
The Use of Dietary Supplements and Amino Acid Restriction Interventions to Reduce Frailty in Pre-Clinical Models


Abstract
1. Introduction
2. Frailty
2.1. Definition of Frailty
2.2. Measuring Frailty
2.3. Putative Frailty Mechanisms
3. Dietary Interventions
3.1. Vitamin Supplements
3.1.1. Vitamin D
3.1.2. Vitamin C
3.1.3. Vitamin E
3.1.4. Nicotinamide

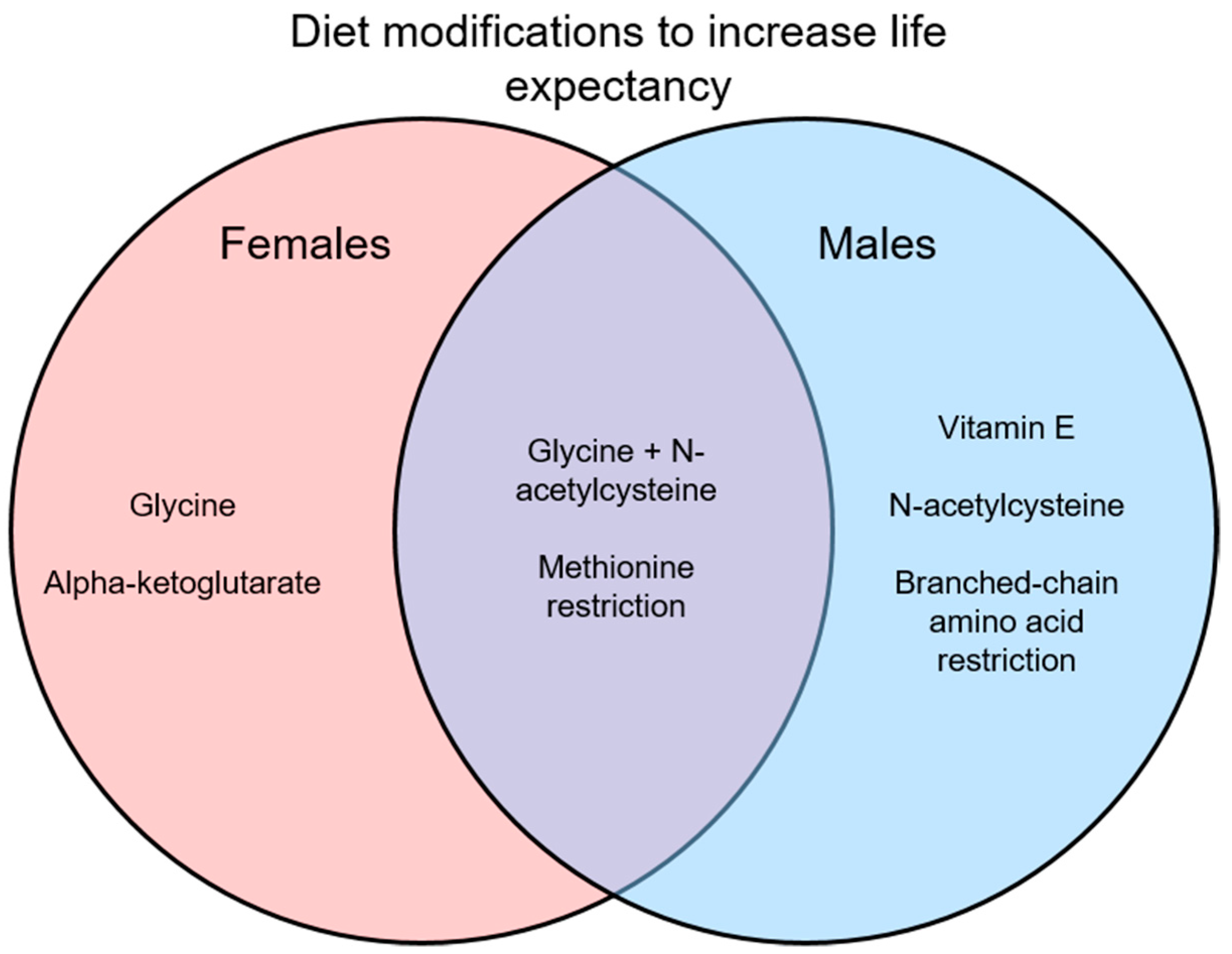{kind=link}
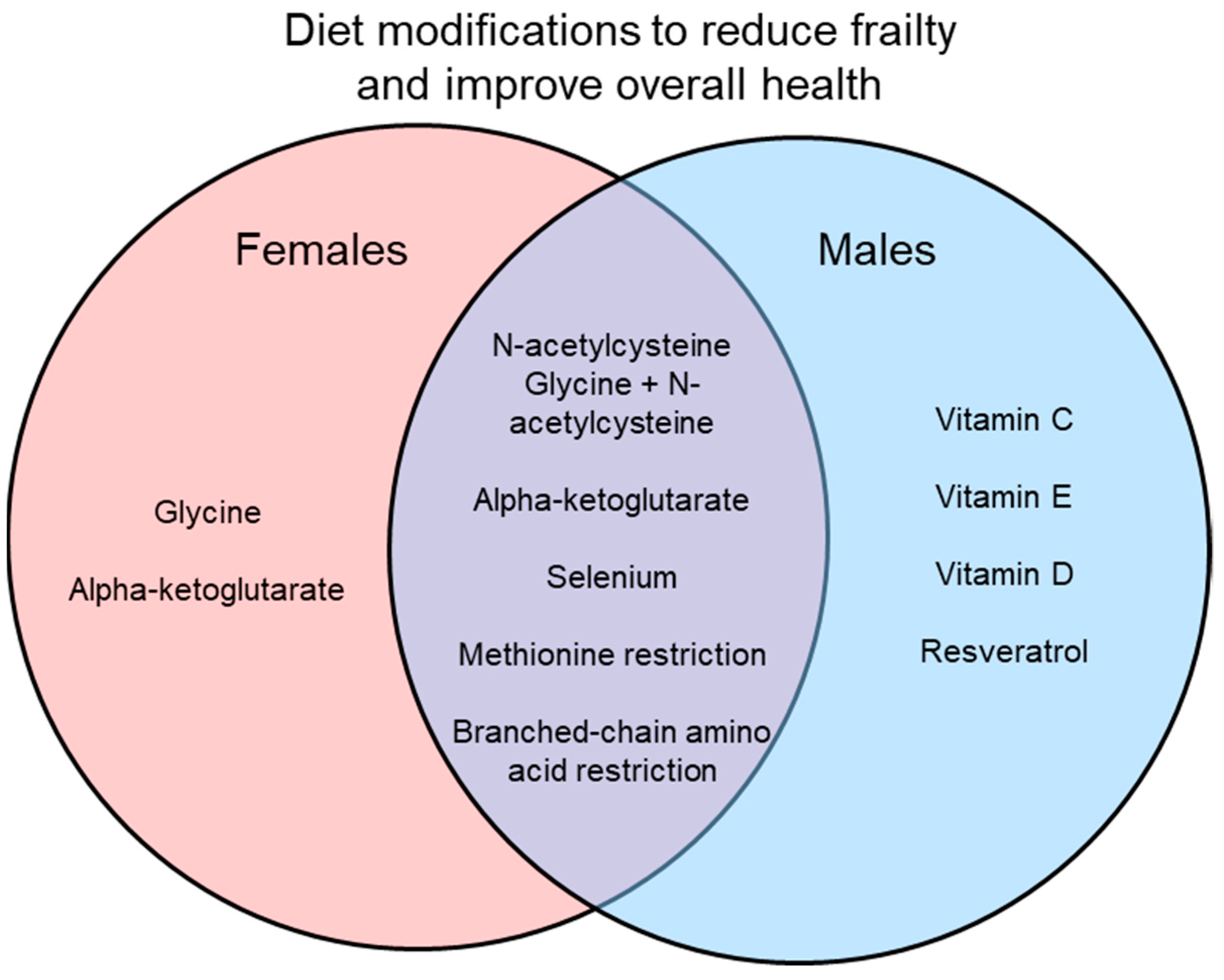{kind=link}
| Intervention 1 | Strain/ Species | Sex | Age (Mos) | Intervention | Health Assessment | Main Results | References |
|---|---|---|---|---|---|---|---|
| Vitamin D3 | C57Bl/6 mice | Male | 6 | 125 IU or 1000 IU of vitamin D3 for 12 months | Physical performance |
| [45] |
| Fischer 344 rats | Male | 6 or 13 | 0.045 µ/kg vitamin D3 for 7 months | 27 item frailty index |
| [47] | |
| C57Bl/6 mice | Male | 24 | 125 IU, 1000 IU or 8000 IU vitamin D3 for 4 months | Frailty phenotype |
| [46] | |
| Vitamin C | Cold stressed C57Bl/6 mice | Female | 4 | 180 mg/kg vitamin C for 18 months | Oxidative damage and lifespan |
| [55] |
| SMP30-KO and C57BL/6 mice | Male | 0.5 | 20 mg/kg or 200 mg/kg vitamin C for 2 months | Immune function |
| [57] | |
| Vitamin E | CD1/UCadiz | Both | 7 | 5.0 g/kg of vitamin E for lifelong | Lifespan, physical and neurological performance |
| [63] |
| Nicotinamide | C57Bl/6 mice | Male | 12 | 0.5 g/kg nicotinamide with high or low-fat diet for 15.5 months | Lifespan and metabolism |
| [70] |
3.2. Non-Vitamin Supplements
3.2.1. Allicin
3.2.2. Glycine
3.2.3. N-Acetylcysteine
3.2.4. Alpha-Ketoglutarate
3.2.5. Selenium
3.2.6. Resveratrol
| Intervention 1 | Strain/ Species | Sex | Age (Mos) | Intervention | Health Assessment | Main Results | References |
|---|---|---|---|---|---|---|---|
| Allicin | Fischer 344 rats | Male | 6 and 13 | 4 mg/kg, 8 mg/kg, and 16 mg/kg allicin for 8 months | 27-item frailty index |
| [47] |
| Glycine | UM-HET3 mice | Both | 9 | 8% glycine for lifelong | Lifespan and body composition |
| [80] |
| N-acetylcysteine | HET3 mice | Both | 7 | 5 g/L or 10 g/L of N-acetylcysteine for lifelong | Lifespan and body composition |
| [86] |
| GlyNAC | C57Bl/6 mice | Both | 14.9 | 1.6 mg/g glycine and 1.6 mg/g N-acetylcysteine for lifelong | Lifespan and metabolism |
| [81] |
| Alpha-ketoglutarate | C57Bl/6 mice | Both | 18 | 2% per weight alpha-ketoglutarate for lifelong | Lifespan and frailty index |
