
Elucidation of Anti-Hypertensive Mechanism by a Novel Lactobacillus rhamnosus AC1 Fermented Soymilk in the Deoxycorticosterone Acetate-Salt Hypertensive Rats


Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacteria Culture
2.2. Preparation of Fermented Soymilk with L. rhamnosus AC1 and Its Extract
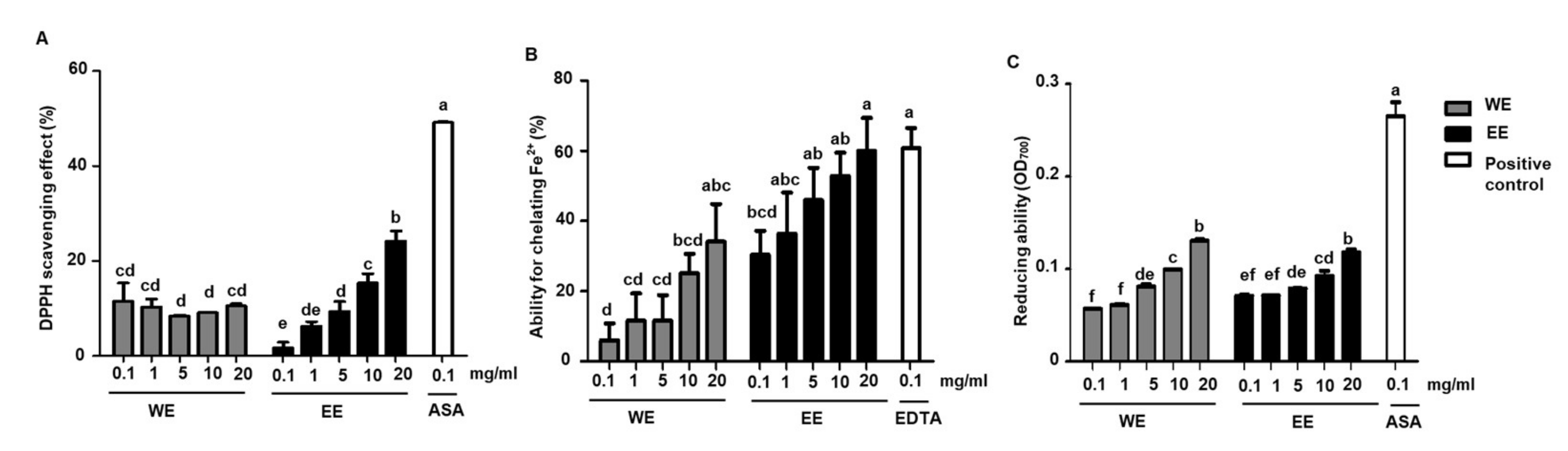
2.3. Measurement of Antioxidative Abilities In Vitro
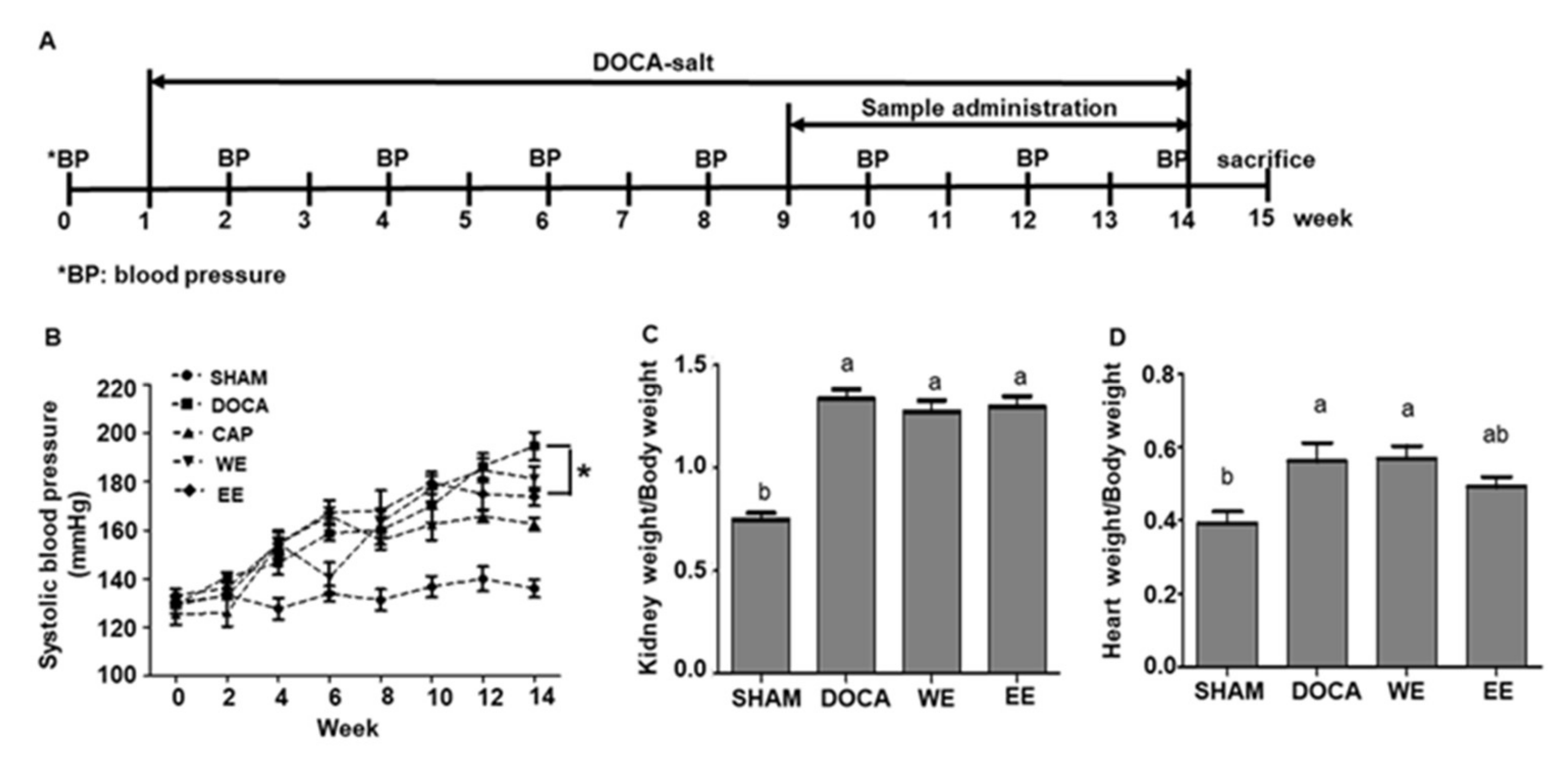
2.4. Animal Experiments
2.5. DOCA-Salt Hypertensive Rat Model
2.6. Biochemical and Histological Assessment
2.7. Analysis of Gut Microbiota
2.8. Metabolomic Profiling of Serum Samples
2.9. Data Analysis
3. Results
3.1. Antioxidative Effects of L. rhamnosus AC1-Fermented Soymilk In Vitro
3.2. Improvement Effects of L. rhamnosus AC1-Fermented Soymilk Extract on Hypertensive Rats
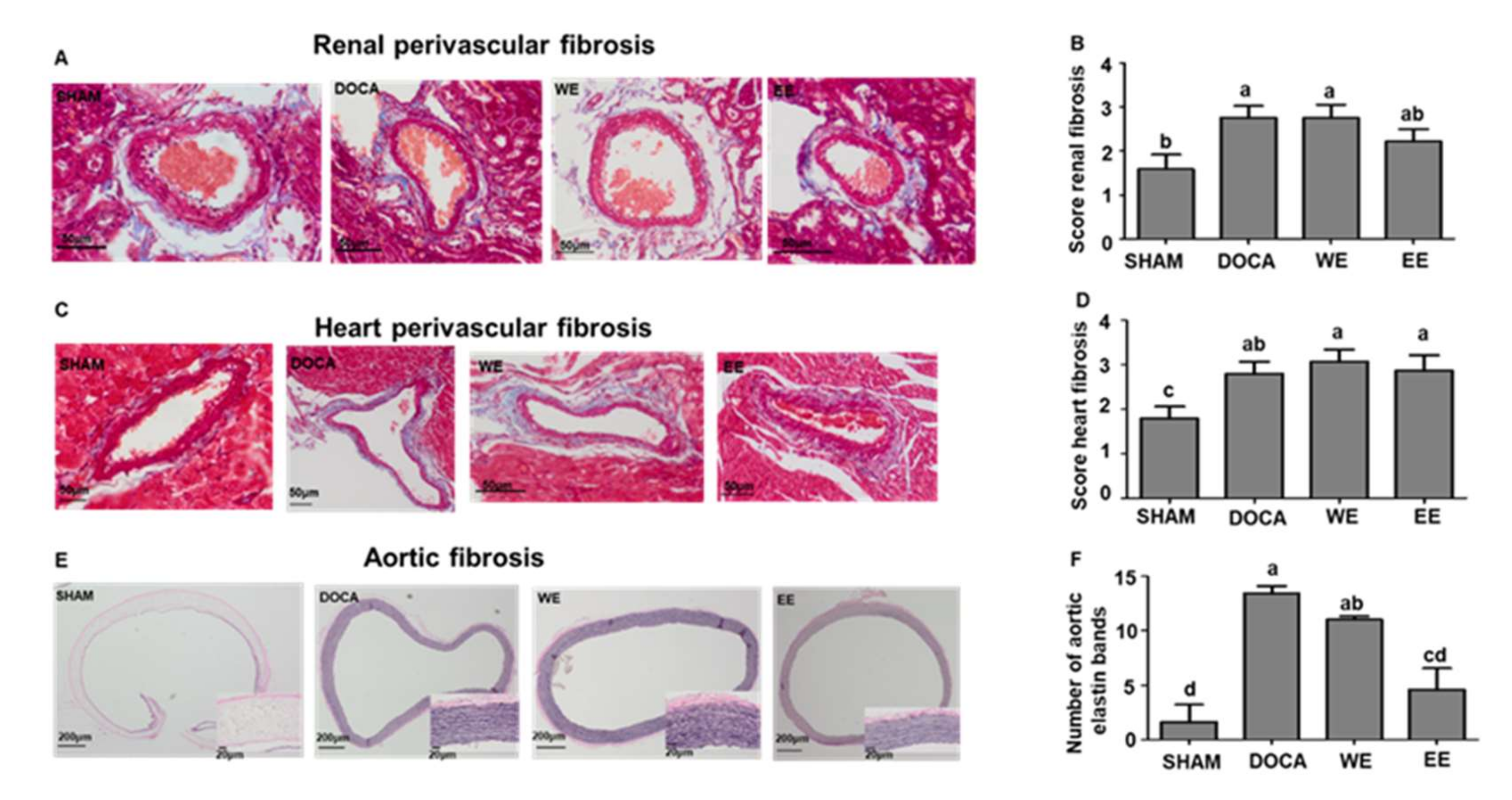
3.3. Improvement Effects of L. rhamnosus AC1-Fermented Soymilk Extract on Tissue Hypertrophy and Fibrosis
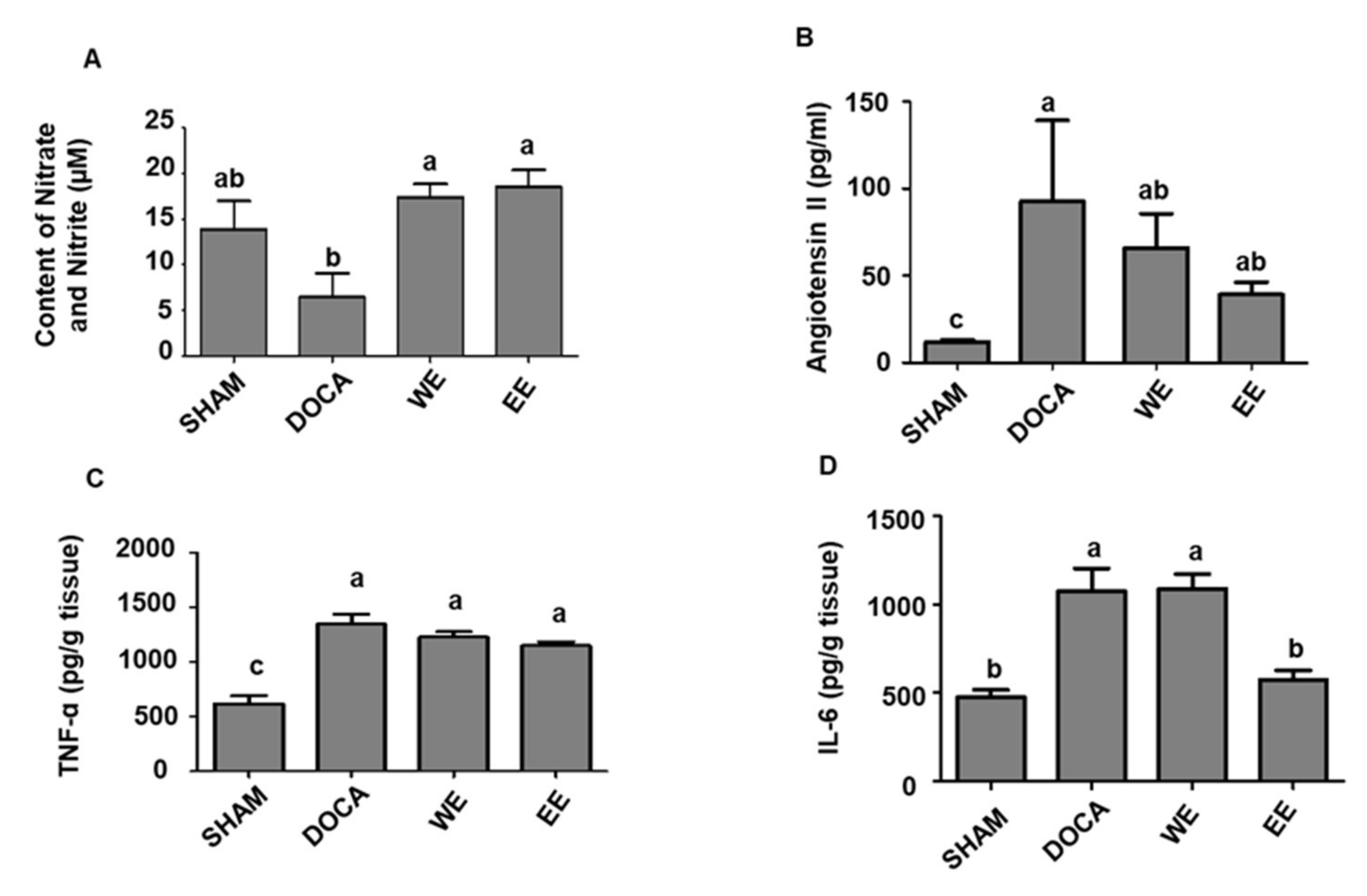
3.4. Regulatory Effects of L. rhamnosus AC1-Fermented Soymilk Extract on DOCA-Salt Hypertensive Rats
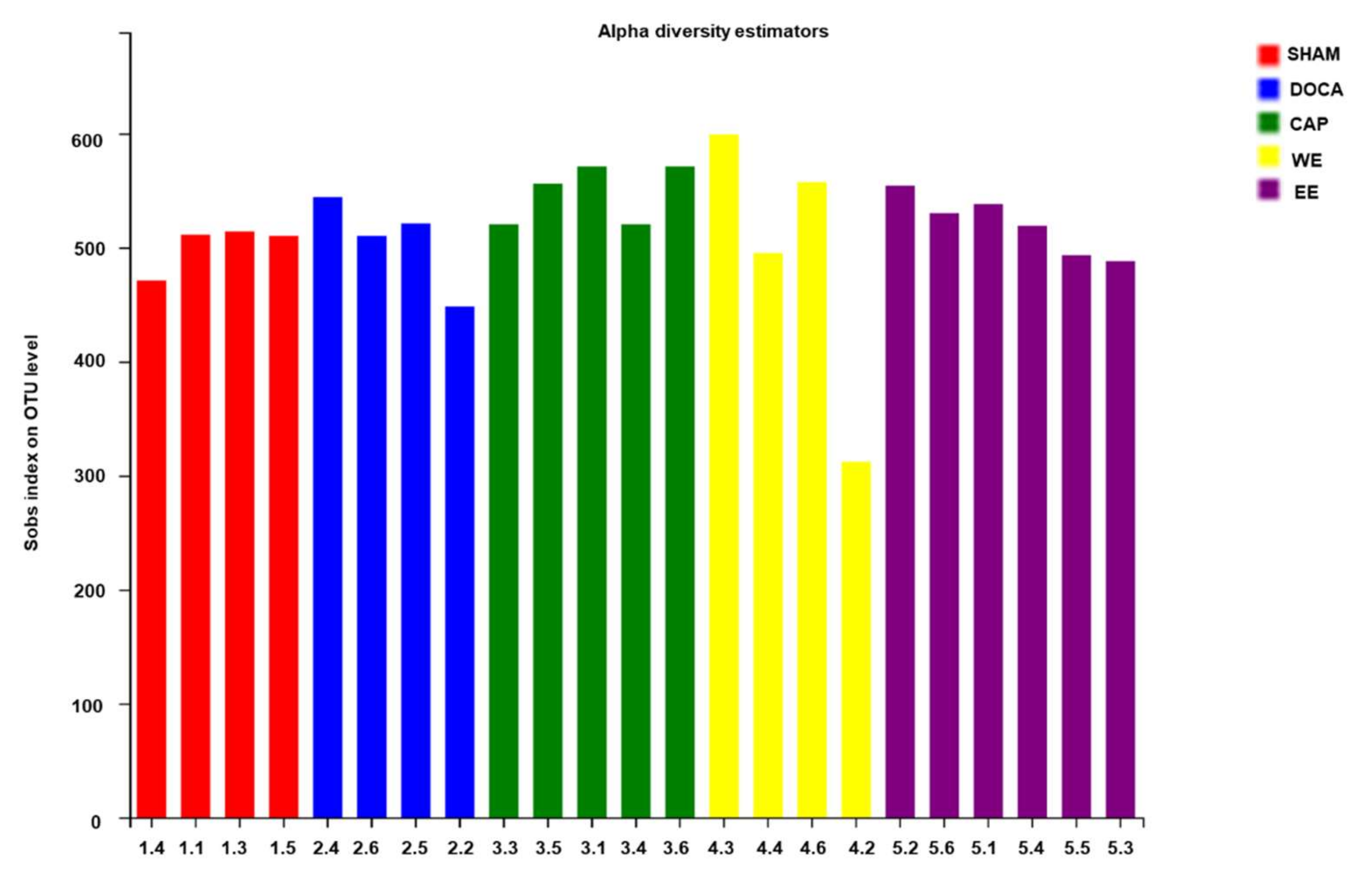
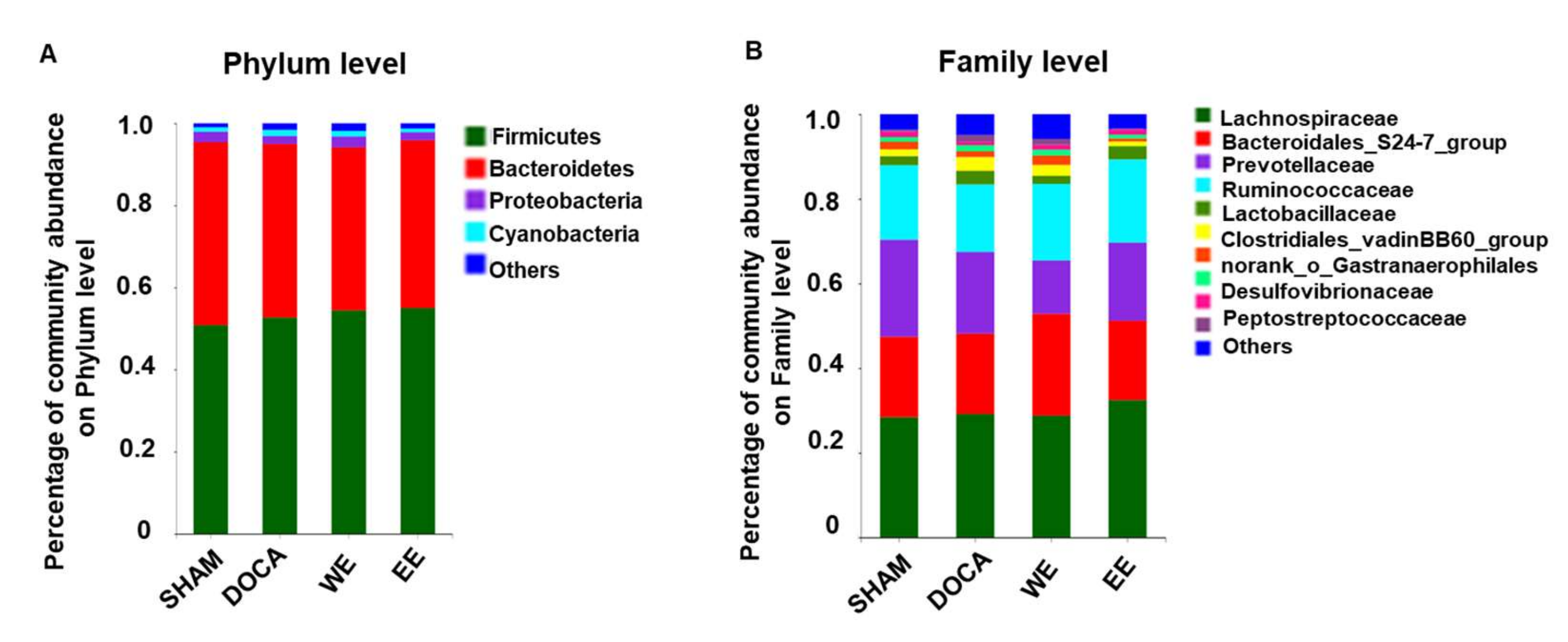
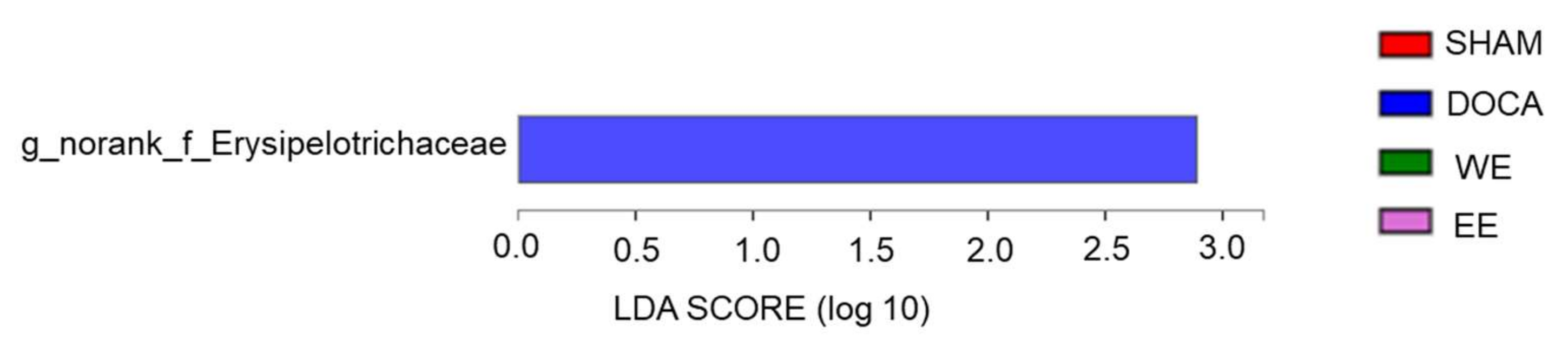
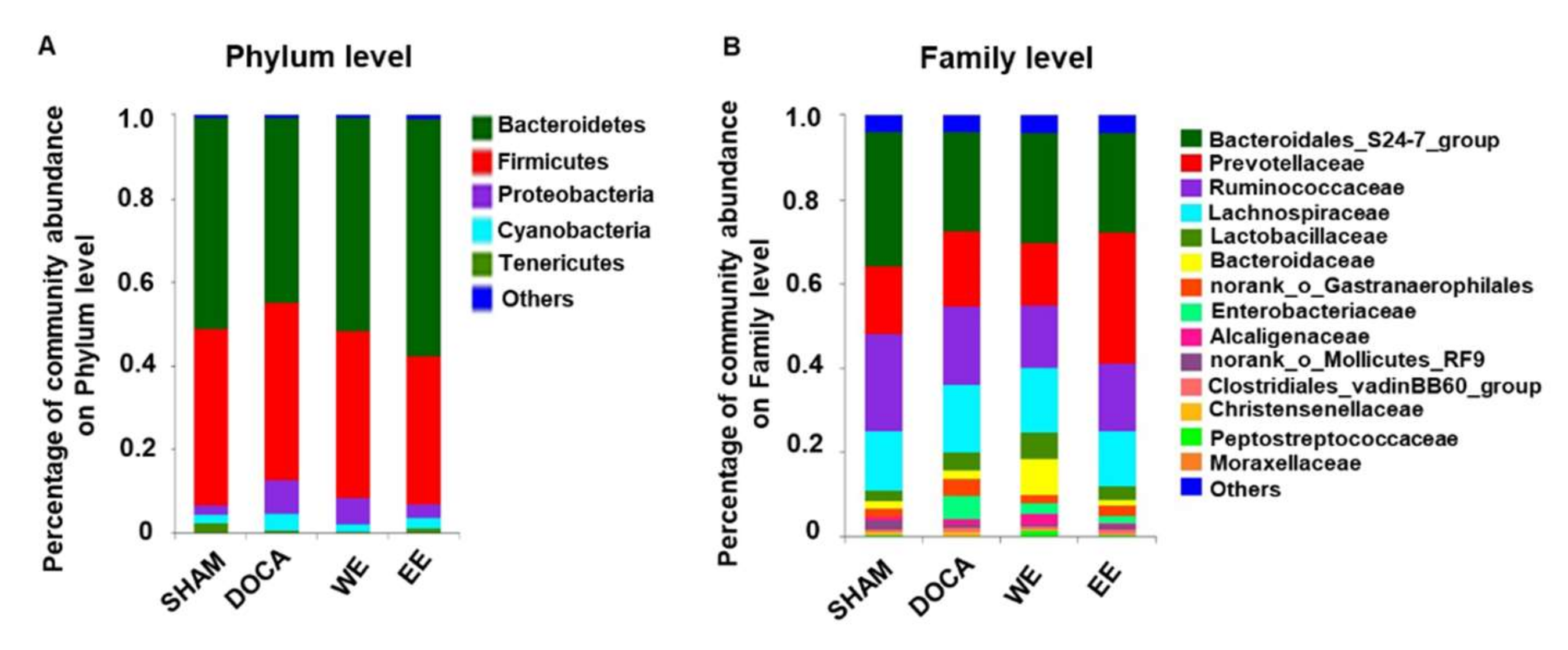
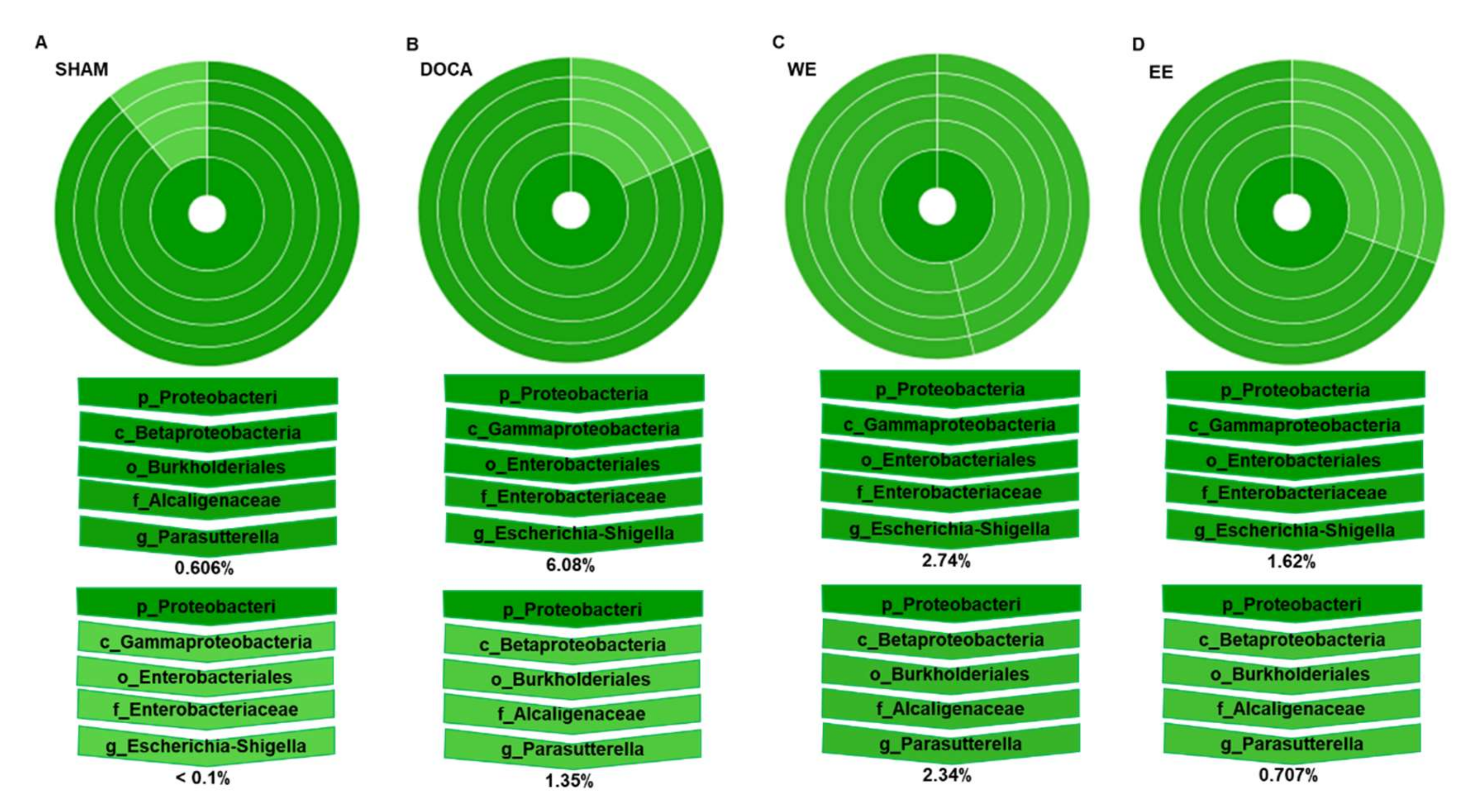
3.5. Effects of L. rhamnosus AC1-Fermented Soymilk Extract on Alteration of Gut Microbiome
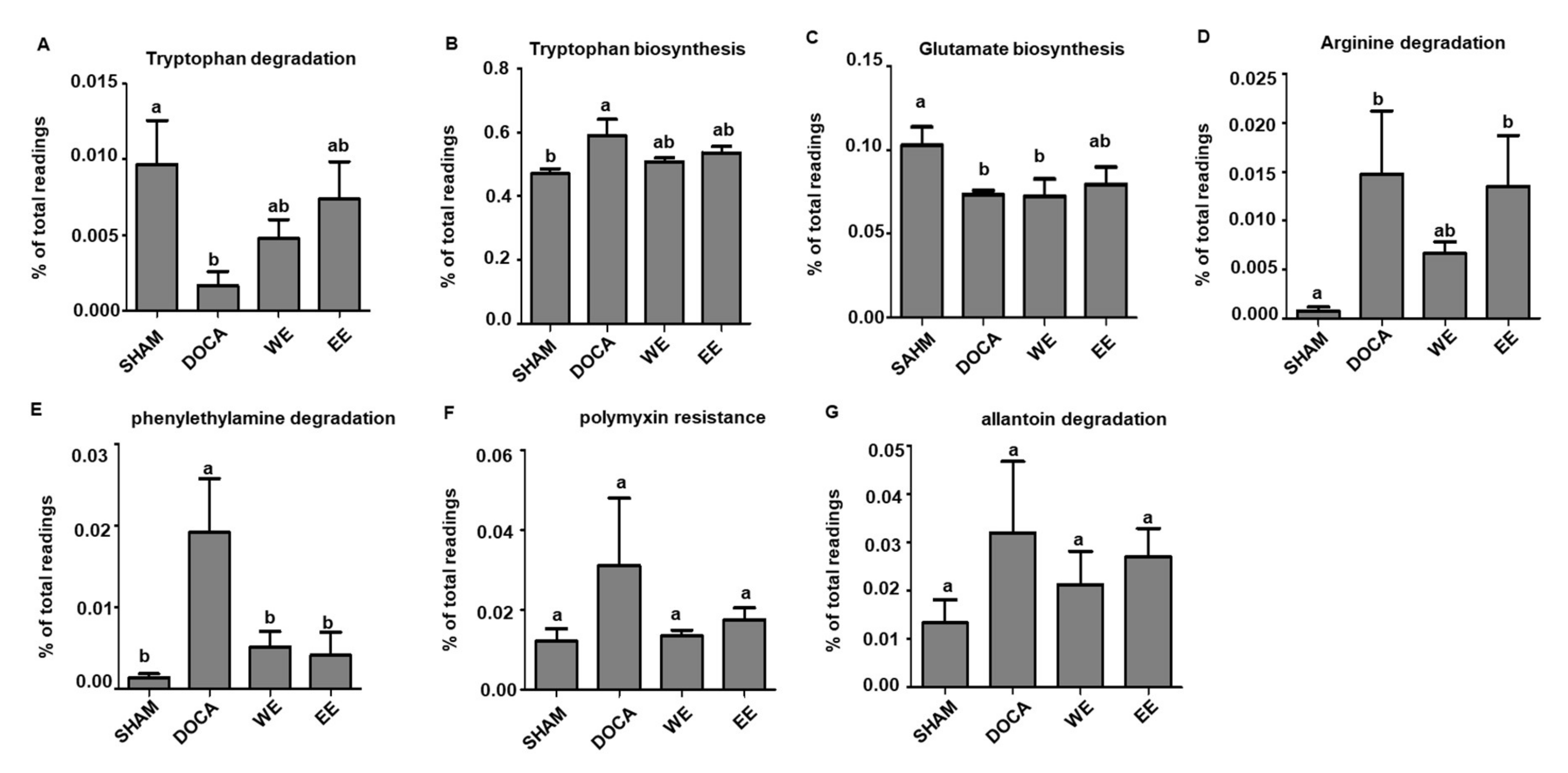
3.6. L. rhamnosus AC1-Fermented Soymilk Extract Alters the Levels of Potential Gut Microbiota-Associated Metabolites in DOCA-Salt-Induced Hypertension
