
Genistein: Dual Role in Women’s Health


Abstract
1. Introduction
1.1. Genistein in Food
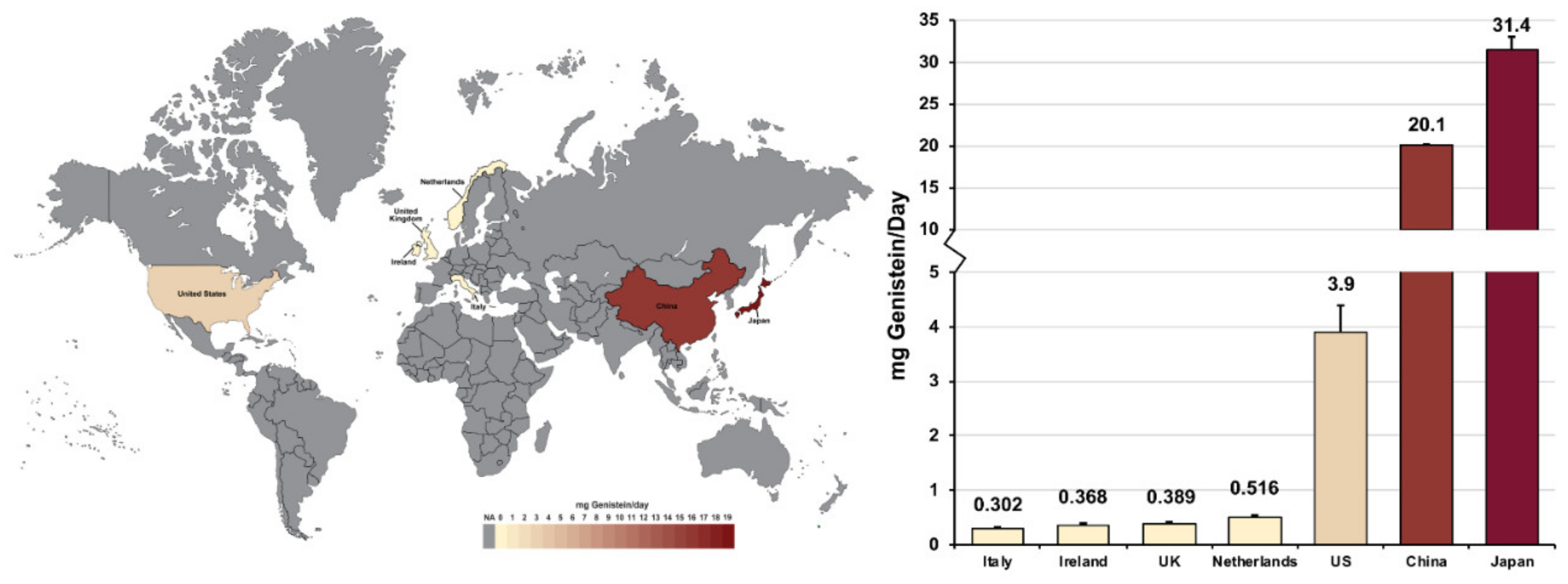
1.2. Genistein Levels in Various Populations
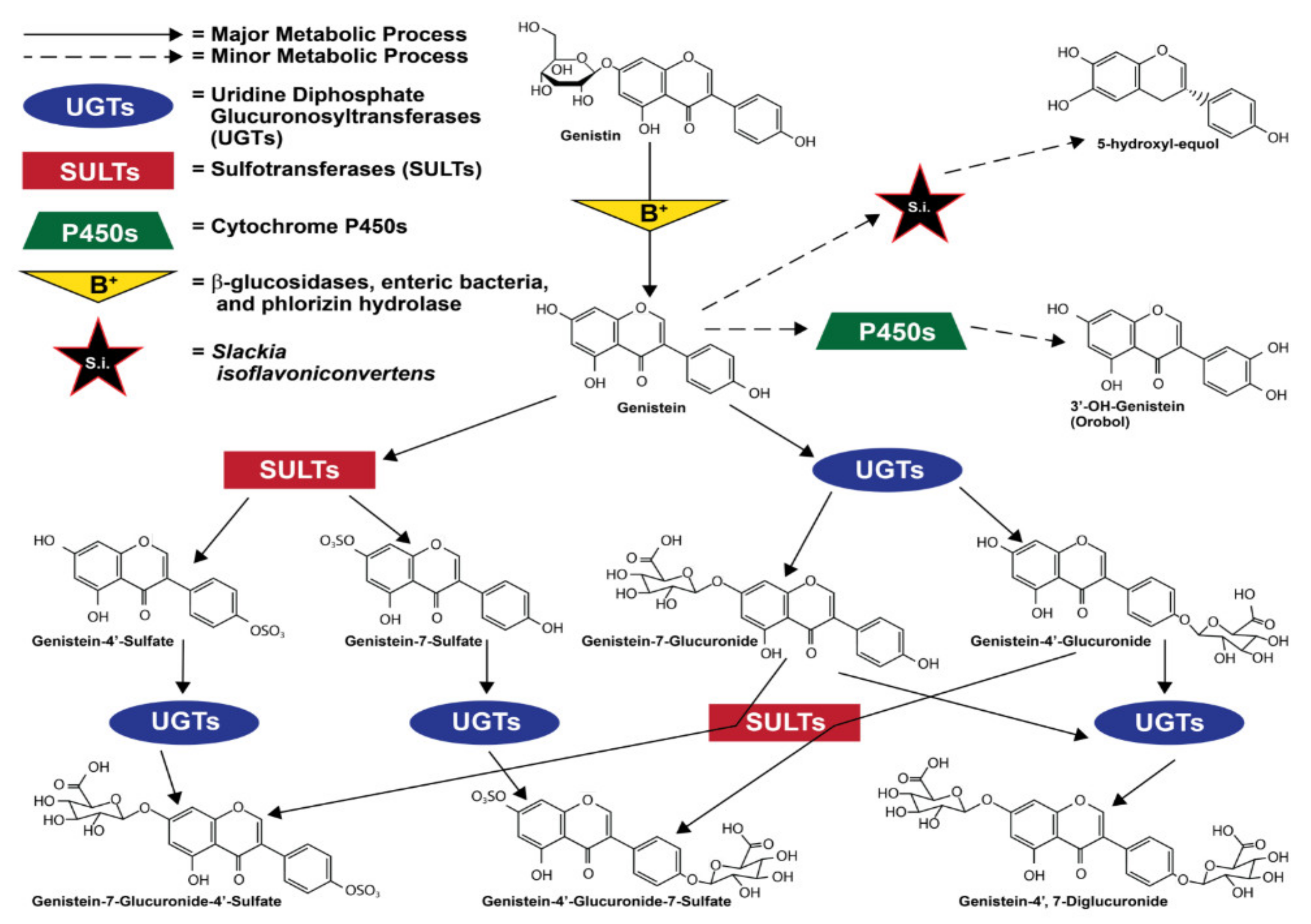
1.3. Metabolism and Metabolites of Genistein
1.4. Estrogenic Effects of Genistein
2. Biological Effects and Mechanism(s) of Genistein
2.1. In Vivo Experimental and Clinical Findings
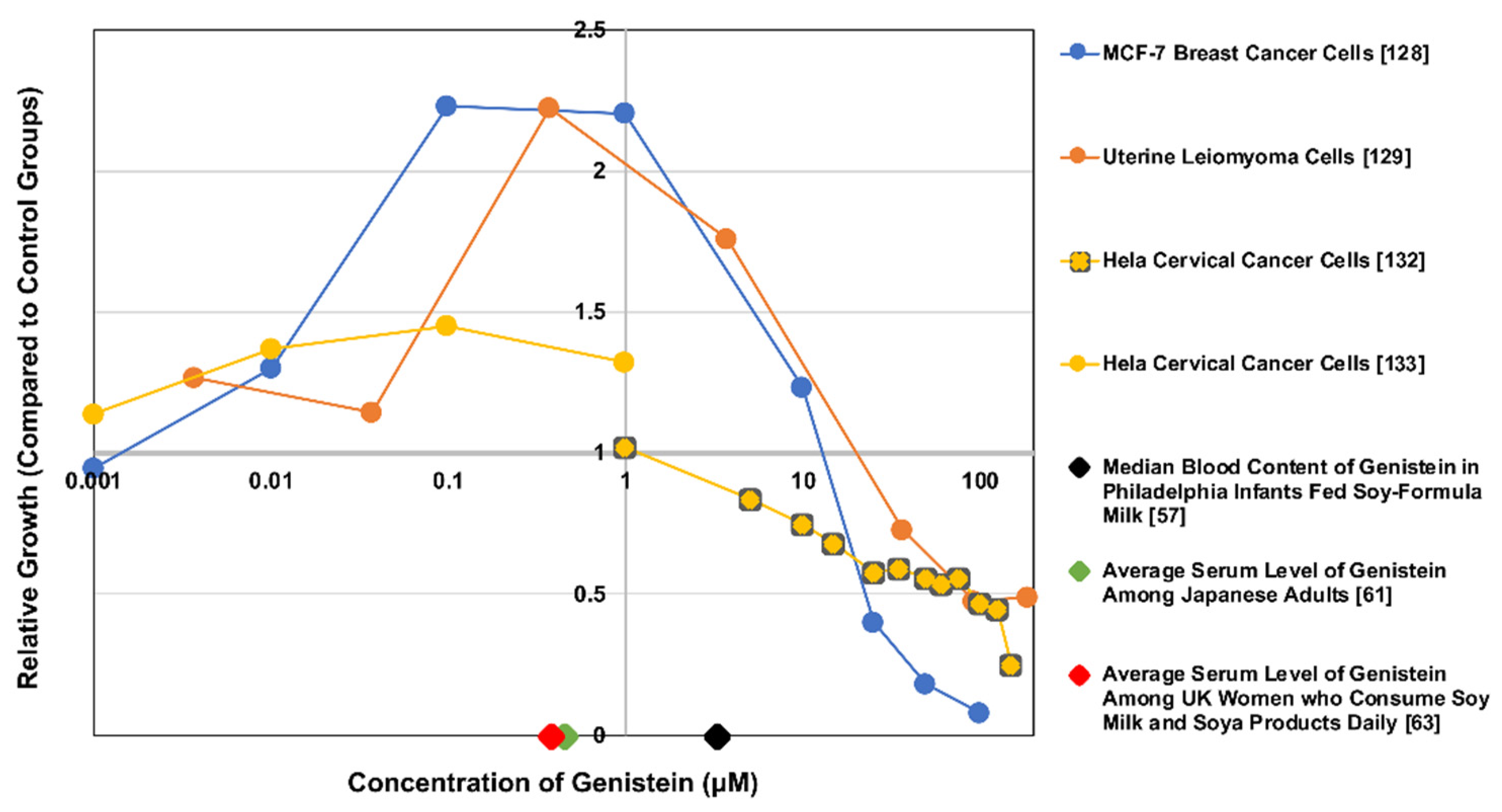
2.2. In Vitro Experimental Findings
3. Genistein and Women’s Diseases
3.1. Genistein and Obesity
3.2. Genistein and Breast Cancer
3.3. Genistein and Uterine Leiomyoma
3.4. Genistein and Endometriosis
3.5. Genistein and Endometrial Cancer
3.6. Genistein and Polycystic Ovarian Syndrome
3.7. Genistein and Cervical Cancer
3.8. Genistein and Menopause (Hormone Regulation)
4. Clinical Therapeutic Options
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- National Center for Biotechnology Information. Compound Summary for CID 5280961, Genistein. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/Genistein (accessed on 17 December 2020).
- UCSF Health. A Guide to Foods Rich in Soy. Available online: https://www.ucsfhealth.org/education/a-guide-to-foods-rich-in-soy (accessed on 17 December 2020).
- The Scientific Advisory Committee on Nutrition (SACN). Statement on the Potential Risks from High Levels of Soya Phytoestrogens in the Infant Diet; Committee on Toxicity of Chemicals in Food, Consumer Products and the Environment: London, UK, 2003. [Google Scholar]
- Bhagwat, S.; Hayowitz, D.B.; Holden, J.M. USDA Database for the Isoflavone Content of Selected Foods, Release 2.0; US Department of Agriculture: Washington, DC, USA, 2008. [CrossRef]
- Economic Research Service. ERS Charts of Note: Soybeans & Oil Crops. Available online: https://www.ers.usda.gov/data-products/charts-of-note/charts-of-note/?topicId=14851#:~:text=Brazil%20is%20a%20leading%20global,the%202018%2F19%20marketing%20year (accessed on 17 December 2020).
- The Good Food Institute. Plant-Based Market Overview. Available online: https://www.gfi.org/marketresearch (accessed on 17 December 2020).