| [92] |
| Selenium | ICR mice | Female | 12 | 0.15 mg/kg or 0.33 mg/kg sodium selenite or selenium yeast for 6 weeks | Reproductive physiology |
| [97] |
| C57Bl/6 mice | Both | 20 | 5 mg/kg nanoSelenium food for 2 months | Physical performance |
| [98] | |
| C57Bl/6 mice | Both | 2 | 0.0073% sodium selenite or 0.0037% selenomethionine for 4 months | Body composition, serum hormones |
| [99] | |
| C57Bl/6 mice | Female | 12 or 18 | 50nM seleno-L-methionine for 28 days or 4 weeks | Neurogenesis, and memory tests |
| [100] | |
| Resveratrol | C57Bl/6 mice | Male | 18 | 100 mg/kg for 6 months | Frailty Index |
| [111] |
| C57Bl/6 mice | Male | 16 | 25 mg/kg for 28 days | Physical performance |
| [112] | |
| C57Bl/6J mice | Male | 2, 12 or 18 | 16–17 mg/kg for 4.5 months | Physical performance |
| [113] | |
| Mice | Male | 12 or 18 | 15 mg/kg for 4 weeks | Physical performance |
| [114] |
3.3. Amino Acid Restriction Diets
3.3.1. Methionine Restriction Diets
3.3.2. Branched-Chain Amino Acid Restriction
| Intervention 1 | Strain/Species | Sex | Age (Mos) | Intervention | Health Assessment | Main Results | References |
|---|---|---|---|---|---|---|---|
| Methionine restriction | HGPS mice | Both | 1.5 | 0.12% methionine restriction, lifetime | Lifespan and metabolism |
| [119] |
| C57Bl/6Nia mice | Male | 21 | 0.1% methionine restriction for 6 months | Rockwood Frailty Index |
| [122] | |
| C57Bl/6 mice | Both | 21 | 0.1% methionine restriction for 6 months | Frailty Index |
| [121] | |
| Branched chain amino acid restriction | C57Bl/6Nia mice | Both | 16 | Reduced amino acid diet or low-branched amino acid (BCAA) diet for life | 26 item frailty index |
| [126] |
4. Conclusions, Limitations and Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Statistics Canada, Government of Canada. Use of Nutritional Supplements. 2015. Available online: https://www150.statcan.gc.ca/n1/pub/82-625-x/2017001/article/14831-eng.htm (accessed on 7 April 2022).
- Hernández Morante, J.J.; Gómez Martínez, C.; Morillas-Ruiz, J.M. Dietary Factors Associated with Frailty in Old Adults: A Review of Nutritional Interventions to Prevent Frailty Development. Nutrients 2019, 11, 102. [Google Scholar] [CrossRef] [PubMed]
- WHO Clinical Consortium on Healthy Ageing. Report of Consortium Meeting 1–2 December 2016 in Geneva, Switzerland; World Health Organization: Geneva, Switzerland, 2017.
