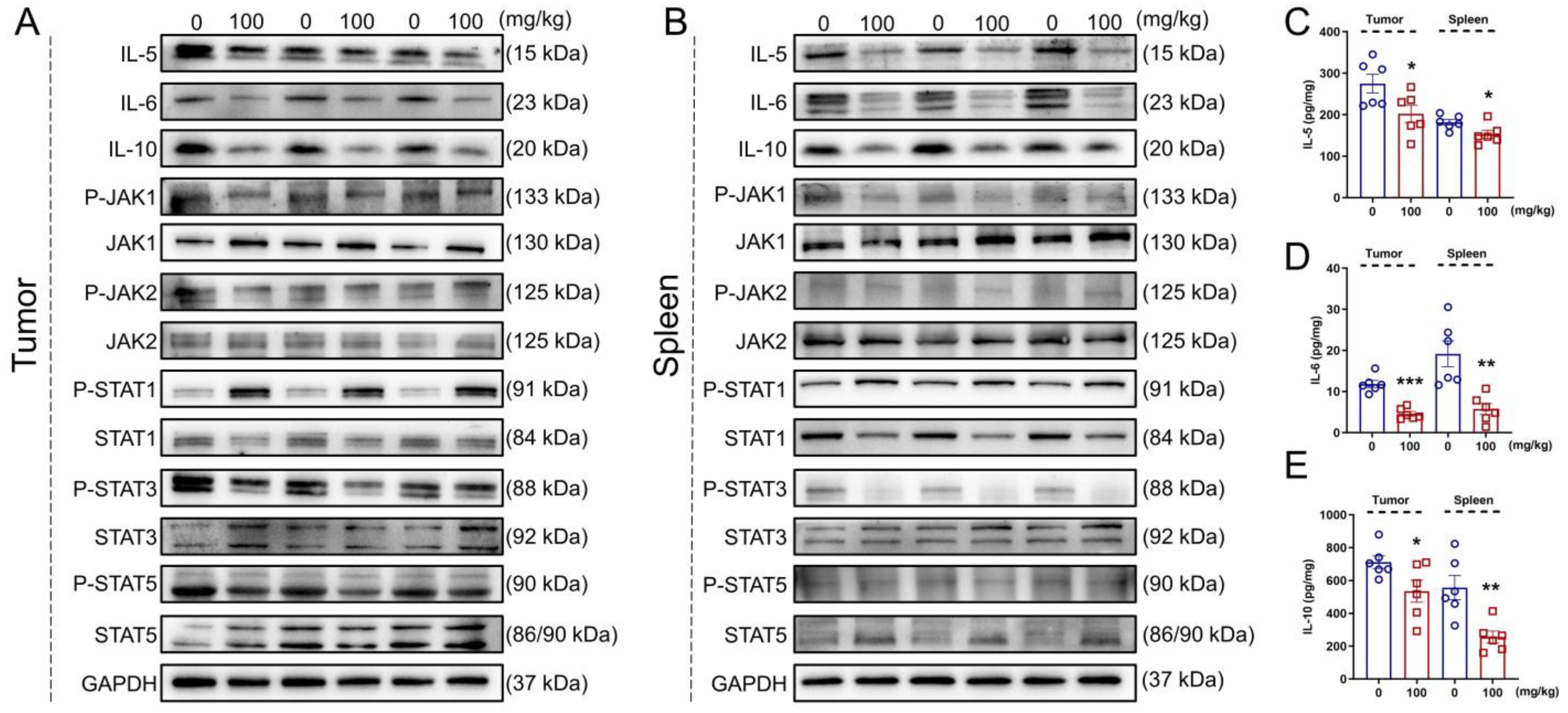
2.8. Cytokine Detection
2.9. Western Blot
2.10. Statistical Analyses
3. Results
3.1. Main IHS Compounds
3.2. IHS Restricts Tumor Growth in ApcMin/+ Mice
3.3. IHS Alters the Gut Microbiota of ApcMin/+ Mice
3.4. IHS Influences Serum Metabolites in ApcMin/+ Mice
3.5. IHS Increases the Abundance of CD8+ T Cells
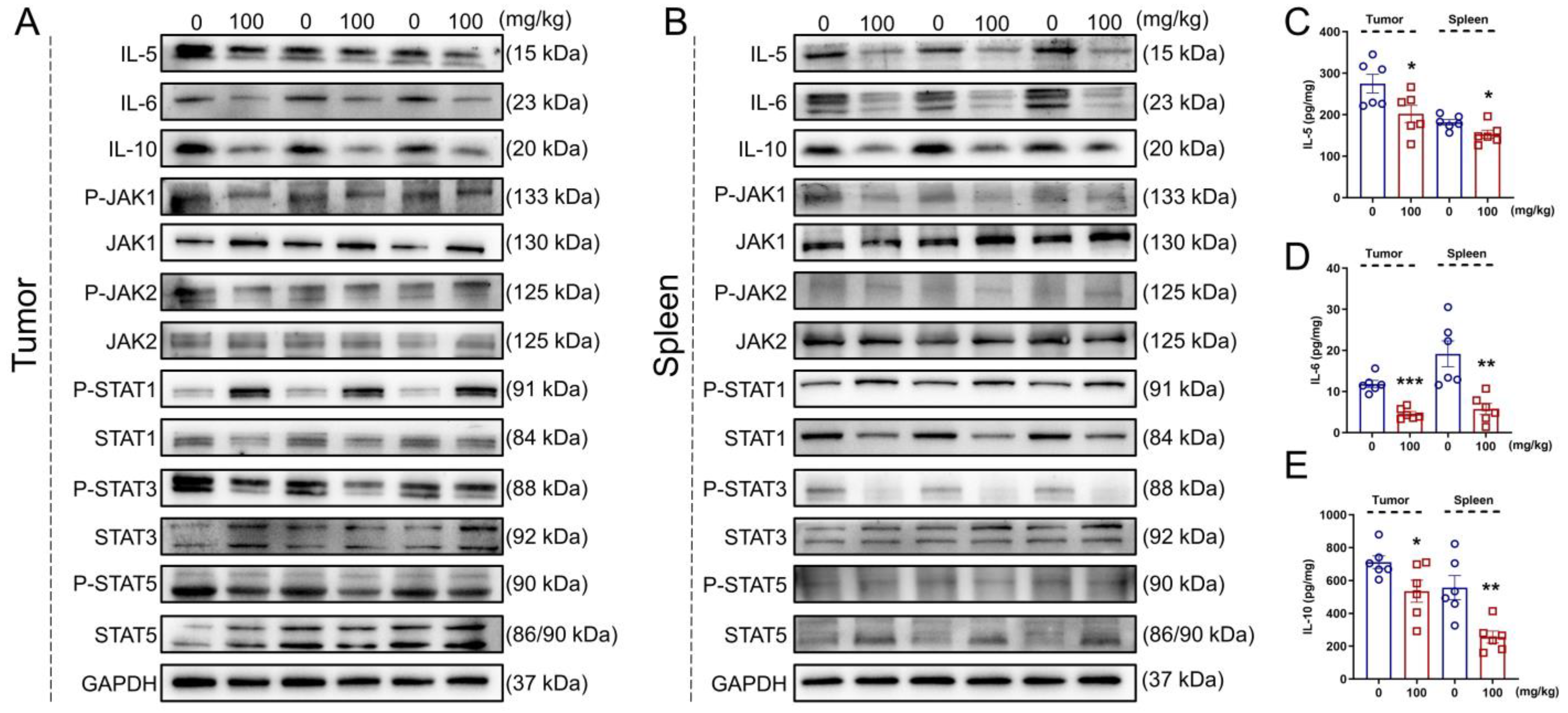
3.6. IHS Regulates JAK/STAT Signaling in ApcMin/+ Mice
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. Ca-A Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Li, Y.; Liang, X.; Yang, L.; Su, M.; Lai, K.P. Network Pharmacology and bioinformatics analyses identify intersection genes of niacin and COVID-19 as potential therapeutic targets. Brief. Bioinform. 2021, 22, 1279–1290. [Google Scholar] [CrossRef]
- Keum, N.; Giovannucci, E. Global burden of colorectal cancer: Emerging trends, risk factors and prevention strategies. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 713–732. [Google Scholar] [CrossRef]
- Song, M.; Chan, A.T.; Sun, J. Influence of the Gut Microbiome, Diet, and Environment on Risk of Colorectal Cancer. Gastroenterology 2020, 158, 322–340. [Google Scholar] [CrossRef] [PubMed]
- Liang, Q.; Chiu, J.; Chen, Y.; Huang, Y.; Higashimori, A.; Fang, J.; Brim, H.; Ashktorab, H.; Ng, S.C.; Ng, S.S.M.; et al. Fecal Bacteria Act as Novel Biomarkers for Noninvasive Diagnosis of Colorectal Cancer. Clin. Cancer Res. 2017, 23, 2061–2070. [Google Scholar] [CrossRef] [PubMed]
- Si, H.; Yang, Q.; Hu, H.; Ding, C.; Wang, H.; Lin, X. Colorectal cancer occurrence and treatment based on changes in intestinal flora. Semin. Cancer Biol. 2021, 70, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Han, S.; Yang, X.; Qi, Q.; Pan, Y.; Da, M.; Zhou, Q. Relationship between intestinal microorganisms and T lymphocytes in colorectal cancer. Future Oncol. 2019, 15, 1655–1666. [Google Scholar] [CrossRef]
- Daester, S.; Eppenberger-Castori, S.; Hirt, C.; Soysal, S.D.; Delko, T.; Nebiker, C.A.; Weixler, B.; Amicarella, F.; Iezzi, G.; Governa, V.; et al. Absence of myeloperoxidase and CD8 positive cells in colorectal cancer infiltrates identifies patients with severe prognosis. Oncoimmunology 2015, 4, e1050574. [Google Scholar] [CrossRef]
- Galon, J.; Costes, A.; Sanchez-Cabo, F.; Kirilovsky, A.; Mlecnik, B.; Lagorce-Pages, C.; Tosolini, M.; Camus, M.; Berger, A.; Wind, P.; et al. Type, density, and location of immune cells within human colorectal tumors predict clinical outcome. Science 2006, 313, 1960–1964. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Li, J.; Gao, Y.; Jia, M.; Li, N.; Yang, G. Study on Screening of Excellent Strains of Inonotus hispidus Parasitizing on Mulberry. North Seric. 2019, 40, 8–12. [Google Scholar] [CrossRef]