- Coward, L.; Smith, M.; Kirk, M.; Barnes, S. Chemical modification of isoflavones in soyfoods during cooking and processing. Am. J. Clin. Nutr. 1998, 68, 1486S–1491S. [Google Scholar] [CrossRef]
- King, R.; Bignell, C.M. Concentrations of isoflavone phytoestrogens and their glucosides in Australian soya beans and soya foods. Aust. J. Nutr. Dietics 2000, 57, 70–78. [Google Scholar]
- Liggins, J.; Bluck, L.J.C.; Runswick, S.; Atkinson, C.; Coward, W.A.; Bingham, S.A. Daidzein and genistein contents of vegetables. Br. J. Nutr. 2007, 84, 717–725. [Google Scholar] [CrossRef]
- Murphy, P.A.; Barua, K.; Hauck, C.C. Solvent extraction selection in the determination of isoflavones in soy foods. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2002, 777, 129–138. [Google Scholar] [CrossRef]
- Nguyenle, T.; Wang, E.; Cheung, A.P. An investigation on the extraction and concentration of isoflavones in soy-based products. J. Pharm. Biomed. Anal. 1995, 14, 221–232. [Google Scholar] [CrossRef]
- Rostagno, M.A.; Palma, M.; Barroso, C.G. Fast analysis of soy isoflavones by high-performance liquid chromatography with monolithic columns. Anal. Chim. Acta 2007, 582, 243–249. [Google Scholar] [CrossRef] [PubMed]
- Umphress, S.T.; Murphy, S.P.; Franke, A.A.; Custer, L.J.; Blitz, C.L. Isoflavone content of foods with soy additives. J. Food Compos. Anal. 2005, 18, 533–550. [Google Scholar] [CrossRef]
- Wang, H.; Murphy, P.A. Isoflavone Content in Commercial Soybean Foods. J. Agric. Food Chem. 1994, 42, 1666–1673. [Google Scholar] [CrossRef]
- Genovese, M.I.; Hassimoto, N.M.A.; Lajolo, F.M. Isoflavone profile and antioxidant activity of Brazilian soybean varieties. Food Sci. Technol. Int. 2005, 11, 205–211. [Google Scholar] [CrossRef]
- Coward, L.; Barnes, N.C.; Setchell, K.D.R.; Barnes, S. Genistein, daidzein, and their beta-glycoside conjugates: Antitumor isoflavones in soybean foods from American and Asian diets. J. Agric. Food Chem. 1993, 41, 1961–1967. [Google Scholar] [CrossRef]
- Chiou, R.Y.; Cheng, S.L. Isoflavone transformation during soybean koji preparation and subsequent miso fermentation supplemented with ethanol and NaCl. J. Agric. Food Chem. 2001, 49, 3656–3660. [Google Scholar] [CrossRef]
- Charron, C.S.; Allen, F.L.; Johnson, R.D.; Pantalone, V.R.; Sams, C.E. Correlations of oil and protein with isoflavone concentration in soybean [Glycine max (L.) Merr.]. J. Agric. Food Chem. 2005, 53, 7128–7135. [Google Scholar] [CrossRef]
- Murphy, P.A.; Barua, K.; Song, T. Soy Isoflavones in Foods: Database Development; American Chemical Society Symposium Series; American Chemical Society: Washington, DC, USA, 1998; pp. 138–149. [Google Scholar]
- Xu, X.; Wang, H.J.; Murphy, P.A.; Cook, L.; Hendrich, S. Daidzein is a more bioavailable soymilk isoflavone than is genistein in adult women. J. Nutr. 1994, 124, 825–832. [Google Scholar] [CrossRef]
- Achouri, A.; Boye, J.I.; Belanger, D. Soybean isoflavones: Efficacy of extraction conditions and effect of food type on extractability. Food Res. Int. 2005, 38, 1199–1204. [Google Scholar] [CrossRef]
- Barnes, S.; Kirk, M.; Coward, L. Isoflavones and their conjugates in soy foods: Extraction conditions and analysis by HPLC-mass spectrometry. J. Agric. Food Chem. 1994, 42, 2466–2474. [Google Scholar] [CrossRef]
- Franke, A.A.; Custer, L.J.; Wang, W.; Shi, C.Y. HPLC analysis of isoflavonoids and other phenolic agents from foods and from human fluids. Proc. Soc. Exp. Biol. Med. 1998, 217, 263–273. [Google Scholar] [CrossRef] [PubMed]
- Coward, L.; Kirk, M.; Albin, N.; Barnes, S. Analysis of plasma isoflavones by reversed-phase HPLC-multiple reaction ion monitoring-mass spectrometry. Clin. Chim. Acta 1996, 247, 121–142. [Google Scholar] [CrossRef]
- Downing, J.M.; Chung, O.K.; Seib, P.A.; Hubbard, J.D. Pressurized Solvent Extraction of Genistein and Its β-Glucoside Conjugates from Soybean Flours and Soy-Based Foods. Cereal Chem. 2007, 84, 44–47. [Google Scholar] [CrossRef]
- Fang, N.; Yu, S.; Badger, T.M. Comprehensive Phytochemical Profile of Soy Protein Isolate. J. Agric. Food Chem. 2004, 52, 4012–4020. [Google Scholar] [CrossRef]
- Seo, A.; Morr, C.V. Improved high-performance liquid chromatographic analysis of phenolic acids and isoflavonoids from soybean protein products. J. Agric. Food Chem. 1984, 32, 530–533. [Google Scholar] [CrossRef]
- Wang, C.; Ma, Q.; Pagadala, S.; Sherrard, M.S.; Krishnan, P.G. Changes of isoflavones during processing of soy protein isolates. J. Am. Oil Chem. Soc. 1998, 75, 337–341. [Google Scholar] [CrossRef]
- Wang, H.-J.; Murphy, P.A. Mass Balance Study of Isoflavones during Soybean Processing. J. Agric. Food Chem. 1996, 44, 2377–2383. [Google Scholar] [CrossRef]
- Franke, A.A.; Hankin, J.H.; Yu, M.C.; Maskarinec, G.; Low, S.H.; Custer, L.J. Isoflavone levels in soy foods consumed by multiethnic populations in Singapore and Hawaii. J. Agric. Food Chem. 1999, 47, 977–986. [Google Scholar] [CrossRef] [PubMed]
- Murphy, P.A.; Song, T.; Buseman, G.; Barua, K.; Beecher, G.R.; Trainer, D.; Holden, J. Isoflavones in Retail and Institutional Soy Foods. J. Agric. Food Chem. 1999, 47, 2697–2704. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.S.; Lee, B.H.; Kim, J.H.; Kim, N.S. Concentration of phytoestrogens in soybeans and soybean products in Korea. J. Sci. Food Agric. 2000, 80, 1709–1712. [Google Scholar] [CrossRef]
- Fukutake, M.; Takahashi, M.; Ishida, K.; Kawamura, H.; Sugimura, T.; Wakabayashi, K. Quantification of genistein and genistin in soybeans and soybean products. Food Chem. Toxicol. 1996, 34, 457–461. [Google Scholar] [CrossRef]
- Murphy, P.A. Green soy beans (Edame, dry), Soy beans (small Jade Black), Natto (DHA), Natto (fermented soy beans), Soy bean butter (full fat), Natto Kibun, Soy nuts (full fat), Soy nuts (plain halves), Soy flakes (white, not roasted), Green soy beans (Edame, fresh). Daidzein, Genistein, Glycitein. Unpublished data.