- Clegg, A.; Young, J.; Iliffe, S.; Rikkert, M.O.; Rockwood, K. Frailty in Older People. Lancet 2013, 381, 752–762. [Google Scholar] [CrossRef]
- Song, X.; Mitnitski, A.; Rockwood, K. Prevalence and 10-Year Outcomes of Frailty in Older Adults in Relation to Deficit Accumulation. J. Am. Geriatr. Soc. 2010, 58, 681–687. [Google Scholar] [CrossRef] [PubMed]
- Kane, A.E.; Howlett, S.E. Sex Differences in Frailty: Comparisons between Humans and Preclinical Models. Mech. Ageing Dev. 2021, 198, 111546. [Google Scholar] [CrossRef] [PubMed]
- Keller, K.; Kane, A.; Heinze-Milne, S.; Grandy, S.A.; Howlett, S.E. Chronic Treatment With the ACE Inhibitor Enalapril Attenuates the Development of Frailty and Differentially Modifies Pro- and Anti-Inflammatory Cytokines in Aging Male and Female C57BL/6 Mice. J. Gerontol. A Biol. Sci. Med. Sci. 2019, 74, 1149–1157. [Google Scholar] [CrossRef] [PubMed]
- Correia-Melo, C.; Birch, J.; Fielder, E.; Rahmatika, D.; Taylor, J.; Chapman, J.; Lagnado, A.; Carroll, B.M.; Miwa, S.; Richardson, G.; et al. Rapamycin Improves Healthspan but Not Inflammaging in Nfκb1−/− Mice. Aging Cell 2019, 18, e12882. [Google Scholar] [CrossRef]
- Bisset, E.S.; Heinze-Milne, S.; Grandy, S.A.; Howlett, S.E. Aerobic Exercise Attenuates Frailty in Aging Male and Female C57Bl/6 Mice and Effects Systemic Cytokines Differentially by Sex. J. Gerontol. A Biol. Sci. Med. Sci. 2022, 77, 41–46. [Google Scholar] [CrossRef]
- Seldeen, K.L.; Lasky, G.; Leiker, M.M.; Pang, M.; Personius, K.E.; Troen, B.R. High Intensity Interval Training Improves Physical Performance and Frailty in Aged Mice. J. Gerontol. A Biol. Sci. Med. Sci. 2018, 73, 429–437. [Google Scholar] [CrossRef]
- Mitnitski, A.B.; Mogilner, A.J.; Rockwood, K. Accumulation of Deficits as a Proxy Measure of Aging. Sci. World J. 2001, 1, 323–336. [Google Scholar] [CrossRef]
- Whitehead, J.C.; Hildebrand, B.A.; Sun, M.; Rockwood, M.R.; Rose, R.A.; Rockwood, K.; Howlett, S.E. A Clinical Frailty Index in Aging Mice: Comparisons with Frailty Index Data in Humans. J. Gerontol. A Biol. Sci. Med. Sci. 2014, 69, 621–632. [Google Scholar] [CrossRef]
- Yorke, A.; Kane, A.E.; Hancock Friesen, C.L.; Howlett, S.E.; O’Blenes, S. Development of a Rat Clinical Frailty Index. J. Gerontol. A Biol. Sci. Med. Sci. 2017, 72, 897–903. [Google Scholar] [CrossRef] [PubMed]
- Banzato, T.; Franzo, G.; Di Maggio, R.; Nicoletto, E.; Burti, S.; Cesari, M.; Canevelli, M. A Frailty Index Based on Clinical Data to Quantify Mortality Risk in Dogs. Sci. Rep. 2019, 9, 16749. [Google Scholar] [CrossRef] [PubMed]
- Howlett, S.E.; Rutenberg, A.D.; Rockwood, K. The Degree of Frailty as a Translational Measure of Health in Aging. Nat. Aging 2021, 1, 651–665. [Google Scholar] [CrossRef]
- Fried, L.P.; Tangen, C.M.; Walston, J.; Newman, A.B.; Hirsch, C.; Gottdiener, J.; Seeman, T.; Tracy, R.; Kop, W.J.; Burke, G.; et al. Frailty in Older Adults: Evidence for a Phenotype. J. Gerontol. A Biol. Sci. Med. Sci. 2001, 56, M146–M156. [Google Scholar] [CrossRef]
- Liu, H.; Graber, T.G.; Ferguson-Stegall, L.; Thompson, L.V. Clinically Relevant Frailty Index for Mice. J. Gerontol. A Biol. Sci. Med. Sci. 2014, 69, 1485–1491. [Google Scholar] [CrossRef]
- Miller, M.G.; Thangthaeng, N.; Shukitt-Hale, B. A Clinically Relevant Frailty Index for Aging Rats. J. Gerontol. A Biol. Sci. Med. Sci. 2017, 72, 892–896. [Google Scholar] [CrossRef]
- Yamada, Y.; Kemnitz, J.W.; Weindruch, R.; Anderson, R.M.; Schoeller, D.A.; Colman, R.J. Caloric Restriction and Healthy Life Span: Frail Phenotype of Nonhuman Primates in the Wisconsin National Primate Research Center Caloric Restriction Study. J. Gerontol. A Biol. Sci. Med. Sci. 2018, 73, 273–278. [Google Scholar] [CrossRef]
- Hua, J.; Hoummady, S.; Muller, C.; Pouchelon, J.-L.; Blondot, M.; Gilbert, C.; Desquilbet, L. Assessment of Frailty in Aged Dogs. Am. J. Vet. Res. 2016, 77, 1357–1365. [Google Scholar] [CrossRef]
- Kane, A.E.; Huizer-Pajkos, A.; Mach, J.; Mitchell, S.J.; de Cabo, R.; Le Couteur, D.G.; Howlett, S.E.; Hilmer, S.N. A Comparison of Two Mouse Frailty Assessment Tools. J. Gerontol. A Biol. Sci. Med. Sci. 2017, 72, 904–909. [Google Scholar] [CrossRef]
- Bisset, E.S.; Howlett, S.E. The Biology of Frailty in Humans and Animals: Understanding Frailty and Promoting Translation. Aging Med. 2019, 2, 27–34. [Google Scholar] [CrossRef]
- Goh, J.; Wong, E.; Soh, J.; Maier, A.B.; Kennedy, B.K. Targeting the Molecular & Cellular Pillars of Human Aging with Exercise. FEBS J. 2021. [Google Scholar] [CrossRef]
- Franceschi, C.; Bonafè, M.; Valensin, S.; Olivieri, F.; De Luca, M.; Ottaviani, E.; De Benedictis, G. Inflamm-Aging: An Evolutionary Perspective on Immunosenescence. Ann. N. Y. Acad. Sci. 2006, 908, 244–254. [Google Scholar] [CrossRef] [PubMed]
- Lai, H.-Y.; Chang, H.; Lee, Y.L.; Hwang, S.-J. Association between Inflammatory Markers and Frailty in Institutionalized Older Men. Maturitas 2014, 79, 329–333. [Google Scholar] [CrossRef] [PubMed]
- Kane, A.E.; Keller, K.M.; Heinze-Milne, S.; Grandy, S.A.; Howlett, S.E. A Murine Frailty Index Based on Clinical and Laboratory Measurements: Links Between Frailty and Pro-Inflammatory Cytokines Differ in a Sex-Specific Manner. J. Gerontol. A Biol. Sci. Med. Sci. 2019, 74, 275–282. [Google Scholar] [CrossRef] [PubMed]
- Márquez, E.J.; Chung, C.; Marches, R.; Rossi, R.J.; Nehar-Belaid, D.; Eroglu, A.; Mellert, D.J.; Kuchel, G.A.; Banchereau, J.; Ucar, D. Sexual-Dimorphism in Human Immune System Aging. Nat. Commun. 2020, 11, 751. [Google Scholar] [CrossRef]
- Franceschi, C.; Campisi, J. Chronic Inflammation (Inflammaging) and Its Potential Contribution to Age-Associated Diseases. J. Gerontol. A Biol. Sci. Med. Sci. 2014, 69 (Suppl. S1), S4–S9. [Google Scholar] [CrossRef]
- Branch, L.S. Consolidated Federal Laws of Canada, Natural Health Products Regulations. Available online: https://laws-lois.justice.gc.ca/eng/regulations/sor-2003-196/FullText.html (accessed on 7 April 2022).