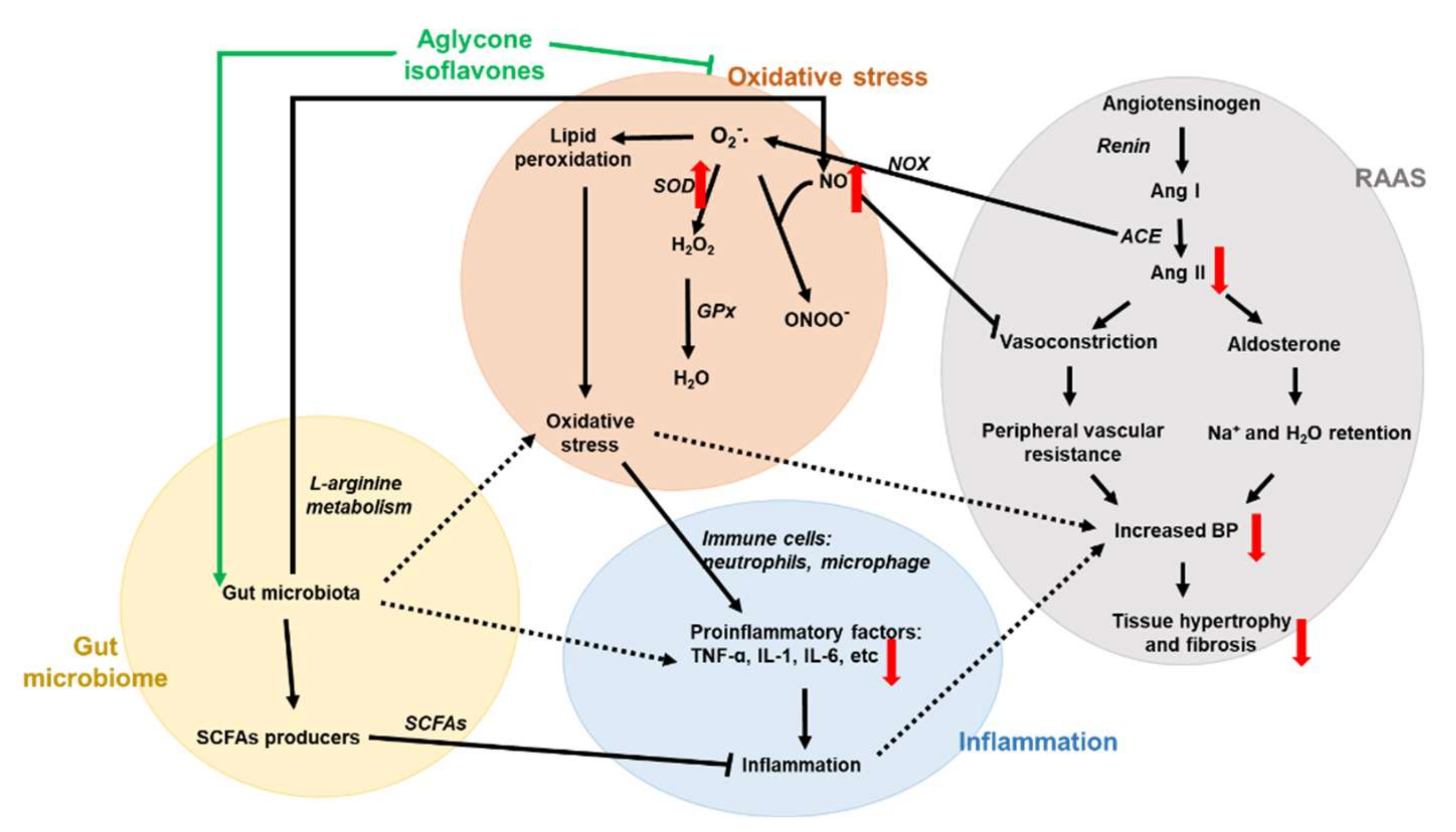
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

References
- Kearney, P.M.; Whelton, M.; Reynolds, K.; Whelton, P.K.; He, J. Worldwide prevalence of hypertension: A systematic review. J. Hypertens. 2004, 22, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Heilpern, K. Pathophysiology of hypertension. Ann. Emerg. Med. 2008, 51 (Suppl. S3), S5–S6. [Google Scholar] [CrossRef] [PubMed]
- Lee, L.K.; Kim, M.Y.; Kim, J.H.; Lee, J.U.; Park, B.S.; Yang, S.M.; Jeon, H.J.; Lee, W.D.; Noh, J.W.; Kwak, T.Y.; et al. A review of deoxycorticosterone acetate-salt hypertension and its relevance for cardiovascular physiotherapy research. J. Phys. Ther. Sci. 2015, 27, 303–307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iyer, A.; Chan, V.; Brown, L. The DOCA-Salt Hypertensive Rat as a Model of Cardiovascular Oxidative and Inflammatory Stress. Curr. Cardiol. Rev. 2010, 6, 291–297. [Google Scholar] [CrossRef] [Green Version]
- Appel, L.J.; Brands, M.W.; Daniels, S.R.; Karanja, N.; Elmer, P.J.; Sacks, F.M. American Heart Association. Dietary approaches to prevent and treat hypertension: A scientific statement from the American Heart Association. Hypertension 2006, 47, 296–308. [Google Scholar] [CrossRef] [Green Version]
- Linden, W.; Moseley, J.V. The efficacy of behavioral treatments for hypertension. Appl. Psychophysiol. Biofeedback 2006, 31, 51–63. [Google Scholar] [CrossRef]
- Linden, W.; Chambers, L. Clinical effectiveness of non-drug treatment for hypertension: A meta-analysis. Ann. Behav. Med. 1994, 16, 35–45. [Google Scholar]