- Nakamura, Y.; Tsuji, S.; Tonogai, Y. Determination of the levels of isoflavonoids in soybeans and soy-derived foods and estimation of isoflavonoids in the Japanese daily intake. J. AOAC Int. 2000, 83, 635–650. [Google Scholar] [CrossRef]
- Toda, T.; Sakamoto, A.; Takayanagi, T.; Yokotsuka, K. Changes in Isoflavone Compositions of Sobean Foods during Cooking Process. Food Sci. Technol. Res. 2000, 6, 314–319. [Google Scholar] [CrossRef]
- Hutabarat, L.S.; Greenfield, H.; Mulholland, M. Isoflavones and Coumestrol in Soybeans and Soybean Products from Australia and Indonesia. J. Food Compos. Anal. 2001, 14, 43–58. [Google Scholar] [CrossRef]
- Hutchins, A.M.; Slavin, J.L.; Lampe, J.W. Urinary isoflavonoid phytoestrogen and lignan Excretion After Consumption of Fermented and Unfermented Soy Products. J. Am. Diet. Assoc. 1995, 95, 545–551. [Google Scholar] [CrossRef]
- Nakajima, N.; Nozaki, N.; Ishihara, K.; Ishikawa, A.; Tsuji, H. Analysis of isoflavone content in tempeh, a fermented soybean, and preparation of a new isoflavone-enriched tempeh. J. Biosci. Bioeng. 2005, 100, 685–687. [Google Scholar] [CrossRef] [PubMed]
- Thompson, L.U.; Boucher, B.A.; Liu, Z.; Cotterchio, M.; Kreiger, N. Phytoestrogen Content of Foods Consumed in Canada, Including Isoflavones, Lignans, and Coumestan. Nutr. Cancer 2006, 54, 184–201. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.J.; Broemeling, L.D.; Marshall, M.V.; Ramanujam, V.M. A simplified method to quantify isoflavones in commercial soybean diets and human urine after legume consumption. Cancer Epidemiol. Biomark. Prev. 1995, 4, 497–503. [Google Scholar]
- Mitani, K.; Narimatsu, S.; Kataoka, H. Determination of daidzein and genistein in soybean foods by automated on-line in-tube solid-phase microextraction coupled to high-performance liquid chromatography. J. Chromatogr. A 2003, 986, 169–177. [Google Scholar] [CrossRef]
- Yamabe, S.; Kobayashi-Hattori, K.; Kaneko, K.; Endo, H.; Takita, T. Effect of soybean varieties on the content and composition of isoflavone in rice-koji miso. Food Chem. 2007, 100, 369–374. [Google Scholar] [CrossRef]
- Pamela, L.H.-R.; Stephen, B.; Marion, L.; Lori, C.; Mandel, J.E.; Jocelyn, K.; Esther, M.J.; Michelle, S. Assessing Phytoestrogen Exposure in Epidemiologic Studies: Development of a Database (United States). Cancer Causes Control 2000, 11, 289–298. [Google Scholar] [CrossRef]
- Lin, P.-Y.; Lai, H.-M. Bioactive Compounds in Legumes and Their Germinated Products. J. Agric. Food Chem. 2006, 54, 3807–3814. [Google Scholar] [CrossRef]
- Müllner, C.; Sontag, G. Determination of some phytoestrogens in soybeans and their processed products with HPLC and coulometric electrode array detection. Fresenius’ J. Anal. Chem. 1999, 364, 261–265. [Google Scholar] [CrossRef]
- Nakamura, Y.; Kaihara, A.; Yoshii, K.; Tsumura, Y.; Ishimitsu, S.; Tonogai, Y. Content and Composition of Isoflavonoids in Mature or Immature Beans and Bean Sprouts Consumed in Japan. J. Health Sci. 2001, 47, 394–406. [Google Scholar] [CrossRef][Green Version]
- Plaza, L.; de Ancos, B.; Cano, P.M. Nutritional and health-related compounds in sprouts and seeds of soybean (Glycine max), wheat (Triticum aestivum.L) and alfalfa (Medicago sativa) treated by a new drying method. Eur. Food Res. Technol. 2003, 216, 138–144. [Google Scholar] [CrossRef]
- Wang, G.; Kuan, S.S.; Francis, O.J.; Ware, G.M.; Carman, A.S. A simplified HPLC method for the determination of phytoestrogens in soybean and its processed products. J. Agric. Food Chem. 1990, 38, 185–190. [Google Scholar] [CrossRef]
- Grün, I.U.; Adhikari, K.; Li, C.; Li, Y.; Lin, B.; Zhang, J.; Fernando, L.N. Changes in the Profile of Genistein, Daidzein, and Their Conjugates during Thermal Processing of Tofu. J. Agric. Food Chem. 2001, 49, 2839–2843. [Google Scholar] [CrossRef] [PubMed]
- Krenn, L.; Unterrieder, I.; Ruprechter, R. Quantification of isoflavones in red clover by high-performance liquid chromatography. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2002, 777, 123–128. [Google Scholar] [CrossRef]
- Gentile, C.; Tesoriere, L.; Butera, D.; Fazzari, M.; Monastero, M.; Allegra, M.; Livrea, M.A. Antioxidant Activity of Sicilian Pistachio (Pistacia vera L. Var. Bronte) Nut Extract and Its Bioactive Components. J. Agric. Food Chem. 2007, 55, 643–648. [Google Scholar] [CrossRef]
- Johns, P.; Dowlati, L.; Wargo, W. Determination of isoflavones in ready-to-feed soy-based infant formula. J. AOAC Int. 2003, 86, 72–78. [Google Scholar] [CrossRef]
- Setchell, K.D.R.; Welsh, M.B.; Lim, C.K. High-performance liquid chromatographic analysis of phytoestrogens in soy protein preparations with ultraviolet, electrochemical and thermospray mass spectrometric detection. J. Chromatogr. A 1987, 386, 315–323. [Google Scholar] [CrossRef]
- Franke, A.A.; Custer, L.J.; Cerna, C.M.; Narala, K. Rapid HPLC analysis of dietary phytoestrogens from legumes and from human urine. Proc. Soc. Exp. Biol. Med. 1995, 208, 18–26. [Google Scholar] [CrossRef]
- Antonelli, M.L.; Faberi, A.; Pastorini, E.; Samperi, R.; Lagana, A. Simultaneous quantitation of free and conjugated phytoestrogens in Leguminosae by liquid chromatography-tandem mass spectrometry. Talanta 2005, 66, 1025–1033. [Google Scholar] [CrossRef]
- Cao, Y.; Calafat, A.M.; Doerge, D.R.; Umbach, D.M.; Bernbaum, J.C.; Twaddle, N.C.; Ye, X.; Rogan, W.J. Isoflavones in urine, saliva, and blood of infants: Data from a pilot study on the estrogenic activity of soy formula. J. Expo. Sci. Environ. Epidemiol. 2008, 19, 223–234. [Google Scholar] [CrossRef]
- Tseng, M.; Olufade, T.; Kurzer, M.S.; Wähälä, K.; Fang, C.Y.; van der Schouw, Y.T.; Daly, M.B. Food Frequency Questionnaires and Overnight Urines Are Valid Indicators of Daidzein and Genistein Intake in U.S. Women Relative to Multiple 24-h Urine Samples. Nutr. Cancer 2008, 60, 619–626. [Google Scholar] [CrossRef]
- Van Erp-Baart, M.-A.J.; Brants, H.A.M.; Kiely, M.; Mulligan, A.; Turrini, A.; Sermoneta, C.; Kilkkinen, A.; Valsta, L.M. Isoflavone intake in four different European countries: The VENUS approach. Br. J. Nutr. 2003, 89, S25–S30. [Google Scholar] [CrossRef]
- Huang, M.-H.; Luetters, C.; Buckwalter, G.J.; Seeman, T.E.; Gold, E.B.; Sternfeld, B.; Greendale, G.A. Dietary genistein intake and cognitive performance in a multiethnic cohort of midlife women. Menopause 2006, 13, 621–630. [Google Scholar] [CrossRef]
- Yamamoto, S.; Sobue, T.; Sasaki, S.; Kobayashi, M.; Arai, Y.; Uehara, M.; Adlercreutz, H.; Watanabe, S.; Takahashi, T.; Iitoi, Y.; et al. Validity and reproducibility of a self-administered food-frequency questionnaire to assess isoflavone intake in a japanese population in comparison with dietary records and blood and urine isoflavones. J. Nutr. 2001, 131, 2741–2747. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.A.; Wen, W.; Xiang, Y.B.; Barnes, S.; Liu, D.; Cai, Q.; Zheng, W.; Shu, X.O. Assessment of dietary isoflavone intake among middle-aged Chinese men. J. Nutr. 2007, 137, 1011–1016. [Google Scholar] [CrossRef]
- Verkasalo, P.K.; Appleby, P.N.; Allen, N.E.; Davey, G.; Adlercreutz, H.; Key, T.J. Soya intake and plasma concentrations of daidzein and genistein: Validity of dietary assessment among eighty British women (Oxford arm of the European Prospective Investigation into Cancer and Nutrition). Br. J. Nutr. 2001, 86, 415–421. [Google Scholar] [CrossRef] [PubMed]
- Adlercreutz, H.; Yamada, T.; Wähälä, K.; Watanabe, S. Maternal and neonatal phytoestrogens in Japanese women during birth. Am. J. Obstet. Gynecol. 1999, 180, 737–743. [Google Scholar] [CrossRef]
- Newton, J.; Nelson, M. China Uses One-Third of World’s Soybeans. Available online: https://www.fb.org/market-intel/china-uses-one-third-of-worlds-soybeans (accessed on 17 December 2020).