- Ren, Q.; Lu, X.Y.; Han, J.X.; Aisa, H.A.; Yuan, T. Triterpenoids and phenolics from the fruiting bodies of Inonotus hispidus and their activations of melanogenesis and tyrosinase. Chin. Chem. Lett. 2017, 28, 1052–1056. [Google Scholar] [CrossRef]
- Gruendemann, C.; Arnhold, M.; Meier, S.; Baecker, C.; Garcia-Kaeufer, M.; Grunewald, F.; Steinborn, C.; Klemd, A.M.; Wille, R.; Huber, R.; et al. Effects of Inonotus hispidus Extracts and Compounds on Human Immunocompetent Cells. Planta Med. 2016, 82, 1359–1367. [Google Scholar] [CrossRef]
- Wang, T.; Bao, H.-Y.; Bau, T.; Li, Y. Antitumor Effect of Solid State Fermentation Powder of Inonotus hispidus on H22 Bearing Mice. Zhong Yao Cai = Zhongyaocai = J. Chin. Med. Mater. 2016, 39, 389–394. [Google Scholar]
- Angelini, P.; Girometta, C.; Tirillini, B.; Moretti, S.; Covino, S.; Cipriani, M.; D’Ellena, E.; Angeles, G.; Federici, E.; Savino, E.; et al. A comparative study of the antimicrobial and antioxidant activities of Inonotus hispidus fruit and their mycelia extracts. Int. J. Food Prop. 2019, 22, 768–783. [Google Scholar] [CrossRef]
- Bal, C.; Akgul, H.; Sevindik, M.; Akata, I.; Yumrutas, O. Determination of the anti-oxidative activities of six mushrooms. Fresenius Environ. Bull. 2017, 26, 6246–6252. [Google Scholar]
- Fearon, E.R. Molecular Genetics of Colorectal Cancer. Annu. Rev. Pathol. Mech. Dis. 2011, 6, 479–507. [Google Scholar] [CrossRef] [PubMed]
- Phesse, T.J.; Buchert, M.; Stuart, E.; Flanagan, D.J.; Faux, M.; Afshar-Sterle, S.; Walker, F.; Zhang, H.-H.; Nowell, C.J.; Jorissen, R.; et al. Partial inhibition of gp130-Jak-Stat3 signaling prevents Wnt-beta-catenin-mediated intestinal tumor growth and regeneration. Sci. Signal. 2014, 7, ra92. [Google Scholar] [CrossRef] [PubMed]
- Musteanu, M.; Blaas, L.; Mair, M.; Schlederer, M.; Bilban, M.; Tauber, S.; Esterbauer, H.; Mueller, M.; Casanova, E.; Kenner, L.; et al. Stat3 Is a Negative Regulator of Intestinal Tumor Progression in Apc(Min) Mice. Gastroenterology 2010, 138, 1003–1011. [Google Scholar] [CrossRef] [PubMed]
- Xiong, H.; Zhang, Z.G.; Tian, X.Q.; Sun, D.F.; Liang, Q.C.; Zhang, Y.J.; Lu, R.; Chen, Y.X.; Fang, J.Y. Inhibition of JAK1, 2/STAT3 signaling induces apoptosis, cell cycle arrest, and reduces tumor cell invasion in colorectal cancer cells. Neoplasia 2008, 10, 287–297. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Li, J.; Fu, M.; Zhao, X.; Wang, W. The JAK/STAT signaling pathway: From bench to clinic. Signal Transduct. Target. Ther. 2021, 6, 402. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Lundqvist, A. Immunomodulatory Effects of IL-2 and IL-15; Implications for Cancer Immunotherapy. Cancers 2020, 12, 3586. [Google Scholar] [CrossRef] [PubMed]