- Mitchell, S.J.; Mitchell, G.J.; Mitchell, J.R. Modulation of Frailty Syndrome by Diet: A Review of Evidence from Mouse Studies. Mech. Ageing Dev. 2019, 180, 82–88. [Google Scholar] [CrossRef]
- Wolf, A.M. Rodent Diet Aids and the Fallacy of Caloric Restriction. Mech. Ageing Dev. 2021, 200, 111584. [Google Scholar] [CrossRef]
- DeLuca, H.F. Overview of General Physiologic Features and Functions of Vitamin D. Am. J. Clin. Nutr. 2004, 80 (Suppl. S6), 1689S–1696S. [Google Scholar] [CrossRef]
- Ceglia, L.; Harris, S.S. Vitamin D and Its Role in Skeletal Muscle. Calcif. Tissue Int. 2013, 92, 151–162. [Google Scholar] [CrossRef]
- Lips, P.; van Schoor, N.M. The Effect of Vitamin D on Bone and Osteoporosis. Best Pract. Res. Clin. Endocrinol. Metab. 2011, 25, 585–591. [Google Scholar] [CrossRef] [PubMed]
- Barbarawi, M.; Kheiri, B.; Zayed, Y.; Barbarawi, O.; Dhillon, H.; Swaid, B.; Yelangi, A.; Sundus, S.; Bachuwa, G.; Alkotob, M.L.; et al. Vitamin D Supplementation and Cardiovascular Disease Risks in More Than 83 000 Individuals in 21 Randomized Clinical Trials: A Meta-Analysis. JAMA Cardiol. 2019, 4, 765. [Google Scholar] [CrossRef] [PubMed]
- Verstuyf, A.; Carmeliet, G.; Bouillon, R.; Mathieu, C. Vitamin D: A Pleiotropic Hormone. Kidney Int. 2010, 78, 140–145. [Google Scholar] [CrossRef] [PubMed]
- Yoshizawa, T.; Handa, Y.; Uematsu, Y.; Takeda, S.; Sekine, K.; Yoshihara, Y.; Kawakami, T.; Arioka, K.; Sato, H.; Uchiyama, Y.; et al. Mice Lacking the Vitamin D Receptor Exhibit Impaired Bone Formation, Uterine Hypoplasia and Growth Retardation after Weaning. Nat. Genet. 1997, 16, 391–396. [Google Scholar] [CrossRef] [PubMed]
- Grundmann, S.M.; Brandsch, C.; Rottstädt, D.; Kühne, H.; Stangl, G.I. The High Calcium, High Phosphorus Rescue Diet Is Not Suitable to Prevent Secondary Hyperparathyroidism in Vitamin D Receptor Deficient Mice. Front. Physiol. 2017, 8, 212. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Dzik, K.P.; Kaczor, J.J. Mechanisms of Vitamin D on Skeletal Muscle Function: Oxidative Stress, Energy Metabolism and Anabolic State. Eur. J. Appl. Physiol. 2019, 119, 825–839. [Google Scholar] [CrossRef]
- Keisala, T.; Minasyan, A.; Lou, Y.-R.; Zou, J.; Kalueff, A.V.; Pyykkö, I.; Tuohimaa, P. Premature Aging in Vitamin D Receptor Mutant Mice. J. Steroid Biochem. Mol. Biol. 2009, 115, 91–97. [Google Scholar] [CrossRef]
- Zhu, J.G.; Ochalek, J.T.; Kaufmann, M.; Jones, G.; DeLuca, H.F. CYP2R1 Is a Major, but Not Exclusive, Contributor to 25-Hydroxyvitamin D Production in Vivo. Proc. Natl. Acad. Sci. USA 2013, 110, 15650–15655. [Google Scholar] [CrossRef]
- Roizen, J.D.; Casella, A.; Lai, M.; Long, C.; Tara, Z.; Caplan, I.; O’Lear, L.; Levine, M.A. Decreased Serum 25-Hydroxyvitamin D in Aging Male Mice Is Associated With Reduced Hepatic Cyp2r1 Abundance. Endocrinology 2018, 159, 3083–3089. [Google Scholar] [CrossRef]
- Cielen, N.; Heulens, N.; Maes, K.; Carmeliet, G.; Mathieu, C.; Janssens, W.; Gayan-Ramirez, G. Vitamin D Deficiency Impairs Skeletal Muscle Function in a Smoking Mouse Model. J. Endocrinol. 2016, 229, 97–108. [Google Scholar] [CrossRef]
- Das, A.; Gopinath, S.D.; Arimbasseri, G.A. Systemic Ablation of Vitamin D Receptor Leads to Skeletal Muscle Glycogen Storage Disorder in Mice. J. Cachexia Sarcopenia Muscle 2022, 13, 467–480. [Google Scholar] [CrossRef] [PubMed]
- Seldeen, K.L.; Pang, M.; Leiker, M.M.; Bard, J.E.; Rodríguez-Gonzalez, M.; Hernandez, M.; Sheridan, Z.; Nowak, N.; Troen, B.R. Chronic Vitamin D Insufficiency Impairs Physical Performance in C57BL/6J Mice. Aging (Albany NY) 2018, 10, 1338–1355. [Google Scholar] [CrossRef] [PubMed]
- Seldeen, K.L.; Berman, R.N.; Pang, M.; Lasky, G.; Weiss, C.; MacDonald, B.A.; Thiyagarajan, R.; Redae, Y.; Troen, B.R. Vitamin D Insufficiency Reduces Grip Strength, Grip Endurance and Increases Frailty in Aged C57Bl/6J Mice. Nutrients 2020, 12, 3005. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; You, M.; Shen, J.; Xu, Y.; Li, L.; Wang, D.; Yang, Y. Allicin Reversed the Process of Frailty in Aging Male Fischer 344 Rats With Osteoporosis. J. Gerontol. A Biol. Sci. Med. Sci. 2020, 75, 821–825. [Google Scholar] [CrossRef] [PubMed]
- Padayatty, S.J.; Levine, M. Vitamin C: The Known and the Unknown and Goldilocks. Oral Dis. 2016, 22, 463–493. [Google Scholar] [CrossRef] [PubMed]
- Carr, A.C.; Maggini, S. Vitamin C and Immune Function. Nutrients 2017, 9, 1211. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Maeda, N.; Beck, M.A. Vitamin C Deficiency Increases the Lung Pathology of Influenza Virus-Infected Gulo−/− Mice. J. Nutr. 2006, 136, 2611–2616. [Google Scholar] [CrossRef]
- Baguley, B.C.; Ding, Q.; Richardson, E. Preliminary Evidence That High-Dose Vitamin C Has a Vascular Disrupting Action in Mice. Front. Oncol. 2014, 4, 310. [Google Scholar] [CrossRef]
- Massie, H.R.; Aiello, V.R.; Doherty, T.J. Dietary Vitamin C Improves the Survival of Mice. Gerontology 1984, 30, 371–375. [Google Scholar] [CrossRef]
- Maeda, N.; Hagihara, H.; Nakata, Y.; Hiller, S.; Wilder, J.; Reddick, R. Aortic Wall Damage in Mice Unable to Synthesize Ascorbic Acid. Proc. Natl. Acad. Sci. USA 2000, 97, 841–846. [Google Scholar] [CrossRef]