- Gilliland, S.E. Health and nutritional benefits from lactic acid bacteria. FEMS. Microbiol. Rev. 1990, 7, 175–188. [Google Scholar] [CrossRef]
- Cheng, C.P.; Tsai, S.W.; Chiu, C.P.; Pan, T.M.; Tsai, T.Y. The effect of probiotic-fermented soy milk on enhancing the NO-mediated vascular relaxation factors. J. Sci. Food Agric. 2013, 93, 1219–1225. [Google Scholar] [CrossRef]
- O’Hara, A.M.; Shanahan, F. The gut flora as a forgotten organ. EMBO Rep. 2006, 7, 688–693. [Google Scholar] [CrossRef] [Green Version]
- Ursell, L.K.; Haiser, H.J.; Van Treuren, W.; Garg, N.; Reddivari, L.; Vanamala, J.; Dorrestein, P.C.; Turnbaugh, P.J.; Knight, R. The intestinal metabolome: An intersection between microbiota and host. Gastroenterology 2014, 146, 1470–1476. [Google Scholar] [CrossRef] [Green Version]
- Ley, R.E.; Turnbaugh, P.J.; Klein, S.; Gordon, J.I. Microbial ecology: Human gut microbes associated with obesity. Nature 2006, 444, 1022–1023. [Google Scholar] [CrossRef]
- Sanapareddy, N.; Legge, R.M.; Jovov, B.; McCoy, A.; Burcal, L.; Araujo-Perez, F.; Randall, T.A.; Galanko, J.; Benson, A.; Sandler, R.S.; et al. Increased rectal microbial richness is associated with the presence of colorectal adenomas in humans. ISME J. 2012, 6, 1858–1868. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Duan, Z.P.; Ha, D.K.; Bengmark, S.; Kurtovic, J.; Riordan, S.M. Synbiotic modulation of gut flora: Effect on minimal hepatic encephalopathy in patients with cirrhosis. Hepatology 2004, 39, 1441–1449. [Google Scholar] [CrossRef]