- Rosell, M.S.; Appleby, P.N.; Spencer, E.A.; Key, T.J. Soy intake and blood cholesterol concentrations: A cross-sectional study of 1033 pre- and postmenopausal women in the Oxford arm of the European Prospective Investigation into Cancer and Nutrition. Am. J. Clin. Nutr. 2004, 80, 1391–1396. [Google Scholar] [CrossRef] [PubMed]
- Tempfer, C.B.M.D.; Bentz, E.-K.M.D.; Leodolter, S.M.D.; Tscherne, G.M.D.; Reuss, F.M.D.; Cross, H.S.; Huber, J.C. Phytoestrogens in clinical practice: A review of the literature. Fertil. Steril. 2007, 87, 1243–1249. [Google Scholar] [CrossRef]
- Kurzer, M.S.; Xu, X. Dietary Phytoestrogens. Annu. Rev. Nutr. 1997, 17, 353–381. [Google Scholar] [CrossRef]
- Barrett, A. Long-Term World Soybean Outlook. Available online: https://ussoy.org/long-term-world-soybean-outlook/#:~:text=The%20increase%20in%20world%20meat,of%20coarse%20grains%20and%20soybeans.&text=Global%20soybean%20utilization%20is%20estimated,in%20second%20at%2061%20MMT (accessed on 17 December 2020).
- Walsh, K.R.; Haak, S.J.; Bohn, T.; Tian, Q.; Schwartz, S.J.; Failla, M.L. Isoflavonoid glucosides are deconjugated and absorbed in the small intestine of human subjects with ileostomies. Am. J. Clin. Nutr. 2007, 85, 1050–1056. [Google Scholar] [CrossRef]
- Bokkenheuser, V.D.; Shackleton, C.H.L.; Winter, J. Hydrolysis of dietary flavonoid glycosides by strains of intestinal Bacteroides from humans. Biochem. J. 1987, 248, 953–956. [Google Scholar] [CrossRef]
- Mattison, D.R.; Karyakina, N.; Goodman, M.; LaKind, J.S. Pharmaco- and toxicokinetics of selected exogenous and endogenous estrogens: A review of the data and identification of knowledge gaps. Crit. Rev. Toxicol. 2014, 44, 696–724. [Google Scholar] [CrossRef]
- Hu, M. Commentary: Bioavailability of Flavonoids and Polyphenols: Call to Arms. Mol. Pharm. 2007, 4, 803–806. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Hu, M. Absorption and metabolism of flavonoids in the caco-2 cell culture model and a perused rat intestinal model. Drug Metab. Dispos. 2002, 30, 370–377. [Google Scholar] [CrossRef]
- Rozman, K.K.; Bhatia, J.; Calafat, A.M.; Chambers, C.; Culty, M.; Etzel, R.A.; Flaws, J.A.; Hansen, D.K.; Hoyer, P.B.; Jeffery, E.H.; et al. NTP-CERHR expert panel report on the reproductive and developmental toxicity of genistein. Birth Defects Res. Part. B Dev. Reprod. Toxicol. 2006, 77, 485–638. [Google Scholar] [CrossRef]
- Yang, Z.; Zhu, W.; Gao, S.; Xu, H.; Wu, B.; Kulkarni, K.; Singh, R.; Tang, L.; Hu, M. Simultaneous determination of genistein and its four phase II metabolites in blood by a sensitive and robust UPLC–MS/MS method: Application to an oral bioavailability study of genistein in mice. J. Pharm. Biomed. Anal. 2010, 53, 81–89. [Google Scholar] [CrossRef]
- Setchell, K.D.R.; Brown, N.M.; Desai, P.; Zimmer-Nechemias, L.; Wolfe, B.E.; Brashear, W.T.; Kirschner, A.S.; Cassidy, A.; Heubi, J.E. Bioavailability of Pure Isoflavones in Healthy Humans and Analysis of Commercial Soy Isoflavone Supplements. J. Nutr. 2001, 131, 1362S–1375S. [Google Scholar] [CrossRef]
- Riches, Z.; Stanley, E.L.; Bloomer, J.C.; Coughtrie, M.W.H. Quantitative Evaluation of the Expression and Activity of Five Major Sulfotransferases (SULTs) in Human Tissues: The SULT “Pie”. Drug Metab. Dispos. 2009, 37, 2255–2261. [Google Scholar] [CrossRef]
- Boonpawa, R.; Spenkelink, A.; Punt, A.; Rietjens, I.M.C.M. In vitro-in silico-based analysis of the dose-dependent in vivo oestrogenicity of the soy phytoestrogen genistein in humans. Br. J. Pharmacol. 2017, 174, 2739–2757. [Google Scholar] [CrossRef]
- Liu, J.; Yu, X.; Zhong, S.; Han, W.; Liang, Z.; Ye, L.; Zhao, J.; Liu, M.; Liu, S.; Wei, Q.; et al. Hepatic and renal metabolism of genistein: An individual-based model to predict glucuronidation behavior of genistein in different organs. J. Pharm. Biomed. Anal. 2017, 139, 252–262. [Google Scholar] [CrossRef]
- Kurkela, M.; Garcia-Horsmant, J.A.; Luukkanen, L.; Mörsky, S.; Taskinen, J.; Baumann, M.; Kostiainen, R.; Hirvonen, J.; Finel, M. Expression and characterization of recombinant human UDP-glucuronosyltransferases (UGTs): UGT1A9 is more resistant to detergent inhibition than the other UGTs and was purified as an active dimeric enzyme. J. Biol. Chem. 2003, 278, 3536–3544. [Google Scholar] [CrossRef]
- Yuan, B.; Zhen, H.; Jin, Y.; Xu, L.; Jiang, X.; Sun, S.; Li, C.; Xu, H. Absorption and Plasma Disposition of Genistin Differ from Those of Genistein in Healthy Women. J. Agric. Food Chem. 2012, 60, 1428–1436. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.-C.; Nair, M.G. Metabolism of Daidzein and Genistein by Intestinal Bacteria. J. Nat. Prod. 1995, 58, 1892–1896. [Google Scholar] [CrossRef] [PubMed]
- Kulling, S.E.; Honig, D.M.; Metzler, M. Oxidative Metabolism of the Soy Isoflavones Daidzein and Genistein in Humans in Vitro and in Vivo. J. Agric. Food Chem. 2001, 49, 3024–3033. [Google Scholar] [CrossRef]
