
Lutein Can Alleviate Oxidative Stress, Inflammation, and Apoptosis Induced by Excessive Alcohol to Ameliorate Reproductive Damage in Male Rats


Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Experimental Design and Animals
2.3. Calculation of the Testis Index and Sperm Analysis
2.4. Histopathological Study of Testis Tissues
2.5. Detection of Sex Hormones and Labeling Enzymes
2.6. Oxidative Stress Levels and Inflammatory Parameters
2.7. Western Blot Analysis
2.8. TUNEL Staining
2.9. Statistical Analysis
3. Results
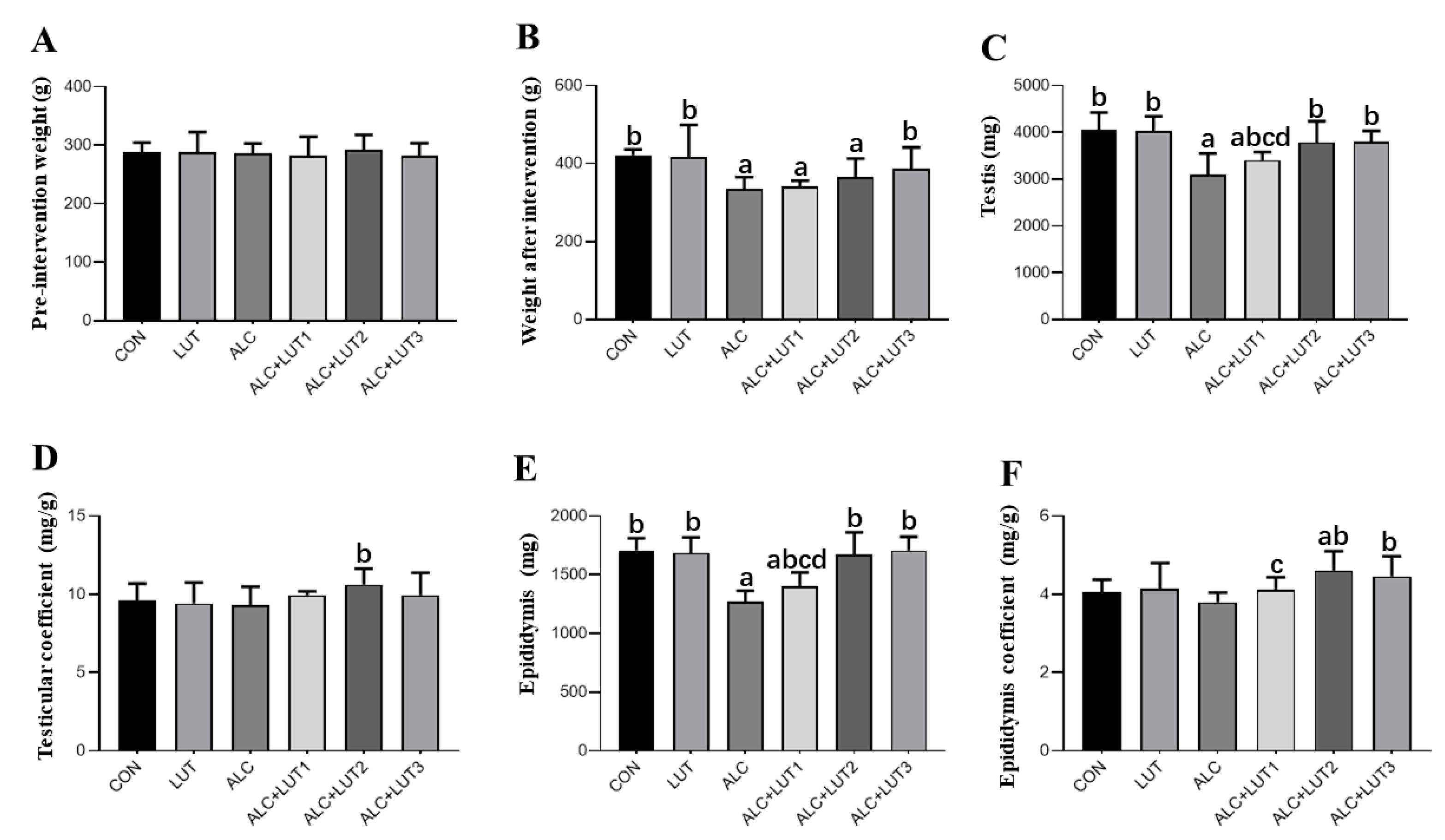
3.1. Body Weights and Organ Weights
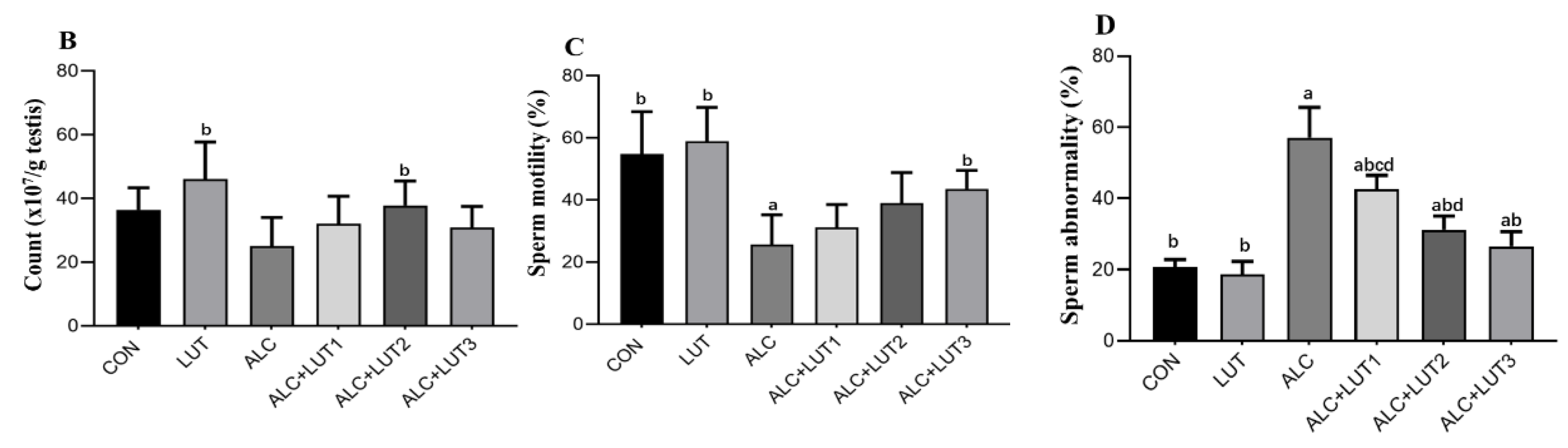
3.2. Effect of Lutein on Sperm Quality in Alcohol-Treated Rats
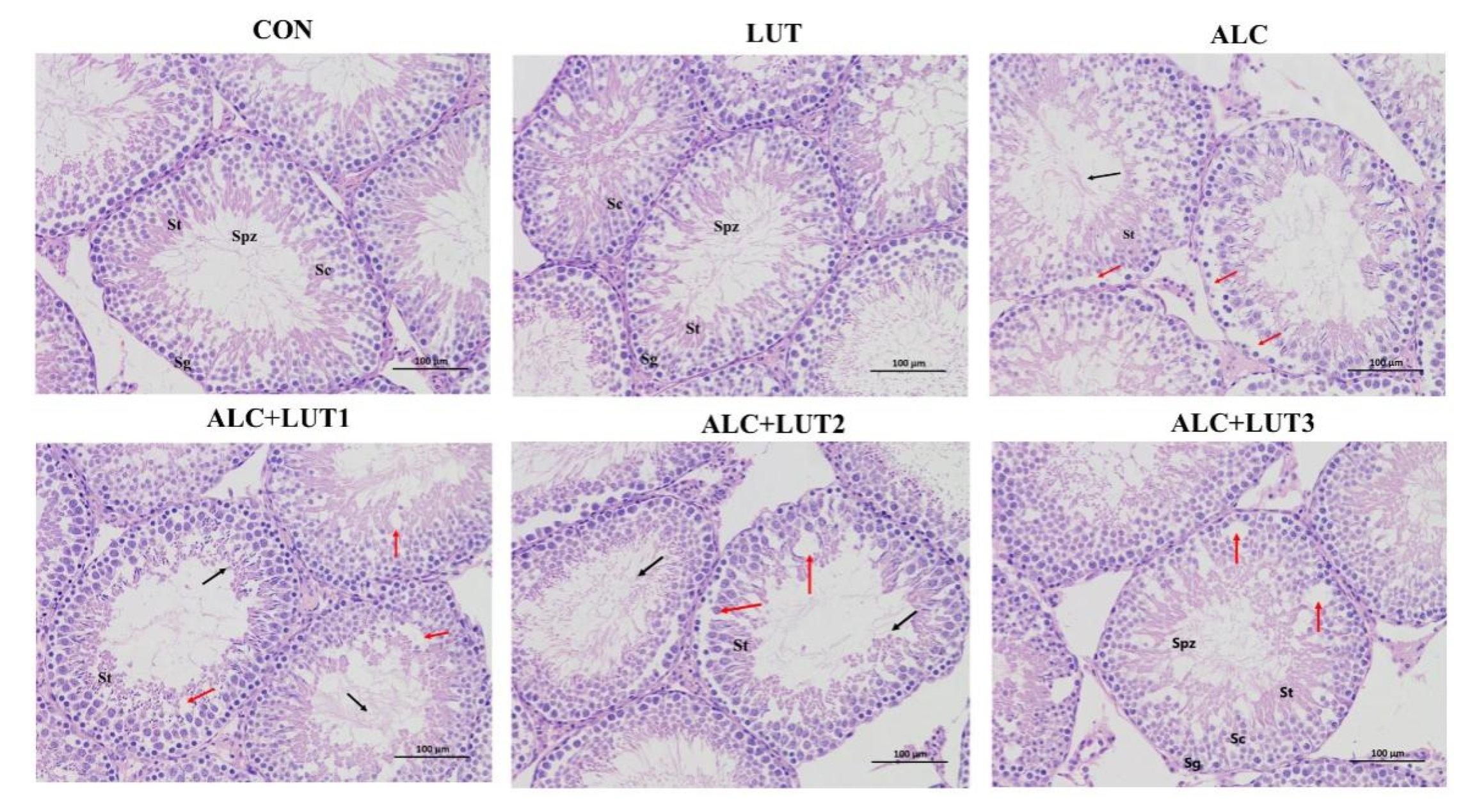
3.3. Pathological Changes of Testicular Tissue
3.4. The Levels of Marker Enzymes in Testis
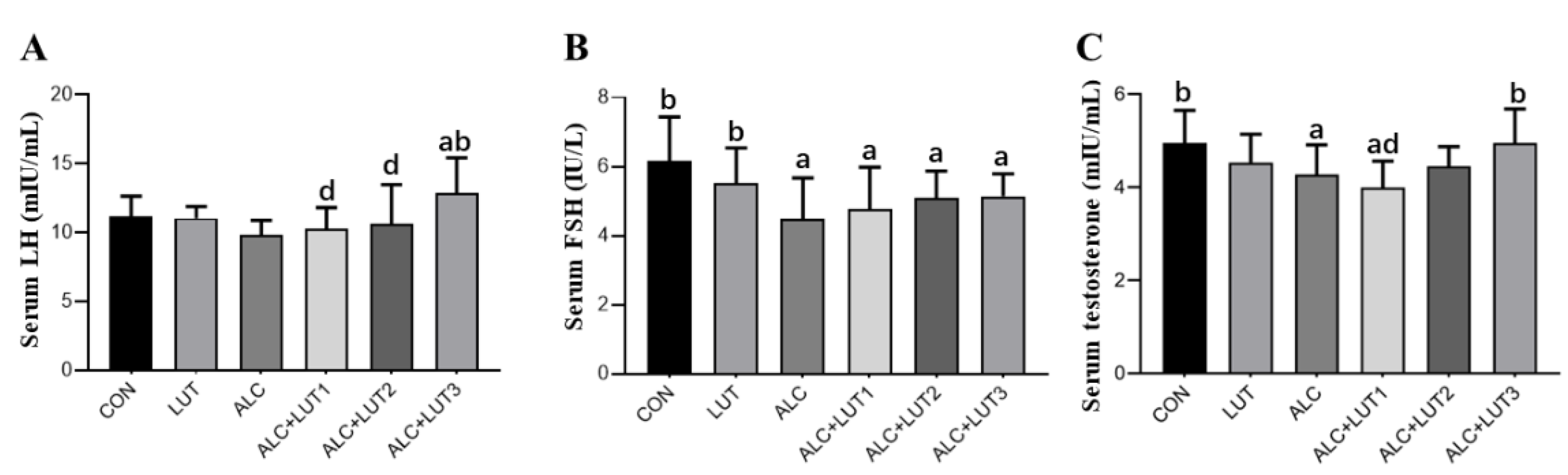
3.5. Male Reproductive Hormone Level
3.6. Changes in Oxidative Stress Levels in Testicular Tissue
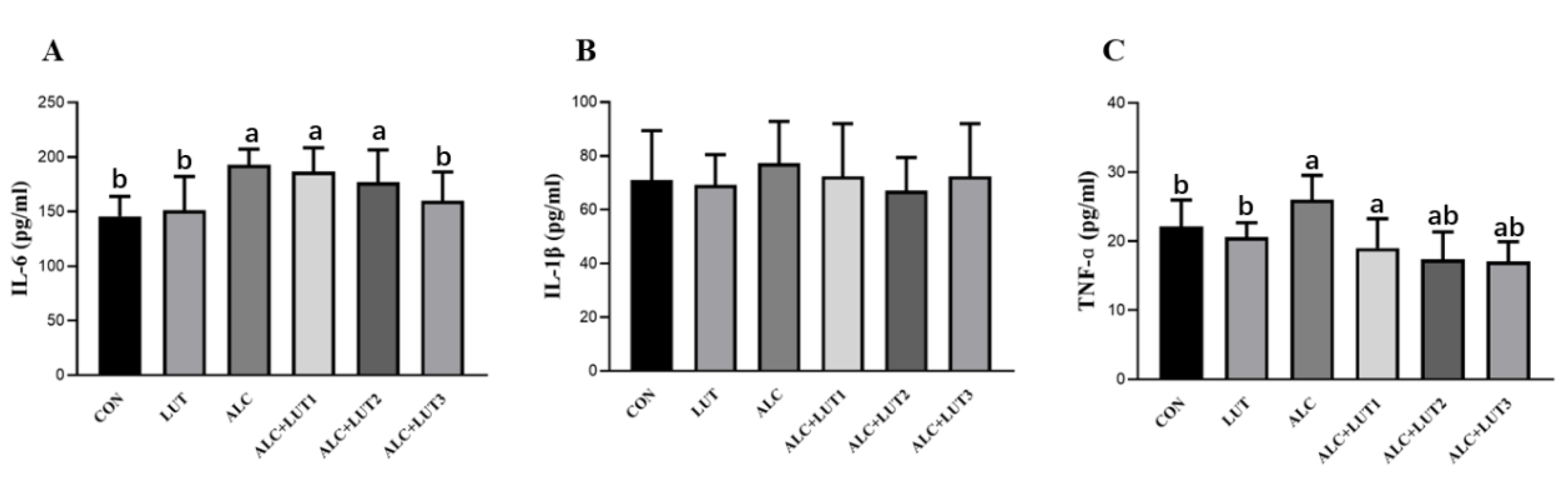
3.7. Changes in the Levels of Inflammatory Markers in Testicular Tissue
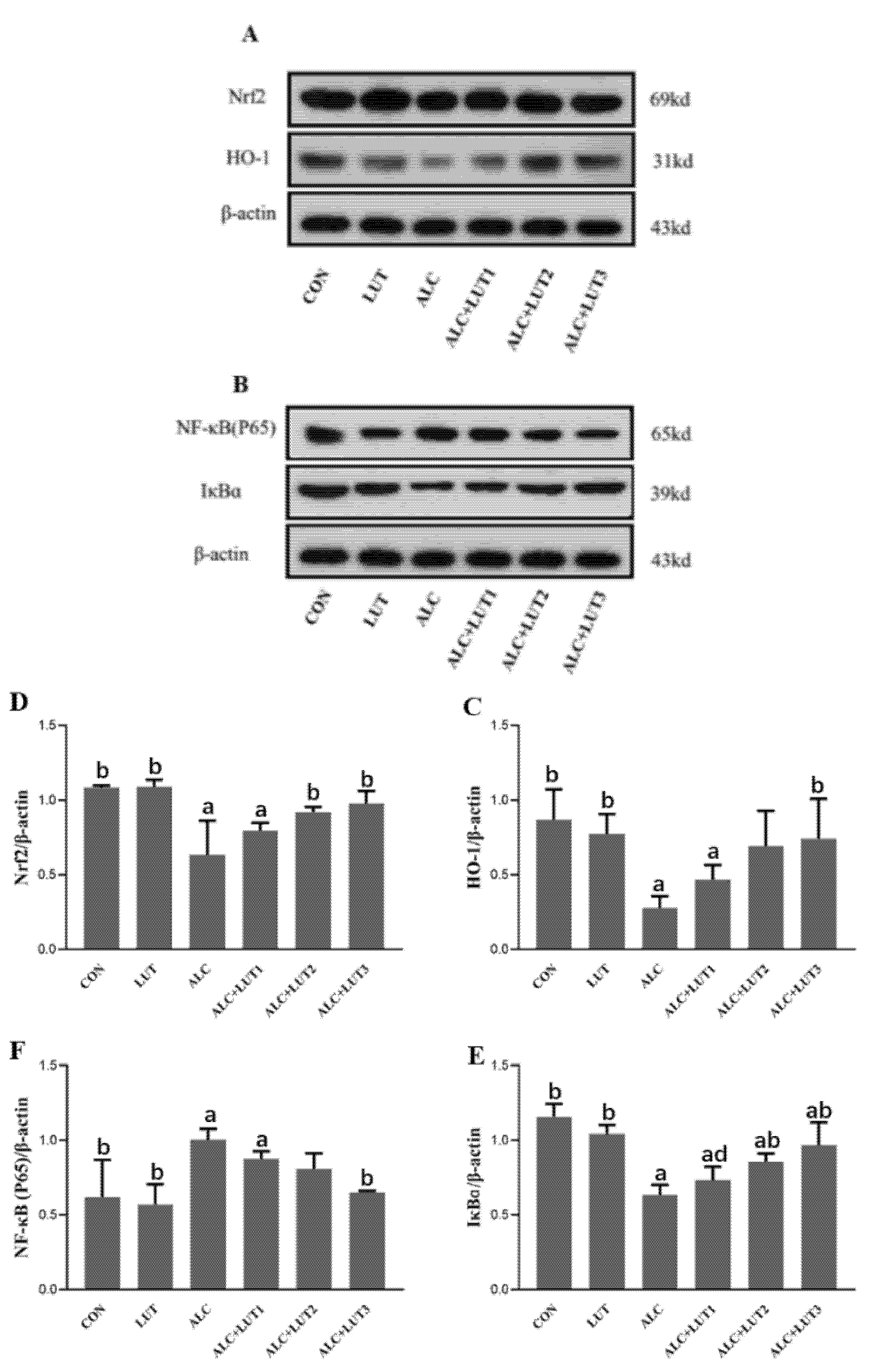
3.8. Changes of Expression of Proteins Related to NF-κB/Nrf2 Pathway
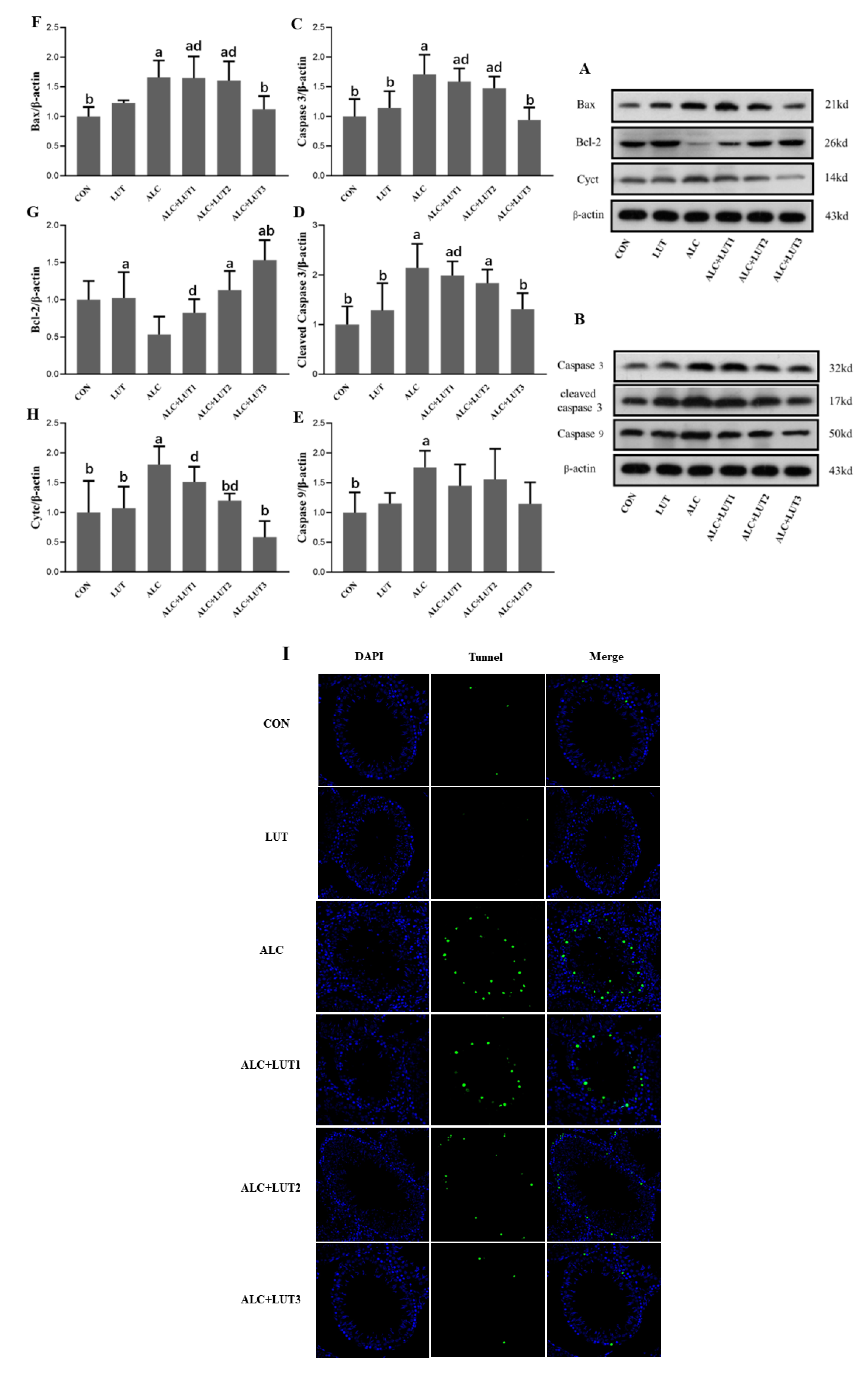
3.9. Lutein Attenuates Chronic Alcohol-Induced Testicular Germ Cell Apoptosis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Agarwal, A.; Majzoub, A.; Parekh, N.; Henkel, R. A Schematic Overview of the Current Status of Male Infertility Practice. World J. Mens. Health 2020, 38, 308–322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siervo, G.E.; Vieira, H.R.; Ogo, F.M.; Fernandez, C.D.; Gonçalves, G.D.; Mesquita, S.F.; Anselmo-Franci, J.A.; Cecchini, R.; Guarnier, F.A.; Fernandes, G.S. Spermatic