- Maeda, Y.; Takeda, K. Role of Gut Microbiota in Rheumatoid Arthritis. J. Clin. Med. 2017, 6, 60. [Google Scholar] [CrossRef]
- Qin, J.; Li, Y.; Cai, Z.; Li, S.; Zhu, J.; Zhang, F.; Liang, S.; Zhang, W.; Guan, Y.; Shen, D.; et al. A metagenome-wide association study of gut microbiota in type 2 diabetes. Nature 2012, 490, 55–60. [Google Scholar] [CrossRef]
- Jose, P.A.; Raj, D. Gut microbiota in hypertension. Curr. Opin. Nephrol. Hypertens. 2015, 4, 403–409. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.; Shum, T.F.; Chiou, J. Characterization of the Probiotic Potential of Lactic Acid Bacteria Isolated from Kimchi, Yogurt, and Baby Feces in Hong Kong and Their Performance in Soymilk Fermentation. Microorganisms 2021, 9, 2544. [Google Scholar] [CrossRef]
- Wu, H.; Lam, T.Y.C.; Shum, T.F.; Tsai, T.Y.; Chiou, J. Hypotensive effect of captopril on deoxycorticosterone acetate-salt-induced hypertensive rat is associated with gut microbiota alteration. Hypertens. Res. 2021, 45, 270–282. [Google Scholar] [CrossRef]
- García, N.; Zazueta, C.; Aguilera-Aguirre, L. Oxidative Stress and Inflammation in Cardiovascular Disease. Oxidative Med. Cell. Longev. 2017, 2017, 5853238. [Google Scholar] [CrossRef] [Green Version]
- Koliada, A.; Syzenko, G.; Moseiko, V.; Budovska, L.; Puchkov, K.; Perederiy, V.; Gavalko, Y.; Dorofeyev, A.; Romanenko, M.; Tkach, S.; et al. Association between body mass index and Firmicutes/Bacteroidetes ratio in an adult Ukrainian population. BMC Microbiol. 