
Advances in the Study of the Mechanism by Which Selenium and Selenoproteins Boost Immunity to Prevent Food Allergies


 and
and 
Abstract
1. Introduction
2. Selenium and Selenoproteins
3. Effects of Selenoproteins on the Immune System

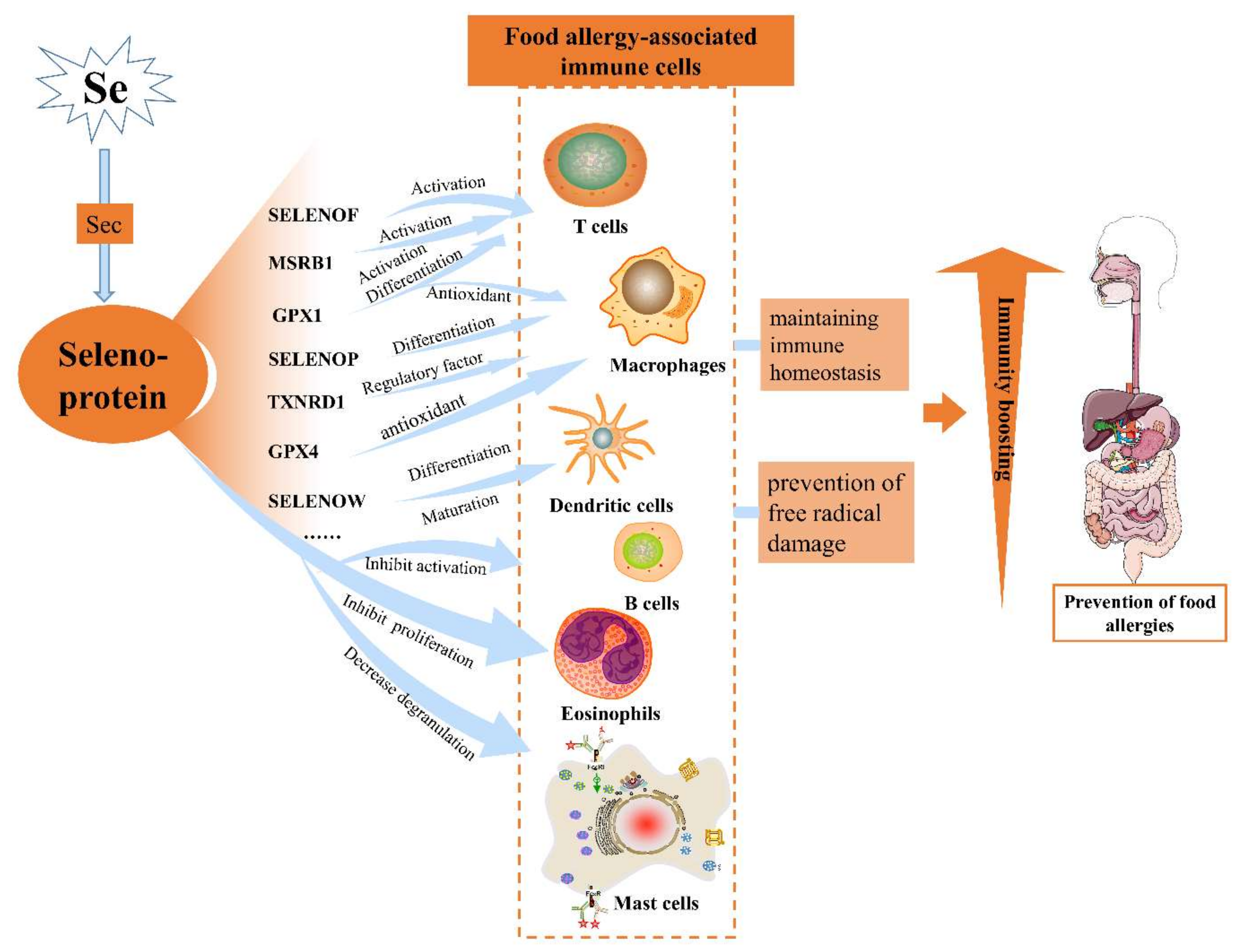{kind=link}
| Selenoprotein | Abbreviation(s) | Immune Function | Species | References |
|---|---|---|---|---|
| Selenoprotein K | SELENOK | Promote effective Ca2+ flux during immune cell activation | Mice | [13,22,23,24] |
| Selenoprotein S | SELENOS, SEPS1 | Retrograde translocation of misfolded proteins from the ER to regulating ER stress; modify immune-cell activity | Mice | [25] |
| Thioredoxin reductase 1 | TXNRD1, TrxR1, TR1 | Act as a regulator and a regulated target in the macrophage gene expression. Maintain redox tone in immune cells through regeneration of reduced cytosolic TXN1 | Mice | [26] |
| Cytosolic glutathione Peroxidase | GPX1 | Highly expressed in macrophages and T cells as an antioxidant. Important in T-cell activation and differentiation | Mice | [27,28] |
| Phospholipid hydroperoxide glutathione peroxidase | GPX4 | Highly expressed in macrophages as an antioxidant | Mice | [4,29] |
| Deiodinases | DIOs | Active thyroid hormone levels may affect systemic Se available for selenoprotein synthesis in tissues involved in the immune response | Mice | [4,30] |
| Selenoprotein P | SELENOP, SEPP | Intracellular antioxidant in phagocytes; affect the differentiation of macrophages; modulate immune cell function | Mice | [31,32] |
| Selenoprotein H | SELENOH | Transcriptional regulation | Mice | [33] |
| Selenoprotein I | SELENOI | Phospholipid synthesis Act as an ethanolamine phosphotransferase affecting T-cell activation | Mice | [4,34] |
| Selenoprotein F | SELENOF, SEP15 | Protein-folding; facilitates antigen presentation via ER-to-Golgi transport | Mice | [34,35] |
| Selenoprotein R | MSRB1 | Promote DC-mediated T-cell priming and Th1 differentiation | Mice | [36,37] |
| Selenophosphate Synthetase | SPS2 | The biosynthesis of selenoproteins | Mice | [38] |
| Selenoprotein T | SELENOT | Strongly correlated with immune-related cytokines, especially IL-1β and IL-2 | Chicken | [39,40] |
| Selenoprotein W | SELENOW | Expressed in the immune system, but the regulatory mechanism is unknown | Chicken | [31,41,42] |
4. Glutathione Peroxidases
5. Thioredoxin Reductases
6. Deiodinases
7. The Thioredoxin-Like Rdx Family
8. Other Selenoproteins
9. The Role of Selenoproteins in the Prevention of Food Allergies
9.1. Food Allergy and the Immune System
9.2. Ig-E-Mediated Food Allergy
9.3. Non-IgE-Mediated Food Allergy
9.4. Se and Selenoproteins for Immunity
9.5. Reasonable Se Supplementation to Prevent Food Allergy
10. Summary and Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
Abbreviations
References
- Rayman, M.P. Selenium and human health. Lancet 2012, 379, 1256–1268. [Google Scholar] [CrossRef]
- Reeves, M.A.; Hoffmann, P.R. The human selenoproteome: Recent insights into functions and regulation. Cell. Mol. Life Sci. 2009, 66, 2457–2478. [Google Scholar] [CrossRef]