and testicular damages in rats exposed to ethanol: Influence of lipid peroxidation but not testosterone. Toxicology 2015, 330, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Emanuele, M.A.; Emanuele, N. Alcohol and the male reproductive system. Alcohol Res. Health 2001, 25, 282–287. [Google Scholar] [PubMed]
- Radhakrishnakartha, H.; Appu, A.P.; Indira, M. Ascorbic acid supplementation enhances recovery from ethanol induced inhibition of Leydig cell steroidogenesis than abstention in male guinea pigs. Eur. J. Pharmacol. 2014, 723, 73–79. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Su, P. The role of the Fas/FasL signaling pathway in environmental toxicant-induced testicular cell apoptosis: An update. Syst. Biol. Reprod. Med. 2018, 64, 93–102. [Google Scholar] [CrossRef] [PubMed]
- Adachi, Y.; Moore, L.E.; Bradford, B.U.; Gao, W.; Thurman, R.G. Antibiotics prevent liver injury in rats following long-term exposure to ethanol. Gastroenterology 1995, 108, 218–224. [Google Scholar] [CrossRef]
- Ranard, K.M.; Jeon, S.; Mohn, E.S.; Griffiths, J.C.; Johnson, E.J.; Erdman, J.W., Jr. Dietary guidance for lutein: Consideration for intake recommendations is scientifically supported. Eur. J. Nutr. 2017, 56 (Suppl. 3), 37–42. [Google Scholar] [CrossRef] [Green Version]
- Diamanti-Kandarakis, E.; Bourguignon, J.P.; Giudice, L.C.; Hauser, R.; Prins, G.S.; Soto, A.M.; Zoeller, R.T.; Gore, A.C. Endocrine-disrupting chemicals: An Endocrine Society scientific statement. Endocr. Rev. 2009, 30, 293–342. [Google Scholar] [CrossRef]