- Bursztyka, J.; Perdu, E.; Tulliez, J.; Debrauwer, L.; Delous, G.; Canlet, C.; De Sousa, G.; Rahmani, R.; Benfenati, E.; Cravedi, J.-P. Comparison of genistein metabolism in rats and humans using liver microsomes and hepatocytes. Food Chem. Toxicol. 2008, 46, 939–948. [Google Scholar] [CrossRef]
- Breinholt, V.M.; Rasmussen, S.E.; Brosen, K.; Friedberg, T.H. In vitro Metabolism of Genistein and Tangeretin by Human and Murine Cytochrome P450s. Pharmacol. Toxicol. 2003, 93, 14–22. [Google Scholar] [CrossRef]
- Hu, M.; Krausz, K.; Chen, J.; Ge, X.; Li, J.; Gelboin, H.L.; Gonzalez, F.J. Identification of cyp1a2 as the main isoform for the phase i hydroxylated metabolism of genistein and a prodrug converting enzyme of methylated isoflavones. Drug Metab. Dispos. 2003, 31, 924–931. [Google Scholar] [CrossRef]
- Matthies, A.; Blaut, M.; Braune, A. Isolation of a human intestinal bacterium capable of daidzein and genistein conversion. Appl. Environ. Microbiol. 2009, 75, 1740–1744. [Google Scholar] [CrossRef]
- Hosoda, K.; Furuta, T.; Yokokawa, A.; Ishii, K. Identification and quantification of daidzein-7-glucuronide-4’-sulfate, genistein-7-glucuronide-4’-sulfate and genistein-4’,7-diglucuronide as major metabolites in human plasma after administration of kinako. Anal. Bioanal. Chem. 2010, 397, 1563–1572. [Google Scholar] [CrossRef]
- Munro, I.C.; Harwood, M.; Hlywka, J.J.; Stephen, A.M.; Doull, J.; Flamm, W.G.; Adlercreutz, H. Soy Isoflavones: A Safety Review. Nutr. Rev. 2003, 61, 1–33. [Google Scholar] [CrossRef]
- Hoey, L.; Rowland, I.R.; Lloyd, A.S.; Clarke, D.B.; Wiseman, H. Influence of soya-based infant formula consumption on isoflavone and gut microflora metabolite concentrations in urine and on faecal microflora composition and metabolic activity in infants and children. Br. J. Nutr. 2004, 91, 607–616. [Google Scholar] [CrossRef] [PubMed]
- Jackson, R.L.; Greiwe, J.S.; Schwen, R.J. Emerging evidence of the health benefits of S-equol, an estrogen receptor beta agonist. Nutr. Rev. 2011, 69, 432–448. [Google Scholar] [CrossRef]
- Choi, E.J.; Kim, G.H. The antioxidant activity of daidzein metabolites, Odesmethylangolensin and equol, in HepG2 cells. Mol. Med. Rep. 2014, 9, 328–332. [Google Scholar] [CrossRef]
- Wei, X.J.; Wu, J.; Ni, Y.D.; Lu, L.Z.; Zhao, R.Q. Antioxidant effect of a phytoestrogen equol on cultured muscle cells of embryonic broilers. Vitr. Cell Dev. Biol. Anim. 2011, 47, 735–741. [Google Scholar] [CrossRef]
- Arora, A.; Nair, M.G.; Strasburg, G.M. Antioxidant Activities of Isoflavones and Their Biological Metabolites in a Liposomal System. Arch. Biochem. Biophys. 1998, 356, 133–141. [Google Scholar] [CrossRef]
- Mayo, B.; Vazquez, L.; Belen Florez, A. Equol: A Bacterial Metabolite from The Daidzein Isoflavone and Its Presumed Beneficial Health Effects. Nutrients 2019, 11, 2231. [Google Scholar] [CrossRef] [PubMed]
- Kuiper, G.G.J.M.; Lemmen, J.G.; Carlsson, B.; Corton, J.C.; Safe, S.H.; van der Saag, P.T.; van der Burg, B.; Gustafsson, J.-Å. Interaction of Estrogenic Chemicals and Phytoestrogens with Estrogen Receptor β. Endocrinology 1998, 139, 4252–4263. [Google Scholar] [CrossRef] [PubMed]
- Koenig, A.; Buskiewicz, I.; Huber, S.A. Age-associated changes in estrogen receptor ratios correlate with increased female susceptibility to coxsackievirus B3-induced myocarditis. Front. Immunol. 2017, 8, 1585. [Google Scholar] [CrossRef] [PubMed]
- Qing, X.; Zhihong, L.; You-Hong, C.; Chiang-Ching, H.; Erica, M.; Ping, Y.; Magdy, P.M.; Edmond, C.; Scott, R.; Joy, I.; et al. Promoter Methylation Regulates Estrogen Receptor 2 in Human Endometrium and Endometriosis. Biol. Reprod. 2007, 77, 681–687. [Google Scholar] [CrossRef]
- Abbasi, S. Estrogen Receptor-Beta Gene Polymorphism in women with Breast Cancer at the Imam Khomeini Hospital Complex, Iran. BMC Med. Genet. 2010, 11, 109. [Google Scholar] [CrossRef]
- Maggiolini, M.; Vivacqua, A.; Fasanella, G.; Recchia, A.G.; Sisci, D.; Pezzi, V.; Montanaro, D.; Musti, A.M.; Picard, D.; Ando, S. The G protein-coupled receptor GPR30 mediates c-fos up-regulation by 17beta-estradiol and phytoestrogens in breast cancer cells. J. Biol. Chem. 2004, 279, 27008–27016. [Google Scholar] [CrossRef] [PubMed]
- Thomas, P.; Pang, Y.; Filardo, E.J.; Dong, J. Identity of an estrogen membrane receptor coupled to a G protein in human breast cancer cells. Endocrinology 2005, 146, 624–632. [Google Scholar] [CrossRef] [PubMed]
- Du, Z.-R.; Feng, X.-Q.; Li, N.; Qu, J.-X.; Feng, L.; Chen, L.; Chen, W.-F. G protein-coupled estrogen receptor is involved in the anti-inflammatory effects of genistein in microglia. Phytomedicine 2018, 43, 11–20. [Google Scholar] [CrossRef]
- Okura, A.; Arakawa, H.; Oka, H.; Yoshinari, T.; Monden, Y. Effect of genistein on topoisomerase activity and on the growth of [VAL 12]Ha- ras-transformed NIH 3T3 cells. Biochem. Biophys. Res. Commun. 1988, 157, 183–189. [Google Scholar] [CrossRef]
- Johnson, A.; Roberts, L.; Elkins, G. Complementary and Alternative Medicine for Menopause. J. Evid. Based Integr. Med. 2019, 24, 2515690X19829380. [Google Scholar] [CrossRef]