- Rotruck, J.; Pope, A.; Ganther, H.; Swanson, A.; Hafeman, D.; Hoekstra, W. Selenium—Biochemical Role as a Component of Glutathione Peroxidase. Science 1973, 179, 588–590. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Rose, A.H.; Hoffmann, P.R. The Role of Selenium in Inflammation and Immunity: From Molecular Mechanisms to Therapeutic Opportunities. Antioxid. Redox Signal. 2012, 16, 705–743. [Google Scholar] [CrossRef]
- Zhao, X.; Hogenkamp, A.; Li, X.; Chen, H.; Garssen, J.; Knippels, L.M.J. Role of selenium in IgE mediated soybean allergy development. Crit. Rev. Food Sci. Nutr. 2022, 20, 1–9. [Google Scholar] [CrossRef]
- Combs, G.F. Selenium in global food systems. Br. J. Nutr. 2001, 85, 517–547. [Google Scholar] [CrossRef] [PubMed]
- Varlamova, E.G.; Turovsky, E.A. The main cytotoxic effects of methylseleninic acid on various cancer cells. Int. J. Mol. Sci. 2021, 22, 6614. [Google Scholar] [CrossRef]
- Mal’tseva, V.N.; Goltyaev, M.V.; Turovsky, E.A.; Varlamova, E.G. Immunomodulatory and Anti-Inflammatory Properties of Selenium-Containing Agents: Their Role in the Regulation of Defense Mechanisms against COVID-19. Int. J. Mol. Sci. 2022, 23, 2360. [Google Scholar] [CrossRef]
- Ren, Y.; Zhao, T.; Mao, G.; Zhang, M.; Li, F.; Zou, Y.; Yang, L.; Wu, X. Antitumor activity of hyaluronic acid–selenium nanoparticles in Heps tumor mice models. Int. J. Biol. Macromol. 2013, 57, 57–62. [Google Scholar] [CrossRef]
- Yanhua, W.; Hao, H.; Li, Y.; Zhang, S. Selenium-substituted hydroxyapatite nanoparticles and their in vivo antitumor effect on hepatocellular carcinoma. Colloids Surf. B Biointerfaces 2016, 140, 297–306. [Google Scholar] [CrossRef]
- Valdiglesias, V.; Pásaro, E.; Méndez, J.; Laffon, B. In vitro evaluation of selenium genotoxic, cytotoxic, and protective effects: A review. Arch. Toxicol. 2010, 84, 337–351. [Google Scholar] [CrossRef]
- Kalishwaralal, K.; Jeyabharathi, S.; Sundar, K.; Muthukumaran, A. A novel one-pot green synthesis of selenium nanoparticles and evaluation of its toxicity in zebrafish embryos. Artif. Cells Nanomed. Biotechnol. 2016, 44, 471–477. [Google Scholar] [CrossRef]
- Menon, S.; Ks, S.D.; Santhiya, R.; Rajeshkumar, S.; Kumar, V. Selenium nanoparticles: A potent chemotherapeutic agent and an elucidation of its mechanism. Colloids Surf. B Biointerfaces 2018, 170, 280–292. [Google Scholar] [CrossRef] [PubMed]
- Sampson, H.A.; O’Mahony, L.; Burks, A.W.; Plaut, M.; Lack, G.; Akdis, C.A. Mechanisms of food allergy. J. Allergy Clin. Immunol. 2018, 141, 11–19. [Google Scholar] [CrossRef]
- Satitsuksanoa, P.; Jansen, K.; Globinska, A.; Veen, W.; Akdis, M. Regulatory Immune Mechanisms in Tolerance to Food Allergy. Front. Immunol. 2018, 9, 2939. [Google Scholar] [CrossRef]
- Roman, M.; Jitaru, P.; Barbante, C. Selenium biochemistry and its role for human health. Metallomics 2014, 6, 25–54. [Google Scholar] [CrossRef] [PubMed]
- Minich, W.B. Selenium Metabolism and Biosynthesis of Selenoproteins in the Human Body. Biochemistry 2022, 87, S168–S177. [Google Scholar] [CrossRef]