- Aumailley, L.; Warren, A.; Garand, C.; Dubois, M.J.; Paquet, E.R.; Le Couteur, D.G.; Marette, A.; Cogger, V.C.; Lebel, M. Vitamin C Modulates the Metabolic and Cytokine Profiles, Alleviates Hepatic Endoplasmic Reticulum Stress, and Increases the Life Span of Gulo−/− Mice. Aging (Albany NY) 2016, 8, 458–483. [Google Scholar] [CrossRef] [PubMed]
- Selman, C.; McLaren, J.S.; Meyer, C.; Duncan, J.S.; Redman, P.; Collins, A.R.; Duthie, G.G.; Speakman, J.R. Life-Long Vitamin C Supplementation in Combination with Cold Exposure Does Not Affect Oxidative Damage or Lifespan in Mice, but Decreases Expression of Antioxidant Protection Genes. Mech. Ageing Dev. 2006, 127, 897–904. [Google Scholar] [CrossRef] [PubMed]
- Lin, M.T.; Flint Beal, M. The Oxidative Damage Theory of Aging. Clin. Neurosci. Res. 2003, 2, 305–315. [Google Scholar] [CrossRef]
- Uchio, R.; Hirose, Y.; Murosaki, S.; Ishigami, A. High Dietary Vitamin C Intake Reduces Glucocorticoid-Induced Immunosuppression and Measures of Oxidative Stress in Vitamin C-Deficient Senescence Marker Protein 30 Knockout Mice. Br. J. Nutr. 2019, 122, 1120–1129. [Google Scholar] [CrossRef]
- Ishigami, A.; Fujita, T.; Handa, S.; Shirasawa, T.; Koseki, H.; Kitamura, T.; Enomoto, N.; Sato, N.; Shimosawa, T.; Maruyama, N. Senescence Marker Protein-30 Knockout Mouse Liver Is Highly Susceptible to Tumor Necrosis Factor-Alpha- and Fas-Mediated Apoptosis. Am. J. Pathol. 2002, 161, 1273–1281. [Google Scholar] [CrossRef]
- Uchio, R.; Hirose, Y.; Murosaki, S.; Yamamoto, Y.; Ishigami, A. High Dietary Intake of Vitamin C Suppresses Age-Related Thymic Atrophy and Contributes to the Maintenance of Immune Cells in Vitamin C-Deficient Senescence Marker Protein-30 Knockout Mice. Br. J. Nutr. 2015, 113, 603–609. [Google Scholar] [CrossRef]
- Han, S.N.; Meydani, S.N. Antioxidants, Cytokines, and Influenza Infection in Aged Mice and Elderly Humans. J. Infect. Dis. 2000, 182, S74–S80. [Google Scholar] [CrossRef]
- Fukui, K.; Nakamura, K.; Shirai, M.; Hirano, A.; Takatsu, H.; Urano, S. Long-Term Vitamin E-Deficient Mice Exhibit Cognitive Dysfunction via Elevation of Brain Oxidation. J. Nutr. Sci. Vitaminol. 2015, 61, 362–368. [Google Scholar] [CrossRef]
- Gohil, K.; Schock, B.C.; Chakraborty, A.A.; Terasawa, Y.; Raber, J.; Farese, R.V.; Packer, L.; Cross, C.E.; Traber, M.G. Gene Expression Profile of Oxidant Stress and Neurodegeneration in Transgenic Mice Deficient in Alpha-Tocopherol Transfer Protein. Free Radic. Biol. Med. 2003, 35, 1343–1354. [Google Scholar] [CrossRef]
- Navarro, A.; Gómez, C.; Sánchez-Pino, M.-J.; González, H.; Bández, M.J.; Boveris, A.D.; Boveris, A. Vitamin E at High Doses Improves Survival, Neurological Performance, and Brain Mitochondrial Function in Aging Male Mice. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2005, 289, R1392–R1399. [Google Scholar] [CrossRef]
- Fricker, R.A.; Green, E.L.; Jenkins, S.I.; Griffin, S.M. The Influence of Nicotinamide on Health and Disease in the Central Nervous System. Int. J. Tryptophan Res. 2018, 11, 1178646918776658. [Google Scholar] [CrossRef] [PubMed]
- Braidy, N.; Poljak, A.; Grant, R.; Jayasena, T.; Mansour, H.; Chan-Ling, T.; Guillemin, G.J.; Smythe, G.; Sachdev, P. Mapping NAD+ Metabolism in the Brain of Ageing Wistar Rats: Potential Targets for Influencing Brain Senescence. Biogerontology 2014, 15, 177–198. [Google Scholar] [CrossRef] [PubMed]
- Nadeeshani, H.; Li, J.; Ying, T.; Zhang, B.; Lu, J. Nicotinamide Mononucleotide (NMN) as an Anti-Aging Health Product—Promises and Safety Concerns. J. Adv. Res. 2022, 37, 267–278. [Google Scholar] [CrossRef] [PubMed]
- Shi, W.; Hegeman, M.A.; Doncheva, A.; van der Stelt, I.; Bekkenkamp-Grovenstein, M.; van Schothorst, E.M.; Brenner, C.; de Boer, V.C.J.; Keijer, J. Transcriptional Response of White Adipose Tissue to Withdrawal of Vitamin B3. Mol. Nutr. Food Res. 2019, 63, e1801100. [Google Scholar] [CrossRef]
- Van der Stelt, I.; Shi, W.; Bekkenkamp-Grovenstein, M.; Zapata-Pérez, R.; Houtkooper, R.H.; de Boer, V.C.J.; Hegeman, M.A.; Keijer, J. The Female Mouse Is Resistant to Mild Vitamin B3 Deficiency. Eur. J. Nutr. 2022, 61, 329–340. [Google Scholar] [CrossRef]
- Palliyaguru, D.L.; Moats, J.M.; di Germanio, C.; Bernier, M.; de Cabo, R. Frailty Index as a Biomarker of Lifespan and Healthspan: Focus on Pharmacological Interventions. Mech. Ageing Dev. 2019, 180, 42–48. [Google Scholar] [CrossRef]
- Mitchell, S.J.; Bernier, M.; Aon, M.A.; Cortassa, S.; Kim, E.Y.; Fang, E.F.; Palacios, H.H.; Ali, A.; Navas-Enamorado, I.; Di Francesco, A.; et al. Nicotinamide Improves Aspects of Healthspan, but Not Lifespan, in Mice. Cell Metab. 2018, 27, 667–676.e4. [Google Scholar] [CrossRef]
- Qi, Z.; Xia, J.; Xue, X.; He, Q.; Ji, L.; Ding, S. Long-Term Treatment with Nicotinamide Induces Glucose Intolerance and Skeletal Muscle Lipotoxicity in Normal Chow-Fed Mice: Compared to Diet-Induced Obesity. J. Nutr. Biochem. 2016, 36, 31–41. [Google Scholar] [CrossRef]