- Heinrich, P.C.; Behrmann, I.; Haan, S.; Hermanns, H.M.; Muller-Newen, G.; Schaper, F. Principles of interleukin (IL)-6-type cytokine signalling and its regulation. Biochem. J. 2003, 374, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Li, L.Z.; Zhao, C.F.; Kong, F.E.; Li, Y.C.; Wang, C.X.; Chen, S.S.; Tan, H.Y.; Liu, Y.; Wang, D. Calf Thymus Polypeptide Restrains the Growth of Colorectal Tumor via Regulating the Intestinal Microbiota-Mediated Immune Function. Front. Pharmacol. 2022, 13, 1645. [Google Scholar] [CrossRef]
- McClellan, J.L.; Davis, J.M.; Steiner, J.L.; Enos, R.T.; Jung, S.H.; Carson, J.A.; Pena, M.M.; Carnevale, K.A.; Berger, F.G.; Murphy, E.A. Linking tumor-associated macrophages, inflammation, and intestinal tumorigenesis: Role of MCP-1. Am. J. Physiol.-Gastrointest. Liver Physiol. 2012, 303, G1087–G1095. [Google Scholar] [CrossRef]
- Kowalska, H.; Kowalska, J.; Ignaczak, A.; Masiarz, E.; Domian, E.; Galus, S.; Ciurzynska, A.; Salamon, A.; Zajac, A.; Marzec, A. Development of a High-Fibre Multigrain Bar Technology with the Addition of Curly Kale. Molecules 2021, 26, 3939. [Google Scholar] [CrossRef] [PubMed]
- Adusei, S.; Otchere, J.K.; Oteng, P.; Mensah, R.Q.; Tei-Mensah, E. Phytochemical analysis, antioxidant and metal chelating capacity of Tetrapleura tetraptera. Heliyon 2019, 5, e02762. [Google Scholar] [CrossRef] [PubMed]
- Braga Resende, L.M.; de Souza, V.R.; Dias Ferreira, G.M.; Nunes, C.A. Changes in quality and phytochemical contents of avocado oil under different temperatures. J. Food Sci. Technol.-Mysore 2019, 56, 401–408. [Google Scholar] [CrossRef] [PubMed]
- Al Rashid, M.H.; Majumder, S.; Mandal, V.; Mandal, S.C.; Thandavarayan, R.A. In search of suitable extraction technique for large scale commercial production of bioactive fraction for the treatment of diabetes: The case Diospyros melanoxylon Roxb. J. Tradit. Complement. Med. 2019, 9, 106–118. [Google Scholar] [CrossRef]
- Cui, Y.; Zhao, X.; Tang, Y.; Zhang, Y.; Sun, L.; Zhang, X. Comparative Study on the Chemical Components and Gastrointestinal Function on Rats of the Raw Product and Licorice-Simmered Product of Polygala tenuifolia. Evid.-Based Complement. Altern. Med. 2021, 2021, 8855536. [Google Scholar] [CrossRef]
