
Effects of Fish Oil on Biomarkers of Axonal Injury and Inflammation in American Football Players: A Placebo-Controlled Randomized Controlled Trial

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
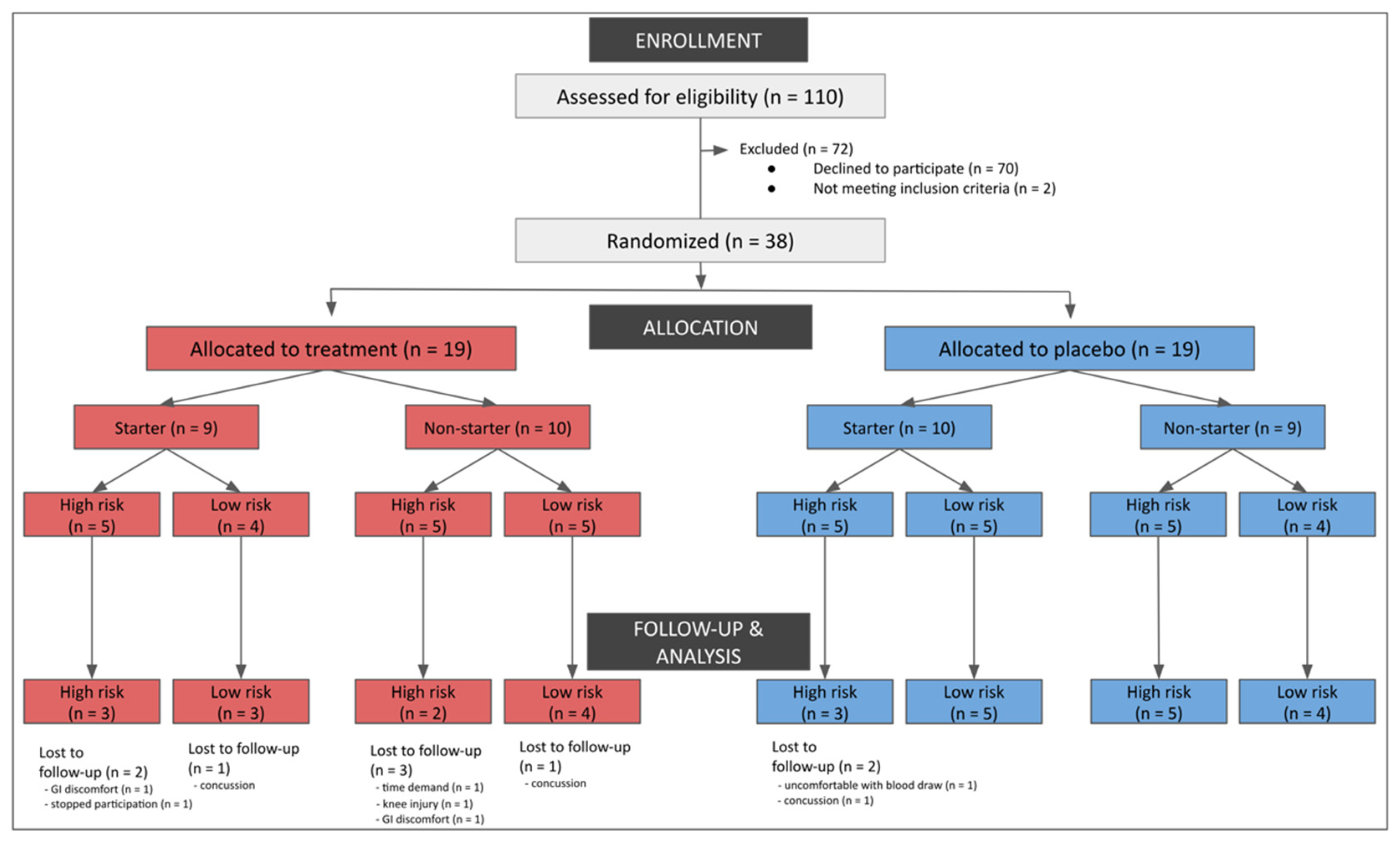
2.1. Participants
2.2. Intervention Design
2.3. Outcomes
2.4. Biospecimen Collection
2.5. Plasma Fatty Acid Characterization
2.6. Neurofilament Light
2.7. Interleukin-6 and Tumor Necrosis Factor-Alpha
2.8. Statistical Analysis
3. Results
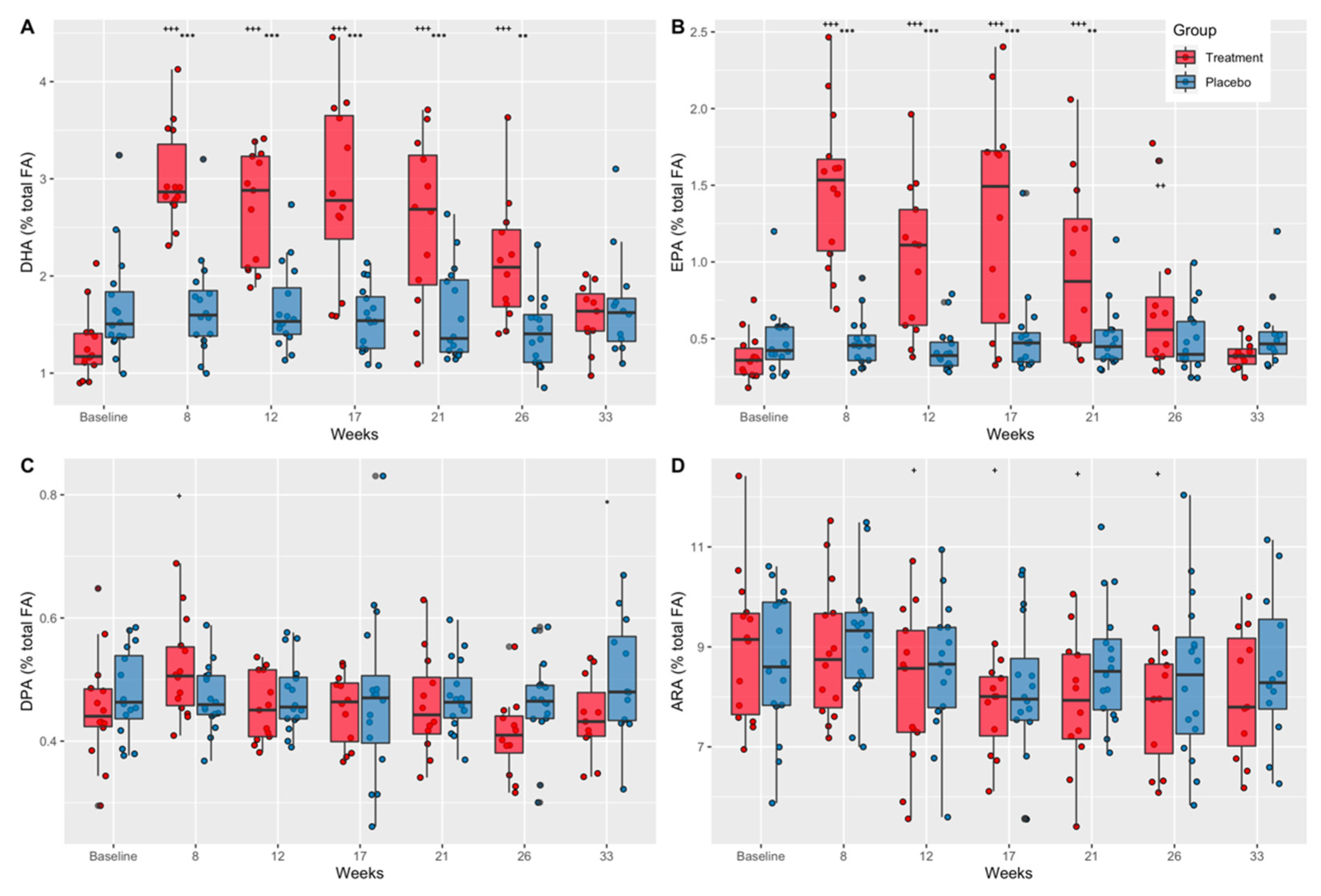
3.1. Effects of Supplementation on Plasma Fatty Acid Concentrations
3.1.1. Docosahexaenoic Acid (DHA)
3.1.2. Eicosapentaenoic Acid (EPA)
3.1.3. Docosapentaenoic Acid (DPA)
3.1.4. Arachidonic Acid (ARA)
3.1.5. ARA:DHA and ARA:EPA Ratios
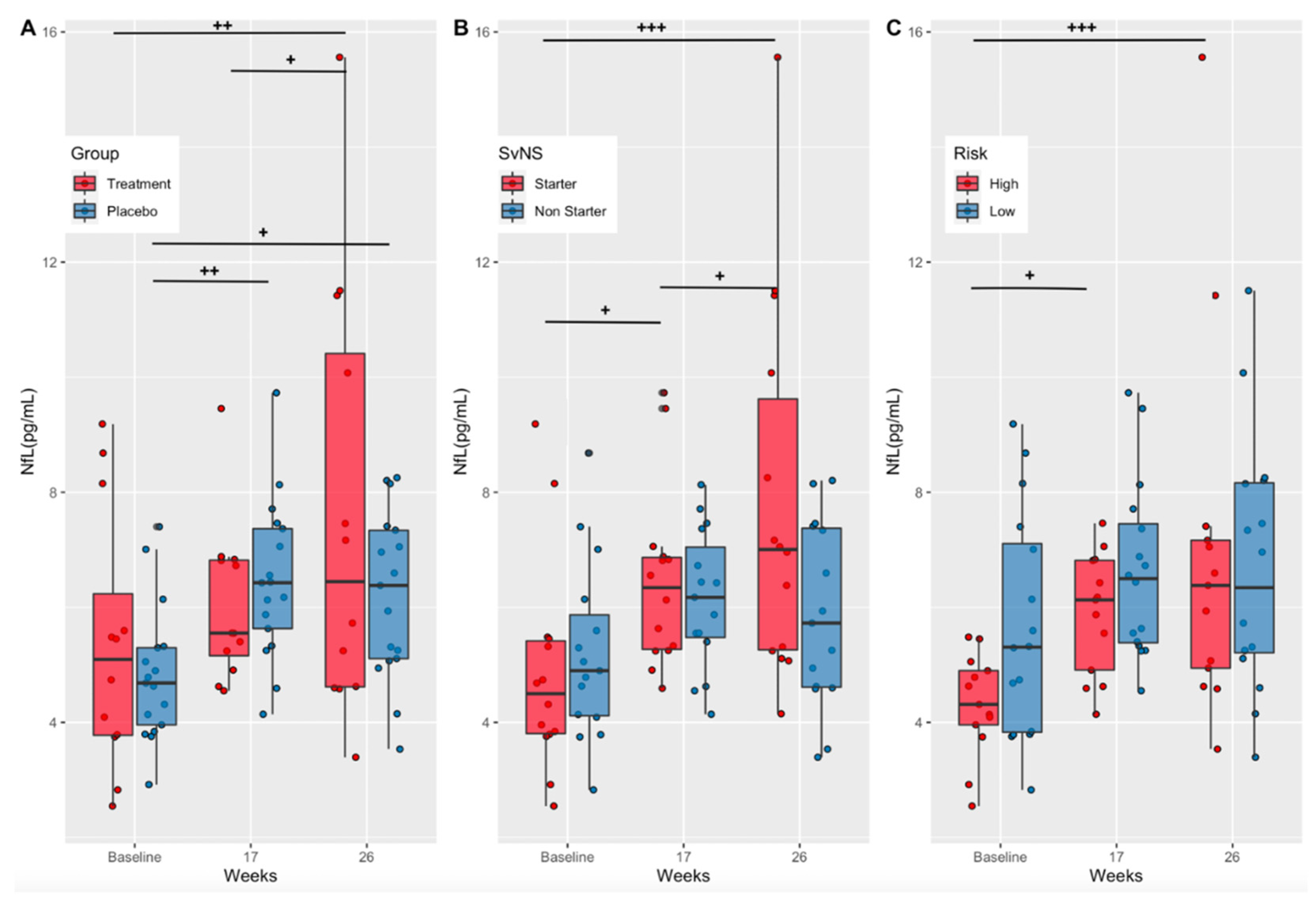
3.2. Effects of Supplementation on a Marker of Axonal Injury (Treatment vs. Placebo)
3.3. Effects of Playing Time and Position Risk on a Marker of Axonal Injury (Starters vs. Non-Starters and High-Risk vs. Low-Risk Positions)
3.4. Effects on Inflammatory Cytokines
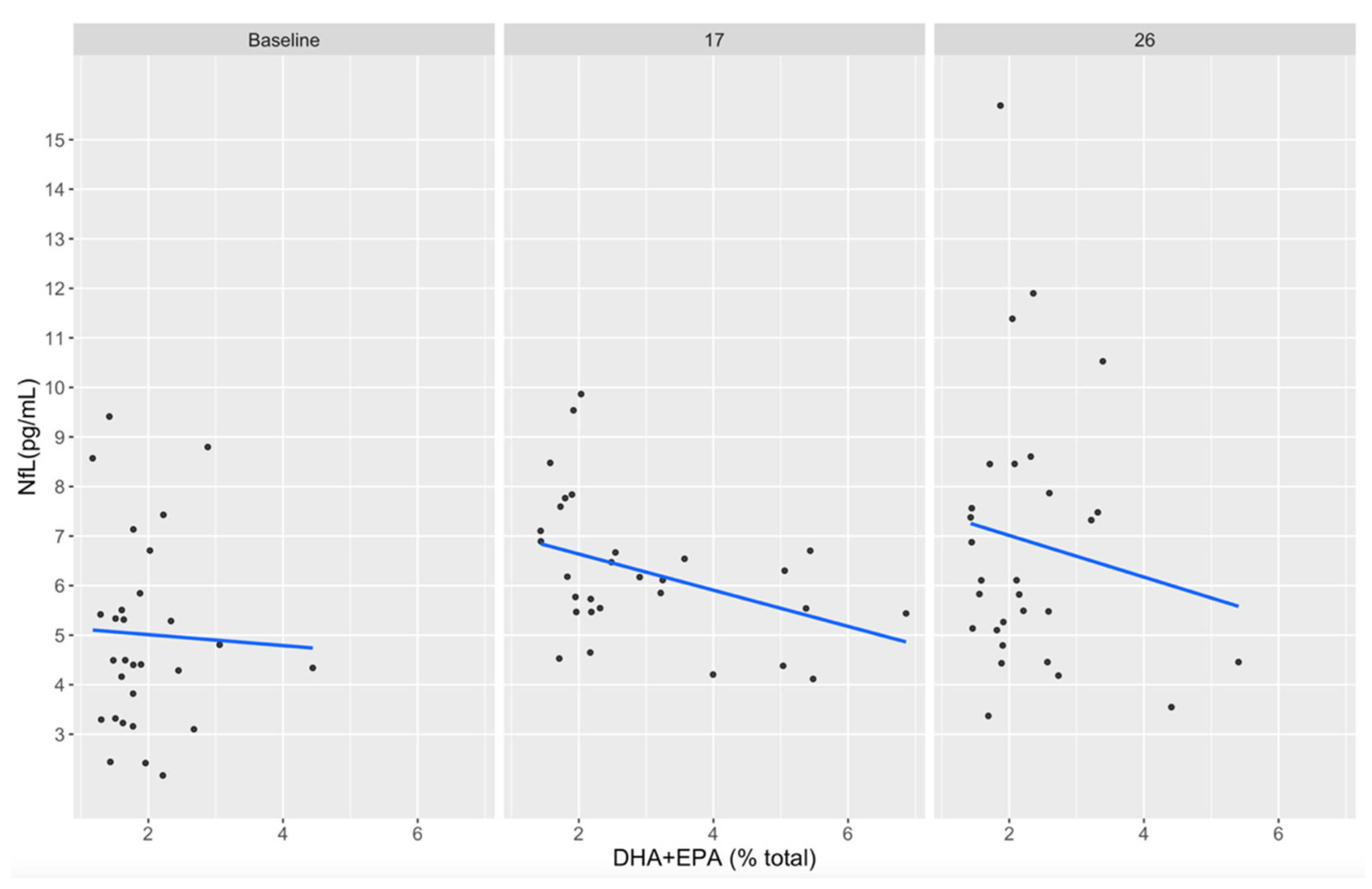
3.5. Associations between DHA+EPA and NfL Concentrations
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Crisco, J.J.; Fiore, R.; Beckwith, J.G.; Chu, J.J.; Brolinson, P.G.; Duma, S.; McAllister, T.W.; Duhaime, A.C.; Greenwald, R.M. Frequency and location of head impact exposures in individual collegiate football players. J. Athl. Train. 2010, 45, 549–559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lust, C.A.C.; Mountjoy, M.; Robinson, L.E.; Oliver, J.M.; Ma, D.W.L. Sports-related concussions and subconcussive impacts in athletes: Incidence, diagnosis, and the emerging role of EPA and DHA. Appl. Physiol. Nutr. Metab. 2020, 45, 886–892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marar, M.; McIlvain, N.M.; Fields, S.K.; Comstock, R.D. Epidemiology of concussions among United States high school athletes in 20 sports. Am. J. Sports Med. 2012, 40, 747–755. [Google Scholar] [CrossRef]