- Serrao, V.H.B.; Silva, I.R.; da Silva, M.T.A.; Scortecci, J.F.; Fernandes, A.; Thiemann, O.H. The unique tRNA(Sec) and its role in selenocysteine biosynthesis. Amino Acids 2018, 50, 1145–1167. [Google Scholar] [CrossRef] [PubMed]
- Gladyshev, V.N.; Hatfield, D.L. Selenocysteine Biosynthesis, Selenoproteins, and Selenoproteomes. In Recoding: Expansion of Decoding Rules Enriches Gene Expression; Atkins, J.F., Gesteland, R.F., Eds.; Springer: Berlin, Germany, 2010; Volume 24, pp. 3–27. [Google Scholar]
- Dalgaard, T.S.; Briens, M.; Engberg, R.M.; Lauridsen, C. The influence of selenium and selenoproteins on immune responses of poultry and pigs. Anim. Feed Sci. Technol. 2018, 238, 73–83. [Google Scholar] [CrossRef]
- Avery, J.C.; Hoffmann, P.R. Selenium, Selenoproteins, and Immunity. Nutrients 2018, 10, 1203. [Google Scholar] [CrossRef] [PubMed]
- Verma, S.; Hoffmann, F.W.; Kumar, M.; Huang, Z.; Roe, K.; Nguyen-Wu, E.; Hashimoto, A.S.; Hoffmann, P.R. Selenoprotein K Knockout Mice Exhibit Deficient Calcium Flux in Immune Cells and Impaired Immune Responses. J. Immunol. 2011, 186, 2127–2137. [Google Scholar] [CrossRef] [PubMed]
- Marciel, M.P.; Hoffmann, P.R. Molecular Mechanisms by Which Selenoprotein K Regulates Immunity and Cancer. Biol. Trace Elem. Res. 2019, 192, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Du, S.; Zhou, J.; Jia, Y.; Huang, K. SelK is a novel ER stress-regulated protein and protects HepG2 cells from ER stress agent-induced apoptosis. Arch. Biochem. Biophys. 2010, 502, 137–143. [Google Scholar] [CrossRef]
- Gao, Y.; Feng, H.C.; Walder, K.; Bolton, K.; Sunderland, T.; Bishara, N.; Quick, M.; Kantham, L.; Collier, G.R. Regulation of the selenoprotein SelS by glucose deprivation and endoplasmic reticulum stress—SelS is a novel glucose-regulated protein. FEBS Lett. 2004, 563, 185–190. [Google Scholar] [CrossRef]
- Carlson, B.A.; Yoo, M.H.; Conrad, M.; Gladyshev, V.N.; Hatfield, D.L.; Park, J.M. Protein kinase-regulated expression and immune function of thioredoxin reductase 1 in mouse macrophages. Mol. Immunol. 2011, 49, 311–316. [Google Scholar] [CrossRef] [PubMed]
- Carlson, B.A.; Yoo, M.-H.; Shrimali, R.K.; Irons, R.; Gladyshev, V.N.; Hatfield, D.L.; Park, J.M. Session 2: Micronutrients and the immune system Role of selenium-containing proteins in T-cell and macrophage function. Proc. Nutr. Soc. 2010, 69, 300–310. [Google Scholar] [CrossRef]
- Hoffmann, F.W.; Hashimoto, A.C.; Shafer, L.A.; Dow, S.; Berry, M.J.; Hoffmann, P.R. Dietary Selenium Modulates Activation and Differentiation of CD4+ T Cells in Mice through a Mechanism Involving Cellular Free Thiols. J. Nutr. 2010, 140, 1155–1161. [Google Scholar] [CrossRef] [PubMed]
- Conrad, M.; Schneider, M.; Seiler, A.; Bornkamm, G.W. Physiological role of phospholipid hydroperoxide glutathione peroxidase in mammals. Biol. Chem. 2007, 388, 1019–1025. [Google Scholar] [CrossRef]
- Mittag, J.; Behrends, T.; Hoefig, C.S.; Vennstrom, B.; Schomburg, L. Thyroid Hormones Regulate Selenoprotein Expression and Selenium Status in Mice. PLoS ONE 2010, 5, e12931. [Google Scholar] [CrossRef]