- Carvalho Santos, W.P.; Ribeiro, N.M.; Muniz Batista Santos, D.C.; Andrade Korn, M.G.; Lopes, M.V. Bioaccessibility assessment of toxic and essential elements in produced pulses, Bahia, Brazil. Food Chem. 2018, 240, 112–122. [Google Scholar] [CrossRef]
- Xu, X.-R.; Xiao, Q.; Hong, Y.-C.; Liu, Y.-H.; Liu, Y.; Tu, J. Activation of dopaminergic VTA inputs to the mPFC ameliorates chronic stress-induced breast tumor progression. Cns Neurosci. Ther. 2021, 27, 206–219. [Google Scholar] [CrossRef]
- Wang, C.; Cai, X.; Wang, R.; Zhai, S.; Zhang, Y.; Hu, W.; Zhang, Y.; Wang, D. Neuroprotective effects of verbascoside against Alzheimer’s disease via the relief of endoplasmic reticulum stress in A beta-exposed U251 cells and APP/PS1 mice. J. Neuroinflamm. 2020, 17, 309. [Google Scholar] [CrossRef]
- Jiang, X.; Hao, J.; Liu, Z.J.; Ma, X.T.; Feng, Y.X.; Teng, L.R.; Li, Y.; Wang, D. Anti-obesity effects of Grifola frondosa through the modulation of lipid metabolism via ceramide in mice fed a high-fat diet. Food Funct. 2021, 12, 6725–6739. [Google Scholar] [CrossRef]
- Fu, Q.; Zhou, S.; Yu, M.; Lu, Y.; He, G.; Huang, X.; Huang, Y. Portulaca oleracea Polysaccharides Modulate Intestinal Microflora in Aged Rats in vitro. Front. Microbiol. 2022, 13, 841397. [Google Scholar] [CrossRef]
- Li, S.; Yang, H.; Li, L.; Wang, W.; Tan, H.-Y.; Qu, Y.; Wang, D. The involvement of gut microbiota in the anti-tumor effect of carnosic acid via IL-17 suppression in colorectal cancer. Chem.-Biol. Interact. 2022, 365, 110080. [Google Scholar] [CrossRef]
- Luo, D.; Deng, T.; Yuan, W.; Deng, H.; Jin, M. Plasma metabolomic study in Chinese patients with wet age-related macular degeneration. BMC Ophthalmol. 2017, 17, 165. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Dai, X.; Zhou, C.-C.; Li, K.-X.; Zhang, Y.-J.; Lou, X.-Y.; Zhu, Y.-M.; Sun, Y.-L.; Peng, B.-X.; Cui, W. Integrated analysis of the faecal metagenome and serum metabolome reveals the role of gut microbiome-associated metabolites in the detection of colorectal cancer and adenoma. Gut 2021, 71, 1315–1325. [Google Scholar] [CrossRef]
- Al-Koussa, H.; El Mais, N.; Maalouf, H.; Abi-Habib, R.; El-Sibai, M. Arginine deprivation: A potential therapeutic for cancer cell metastasis? A review. Cancer Cell Int. 2020, 20, 150. [Google Scholar] [CrossRef]
- Ren, B.; Van Kampen, E.; Van Berkel, T.J.C.; Cruickshank, S.M.; Van Eck, M. Hematopoietic arginase 1 deficiency results in decreased leukocytosis and increased foam cell formation but does not affect atherosclerosis. Atherosclerosis 2017, 256, 35–46. [Google Scholar] [CrossRef]