- DeBerardinis, R.J.; Lum, J.J.; Hatzivassiliou, G.; Thompson, C.B. The biology of cancer: Metabolic reprogramming fuels cell growth and proliferation. Cell Metab. 2008, 7, 11–20. [Google Scholar] [CrossRef] [Green Version]
- Sindhu, E.R.; Firdous, A.P.; Preethi, K.C.; Kuttan, R. Carotenoid lutein protects rats from paracetamol-, carbon tetrachloride- and ethanol-induced hepatic damage. J. Pharm Pharmacol. 2010, 62, 1054–1060. [Google Scholar] [CrossRef]
- Wang, D.; Kamijima, M.; Okamura, A.; Ito, Y.; Yanagiba, Y.; Jia, X.F.; Naito, H.; Ueyama, J.; Nakajima, T. Evidence for diazinon-mediated inhibition of cis-permethrin metabolism and its effects on reproductive toxicity in adult male mice. Reprod. Toxicol. 2012, 34, 489–497. [Google Scholar] [CrossRef] [PubMed]
- Harikrishnan, R.; Abhilash, P.A.; Syam Das, S.; Prathibha, P.; Rejitha, S.; John, F.; Kavitha, S.; Indira, M. Protective effect of ascorbic acid against ethanol-induced reproductive toxicity in male guinea pigs. Br. J. Nutr. 2013, 110, 689–698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Felemban, S.G.; Aldubayan, M.A.; Alhowail, A.H.; Almami, I.S. Vitamin B17 Ameliorates Methotrexate-Induced Reproductive Toxicity, Oxidative Stress, and Testicular Injury in Male Rats. Oxid. Med. Cell Longev. 2020, 2020, 4372719. [Google Scholar] [CrossRef] [PubMed]
- Muthusami, K.R.; Chinnaswamy, P. Effect of chronic alcoholism on male fertility hormones and semen quality. Fertil Steril 2005, 84, 919–924. [Google Scholar] [CrossRef]