- Mez, J.; Daneshvar, D.H.; Kiernan, P.T.; Abdolmohammadi, B.; Alvarez, V.E.; Huber, B.R.; Alosco, M.L.; Solomon, T.M.; Nowinski, C.J.; McHale, L.; et al. Clinicopathological Evaluation of Chronic Traumatic Encephalopathy in Players of American Football. JAMA 2017, 318, 360–370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harmon, K.G.; Clugston, J.R.; Dec, K.; Hainline, B.; Herring, S.A.; Kane, S.; Kontos, A.P.; Leddy, J.J.; McCrea, M.A.; Poddar, S.K.; et al. American Medical Society for Sports Medicine Position Statement on Concussion in Sport. Clin. J. Sport Med. 2019, 29, 87–100. [Google Scholar] [CrossRef] [PubMed]
- Montenigro, P.H.; Alosco, M.L.; Martin, B.M.; Daneshvar, D.H.; Mez, J.; Chaisson, C.E.; Nowinski, C.J.; Au, R.; McKee, A.C.; Cantu, R.C.; et al. Cumulative Head Impact Exposure Predicts Later-Life Depression, Apathy, Executive Dysfunction, and Cognitive Impairment in Former High School and College Football Players. J. Neurotrauma 2017, 34, 328–340. [Google Scholar] [CrossRef]
- Baugh, C.M.; Kiernan, P.T.; Kroshus, E.; Daneshvar, D.H.; Montenigro, P.H.; McKee, A.C.; Stern, R.A. Frequency of head-impact-related outcomes by position in NCAA division I collegiate football players. J. Neurotrauma 2015, 32, 314–326. [Google Scholar] [CrossRef] [Green Version]
- Ocwieja, K.E.; Mihalik, J.P.; Marshall, S.W.; Schmidt, J.D.; Trulock, S.C.; Guskiewicz, K.M. The effect of play type and collision closing distance on head impact biomechanics. Ann. Biomed. Eng. 2012, 40, 90–96. [Google Scholar] [CrossRef]
- Oliver, J.M.; Jones, M.T.; Kirk, K.M.; Gable, D.A.; Repshas, J.T.; Johnson, T.A.; Andreasson, U.; Norgren, N.; Blennow, K.; Zetterberg, H. Effect of Docosahexaenoic Acid on a Biomarker of Head Trauma in American Football. Med. Sci. Sports Exerc. 2016, 48, 974–982. [Google Scholar] [CrossRef]