2017, 17, 120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rowin, J.; Xia, Y.; Jung, B.; Sun, J. Gut inflammation and dysbiosis in human motor neuron disease. Physiol. Rep. 2017, 5, e13443. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Santisteban, M.M.; Rodriguez, V.; Li, E.; Ahmari, N.; Carvajal, J.M.; Zadeh, M.; Gong, M.; Qi, Y.; Zubcevic, J.; et al. Gut dysbiosis is linked to hypertension. Hypertension 2015, 65, 1331–1340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Zhao, F.; Wang, Y.; Chen, J.; Tao, J.; Tian, G.; Wu, S.; Liu, W.; Cui, Q.; Geng, B.; et al. Gut microbiota dysbiosis contributes to the development of hypertension. Microbiome 2017, 5, 14. [Google Scholar] [CrossRef] [Green Version]
- Rossier, B.C.; Bochud, M.; Devuyst, O. The Hypertension Pandemic: An Evolutionary Perspective. Physiology 2017, 32, 112–125. [Google Scholar] [CrossRef]
- Basting, T.; Lazartigues, E. DOCA-Salt Hypertension: An Update. Curr. Hypertens. Rep. 2017, 19, 32. [Google Scholar] [CrossRef]
- Selye, H.; Hall, C.E.; Rowley, E.M. Malignant Hypertension Produced by Treatment with Desoxycorticosterone Acetate and Sodium Chloride. Can. Med. Assoc. J. 1943, 49, 88–92. [Google Scholar]
- Gavras, H.; Brunner, H.R.; Laragh, J.H.; Vaughan, E.D., Jr.; Koss, M.; Cote, L.J.; Gavras, I. Malignant hypertension resulting from deoxycorticosterone acetate and salt excess: Role of renin and sodium in vascular changes. Circ. Res. 1975, 36, 300–309. [Google Scholar] [CrossRef] [Green Version]
- De Champlain, J.; Krakoff, L.; Axelrod, J. Interrelationships of sodium intake, hypertension, and norepinephrine storage in the rat. Circ. Res. 1969, 24, 75–92. [Google Scholar]