- Jensen, T.K.; Gottschau, M.; Madsen, J.O.; Andersson, A.M.; Lassen, T.H.; Skakkebæk, N.E.; Swan, S.H.; Priskorn, L.; Juul, A.; Jørgensen, N. Habitual alcohol consumption associated with reduced semen quality and changes in reproductive hormones; a cross-sectional study among 1221 young Danish men. BMJ Open 2014, 4, e005462. [Google Scholar] [CrossRef]
- Long, M.; Yang, S.; Dong, S.; Chen, X.; Zhang, Y.; He, J. Characterization of semen quality, testicular marker enzyme activities and gene expression changes in the blood testis barrier of Kunming mice following acute exposure to zearalenone. Environ. Sci. Pollut. Res. Int. 2017, 24, 27235–27243. [Google Scholar] [CrossRef]
- Benzoni, E.; Minervini, F.; Giannoccaro, A.; Fornelli, F.; Vigo, D.; Visconti, A. Influence of in vitro exposure to mycotoxin zearalenone and its derivatives on swine sperm quality. Reprod. Toxicol. 2008, 25, 461–467. [Google Scholar] [CrossRef]
- Pant, N.; Prasad, A.K.; Srivastava, S.C.; Shankar, R.; Srivastava, S.P. Effect of oral administration of carbofuran on male reproductive system of rat. Hum. Exp. Toxicol. 1995, 14, 889–894. [Google Scholar] [CrossRef]
- Boeira, S.P.; Funck, V.R.; Borges Filho, C.; Del’Fabbro, L.; de Gomes, M.G.; Donato, F.; Royes, L.F.; Oliveira, M.S.; Jesse, C.R.; Furian, A.F. Lycopene protects against acute zearalenone-induced oxidative, endocrine, inflammatory and reproductive damages in male mice. Chem. Biol. Interact. 2015, 230, 50–57. [Google Scholar] [CrossRef]
- Sun, X.; Sun, H.; Yu, K.; Wang, Z.; Liu, Y.; Liu, K.; Zhu, Y.; Li, Y. Aluminum Chloride Causes the Dysfunction of Testes Through Inhibiting the ATPase Enzyme Activities and Gonadotropin Receptor Expression in Rats. Biol. Trace Elem. Res. 2018, 183, 296–304. [Google Scholar] [CrossRef]