- Sobhy, M.M.K.; Mahmoud, S.S.; El-Sayed, S.H.; Rizk, E.M.A.; Raafat, A.; Negm, M.S.I. Impact of treatment with a Protein Tyrosine Kinase Inhibitor (Genistein) on acute and chronic experimental Schistosoma mansoni infection. Exp. Parasitol. 2018, 185, 115–123. [Google Scholar] [CrossRef]
- Evans, B.A.; Griffiths, K.; Morton, M.S. Inhibition of 5 alpha-reductase in genital skin fibroblasts and prostate tissue by dietary lignans and isoflavonoids. J. Endocrinol. 1995, 147, 295–302. [Google Scholar] [CrossRef]
- Barnes, S. Effect of genistein on in vitro and in vivo models of cancer. J. Nutr. 1995, 125, 777S–783S. [Google Scholar] [CrossRef]
- Messing, E.; Gee, J.R.; Saltzstein, D.R.; Kim, K.; DiSant’Agnese, A.; Kolesar, J.; Harris, L.; Faerber, A.; Havighurst, T.; Young, J.M.; et al. A phase 2 cancer chemoprevention biomarker trial of isoflavone G-2535 (genistein) in presurgical bladder cancer patients. Cancer Prev. Res. 2012, 5, 621–630. [Google Scholar] [CrossRef]
- Rüfer, C.E.; Kulling, S.E. Antioxidant Activity of Isoflavones and Their Major Metabolites Using Different in Vitro Assays. J. Agric. Food Chem. 2006, 54, 2926–2931. [Google Scholar] [CrossRef]
- De Gregorio, C.; Marini, H.; Alibrandi, A.; Di Benedetto, A.; Bitto, A.; Adamo, E.B.; Altavilla, D.; Irace, C.; Di Vieste, G.; Pancaldo, D.; et al. Genistein supplementation and cardiac function in postmenopausal women with metabolic syndrome: Results from a pilot strain-echo study. Nutrients 2017, 9, 584. [Google Scholar] [CrossRef]
- Borradaile, N.M.; De Dreu, L.E.; Wilcox, L.J.; Edwards, J.Y.; Huff, M.W. Soya phytoestrogens, genistein and daidzein, decrease apolipoprotein B secretion from HepG2 cells through multiple mechanisms. Biochem. J. 2002, 366, 531–539. [Google Scholar] [CrossRef]
- Turner, R.T.; Iwaniec, U.T.; Andrade, J.E.; Branscum, A.J.; Neese, S.L.; Olson, D.A.; Wagner, L.; Wang, V.C.; Schantz, S.L.; Helferich, W.G. Genistein administered as a once-daily oral supplement had no beneficial effect on the tibia in rat models for postmenopausal bone loss. Menopause 2013, 20, 677–686. [Google Scholar] [CrossRef]
- Thangavel, P.; Puga-Olguín, A.; Rodríguez-Landa, J.F.; Zepeda, R.C. Genistein as Potential Therapeutic Candidate for Menopausal Symptoms and Other Related Diseases. Molecules 2019, 24, 3892. [Google Scholar] [CrossRef]
- Odle, B.; Dennison, N.; Al-Nakkash, L.; Broderick, T.L.; Plochocki, J.H. Genistein treatment improves fracture resistance in obese diabetic mice. BMC Endocr. Disord. 2017, 17, 55. [Google Scholar] [CrossRef]
- Singh, P.; Sharma, S.; Kumar Rath, S. Genistein Induces Deleterious Effects during Its Acute Exposure in Swiss Mice. Biomed. Res. Int. 2014, 2014, 619617. [Google Scholar] [CrossRef] [PubMed]
- Wisniewski, A.B.; Klein, S.L.; Lakshmanan, Y.; Gearhart, J.P. Exposure to Genistein During Gestation and Lactation Demasculinizes the Reproductive System in Rats. J. Urol. 2003, 169, 1582–1586. [Google Scholar] [CrossRef]
- Lewis, R.W.; Brooks, N.; Milburn, G.M.; Soames, A.; Stone, S.; Hall, M.; Ashby, J. The effects of the phytoestrogen genistein on the postnatal development of the rat. Toxicol. Sci. 2003, 71, 74–83. [Google Scholar] [CrossRef] [PubMed]
- Kerrie, B.B.; Leena, H.-C. Genistein: Does It Prevent or Promote Breast Cancer? Environ. Health Perspect. 2000, 108, 701–708. [Google Scholar] [CrossRef]
- Korde, L.A.; Wu, A.H.; Fears, T.; Nomura, A.M.Y.; West, D.W.; Kolonel, L.N.; Pike, M.C.; Hoover, R.N.; Ziegler, R.G. Childhood Soy Intake and Breast Cancer Risk in Asian American Women. Cancer Epidemiol. Biomark. Prev. 2009, 18, 1050–1059. [Google Scholar] [CrossRef] [PubMed]
- Joanne, T.; Michelle, C.; Beatrice, A.B.; Nancy, K.; Lilian, U.T. Adolescent Dietary Phytoestrogen Intake and Breast Cancer Risk (Canada). Cancer Causes Control 2006, 17, 1253–1261. [Google Scholar] [CrossRef]
- Yatani, R.; Chigusa, I.; Akazaki, K.; Stemmermann, G.N.; Welsh, R.A.; Correa, P. Geographic pathology of latent prostatic carcinoma. Int. J. Cancer 1982, 29, 611–616. [Google Scholar] [CrossRef]
- Messina, M.; Nagata, C.; Wu, A.H. Estimated Asian Adult Soy Protein and Isoflavone Intakes. Nutr. Cancer 2006, 55, 1–12. [Google Scholar] [CrossRef]
- Tham, D.M.; Gardner, C.D.; Haskell, W.L. Clinical review 97-Potential health benefits of dietary phytoestrogens: A review of the clinical, epidemiological, and mechanistic evidence. J. Clin. Endocrinol. Metab. 1998, 83, 2223–2235. [Google Scholar] [CrossRef] [PubMed]
- Akiyama, T.; Ishida, J.; Nakagawa, S.; Ogawara, H.; Watanabe, S.; Itoh, N.; Shibuya, M.; Fukami, Y. Genistein, a specific inhibitor of tyrosine-specific protein kinases. J. Biol. Chem. 1987, 262, 5592–5595. [Google Scholar] [CrossRef]
- Agarwal, R. Cell signaling and regulators of cell cycle as molecular targets for prostate cancer prevention by dietary agents. Biochem. Pharmacol. 2000, 60, 1051–1059. [Google Scholar] [CrossRef]
- Chen, W.-F.; Wong, M.-S. Genistein Enhances Insulin-Like Growth Factor Signaling Pathway in Human Breast Cancer (MCF-7) Cells. J. Clin. Endocrinol. Metab. 2004, 89, 2351–2359. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.T.Y.; Sathyamoorthy, N.; Phang, J.M. Molecular effects of genistein on estrogen receptor mediated pathways. Carcinogenesis 1996, 17, 271–275. [Google Scholar] [CrossRef] [PubMed]