- Stamm, J.M.; Bourlas, A.P.; Baugh, C.M.; Fritts, N.G.; Daneshvar, D.H.; Martin, B.M.; McClean, M.D.; Tripodis, Y.; Stern, R.A. Age of first exposure to football and later-life cognitive impairment in former NFL players. Neurology 2015, 84, 1114–1120. [Google Scholar] [CrossRef] [Green Version]
- Oliver, J.M.; Jones, M.T.; Kirk, K.M.; Gable, D.A.; Repshas, J.T.; Johnson, T.A.; Andreasson, U.; Norgren, N.; Blennow, K.; Zetterberg, H. Serum Neurofilament Light in American Football Athletes over the Course of a Season. J. Neurotrauma 2016, 33, 1784–1789. [Google Scholar] [CrossRef] [PubMed]
- Rubin, L.H.; Tierney, R.; Kawata, K.; Wesley, L.; Lee, J.H.; Blennow, K.; Zetterberg, H.; Langford, D. NFL blood levels are moderated by subconcussive impacts in a cohort of college football players. Brain Inj. 2019, 33, 456–462. [Google Scholar] [CrossRef] [PubMed]
- Shahim, P.; Tegner, Y.; Marklund, N.; Blennow, K.; Zetterberg, H. Neurofilament light and tau as blood biomarkers for sports-related concussion. Neurology 2018, 90, e1780–e1788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wallace, C.; Smirl, J.D.; Zetterberg, H.; Blennow, K.; Bryk, K.; Burma, J.; Dierijck, J.; Wright, A.D.; van Donkelaar, P. Heading in soccer increases serum neurofilament light protein and SCAT3 symptom metrics. BMJ Open Sport Exerc. Med. 2018, 4, e000433. [Google Scholar] [CrossRef] [PubMed]
- Wirsching, A.; Chen, Z.; Bevilacqua, Z.W.; Huibregtse, M.E.; Kawata, K. Association of Acute Increase in Plasma Neurofilament Light with Repetitive Subconcussive Head Impacts: A Pilot Randomized Control Trial. J. Neurotrauma 2019, 36, 548–553. [Google Scholar] [CrossRef] [PubMed]
- Woodcock, T.; Morganti-Kossmann, M.C. The role of markers of inflammation in traumatic brain injury. Front. Neurol. 2013, 4, 18. [Google Scholar] [CrossRef] [Green Version]