- Zeng, Q.; Yi, H.; Huang, L.; An, Q.; Wang, H. Reduced testosterone and Ddx3y expression caused by long-term exposure to arsenic and its effect on spermatogenesis in mice. Environ. Toxicol. Pharmacol. 2018, 63, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Rachdaoui, N.; Sarkar, D.K. Effects of alcohol on the endocrine system. Endocrinol. Metab. Clin. N. Am. 2013, 42, 593–615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finelli, R.; Mottola, F.; Agarwal, A. Impact of Alcohol Consumption on Male Fertility Potential: A Narrative Review. Int. J. Environ. Res. Public Health 2021, 19, 328. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, A.; Virk, G.; Ong, C.; du Plessis, S.S. Effect of oxidative stress on male reproduction. World J. Mens. Health 2014, 32, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zareba, P.; Colaci, D.S.; Afeiche, M.; Gaskins, A.J.; Jørgensen, N.; Mendiola, J.; Swan, S.H.; Chavarro, J.E. Semen quality in relation to antioxidant intake in a healthy male population. Fertil Steril 2013, 100, 1572–1579. [Google Scholar] [CrossRef] [Green Version]
- Ge, N.; Liang, H.; Liu, Y.; Ma, A.G.; Han, L. Protective effect of Aplysin on hepatic injury in ethanol-treated rats. Food Chem. Toxicol. 2013, 62, 361–372. [Google Scholar] [CrossRef]
- Ballway, J.W.; Song, B.J. Translational Approaches with Antioxidant Phytochemicals against Alcohol-Mediated Oxidative Stress, Gut Dysbiosis, Intestinal Barrier Dysfunction, and Fatty Liver Disease. Antioxidants 2021, 10, 384. [Google Scholar] [CrossRef]