- Hibbs, J.B.; Westenfelder, C.; Taintor, R.; Vavrin, Z.; Kablitz, C.; Baranowski, R.L.; Ward, J.H.; Menlove, R.L.; McMurry, M.P.; Kushner, J.P.; et al. Evidence for Cytokine-Inducible Nitric-Oxide Synthesis From L-Arginine in Patients Receiving Interleukin-2 Therapy. J. Clin. Investig. 1992, 89, 867–877. [Google Scholar] [CrossRef] [PubMed]
- Chulpanova, D.S.; Gilazieva, Z.E.; Kletukhina, S.K.; Aimaletdinov, A.M.; Garanina, E.E.; James, V.; Rizvanov, A.A.; Solovyeva, V.V. Cytochalasin B-Induced Membrane Vesicles from Human Mesenchymal Stem Cells Overexpressing IL2 Are Able to Stimulate CD8(+) T-Killers to Kill Human Triple Negative Breast Cancer Cells. Biology 2021, 10, 141. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.W.; Qi, S.Z.; Xue, X.F.; Al Naggar, Y.; Wu, L.M.; Wang, K. Understanding the Gastrointestinal Protective Effects of Polyphenols using Foodomics-Based Approaches. Front. Immunol. 2021, 12, 141. [Google Scholar] [CrossRef]
- Wang, G.; Wang, Y.Z.; Yu, Y.; Wang, J.J.; Yin, P.H.; Xu, K. Triterpenoids Extracted from Rhus chinensis Mill Act Against Colorectal Cancer by Inhibiting Enzymes in Glycolysis and Glutaminolysis: Network Analysis and Experimental Validation. Nutr. Cancer-Int. J. 2020, 72, 293–319. [Google Scholar] [CrossRef]
- Yang, T.W.; Lee, W.H.; Tu, S.J.; Huang, W.C.; Chen, H.M.; Sun, T.H.; Tsai, M.C.; Wang, C.C.; Chen, H.Y.; Huang, C.C.; et al. Enterotype-based Analysis of Gut Microbiota along the Conventional Adenoma-Carcinoma Colorectal Cancer Pathway. Sci. Rep. 2019, 9, 10923. [Google Scholar] [CrossRef]
- Thomas, A.M.; Jesus, E.C.; Lopes, A.; Aguiar, S., Jr.; Begnami, M.D.; Rocha, R.M.; Carpinetti, P.A.; Camargo, A.A.; Hoffmann, C.; Freitas, H.C.; et al. Tissue-Associated Bacterial Alterations in Rectal Carcinoma Patients Revealed by 16S rRNA Community Profiling. Front. Cell. Infect. Microbiol. 2016, 6, 179. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.G.; Liu, F.L.; Ling, Z.X.; Tong, X.J.; Xiang, C. Human Intestinal Lumen and Mucosa-Associated Microbiota in Patients with Colorectal Cancer. PLoS ONE 2012, 7, e39743. [Google Scholar] [CrossRef]
- Chassard, C.; Delmas, E.; Robert, C.; Lawson, P.A.; Bernalier-Donadille, A. Ruminococcus champanellensis sp nov., a cellulose-degrading bacterium from human gut microbiota. Int. J. Syst. Evol. Microbiol. 2012, 62, 138–143. [Google Scholar] [CrossRef]
- Flint, H.J.; Duncan, S.H.; Scott, K.P.; Louis, P. Links between diet, gut microbiota composition and gut metabolism. Proc. Nutr. Soc. 2015, 74, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Sheridan, P.O.; Louis, P.; Tsompanidou, E.; Shaw, S.; Harmsen, H.J.; Duncan, S.H.; Flint, H.J.; Walker, A.W. Distribution, organization and expression of genes concerned with anaerobic lactate utilization in human intestinal bacteria. Microb. Genom. 2022, 8, 000739. [Google Scholar] [CrossRef] [PubMed]