- Nitta, M.E.; Savitz, J.; Nelson, L.D.; Teague, T.K.; Hoelzle, J.B.; McCrea, M.A.; Meier, T.B. Acute elevation of serum inflammatory markers predicts symptom recovery after concussion. Neurology 2019, 93, e497–e507. [Google Scholar] [CrossRef]
- Kitson, A.P.; Metherel, A.H.; Chen, C.T.; Domenichiello, A.F.; Trepanier, M.O.; Berger, A.; Bazinet, R.P. Effect of dietary docosahexaenoic acid (DHA) in phospholipids or triglycerides on brain DHA uptake and accretion. J. Nutr. Biochem. 2016, 33, 91–102. [Google Scholar] [CrossRef] [Green Version]
- Lewis, M.D. Concussions, Traumatic Brain Injury, and the Innovative Use of Omega-3s. J. Am. Coll. Nutr. 2016, 35, 469–475. [Google Scholar] [CrossRef]
- Weiser, M.J.; Mucha, B.; Denheyer, H.; Atkinson, D.; Schanz, N.; Vassiliou, E.; Benno, R.H. Dietary docosahexaenoic acid alleviates autistic-like behaviors resulting from maternal immune activation in mice. Prostaglandins Leukot. Essent. Fat. Acids 2016, 106, 27–37. [Google Scholar] [CrossRef]
- Weiser, M.J.; Butt, C.M.; Mohajeri, M.H. Docosahexaenoic Acid and Cognition throughout the Lifespan. Nutrients 2016, 8, 99. [Google Scholar] [CrossRef] [PubMed]
- DiNicolantonio, J.J.; O′Keefe, J.H. Importance of maintaining a low omega-6/omega-3 ratio for reducing inflammation. Open Heart 2018, 5, e000946. [Google Scholar] [CrossRef] [PubMed]
- Deyama, S.; Shimoda, K.; Suzuki, H.; Ishikawa, Y.; Ishimura, K.; Fukuda, H.; Hitora-Imamura, N.; Ide, S.; Satoh, M.; Kaneda, K.; et al. Resolvin E1/E2 ameliorate lipopolysaccharide-induced depression-like behaviors via ChemR23. Psychopharmacology 2018, 235, 329–336. [Google Scholar] [CrossRef] [PubMed]
- Rey, C.; Nadjar, A.; Buaud, B.; Vaysse, C.; Aubert, A.; Pallet, V.; Laye, S.; Joffre, C. Resolvin D1 and E1 promote resolution of inflammation in microglial cells in vitro. Brain Behav. Immun. 2016, 55, 249–259. [Google Scholar] [CrossRef] [PubMed]