- Aladaileh, S.H.; Hussein, O.E.; Abukhalil, M.H.; Saghir, S.A.M.; Bin-Jumah, M.; Alfwuaires, M.A.; Germoush, M.O.; Almaiman, A.A.; Mahmoud, A.M. Formononetin Upregulates Nrf2/HO-1 Signaling and Prevents Oxidative Stress, Inflammation, and Kidney Injury in Methotrexate-Induced Rats. Antioxidants 2019, 8, 430. [Google Scholar] [CrossRef] [Green Version]
- Han, X.X.; Jiang, Y.P.; Liu, N.; Wu, J.; Yang, J.M.; Li, Y.X.; Sun, M.; Sun, T.; Zheng, P.; Jian-Qiang, Y.u. Protective effects of Astragalin on spermatogenesis in streptozotocin-induced diabetes in male mice by improving antioxidant activity and inhibiting inflammation. Biomed Pharm. 2019, 110, 561–570. [Google Scholar] [CrossRef]
- Lustig, L.; Guazzone, V.A.; Theas, M.S.; Pleuger, C.; Jacobo, P.; Pérez, C.V.; Meinhardt, A.; Fijak, M. Pathomechanisms of Autoimmune Based Testicular Inflammation. Front. Immunol. 2020, 11, 583135. [Google Scholar] [CrossRef]
- Sparacio, S.M.; Zhang, Y.; Vilcek, J.; Benveniste, E.N. Cytokine regulation of interleukin-6 gene expression in astrocytes involves activation of an NF-kappa B-like nuclear protein. J. Neuroimmunol. 1992, 39, 231–242. [Google Scholar] [CrossRef]
- Gadjeva, M.; Tomczak, M.F.; Zhang, M.; Wang, Y.Y.; Dull, K.; Rogers, A.B.; Erdman, S.E.; Fox, J.G.; Carroll, M.; Horwitz, B.H. A role for NF-kappa B subunits p50 and p65 in the inhibition of lipopolysaccharide-induced shock. J. Immunol. 2004, 173, 5786–5793. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Li, Y.; Wu, Y.; Zhang, Y.; Wang, Z.; Liu, X. Lutein suppresses inflammatory responses through Nrf2 activation and NF-κB inactivation in lipopolysaccharide-stimulated BV-2 microglia. Mol. Nutr. Food Res. 2015, 59, 1663–1673. [Google Scholar] [CrossRef] [PubMed]