- Campbell, S.E.; Janicki, J.S.; Matsubara, B.B.; Weber, K.T. Myocardial fibrosis in the rat with mineralocorticoid excess. Prevention of scarring by amiloride. Am. J. Hypertens. 1993, 6 Pt 1, 487–495. [Google Scholar] [CrossRef]
- De Champlain, J.; Krakoff, L.R.; Axelrod, J. Catecholamine metabolism in experimental hypertension in the rat. Circ. Res. 1967, 20, 136–145. [Google Scholar] [CrossRef] [Green Version]
- Molteni, A.; Nickerson, P.A.; Latta, J.; Brownie, A.C. Deveolpment of hypertension in rats maintained on a sodium deficient diet and bearing a mammotropic tumor (MtTF4). Virchows Arch. B Cell Pathol. 1975, 19, 191–203. [Google Scholar] [CrossRef]
- Brown, L.; Duce, B.; Miric, G.; Sernia, C. Reversal of cardiac fibrosis in deoxycorticosterone acetate-salt hypertensive rats by inhibition of the renin-angiotensin system. J. Am. Soc. Nephrol. 1999, 10 (Suppl. S11), S143–S148. [Google Scholar]
- Adnan, S.; Nelson, J.W.; Ajami, N.J.; Venna, V.R.; Petrosino, J.F.; Bryan, R.M., Jr.; Durgan, D.J. Alterations in the gut microbiota can elicit hypertension in rats. Physiol. Genom. 2017, 49, 96–104. [Google Scholar] [CrossRef] [Green Version]
- Richards, E.M.; Pepine, C.J.; Raizada, M.K.; Kim, S. The Gut, Its Microbiome, and Hypertension. Curr. Hypertens. Rep. 2017, 19, 36. [Google Scholar] [CrossRef] [Green Version]
- Shah, N.P. Probiotic bacteria: Selective enumeration and survival in dairy foods. J. Dairy Sci. 2000, 83, 894–907. [Google Scholar] [CrossRef]