- Mills, J.D.; Bailes, J.E.; Sedney, C.L.; Hutchins, H.; Sears, B. Omega-3 fatty acid supplementation and reduction of traumatic axonal injury in a rodent head injury model. J. Neurosurg. 2011, 114, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.; Ding, Y.; Kong, W.; Li, T.; Chen, H. Docosahexaenoic Acid (DHA) Provides Neuroprotection in Traumatic Brain Injury Models via Activating Nrf2-ARE Signaling. Inflammation 2018, 41, 1182–1193. [Google Scholar] [CrossRef]
- Heileson, J.L.; Anzalone, A.J.; Carbuhn, A.F.; Askow, A.T.; Stone, J.D.; Turner, S.M.; Hillyer, L.M.; Ma, D.W.L.; Luedke, J.A.; Jagim, A.R.; et al. The effect of omega-3 fatty acids on a biomarker of head trauma in NCAA football athletes: A multi-site, non-randomized study. J. Int. Soc. Sports Nutr. 2021, 18, 65. [Google Scholar] [CrossRef]
- Weaver, K.L.; Ivester, P.; Seeds, M.; Case, L.D.; Arm, J.P.; Chilton, F.H. Effect of dietary fatty acids on inflammatory gene expression in healthy humans. J. Biol. Chem. 2009, 284, 15400–15407. [Google Scholar] [CrossRef] [Green Version]
- Metcalfe, L.D.; Schmitz, A.A.; Pelka, J.R. Rapid preparation of fatty acid esters from lipids for gas chromatographic analysis. Anal. Chem. 1966, 38, 514–515. [Google Scholar] [CrossRef]
- Sergeant, S.; Ruczinski, I.; Ivester, P.; Lee, T.C.; Morgan, T.M.; Nicklas, B.J.; Mathias, R.A.; Chilton, F.H. Impact of methods used to express levels of circulating fatty acids on the degree and direction of associations with blood lipids in humans. Br. J. Nutr. 2016, 115, 251–261. [Google Scholar] [CrossRef] [Green Version]
- Bates, D.; Maechler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 5823. [Google Scholar] [CrossRef]
- Kuznetsova, A.; Brockhoff, P.B.; Christensen, R.H.B. lmerTest Package: Tests in Linear Mixed Effects Models. J. Stat. Softw. 2016, 82, 1–26. [Google Scholar]