- Hou, B.; Wang, F.; Liu, T.; Wang, Z. Reproductive toxicity of polystyrene microplastics: In vivo experimental study on testicular toxicity in mice. J. Hazard. Mater. 2021, 405, 124028. [Google Scholar] [CrossRef]
- Dai, C.; Li, B.; Zhou, Y.; Li, D.; Zhang, S.; Li, H.; Xiao, X.; Tang, S. Curcumin attenuates quinocetone induced apoptosis and inflammation via the opposite modulation of Nrf2/HO-1 and NF-kB pathway in human hepatocyte L02 cells. Food Chem. Toxicol. 2016, 95, 52–63. [Google Scholar] [CrossRef]
- Li, P.; Nijhawan, D.; Budihardjo, I.; Srinivasula, S.M.; Ahmad, M.; Alnemri, E.S.; Wang, X. Cytochrome c and dATP-dependent formation of Apaf-1/caspase-9 complex initiates an apoptotic protease cascade. Cell 1997, 91, 479–489. [Google Scholar] [CrossRef] [Green Version]
- El-Khadragy, M.F.; Al-Megrin, W.A.; Alomar, S.; Alkhuriji, A.F.; Metwally, D.M.; Mahgoub, S.; Amin, H.K.; Habotta, O.A.; Abdel Moneim, A.E.; Albeltagy, R.S. Chlorogenic acid abates male reproductive dysfunction in arsenic-exposed mice via attenuation of testicular oxido-inflammatory stress and apoptotic responses. Chem. Biol. Interact. 2021, 333, 109333. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | LDH (U/g Protein) | AKP (U/g Protein) | ACP (U/g Protein) | SDH (U/mg Protein) |
|---|---|---|---|---|
| CON | 396.26 ± 74.62 | 82.77 ± 20.83 b | 50.33 ± 6.65 b | 5.11 ± 0.88 b |
| LUT | 409.00 + 115.56 | 71.82 ± 19.86 | 46.08 ± 5.63 | 4.56 ± 0.88 |
| ALC | 366.91 ± 114.79 | 61.10 ± 23.12 a | 39.90 ± 6.75 a | 3.99 ± 0.65 a |
| ALC + LUT1 | 344.48 ± 155.26 | 58.37 ± 20.53 ad | 41.42 ± 4.54 a | 4.26 ± 0.61 a |
| ALC + LUT2 | 382.60 ± 176.02 | 70.51 ± 30.88 | 43.32 ± 7.41 a | 4.08 ± 0.55 a |