- Wildman, R. Handbook of Nutraceuticals and Functional Foods, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2016. [Google Scholar]
- Tsangalis, D.; Ashton, J.F.; Mcgill, A.E.J.; Shah, N.P. Enzymic Transformation of Isoflavone Phytoestrogens in Soymilk by β-Glucosidase-Producing Bifidobacteria. J. Food Sci. 2002, 67, 3104–3113. [Google Scholar] [CrossRef]
- Wang, Y.C.; Yu, R.C.; Chou, C.C. Antioxidative activities of soymilk fermented with lactic acid bacteria and bifidobacteria. Food Microbiol. 2006, 23, 128–135. [Google Scholar] [CrossRef]
- Hsieh, M.L.; Chou, C.C. Mutagenicity and antimutagenic effect of soymilk fermented with lactic acid bacteria and bifidobacteria. Int. J. Food Microbiol. 2006, 111, 43–47. [Google Scholar] [CrossRef]
- Karbach, S.H.; Schönfelder, T.; Brandão, I.; Wilms, E.; Hörmann, N.; Jäckel, S.; Schüler, R.; Finger, S.; Knorr, M.; Lagrange, J.; et al. Gut Microbiota Promote Angiotensin II-Induced Arterial Hypertension and Vascular Dysfunction. J. Am. Heart Assoc. 2016, 5, e003698. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Systolic BP (mmHg) | |||||||
|---|---|---|---|---|---|---|---|---|
| Week 0 | Week 2 | Week 4 | Week 6 | Week 8 | Week 10 | Week 12 | Week 14 | |
| SHAM | 129.24 ± 2.55 Aa | 133.42 ± 3.14 ABa | 128.06 ± 2.59 Ba | 138.30 ± 4.11 Ba | 131.38 ± 5.62 Ba | 139.91 ± 5.22 Ba | 140.09 ± 4.32 Ca | 136.1 ± 5.06 Ca |
| DOCA | 129.40 ± 3.26 Ae | 126.03 ± 4.19 Ade | 147.49 ± 5.26 Acd | 161.09 ± 6.89 Abc | 163.66 ± 6.99 Abc | 170.13 ± 8.69 Ab | 186.42 ± 8.02 Aa | 192.14 ± 9.54 Aa |