- Short, S.P.; Pilat, J.M.; Williams, C.S. Roles for selenium and selenoprotein P in the development, progression, and prevention of intestinal disease. Free Radic. Biol. Med. 2018, 127, 26–35. [Google Scholar] [CrossRef]
- Bosschaerts, T.; Guilliams, M.; Noel, W.; Herin, M.; Burk, R.F.; Hill, K.E.; Brys, L.; Raes, G.; Ghassabeh, G.H.; de Baetselier, P.; et al. Alternatively activated myeloid cells limit pathogenicity associated with African trypanosomiasis through the IL-10 inducible gene selenoprotein P. J. Immunol. 2008, 180, 6168–6175. [Google Scholar] [CrossRef] [PubMed]
- Panee, J.; Stoytcheva, Z.R.; Liu, W.; Berry, M.J. Selenoprotein H is redox-sensing high mobility group family DNA-binding protein that up-regulates genes involved in glutathione synthesis and phase II detoxification. J. Biol. Chem. 2007, 282, 23759–23765. [Google Scholar] [CrossRef] [PubMed]
- Ren, B.; Liu, M.; Ni, J.; Tian, J. Role of Selenoprotein F in Protein Folding and Secretion: Potential Involvement in Human Disease. Nutrients 2018, 10, 1619. [Google Scholar] [CrossRef] [PubMed]
- Yim, S.H.; Everley, R.A.; Schildberg, F.A.; Lee, S.G.; Orsi, A.; Barbati, Z.R.; Karatepe, K.; Fomenko, D.E.; Tsuji, P.A.; Luo, H.R.; et al. Role of Selenof as a Gatekeeper of Secreted Disulfide-Rich Glycoproteins. Cell Rep. 2018, 23, 1387–1398. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.J.; Park, J.S.; Yoo, H.J.; Lee, H.M.; Lee, B.C.; Kim, J.H. The Selenoprotein MSRB1 Instructs Dendritic Cells to Induce T-Helper 1 Immune Responses. Antioxidants 2020, 9, 1021. [Google Scholar] [CrossRef]
- Ma, C.; Hoffmann, F.W.; Marciel, M.P.; Page, K.E.; Williams-Aduja, M.A.; Akana, E.N.L.; Gojanovich, G.S.; Gerschenson, M.; Urschitz, J.; Moisyadi, S.; et al. Upregulated ethanolamine phospholipid synthesis via selenoprotein I is required for effective metabolic reprogramming during T cell activation. Mol. Metab. 2021, 47, 101170. [Google Scholar] [CrossRef]
- Xu, X.M.; Carlson, B.A.; Irons, R.; Mix, H.; Zhong, N.; Gladyshev, V.N.; Hatfield, D.L. Selenophosphate synthetase 2 is essential for selenoprotein biosynthesis. Biochem. J. 2007, 404, 115–120. [Google Scholar] [CrossRef]
- Sengupta, A.; Carlson, B.A.; Labunskyy, V.; Gladyshev, V.N.; Hatfield, D.L. Knocking down selenoprotein T (SelT) alters cell adhesion and elevates selenoprotein W (SelW) expression in murine fibroblast cells. FASEB J. 2009, 23, 728. [Google Scholar] [CrossRef]
- Pan, T.; Liu, T.; Tan, S.; Wan, N.; Zhang, Y.; Li, S. Lower Selenoprotein T Expression and Immune Response in the Immune Organs of Broilers with Exudative Diathesis Due to Selenium Deficiency. Biol. Trace Elem. Res. 2018, 182, 364–372. [Google Scholar] [CrossRef] [PubMed]
- Yu, D.; Li, J.-L.; Zhang, J.; Gao, X.; Xu, S. Effects of Dietary Selenium on Selenoprotein W Gene Expression in the Chicken Immune Organs. Biol. Trace Elem. Res. 2011, 144, 678–687. [Google Scholar] [CrossRef]
- Yu, D.; Zhang, Z.; Yao, H.; Li, S.; Xu, S.W. The role of selenoprotein W in inflammatory injury in chicken immune tissues and cultured splenic lymphocyte. Biometals 2015, 28, 75–87. [Google Scholar] [CrossRef] [PubMed]