| ALC + LUT3 | 391.11 ± 141.63 | 86.01 ± 16.73 b | 45.32 ± 10.77 | 4.65 ± 0.38 b |
| Group | SOD (U/mg Protein) | MDA (nmol/mL) | GSH-Px (U/mg Protein) |
|---|---|---|---|
| CON | 153.58 ± 28.73 | 3.00 ± 1.35 b | 11.14 ± 1.43 b |
| LUT | 146.00 + 15.99 | 3.11 ± 0.70 b | 9.98 ± 2.48 |
| ALC | 136.66 ± 21.89 | 4.71 ± 1.85 a | 8.25 ± 2.70 a |
| ALC + LUT1 | 139.63 ± 29.63 | 3.57 ± 1.30 | 8.41 ± 1.69 a |
| ALC + LUT2 | 136.51 ± 27.34 | 3.70 ± 1.53 | 9.05 ± 2.30 a |
| ALC + LUT3 | 144.75 ± 16.81 | 2.70 ± 0.94 b | 9.73 ± 1.90 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Ding, H.; Xu, L.; Zhao, S.; Hu, S.; Ma, A.; Ma, Y. Lutein Can Alleviate Oxidative Stress, Inflammation, and Apoptosis Induced by Excessive Alcohol to Ameliorate Reproductive Damage in Male Rats. Nutrients 2022, 14, 2385. https://doi.org/10.3390/nu14122385
Zhang Y, Ding H, Xu L, Zhao S, Hu S, Ma A, Ma Y. Lutein Can Alleviate Oxidative Stress, Inflammation, and Apoptosis Induced by Excessive Alcohol to Ameliorate Reproductive Damage in Male Rats. Nutrients. 2022; 14(12):2385. https://doi.org/10.3390/nu14122385
Chicago/Turabian StyleZhang, Yebing, Haoyue Ding, Lei Xu, Suli Zhao, Shouna Hu, Aiguo Ma, and Yan Ma. 2022. "Lutein Can Alleviate Oxidative Stress, Inflammation, and Apoptosis Induced by Excessive Alcohol to Ameliorate Reproductive Damage in Male Rats" Nutrients 14, no. 12: 2385. https://doi.org/10.3390/nu14122385
APA StyleZhang, Y., Ding, H., Xu, L., Zhao, S., Hu, S., Ma, A., & Ma, Y. (2022). Lutein Can Alleviate Oxidative Stress, Inflammation, and Apoptosis Induced by Excessive Alcohol to Ameliorate Reproductive Damage in Male Rats. Nutrients, 14(12), 2385. https://doi.org/10.3390/nu14122385