- Salehi, B.; Zucca, P.; Orhan, I.E.; Azzini, E.; Adetunji, C.O.; Mohammed, S.A.; Banerjee, S.K.; Sharopov, F.; Rigano, D.; Sharifi-Rad, J.; et al. Allicin and Health: A Comprehensive Review. Trends Food Sci. Technol. 2019, 86, 502–516. [Google Scholar] [CrossRef]
- Moriguchi, T.; Saito, H.; Nishiyama, N. Aged Garlic Extract Prolongs Longevity and Improves Spatial Memory Deficit in Senescence-Accelerated Mouse. Biol. Pharm. Bull. 1996, 19, 305–307. [Google Scholar] [CrossRef]
- Fujisawa, H.; Suma, K.; Origuchi, K.; Kumagai, H.; Seki, T.; Ariga, T. Biological and Chemical Stability of Garlic-Derived Allicin. J. Agric. Food Chem. 2008, 56, 4229–4235. [Google Scholar] [CrossRef] [PubMed]
- Alves, A.; Bassot, A.; Bulteau, A.-L.; Pirola, L.; Morio, B. Glycine Metabolism and Its Alterations in Obesity and Metabolic Diseases. Nutrients 2019, 11, 1356. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Sun, L.; Zhang, W.; Li, H.; Wang, S.; Mu, H.; Zhou, Q.; Zhang, Y.; Tang, Y.; Wang, Y.; et al. Association of Serum Glycine Levels with Metabolic Syndrome in an Elderly Chinese Population. Nutr. Metab. 2018, 15, 89. [Google Scholar] [CrossRef] [PubMed]
- Kouchiwa, T.; Wada, K.; Uchiyama, M.; Kasezawa, N.; Niisato, M.; Murakami, H.; Fukuyama, K.; Yokogoshi, H. Age-Related Changes in Serum Amino Acids Concentrations in Healthy Individuals. Clin. Chem. Lab. Med. 2012, 50, 861–870. [Google Scholar] [CrossRef]
- Liu, Y.J.; Janssens, G.E.; McIntyre, R.L.; Molenaars, M.; Kamble, R.; Gao, A.W.; Jongejan, A.; van Weeghel, M.; MacInnes, A.W.; Houtkooper, R.H. Glycine Promotes Longevity in Caenorhabditis Elegans in a Methionine Cycle-Dependent Fashion. PLoS Genet. 2019, 15, e1007633. [Google Scholar] [CrossRef]
- Brind, J.; Malloy, V.; Augie, I.; Caliendo, N.; Vogelman, J.H.; Zimmerman, J.A.; Orentreich, N. Dietary Glycine Supplementation Mimics Lifespan Extension by Dietary Methionine Restriction in Fisher 344 Rats. FASEB J. 2011, 25, 528.2. [Google Scholar] [CrossRef]
- Miller, R.A.; Harrison, D.E.; Astle, C.M.; Bogue, M.A.; Brind, J.; Fernandez, E.; Flurkey, K.; Javors, M.; Ladiges, W.; Leeuwenburgh, C.; et al. Glycine Supplementation Extends Lifespan of Male and Female Mice. Aging Cell 2019, 18, e12953. [Google Scholar] [CrossRef]
- Kumar, P.; Osahon, O.W.; Sekhar, R.V. GlyNAC (Glycine and N-Acetylcysteine) Supplementation in Mice Increases Length of Life by Correcting Glutathione Deficiency, Oxidative Stress, Mitochondrial Dysfunction, Abnormalities in Mitophagy and Nutrient Sensing, and Genomic Damage. Nutrients 2022, 14, 1114. [Google Scholar] [CrossRef]
- Šalamon, Š.; Kramar, B.; Marolt, T.P.; Poljšak, B.; Milisav, I. Medical and Dietary Uses of N-Acetylcysteine. Antioxidants 2019, 8, 111. [Google Scholar] [CrossRef]
- Ershad, M.; Naji, A.; Vearrier, D. N Acetylcysteine. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar]
- Shaposhnikov, M.V.; Zemskaya, N.V.; Koval, L.A.; Schegoleva, E.V.; Zhavoronkov, A.; Moskalev, A.A. Effects of N-Acetyl-L-Cysteine on Lifespan, Locomotor Activity and Stress-Resistance of 3 Drosophila Species with Different Lifespans. Aging (Albany NY) 2018, 10, 2428–2458. [Google Scholar] [CrossRef]
- Oh, S.-I.; Park, J.-K.; Park, S.-K. Lifespan Extension and Increased Resistance to Environmental Stressors by N-Acetyl-L-Cysteine in Caenorhabditis Elegans. Clinics 2015, 70, 380–386. [Google Scholar] [CrossRef]
- Flurkey, K.; Astle, C.M.; Harrison, D.E. Life Extension by Diet Restriction and N-Acetyl-L-Cysteine in Genetically Heterogeneous Mice. J. Gerontol. A Biol. Sci. Med. Sci. 2010, 65A, 1275–1284. [Google Scholar] [CrossRef]
- Harrison, A.P.; Pierzynowski, S.G. Biological Effects of 2-Oxoglutarate with Particular Emphasis on the Regulation of Protein, Mineral and Lipid Absorption/Metabolism, Muscle Performance, Kidney Function, Bone Formation and Cancerogenesis, All Viewed from a Healthy Ageing Perspective State of the Art--Review Article. J. Physiol. Pharmacol. 2008, 59 (Suppl. S1), 91–106. [Google Scholar] [PubMed]
- Chin, R.M.; Fu, X.; Pai, M.Y.; Vergnes, L.; Hwang, H.; Deng, G.; Diep, S.; Lomenick, B.; Meli, V.S.; Monsalve, G.C.; et al. The Metabolite α-Ketoglutarate Extends Lifespan by Inhibiting ATP Synthase and TOR. Nature 2014, 510, 397–401. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.; Wang, T.; Wu, N.; Li, D.; Fan, X.; Xu, Z.; Mishra, S.K.; Yang, M. Alpha-Ketoglutarate Extends Drosophila Lifespan by Inhibiting MTOR and Activating AMPK. Aging 2019, 11, 4183–4197. [Google Scholar] [CrossRef]
- Heitman, J.; Movva, N.R.; Hall, M.N. Targets for Cell Cycle Arrest by the Immunosuppressant Rapamycin in Yeast. Science 1991, 253, 905–909. [Google Scholar] [CrossRef]
- Shindyapina, A.V.; Cho, Y.; Kaya, A.; Tyshkovskiy, A.; Castro, J.P.; Gordevicius, J.; Poganik, J.R.; Horvath, S.; Peshkin, L.; Gladyshev, V.N. Rapamycin Treatment during Development Extends Lifespan and Healthspan. bioRxiv 2022. [Google Scholar] [CrossRef]
- Asadi Shahmirzadi, A.; Edgar, D.; Liao, C.-Y.; Hsu, Y.-M.; Lucanic, M.; Asadi Shahmirzadi, A.; Wiley, C.D.; Gan, G.; Kim, D.E.; Kasler, H.G.; et al. Alpha-Ketoglutarate, an Endogenous Metabolite, Extends Lifespan and Compresses Morbidity in Aging Mice. Cell Metab. 2020, 32, 447–456.e6. [Google Scholar] [CrossRef]