- Lenth, R.V. Least-squares means: The R package lsmeans. J. Stat. Softw. 2016, 69, 1–33. [Google Scholar] [CrossRef] [Green Version]
- Wickham, H.; Hester, J.; Chang, W. Devtools: Tools to Make Developing R Packages Easier. 2020. R Package Version 1, 0, 9000. Available online: https://devtools.r-lib.org/ (accessed on 30 March 2022).
- Ben-Shachar, M.; Lüdecke, D.; Makowski, D. Estimation of Effect Size Indices and Standardized Parameters. J. Open Source Softw. 2020, 5, 2815. [Google Scholar] [CrossRef]
- Cohen, J. Statistical Power Analysis for the Behavioral Sciences, 2nd ed.; Erlbaum: Hillsdale, NJ, USA, 1988. [Google Scholar]
- Kassambara, A. ggpubr: ‘ggplot2′ Based Publication Ready Plots. 2020. R Package Version 0.4, 0, 438. Available online: https://rpkgs.datanovia.com/ggpubr/ (accessed on 30 March 2022).
- Di Battista, A.P.; Churchill, N.; Rhind, S.G.; Richards, D.; Hutchison, M.G. Evidence of a distinct peripheral inflammatory profile in sport-related concussion. J. Neuroinflamm. 2019, 16, 17. [Google Scholar] [CrossRef]
- Meier, T.B.; Nelson, L.D.; Huber, D.L.; Bazarian, J.J.; Hayes, R.L.; McCrea, M.A. Prospective Assessment of Acute Blood Markers of Brain Injury in Sport-Related Concussion. J. Neurotrauma 2017, 34, 3134–3142. [Google Scholar] [CrossRef] [PubMed]
- Anzalone, A.; Carbuhn, A.; Jones, L.; Gallop, A.; Smith, A.; Johnson, P.; Swearingen, L.; Moore, C.; Rimer, E.; McBeth, J.; et al. The Omega-3 Index in National Collegiate Athletic Association Division I Collegiate Football Athletes. J. Athl. Train. 2019, 54, 7–11. [Google Scholar] [CrossRef] [Green Version]
- Von Schacky, C.; Kemper, M.; Haslbauer, R.; Halle, M. Low Omega-3 Index in 106 German elite winter endurance athletes: A pilot study. Int. J. Sport Nutr. Exerc. Metab. 2014, 24, 559–564. [Google Scholar] [CrossRef] [Green Version]