- Labunskyy, V.M.; Hatfield, D.L.; Gladyshev, V.N. Selenoproteins: Molecular Pathways and Physiological Roles. Physiol. Rev. 2014, 94, 739–777. [Google Scholar] [CrossRef]
- Bjørklund, G.; Zou, L.; Wang, J.; Chasapis, C.T.; Peana, M. Thioredoxin reductase as a pharmacological target. Pharmacol. Res. 2021, 174, 105854. [Google Scholar] [CrossRef]
- Chandler, J.D.; Nichols, D.P.; Nick, J.A.; Hondal, R.J.; Day, B.J. Selective Metabolism of Hypothiocyanous Acid by Mammalian Thioredoxin Reductase Promotes Lung Innate Immunity and Antioxidant Defense. J. Biol. Chem. 2013, 288, 18421–18428. [Google Scholar] [CrossRef]
- Rayman, M.P. Multiple nutritional factors and thyroid disease, with particular reference to autoimmune thyroid disease. Proc. Nutr. Soc. 2019, 78, 34–44. [Google Scholar] [CrossRef]
- Lobanov, A.V.; Hatfield, D.L.; Gladyshev, V.N. Eukaryotic selenoproteins and selenoproteomes. Biochim. Biophys. Acta-Gen. Subj. 2009, 1790, 1424–1428. [Google Scholar] [CrossRef]
- Anouar, Y.; Lihrmann, I.; Falluel-Morel, A.; Boukhzar, L. Selenoprotein T is a key player in ER proteostasis, endocrine homeostasis and neuroprotection. Free Radic. Biol. Med. 2018, 127, 145–152. [Google Scholar] [CrossRef]
- Sun, Z.; Liu, C.; Pan, T.; Yao, H.; Li, S. Selenium accelerates chicken dendritic cells differentiation and affects selenoproteins expression. Dev. Comp. Immunol. 2017, 77, 30–37. [Google Scholar] [CrossRef]
- Jiang, B.; Moskovitz, J. The Functions of the Mammalian Methionine Sulfoxide Reductase System and Related Diseases. Antioxidants 2018, 7, 122. [Google Scholar] [CrossRef]
- Jia, Y.; Zhang, L.; Liu, X.; Zhang, S.; Dai, J.; Huang, J.; Chen, J.; Wang, Y.; Zhou, J.; Zeng, Z. Selenium can regulate the differentiation and immune function of human dendritic cells. Biometals 2021, 34, 1365–1379. [Google Scholar] [CrossRef]
- Lee, B.C.; Peterfi, Z.; Hoffmann, F.W.; Moore, R.E.; Kaya, A.; Avanesov, A.; Tarrago, L.; Zhou, Y.; Weerapana, E.; Fomenko, D.E.; et al. MSRB1 and MICALs Regulate Actin Assembly and Macrophage Function via Reversible Stereoselective Methionine Oxidation. Mol. Cell 2013, 51, 397–404. [Google Scholar] [CrossRef]
- Fredericks, G.J.; Hoffmann, F.W.; Hondal, R.J.; Rozovsky, S.; Urschitz, J.; Hoffmann, P.R. Selenoprotein K Increases Efficiency of DHHC6 Catalyzed Protein Palmitoylation by Stabilizing the Acyl-DHHC6 Intermediate. Antioxidants 2018, 7, 4. [Google Scholar] [CrossRef] [PubMed]
- Fredericks, G.J.; Hoffmann, F.W.; Rose, A.H.; Osterheld, H.J.; Hess, F.M.; Mercier, F.; Hoffmann, P.R. Stable expression and function of the inositol 1,4,5-triphosphate receptor requires palmitoylation by a DHHC6/selenoprotein K complex. Proc. Natl. Acad. Sci. USA 2014, 111, 16478–16483. [Google Scholar] [CrossRef]
- Lee, B.C.; Lee, S.G.; Choo, M.K.; Kim, J.H.; Lee, H.M.; Kim, S.; Fomenko, D.E.; Kim, H.Y.; Park, J.M.; Gladyshev, V.N. Selenoprotein MSRB1 promotes anti-inflammatory cytokine gene expression in macrophages and controls immune response in vivo. Sci. Rep. 2017, 7, 5119. [Google Scholar] [CrossRef]