- Ohashi, T.; Akazawa, T.; Aoki, M.; Kuze, B.; Mizuta, K.; Ito, Y.; Inoue, N. Dichloroacetate improves immune dysfunction caused by tumor-secreted lactic acid and increases antitumor immunoreactivity. Int. J. Cancer 2013, 133, 1107–1118. [Google Scholar] [CrossRef]
- Munder, M. Arginase: An emerging key player in the mammalian immune system. Br. J. Pharmacol. 2009, 158, 638–651. [Google Scholar] [CrossRef] [PubMed]
- Bronte, V.; Serafini, P.; Mazzoni, A.; Segal, D.M.; Zanovello, P. L-arginine metabolism in myeloid cells controls T-lymphocyte functions. Trends Immunol. 2003, 24, 302–306. [Google Scholar] [CrossRef]
- Qiu, Y.P.; Cai, G.X.; Su, M.M.; Chen, T.L.; Zheng, X.J.; Xu, Y.; Ni, Y.; Zhao, A.H.; Xu, L.X.; Cai, S.J.; et al. Serum Metabolite Profiling of Human Colorectal Cancer Using GC-TOFMS and UPLC-QTOFMS. J. Proteome Res. 2009, 8, 4844–4850. [Google Scholar] [CrossRef] [PubMed]
- Bronte, V.; Zanovello, P. Regulation of immune responses by L- arginine metabolism. Nat. Rev. Immunol. 2005, 5, 641–654. [Google Scholar] [CrossRef] [PubMed]
- Dai, Z.L.; Wu, G.Y.; Zhu, W.Y. Amino acid metabolism in intestinal bacteria: Links between gut ecology and host health. Front. Biosci.-Landmark 2011, 16, 1768–1786. [Google Scholar] [CrossRef] [PubMed]
- Komori, K.; Tsujimura, A.; Takao, T.; Matsuoka, Y.; Miyagawa, Y.; Takada, S.; Nonomura, N.; Okuyama, A. Nitric oxide synthesis leads to vascular endothelial growth factor synthesis via the NO/cyclic guanosine 3′,5′-monophosphate (cGMP) pathway in human corpus cavernosal smooth muscle cells. J. Sex. Med. 2008, 5, 1623–1635. [Google Scholar] [CrossRef] [PubMed]
- Samlowski, W.E.; Kondapaneni, M.; Tharkar, S.; McGregor, J.R.; Laubach, V.E.; Salvemini, D. Endothelial Nitric Oxide Synthase Is a Key Mediator of Interleukin-2-Induced Hypotension and Vascular Leak Syndrome. J. Immunother. 2011, 34, 419–427. [Google Scholar] [CrossRef]
- Perdigon, G.; Galdeano, C.M.; Valdez, J.C.; Medici, M. Interaction of lactic acid bacteria with the gut immune system. Eur. J. Clin. Nutr. 2002, 56, S21–S26. [Google Scholar] [CrossRef] [PubMed]
- Spolski, R.; Li, P.; Leonard, W.J. Biology and regulation of IL-2: From molecular mechanisms to human therapy. Nat. Rev. Immunol. 2018, 18, 648–659. [Google Scholar] [CrossRef] [PubMed]