- Rayman, M.P. Selenium and Human Health. Lancet 2012, 379, 1256–1268. [Google Scholar] [CrossRef]
- Cai, Z.; Zhang, J.; Li, H. Selenium, Aging and Aging-Related Diseases. Aging Clin. Exp. Res. 2019, 31, 1035–1047. [Google Scholar] [CrossRef]
- Hao, Z.; Liu, Y.; Li, Y.; Song, W.; Yu, J.; Li, H.; Wang, W. Association between Longevity and Element Levels in Food and Drinking Water of Typical Chinese Longevity Area. J. Nutr. Health Aging 2016, 20, 897–903. [Google Scholar] [CrossRef] [PubMed]
- Mickiewicz, B.; Villemaire, M.L.; Sandercock, L.E.; Jirik, F.R.; Vogel, H.J. Metabolic Changes Associated with Selenium Deficiency in Mice. Biometals 2014, 27, 1137–1147. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Qazi, I.H.; Pan, B.; Angel, C.; Guo, S.; Yang, J.; Zhang, Y.; Ming, Z.; Zeng, C.; Meng, Q.; et al. Dietary Selenium Supplementation Ameliorates Female Reproductive Efficiency in Aging Mice. Antioxidants 2019, 8, 634. [Google Scholar] [CrossRef]
- Fodor, J.; Al-Gaadi, D.; Czirják, T.; Oláh, T.; Dienes, B.; Csernoch, L.; Szentesi, P. Improved Calcium Homeostasis and Force by Selenium Treatment and Training in Aged Mouse Skeletal Muscle. Sci. Rep. 2020, 10, 1707. [Google Scholar] [CrossRef]
- Plummer, J.D.; Postnikoff, S.D.; Tyler, J.K.; Johnson, J.E. Selenium Supplementation Inhibits IGF-1 Signaling and Confers Methionine Restriction-like Healthspan Benefits to Mice. Elife 2021, 10, e62483. [Google Scholar] [CrossRef]
- Leiter, O.; Zhuo, Z.; Rust, R.; Wasielewska, J.M.; Grönnert, L.; Kowal, S.; Overall, R.W.; Adusumilli, V.S.; Blackmore, D.G.; Southon, A.; et al. Selenium Mediates Exercise-Induced Adult Neurogenesis and Reverses Learning Deficits Induced by Hippocampal Injury and Aging. Cell Metabolism. 2022, 34, 408–423.e8. [Google Scholar] [CrossRef]
- Van Dijk, M.; Dijk, F.J.; Hartog, A.; van Norren, K.; Verlaan, S.; van Helvoort, A.; Jaspers, R.T.; Luiking, Y. Reduced Dietary Intake of Micronutrients with Antioxidant Properties Negatively Impacts Muscle Health in Aged Mice: Reduced Dietary Intake of Micronutrients. J. Cachexia Sarcopenia Muscle 2018, 9, 146–159. [Google Scholar] [CrossRef] [PubMed]
- Kalantari, H.; Das, D.K. Physiological Effects of Resveratrol. BioFactors 2010, 36, 401–406. [Google Scholar] [CrossRef]
- Jang, M.; Cai, L.; Udeani, G.O.; Slowing, K.V.; Thomas, C.F.; Beecher, C.W.W.; Fong, H.H.S.; Farnsworth, N.R.; Kinghorn, A.D.; Mehta, R.G.; et al. Cancer Chemopreventive Activity of Resveratrol, a Natural Product Derived from Grapes. Science 1997, 275, 218–220. [Google Scholar] [CrossRef]
- Howitz, K.T.; Bitterman, K.J.; Cohen, H.Y.; Lamming, D.W.; Lavu, S.; Wood, J.G.; Zipkin, R.E.; Chung, P.; Kisielewski, A.; Zhang, L.-L.; et al. Small Molecule Activators of Sirtuins Extend Saccharomyces Cerevisiae Lifespan. Nature 2003, 425, 191–196. [Google Scholar] [CrossRef]
- Pallauf, K.; Rimbach, G.; Maria Rupp, P.; Chin, D.; Wolf, I. Resveratrol and Lifespan in Model Organisms. Curr. Med. Chem. 2016, 23, 4639–4680. [Google Scholar] [CrossRef] [PubMed]
- Cantó, C.; Gerhart-Hines, Z.; Feige, J.N.; Lagouge, M.; Noriega, L.; Milne, J.C.; Elliott, P.J.; Puigserver, P.; Auwerx, J. AMPK Regulates Energy Expenditure by Modulating NAD+ Metabolism and SIRT1 Activity. Nature 2009, 458, 1056–1060. [Google Scholar] [CrossRef] [PubMed]
- Lai, X.; Cao, M.; Song, X.; Jia, R.; Zou, Y.; Li, L.; Liang, X.; He, C.; Yin, L.; Yue, G.; et al. Resveratrol Promotes Recovery of Immune Function of Immunosuppressive Mice by Activating JNK/NF-ΚB Pathway in Splenic Lymphocytes. Can. J. Physiol. Pharmacol. 2017, 95, 763–767. [Google Scholar] [CrossRef] [PubMed]
- Sung, M.M.; Das, S.K.; Levasseur, J.; Byrne, N.J.; Fung, D.; Kim, T.T.; Masson, G.; Boisvenue, J.; Soltys, C.-L.; Oudit, G.Y.; et al. Resveratrol Treatment of Mice With Pressure-Overload–Induced Heart Failure Improves Diastolic Function and Cardiac Energy Metabolism. Circ. Heart Fail. 2015, 8, 128–137. [Google Scholar] [CrossRef]
- Vilar-Pereira, G.; Carneiro, V.C.; Mata-Santos, H.; Vicentino, A.R.R.; Ramos, I.P.; Giarola, N.L.L.; Feijó, D.F.; Meyer-Fernandes, J.R.; Paula-Neto, H.A.; Medei, E.; et al. Resveratrol Reverses Functional Chagas Heart Disease in Mice. PLoS Pathog. 2016, 12, e1005947. [Google Scholar] [CrossRef]
- Saud, S.M.; Li, W.; Morris, N.L.; Matter, M.S.; Colburn, N.H.; Kim, Y.S.; Young, M.R. Resveratrol Prevents Tumorigenesis in Mouse Model of Kras Activated Sporadic Colorectal Cancer by Suppressing Oncogenic Kras Expression. Carcinogenesis 2014, 35, 2778–2786. [Google Scholar] [CrossRef]
- Kane, A.E.; Hilmer, S.N.; Boyer, D.; Gavin, K.; Nines, D.; Howlett, S.E.; de Cabo, R.; Mitchell, S.J. Impact of Longevity Interventions on a Validated Mouse Clinical Frailty Index. J. Gerontol. A Biol. Sci. Med. Sci. 2016, 71, 333–339. [Google Scholar] [CrossRef]
- Kan, N.-W.; Ho, C.-S.; Chiu, Y.-S.; Huang, W.-C.; Chen, P.-Y.; Tung, Y.-T.; Huang, C.-C. Effects of Resveratrol Supplementation and Exercise Training on Exercise Performance in Middle-Aged Mice. Molecules 2016, 21, 661. [Google Scholar] [CrossRef]