- Walker, R.E.; Jackson, K.H.; Tintle, N.L.; Shearer, G.C.; Bernasconi, A.; Masson, S.; Latini, R.; Heydari, B.; Kwong, R.Y.; Flock, M.; et al. Predicting the effects of supplemental EPA and DHA on the omega-3 index. Am. J. Clin. Nutr. 2019, 110, 1034–1040. [Google Scholar] [CrossRef]
- Sparkes, C.; Sinclair, A.J.; Gibson, R.A.; Else, P.L.; Meyer, B.J. High variability in erythrocyte, plasma and whole blood EPA and DHA levels in response to supplementation. Nutrients 2020, 12, 1017. [Google Scholar] [CrossRef] [Green Version]
- Giza, C.C.; Hovda, D.A. The neurometabolic cascade of concussion. J. Athl. Train. 2001, 36, 228. [Google Scholar] [CrossRef] [Green Version]
- Hviid, C.V.B.; Knudsen, C.S.; Parkner, T. Reference interval and preanalytical properties of serum neurofilament light chain in Scandinavian adults. Scand. J. Clin. Lab. Investig. 2020, 80, 291–295. [Google Scholar] [CrossRef] [PubMed]
- Hachem, M.B.; Van, A.L.; Picq, M.; Bernoud-Hubac, N. Brain targeting with docosahexaenoic acid as a prospective therapy for neurodegenerative diseases and its passage across blood brain barrier. Biochimie 2020, 170, 208–211. [Google Scholar] [CrossRef] [PubMed]
- Bazinet, R.P.; Bernoud-Hubac, N.; Lagarde, M. How the plasma lysophospholipid and unesterified fatty acid pools supply the brain with docosahexaenoic acid. Prostaglandins Leukot. Essent. Fat. Acids 2019, 142, 1–3. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Adverse Event | Placebo (n = 19) | Treatment (n = 19) |
|---|---|---|
| Any, N (%) | 10 (52) | 12 (63) |
| >2 participants, N, (%) | ||
| Respiratory infection | 9 (47) | 3 (15) |
| Acne | 1 (5) | 2 (10) |
| GI distress (stomach upset) | 0 (0) | 3 (15) |
| Halitosis and eructation | 0 (0) | 4 (21) |
| Concussion * | 1 (5) | 2 (10) |
| Serious AEs, N = 0 (0%) | ||
| Baseline | Week 8 | Week 12 | Week 17 | Week 21 | Week 26 | Week 33 | |