- Ferguson, A.D.; Labunskyy, V.M.; Fomenko, D.E.; Arac, D.; Chelliah, Y.; Amezcua, C.A.; Rizo, J.; Gladyshev, V.N.; Deisenhofer, J. NMR structures of the selenoproteins Sep15 and SelM reveal redox activity of a new thioredoxin-like family. J. Biol. Chem. 2006, 281, 3536–3543. [Google Scholar] [CrossRef]
- Turcanu, V.; Brough, H.A.; Toit, G.D.; Foong, R.X.; Marrs, T.; Santos, A.F.; Lack, G. Immune mechanisms of food allergy and its prevention by early intervention. Curr. Opin. Immunol. 2017, 48, 92–98. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Xiao, H.; Zhang, X.; Zhou, P. Insight into the effects of deglycosylation and glycation of shrimp tropomyosin on in vivo allergenicity and mast cell function. Food Funct. 2019, 10, 3934–3941. [Google Scholar] [CrossRef] [PubMed]
- Skoczylas, D.; Gujski, M.; Bojar, I.; Raciborski, F. Importance of food allergy and food intolerance in allergic multimorbidity. Ann. Agric. Environ. Med. 2020, 27, 413–417. [Google Scholar] [CrossRef]
- Frischmeyer-Guerrerio, P.A.; Guerrerio, A.L.; Chichester, K.L.; Bieneman, A.P.; Hamilton, R.A.; Wood, R.A.; Schroeder, J.T. Dendritic cell and T cell responses in children with food allergy. Clin. Exp. Allergy 2011, 41, 61–71. [Google Scholar] [CrossRef][Green Version]
- Safaralizadeh, R.; Nourizadeh, M.; Zare, A.; Kardar, G.A.; Pourpak, Z. Influence of selenium on mast cell mediator release. Biol. Trace Elem. Res. 2013, 154, 299–303. [Google Scholar] [CrossRef]
- Jian, S.-W.; Mei, C.-E.; Liang, Y.-N.; Li, D.; Chen, Q.-L.; Luo, H.-L.; Li, Y.-Q.; Cai, T.-Y. Influence of selenium-rich rice on transformation of umbilical blood B lymphocytes by Epstein-Barr virus and Epstein-Barr virus early antigen expression. Chin. J. Cancer 2003, 22, 26–29. [Google Scholar]
- Li, S.; Sun, W.; Zhang, K.; Zhu, J.; Jia, X.; Guo, X.; Zhao, Q.; Tang, C.; Yin, J.; Zhang, J. Selenium deficiency induces spleen pathological changes in pigs by decreasing selenoprotein expression, evoking oxidative stress, and activating inflammation and apoptosis. J. Anim. Sci. Biotechnol. 2021, 12, 65. [Google Scholar] [CrossRef] [PubMed]
- Minetti, G. Mevalonate pathway, selenoproteins, redox balance, immune system, COVID-19: Reasoning about connections. Med. Hypotheses 2020, 144, 110128. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Xu, Z.; Wang, D.; Yao, H.; Li, S. Selenium deficiency inhibits differentiation and immune function and imbalances the Th1/Th2 of dendritic cells. Metallomics 2018, 10, 759–767. [Google Scholar] [CrossRef] [PubMed]
- Xia, H.; Zhang, L.; Dai, J.; Liu, X.; Zhang, X.; Zeng, Z.; Jia, Y. Effect of Selenium and Peroxynitrite on Immune Function of Immature Dendritic Cells in Humans. Med. Sci. Monit. 2021, 27, e929004. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Xia, H.; Xia, K.; Liu, X.; Zhang, X.; Dai, J.; Zeng, Z.; Jia, Y. Selenium Regulation of the Immune Function of Dendritic Cells in Mice Through the ERK, Akt and RhoA/ROCK Pathways. Biol. Trace Elem. Res. 2021, 199, 3360–3370. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Zhao, Q.; Zhang, K.; Sun, W.; Jia, X.; Yang, Y.; Yin, J.; Tang, C.; Zhang, J. Se deficiency induces renal pathological changes by regulating selenoprotein expression, disrupting redox balance and activating inflammation. Metallomics 2020, 12, 1576–1584. [Google Scholar] [CrossRef]