- Lv, J.; Jia, Y.T.; Li, J.; Kuai, W.T.; Li, Y.; Guo, F.; Xu, X.J.; Zhao, Z.L.; Lv, J.; Li, Z.X. Gegen Qinlian decoction enhances the effect of PD-1 blockade in colorectal cancer with microsatellite stability by remodelling the gut microbiota and the tumour microenvironment. Cell Death Dis. 2019, 10, 415. [Google Scholar] [CrossRef] [PubMed]
- Cai, G.F.; Wusiman, A.; Gu, P.F.; Mao, N.N.; Xu, S.W.; Zhu, T.Y.; He, J.; Liu, Z.G.; Wang, D.Y. Supplementation of Alhagi honey polysaccharides contributes to the improvement of the intestinal immunity regulating the structure of intestinal flora in mice. Food Funct. 2021, 12, 9693–9707. [Google Scholar] [CrossRef] [PubMed]
- Braun, M.Y.; Desalle, F.; Le Moine, A.; Pretolani, M.; Matthys, P.; Kiss, R.; Goldman, M. IL-5 and eosinophils mediate the rejection of fully histoincompatible vascularized cardiac allografts: Regulatory role of alloreactive CD8+ T lymphocytes and IFN-gamma. Eur. J. Immunol. 2000, 30, 1290–1296. [Google Scholar] [CrossRef]
- Li, J.B.; Xu, J.; Yan, X.F.; Jin, K.; Li, W.Y.; Zhang, R. Targeting Interleukin-6 (IL-6) Sensitizes Anti-PD-L1 Treatment in a Colorectal Cancer Preclinical Model. Med. Sci. Monit. 2018, 24, 5501–5508. [Google Scholar] [CrossRef] [PubMed]
- Diao, B.; Wang, C.H.; Tan, Y.J.; Chen, X.W.; Liu, Y.; Ning, L.F.; Chen, L.; Li, M.; Liu, Y.P.; Wang, G.; et al. Reduction and Functional Exhaustion of T Cells in Patients With Coronavirus Disease 2019 (COVID-19). Front. Immunol. 2020, 11, 827. [Google Scholar] [CrossRef]
- Kaminskiy, Y.; Melenhorst, J.J. STAT3 Role in T-Cell Memory Formation. Int. J. Mol. Sci. 2022, 23, 2878. [Google Scholar] [CrossRef] [PubMed]
- Cui, W.G.; Liu, Y.; Weinstein, J.S.; Craft, J.; Kaech, S.M. An Interleukin-21-Interleukin-10-STAT3 Pathway Is Critical for Functional Maturation of Memory CD8(+) T Cells. Immunity 2011, 35, 792–805. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Pardoll, D.; Jove, R. STATs in cancer inflammation and immunity: A leading role for STAT3. Nat. Rev. Cancer 2009, 9, 798–809. [Google Scholar] [CrossRef] [PubMed]
- Suwanpradid, J.; Shih, M.; Pontius, L.; Yang, B.; Birukova, A.; Guttman-Yassky, E.; Corcoran, D.L.; Que, L.G.; Tighe, R.M.; MacLeod, A.S. Arginase1 Deficiency in Monocytes/Macrophages Upregulates Inducible Nitric Oxide Synthase To Promote Cutaneous Contact Hypersensitivity. J. Immunol. 2017, 199, 1827–1834. [Google Scholar] [CrossRef] [PubMed]
- Ohno, Y.; Kitamura, H.; Takahashi, N.; Ohtake, J.; Kaneumi, S.; Sumida, K.; Homma, S.; Kawamura, H.; Minagawa, N.; Shibasaki, S.; et al. IL-6 down-regulates HLA class II expression and IL-12 production of human dendritic cells to impair activation of antigen-specific CD4(+) T cells. Cancer Immunol. Immunother. 2016, 65, 193–204. [Google Scholar] [CrossRef] [PubMed]