- Rodríguez-Bies, E.; Tung, B.T.; Navas, P.; López-Lluch, G. Resveratrol Primes the Effects of Physical Activity in Old Mice. Br. J. Nutr. 2016, 116, 979–988. [Google Scholar] [CrossRef]
- Muhammad, M.H.; Allam, M.M. Resveratrol and/or Exercise Training Counteract Aging-Associated Decline of Physical Endurance in Aged Mice; Targeting Mitochondrial Biogenesis and Function. J. Physiol. Sci. 2018, 68, 681–688. [Google Scholar] [CrossRef]
- Lee, B.C.; Kaya, A.; Gladyshev, V.N. Methionine Restriction and Life-Span Control. Ann. N. Y. Acad. Sci. 2016, 1363, 116–124. [Google Scholar] [CrossRef] [PubMed]
- Orentreich, N.; Matias, J.R.; DeFelice, A.; Zimmerman, J.A. Low Methionine Ingestion by Rats Extends Life Span. J. Nutr. 1993, 123, 269–274. [Google Scholar] [CrossRef]
- Plummer, J.D.; Johnson, J.E. Extension of Cellular Lifespan by Methionine Restriction Involves Alterations in Central Carbon Metabolism and Is Mitophagy-Dependent. Front. Cell Dev. Biol. 2019, 7, M146–M157. [Google Scholar] [CrossRef] [PubMed]
- Bin, P.; Zhu, C.; Liu, S.; Li, Z.; Ren, W.; Zhu, G. Perspective: Methionine Restriction–Induced Longevity—A Possible Role for Inhibiting the Synthesis of Bacterial Quorum Sensing Molecules. Adv. Nutr. 2020, 11, 773–783. [Google Scholar] [CrossRef] [PubMed]
- Bárcena, C.; Quirós, P.M.; Durand, S.; Mayoral, P.; Rodríguez, F.; Caravia, X.M.; Mariño, G.; Garabaya, C.; Fernández-García, M.T.; Kroemer, G.; et al. Methionine Restriction Extends Lifespan in Progeroid Mice and Alters Lipid and Bile Acid Metabolism. Cell Rep. 2018, 24, 2392–2403. [Google Scholar] [CrossRef]
- Osorio, F.G.; Navarro, C.L.; Cadiñanos, J.; López-Mejía, I.C.; Quirós, P.M.; Bartoli, C.; Rivera, J.; Tazi, J.; Guzmán, G.; Varela, I.; et al. Splicing-Directed Therapy in a New Mouse Model of Human Accelerated Aging. Sci. Transl. Med. 2011, 3, 106ra107. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, S.; MacArthur, M.; Kane, A.; Torrence, M.; Mehmet, H.; Vath, J.; Manning, B.; Mitchell, J. Late-Onset Pharmacological or Dietary Interventions Improve Healthspan and Lifespan in Male and Female Mice. Innov. Aging 2020, 4 (Suppl. S1), 125. [Google Scholar] [CrossRef]
- Schultz, M.B.; Kane, A.E.; Mitchell, S.J.; MacArthur, M.R.; Warner, E.; Vogel, D.S.; Mitchell, J.R.; Howlett, S.E.; Bonkowski, M.S.; Sinclair, D.A. Age and Life Expectancy Clocks Based on Machine Learning Analysis of Mouse Frailty. Nat. Commun. 2020, 11, 4618. [Google Scholar] [CrossRef]
- Levine, M.E.; Suarez, J.A.; Brandhorst, S.; Balasubramanian, P.; Cheng, C.-W.; Madia, F.; Fontana, L.; Mirisola, M.G.; Guevara-Aguirre, J.; Wan, J.; et al. Low Protein Intake Is Associated with a Major Reduction in IGF-1, Cancer, and Overall Mortality in the 65 and Younger but Not Older Population. Cell Metab. 2014, 19, 407–417. [Google Scholar] [CrossRef]
- Anderson, J.G.; Hintze, K.; Marchant, E.D. Restricting Branched-chain Amino Acids: An Approach to Improve Metabolic Health. J. Physiol. 2018, 596, 2469–2470. [Google Scholar] [CrossRef]
- McGarrah, R.W.; Zhang, G.-F.; Christopher, B.A.; Deleye, Y.; Walejko, J.M.; Page, S.; Ilkayeva, O.; White, P.J.; Newgard, C.B. Dietary Branched-Chain Amino Acid Restriction Alters Fuel Selection and Reduces Triglyceride Stores in Hearts of Zucker Fatty Rats. Am. J. Physiol. Endocrinol. Metab. 2020, 318, E216–E223. [Google Scholar] [CrossRef] [PubMed]
- Richardson, N.E.; Konon, E.N.; Schuster, H.S.; Mitchell, A.T.; Boyle, C.; Rodgers, A.C.; Finke, M.; Haider, L.R.; Yu, D.; Flores, V.; et al. Lifelong Restriction of Dietary Branched-Chain Amino Acids Has Sex-Specific Benefits for Frailty and Life Span in Mice. Nat. Aging 2021, 1, 73–86. [Google Scholar] [CrossRef]
- Lorenzo-López, L.; Maseda, A.; de Labra, C.; Regueiro-Folgueira, L.; Rodríguez-Villamil, J.L.; Millán-Calenti, J.C. Nutritional Determinants of Frailty in Older Adults: A Systematic Review. BMC Geriatr. 2017, 17, 108. [Google Scholar] [CrossRef] [PubMed]
- Coelho-Júnior, H.; Rodrigues, B.; Uchida, M.; Marzetti, E. Low Protein Intake Is Associated with Frailty in Older Adults: A Systematic Review and Meta-Analysis of Observational Studies. Nutrients 2018, 10, 1334. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bisset, E.S.; Howlett, S.E. The Use of Dietary Supplements and Amino Acid Restriction Interventions to Reduce Frailty in Pre-Clinical Models. Nutrients 2022, 14, 2806. https://doi.org/10.3390/nu14142806
Bisset ES, Howlett SE. The Use of Dietary Supplements and Amino Acid Restriction Interventions to Reduce Frailty in Pre-Clinical Models. Nutrients. 2022; 14(14):2806. https://doi.org/10.3390/nu14142806
Chicago/Turabian StyleBisset, Elise S., and Susan E. Howlett. 2022. "The Use of Dietary Supplements and Amino Acid Restriction Interventions to Reduce Frailty in Pre-Clinical Models" Nutrients 14, no. 14: 2806. https://doi.org/10.3390/nu14142806
APA StyleBisset, E. S., & Howlett, S. E. (2022). The Use of Dietary Supplements and Amino Acid Restriction Interventions to Reduce Frailty in Pre-Clinical Models. Nutrients, 14(14), 2806. https://doi.org/10.3390/nu14142806