- Mou, D.; Ding, D.; Yang, M.; Jiang, X.; Zhao, L.; Che, L.; Fang, Z.; Xu, S.; Lin, Y.; Zhuo, Y.; et al. Maternal organic selenium supplementation during gestation improves the antioxidant capacity and reduces the inflammation level in the intestine of offspring through the NF-kappa B and ERK/Beclin-1 pathways. Food Funct. 2021, 12, 315–327. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Wang, J.; Huang, K.; Liu, Q.; Liu, G.; Xu, X.; Zhang, H.; Zhu, M. Selenium-enriched Bacillus subtilis yb-114246 improved growth and immunity of broiler chickens through modified ileal bacterial composition. Sci. Rep. 2021, 11, 21690. [Google Scholar] [CrossRef]
- Zhang, R.; Guo, R.; Liu, Q.; Li, G.; Sun, B.; Huang, X. Selenium Deficiency via the TLR4/TRIF/NF-kappa B Signaling Pathway Leading to Inflammatory Injury in Chicken Spleen. Biol. Trace Elem. Res. 2021, 199, 693–702. [Google Scholar] [CrossRef] [PubMed]
- Yiming, Z.; Qingqing, L.; Hang, Y.; Yahong, M.; Li, S. Selenium deficiency causes immune damage by activating the DUSP1/NF-kappa B pathway and endoplasmic reticulum stress in chicken spleen. Food Funct. 2020, 11, 6467–6475. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhang, L.; Xia, K.; Dai, J.; Huang, J.; Wang, Y.; Zhu, G.; Hu, Z.; Zeng, Z.; Jia, Y. Effects of dietary selenium on immune function of spleen in mice. J. Funct. Food. 2022, 89, 104914. [Google Scholar] [CrossRef]
- Yang, Z.; Liu, C.; Liu, C.; Teng, X.; Li, S. Selenium Deficiency Mainly Influences Antioxidant Selenoproteins Expression in Broiler Immune Organs. Biol. Trace Elem. Res. 2016, 172, 209–221. [Google Scholar] [CrossRef]
- Mittal, M.; Siddiqui, M.R.; Tran, K.; Reddy, S.P.; Malik, A.B. Reactive oxygen species in inflammation and tissue injury. Antioxid. Redox Signal. 2014, 20, 1126–1167. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Roh, Y.J.; Han, S.-J.; Park, I.; Lee, H.M.; Ok, Y.S.; Lee, B.C.; Lee, S.-R. Role of Selenoproteins in Redox Regulation of Signaling and the Antioxidant System: A Review. Antioxidants 2020, 9, 383. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.Z.; Lee, S.H.; Lillehoj, H.S.; Bravo, D. Dietary sodium selenite affects host intestinal and systemic immune response and disease susceptibility to necrotic enteritis in commercial broilers. Br. Poult. Sci. 2015, 56, 103–112. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.H.; Pi, D.A.; Zhao, L.; Wang, X.Y.; Zhu, L.Y.; Qi, D.S.; Liu, Y.L. Response of Selenium and Selenogenome in Immune Tissues to LPS-Induced Inflammatory Reactions in Pigs. Biol. Trace Elem. Res. 2017, 177, 90–96. [Google Scholar] [CrossRef] [PubMed]
- Sicherer, S.H.; Sampson, H.A. Food allergy: A review and update on epidemiology, pathogenesis, diagnosis, prevention, and management. J. Allergy Clin. Immunol. 2018, 141, 41–58. [Google Scholar] [CrossRef] [PubMed]