- Parker, K.D.; Maurya, A.K.; Ibrahim, H.; Rao, S.; Hove, P.R.; Kumar, D.; Kant, R.; Raina, B.; Agarwal, R.; Kuhn, K.A.; et al. Dietary Rice Bran-Modified Human Gut Microbial Consortia Confers Protection against Colon Carcinogenesis Following Fecal Transfaunation. Biomedicines 2021, 9, 144. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Maurya, A.K.; Parker, K.D.; Kant, R.; Ibrahim, H.; Kabir, M.I.; Kumar, D.; Weber, A.M.; Agarwal, R.; Kuhn, K.A.; et al. Gender-based effect of absence of gut microbiota on the protective efficacy of Bifidobacterium longum-fermented rice bran diet against inflammation-associated colon tumorigenesis. Mol. Carcinog. 2022. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | Contents (%) |
|---|---|
| Protein | 15.3 |
| Fat | 0.8 |
| Total sugar | 39 |
| Total dietary fiber | 70.23 |
| Total sterol | 0.15 |
| Total alkaloid | 0.07 |
| Total triterpenoids | 0.39 |
| Total flavonoids | 2.07 |
| Total saponin | 1.34 |
| Total polyphenols | 2.65 |
| Compounds | Contents (mg/kg) | |
|---|---|---|
| Heavy metals | Lead (Pb) | 0.368 |
| Mercury (Hg) | 0.0189 | |
| Arsenic (As) | 0.117 | |
| Cadmium (Cd) | 0.064 | |
| Chromium (Cr) | 0.0939 | |
| Copper (Cu) | 9.46 | |
| Metals | Manganese (Mn) | 2.4 |
| Iron (Fe) | 27.9 | |
| Zinc (Zn) | 11.4 | |
| Selenium (Se) | 0.0823 | |
| Potassium (K) | 7.18 × 103 | |
| Calcium (Ca) | 124 | |
| Sodium (Na) | UD * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, H.; Li, S.; Qu, Y.; Li, L.; Li, Y.; Wang, D. Anti-Colorectal Cancer Effects of Inonotus hispidus (Bull.: Fr.) P. Karst. Spore Powder through Regulation of Gut Microbiota-Mediated JAK/STAT Signaling. Nutrients 2022, 14, 3299. https://doi.org/10.3390/nu14163299
Yang H, Li S, Qu Y, Li L, Li Y, Wang D. Anti-Colorectal Cancer Effects of Inonotus hispidus (Bull.: Fr.) P. Karst. Spore Powder through Regulation of Gut Microbiota-Mediated JAK/STAT Signaling. Nutrients. 2022; 14(16):3299. https://doi.org/10.3390/nu14163299
Chicago/Turabian StyleYang, Hongxin, Siyu Li, Yidi Qu, Lanzhou Li, Yu Li, and Di Wang. 2022. "Anti-Colorectal Cancer Effects of Inonotus hispidus (Bull.: Fr.) P. Karst. Spore Powder through Regulation of Gut Microbiota-Mediated JAK/STAT Signaling" Nutrients 14, no. 16: 3299. https://doi.org/10.3390/nu14163299
APA StyleYang, H., Li, S., Qu, Y., Li, L., Li, Y., & Wang, D. (2022). Anti-Colorectal Cancer Effects of Inonotus hispidus (Bull.: Fr.) P. Karst. Spore Powder through Regulation of Gut Microbiota-Mediated JAK/STAT Signaling. Nutrients, 14(16), 3299. https://doi.org/10.3390/nu14163299