| CAP | 125.39 ± 3.59 Ab | 140.73 ± 5.29 Bb | 156.43 ± 5.69 Aa | 166.15 ± 7.89 Aa | 155.78 ± 7.96 Aa | 162.14 ± 10.20 Aa | 164.25 ± 8.65 Ba | 162.57 ± 7.23 Ba |
| WE | 130.43 ± 6.52 Ac | 133.95 ± 5.23 ABc | 155.77 ± 7.85 Ac | 147.50 ± 9.46 Bb | 166.97 ± 5.87 Ab | 178.63 ± 5.99 Aa | 190.94 ± 9.84 Aa | 181.26 ± 8.32 Aa |
| EE | 133.34 ± 5.41 Ad | 136.14 ± 4.69 Ad | 159.40 ± 7.23 Ac | 163.34 ± 8.26 Abc | 164.70 ± 7.46 Abc | 178.19 ± 9.56 Aa | 175.03 ± 10.21 ABa | 173.84 ± 9.65 Ba |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, H.; Jiang, L.; Shum, T.-F.; Chiou, J. Elucidation of Anti-Hypertensive Mechanism by a Novel Lactobacillus rhamnosus AC1 Fermented Soymilk in the Deoxycorticosterone Acetate-Salt Hypertensive Rats. Nutrients 2022, 14, 3174. https://doi.org/10.3390/nu14153174
Wu H, Jiang L, Shum T-F, Chiou J. Elucidation of Anti-Hypertensive Mechanism by a Novel Lactobacillus rhamnosus AC1 Fermented Soymilk in the Deoxycorticosterone Acetate-Salt Hypertensive Rats. Nutrients. 2022; 14(15):3174. https://doi.org/10.3390/nu14153174
Chicago/Turabian StyleWu, Haicui, Lilong Jiang, Tim-Fat Shum, and Jiachi Chiou. 2022. "Elucidation of Anti-Hypertensive Mechanism by a Novel Lactobacillus rhamnosus AC1 Fermented Soymilk in the Deoxycorticosterone Acetate-Salt Hypertensive Rats" Nutrients 14, no. 15: 3174. https://doi.org/10.3390/nu14153174
APA StyleWu, H., Jiang, L., Shum, T.-F., & Chiou, J. (2022). Elucidation of Anti-Hypertensive Mechanism by a Novel Lactobacillus rhamnosus AC1 Fermented Soymilk in the Deoxycorticosterone Acetate-Salt Hypertensive Rats. Nutrients, 14(15), 3174. https://doi.org/10.3390/nu14153174