|---|---|---|---|---|---|---|---|
| ARA:DHA ratio treatment | 7:1 | 3:1 | 3:1 | 3:1 | 3:1 | 4:1 | 5:1 |
| ARA:DHA ratio placebo | 5:1 | 5:1 | 5:1 | 5:1 | 5:1 | 6:1 | 5:1 |
| ARA:EPA ratio treatment | 23:1 | 6:1 | 8:1 | 6:1 | 8:1 | 11:1 | 21:1 |
| ARA:EPA ratio placebo | 19:1 | 20:1 | 21:1 | 16:1 | 18:1 | 17:1 | 16:1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mullins, V.A.; Graham, S.; Cummings, D.; Wood, A.; Ovando, V.; Skulas-Ray, A.C.; Polian, D.; Wang, Y.; Hernandez, G.D.; Lopez, C.M.; et al. Effects of Fish Oil on Biomarkers of Axonal Injury and Inflammation in American Football Players: A Placebo-Controlled Randomized Controlled Trial. Nutrients 2022, 14, 2139. https://doi.org/10.3390/nu14102139
Mullins VA, Graham S, Cummings D, Wood A, Ovando V, Skulas-Ray AC, Polian D, Wang Y, Hernandez GD, Lopez CM, et al. Effects of Fish Oil on Biomarkers of Axonal Injury and Inflammation in American Football Players: A Placebo-Controlled Randomized Controlled Trial. Nutrients. 2022; 14(10):2139. https://doi.org/10.3390/nu14102139
Chicago/Turabian StyleMullins, Veronica A., Sarah Graham, Danielle Cummings, Alva Wood, Vanessa Ovando, Ann C. Skulas-Ray, Dennis Polian, Yiwei Wang, Gerson D. Hernandez, Claudia M. Lopez, and et al. 2022. "Effects of Fish Oil on Biomarkers of Axonal Injury and Inflammation in American Football Players: A Placebo-Controlled Randomized Controlled Trial" Nutrients 14, no. 10: 2139. https://doi.org/10.3390/nu14102139
APA StyleMullins, V. A., Graham, S., Cummings, D., Wood, A., Ovando, V., Skulas-Ray, A. C., Polian, D., Wang, Y., Hernandez, G. D., Lopez, C. M., Raikes, A. C., Brinton, R. D., & Chilton, F. H. (2022). Effects of Fish Oil on Biomarkers of Axonal Injury and Inflammation in American Football Players: A Placebo-Controlled Randomized Controlled Trial. Nutrients, 14(10), 2139. https://doi.org/10.3390/nu14102139