- Chinthrajah, R.S.; Hernandez, J.D.; Boyd, S.D.; Galli, S.J.; Nadeau, K.C. Molecular and cellular mechanisms of food allergy and food tolerance. J. Allergy Clin. Immunol. 2016, 137, 984–997. [Google Scholar] [CrossRef]
- Hoffmann, P.R.; Saux, C.J.; Hoffmann, F.W.; Chang, P.S.; Bollt, O.; He, Q.; Tam, E.K.; Berry, M.J. A role for dietary selenium and selenoproteins in allergic airway inflammation. J. Immunol. 2007, 179, 3258–3267. [Google Scholar] [CrossRef]
- Kalita, B.; Nowak, P.; Slimok, M.; Sikora, A.; Szkilnik, R.; Obuchowicz, A.; Sulej, J.; Sabat, D. Selenium plasma concentration level in children with food allergy. Polski. Merkur. Lek. Organ Pol. Tow. Lek. 2001, 10, 411–413. [Google Scholar]
- Jia, S.Z.; Xu, X.W.; Zhang, Z.H.; Chen, C.; Chen, Y.B.; Huang, S.L.; Liu, Q.; Hoffmann, P.R.; Song, G.L. Selenoprotein K deficiency-induced apoptosis: A role for calpain and the ERS pathway. Redox Biol. 2021, 47, 102154. [Google Scholar] [CrossRef]
- Ivory, K.; Prieto, E.; Spinks, C.; Armah, C.N.; Goldson, A.J.; Dainty, J.R.; Nicoletti, C. Selenium supplementation has beneficial and detrimental effects on immunity to influenza vaccine in older adults. Clin. Nutr. 2017, 36, 407–415. [Google Scholar] [CrossRef] [PubMed]
- Vitamin and Mineral Requirements in Human Nutrition: Report of a Joint FAO/WHO Expert Consultation, Bangkok, Thailand, 21–30 September 1998. Available online: https://www.webofscience.com/wos/alldb/full-record/CABI:20053144499 (accessed on 21 June 2022).
- Wang, Y.; Jiang, L.; He, J.; Hu, M.; Zeng, F.; Li, Y.; Tian, H.; Luo, X. The Adverse Effects of Se Toxicity on Inflammatory and Immune Responses in Chicken Spleens. Biol. Trace Elem. Res. 2018, 185, 170–176. [Google Scholar] [CrossRef] [PubMed]
- Wadhwani, S.A.; Shedbalkar, U.U.; Singh, R.; Chopade, B.A. Biogenic selenium nanoparticles: Current status and future prospects. Appl. Microbiol. Biotechnol. 2016, 100, 2555–2566. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, K.; Yang, S.; Li, T.; Huang, X.; Dong, Y.; Wang, P.; Huang, J. Advances in the Study of the Mechanism by Which Selenium and Selenoproteins Boost Immunity to Prevent Food Allergies. Nutrients 2022, 14, 3133. https://doi.org/10.3390/nu14153133
Zhu K, Yang S, Li T, Huang X, Dong Y, Wang P, Huang J. Advances in the Study of the Mechanism by Which Selenium and Selenoproteins Boost Immunity to Prevent Food Allergies. Nutrients. 2022; 14(15):3133. https://doi.org/10.3390/nu14153133
Chicago/Turabian StyleZhu, Kongdi, Shihui Yang, Tong Li, Xin Huang, Yulan Dong, Pengjie Wang, and Jiaqiang Huang. 2022. "Advances in the Study of the Mechanism by Which Selenium and Selenoproteins Boost Immunity to Prevent Food Allergies" Nutrients 14, no. 15: 3133. https://doi.org/10.3390/nu14153133
APA StyleZhu, K., Yang, S., Li, T., Huang, X., Dong, Y., Wang, P., & Huang, J. (2022). Advances in the Study of the Mechanism by Which Selenium and Selenoproteins Boost Immunity to Prevent Food Allergies. Nutrients, 14(15), 3133. https://doi.org/10.3390/nu14153133