- Moore, A.B.; Castro, L.; Yu, L.; Zheng, X.; Di, X.; Sifre, M.I.; Kissling, G.E.; Newbold, R.R.; Bortner, C.D.; Dixon, D. Stimulatory and inhibitory effects of genistein on human uterine leiomyoma cell proliferation are influenced by the concentration. Hum. Reprod. 2007, 22, 2623–2631. [Google Scholar] [CrossRef] [PubMed]
- Kohara, Y.; Kuwahara, R.; Kawaguchi, S.; Jojima, T.; Yamashita, K. Perinatal exposure to genistein, a soy phytoestrogen, improves spatial learning and memory but impairs passive avoidance learning and memory in offspring. Physiol. Behav. 2014, 130, 40–46. [Google Scholar] [CrossRef] [PubMed]
- Vandenberg, L.N. Non-Monotonic Dose Responses in Studies of Endocrine Disrupting Chemicals: Bisphenol a as a Case Study. Dose-Response 2013, 12, 259–276. [Google Scholar] [CrossRef] [PubMed]
- Hussain, A.; Harish, G.; Prabhu, S.A.; Mohsin, J.; Khan, M.A.; Rizvi, T.A.; Sharma, C. Inhibitory effect of genistein on the invasive potential of human cervical cancer cells via modulation of matrix metalloproteinase-9 and tissue inhibitiors of matrix metalloproteinase-1 expression. Cancer Epidemiol. 2012, 36, e387–e393. [Google Scholar] [CrossRef]
- Chen, H.-H.; Chen, S.-P.; Zheng, Q.-L.; Nie, S.-P.; Li, W.-J.; Hu, X.-J.; Xie, M.-Y. Genistein promotes proliferation of human cervical cancer cells through estrogen receptor-mediated PI3K/Akt-NF-κB pathway. J. Cancer 2018, 9, 288–295. [Google Scholar] [CrossRef]
- Shen, H.-H.; Huang, S.-Y.; Kung, C.-W.; Chen, S.-Y.; Chen, Y.-F.; Cheng, P.-Y.; Lam, K.-K.; Lee, Y.-M. Genistein ameliorated obesity accompanied with adipose tissue browning and attenuation of hepatic lipogenesis in ovariectomized rats with high-fat diet. J. Nutr. Biochem. 2019, 67, 111–122. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.K.; Nelson-Dooley, C.; Della-Fera, M.A.; Yang, J.Y.; Zhang, W.; Duan, J.; Hartzell, D.L.; Hamrick, M.W.; Baile, C.A. Genistein decreases food intake, body weight, and fat pad weight and causes adipose tissue apoptosis in ovariectomized female mice. J. Nutr. 2006, 136, 409–414. [Google Scholar] [CrossRef] [PubMed]
- Naaz, A.; Yellayi, S.; Zakroczymski, M.A.; Bunick, D.; Doerge, D.R.; Lubahn, D.B.; Helferich, W.G.; Cooke, P.S. The Soy Isoflavone Genistein Decreases Adipose Deposition in Mice. Endocrinology 2003, 144, 3315–3320. [Google Scholar] [CrossRef]
- Dolinoy, D.C.; Weidman, J.R.; Waterland, R.A.; Jirtle, R.L. Maternal genistein alters coat color and protects A(vy) mouse offspring from obesity by modifying the fetal epigenome. Environ. Health Perspect. 2006, 114, 567–572. [Google Scholar] [CrossRef]
- Chang, H.C.; Doerge, D.R. Dietary genistein inactivates rat thyroid peroxidase in vivo without an apparent hypothyroid effect. Toxicol. Appl. Pharm. 2000, 168, 244–252. [Google Scholar] [CrossRef] [PubMed]
- Nogowski, L.; Nowak, K.W.; Kaczmarek, P.; Mackowiak, P. The influence of coumestrol, zearalenone, and genistein administration on insulin receptors and insulin secretion in ovariectomized rats. J. Recept. Signal. Transduct. Res. 2002, 22, 449–457. [Google Scholar] [CrossRef] [PubMed]
- Tsai, Y.C.; Leu, S.Y.; Peng, Y.J.; Lee, Y.M.; Hsu, C.H.; Chou, S.C.; Yen, M.H.; Cheng, P.Y. Genistein suppresses leptin-induced proliferation and migration of vascular smooth muscle cells and neointima formation. J. Cell. Mol. Med. 2017, 21, 422–431. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Lee, S.H.; Ji, H.; Kim, J.-E.; Yoo, R.; Kim, J.H.; Suk, S.; Huh, C.S.; Park, J.H.Y.; Heo, Y.-S.; et al. Orobol, an Enzyme-Convertible Product of Genistein, exerts Anti-Obesity Effects by Targeting Casein Kinase 1 Epsilon. Sci. Rep. 2019, 9, 8942. [Google Scholar] [CrossRef]
- Skov, A.R.; Toubro, S.; Ronn, B.; Holm, L.; Astrup, A. Randomized trial on protein vs carbohydrate in ad libitum fat reduced diet for the treatment of obesity. Int. J. Obes. Relat. Metab. Disord. 1999, 23, 528–536. [Google Scholar] [CrossRef]
- Velasquez, M.T.; Bhathena, S.J. Role of Dietary Soy Protein in Obesity. Int. J. Med. Sci. 2007, 4, 72–82. [Google Scholar] [CrossRef]
- Mikkelsen, P.B.; Toubro, S.; Astrup, A. Effect of fat-reduced diets on 24-h energy expenditure: Comparisons between animal protein, vegetable protein, and carbohydrate. Am. J. Clin. Nutr. 2000, 72, 1135–1141. [Google Scholar] [CrossRef]
- Bosello, O.; Cominacini, L.; Zocca, I.; Garbin, U.; Compri, R.; Davoli, A.; Brunetti, L. Short- and long-term effects of hypocaloric diets containing proteins of different sources on plasma lipids and apoproteins of obese subjects. Ann. Nutr. Metab. 1988, 32, 206–214. [Google Scholar] [CrossRef]
- Yamashita, T.; Sasahara, T.; Pomeroy, S.E.; Collier, G.; Nestel, P.J. Arterial compliance, blood pressure, plasma leptin, and plasma lipids in women are improved with weight reduction equally with a meat-based diet and a plant-based diet. Metabolism 1998, 47, 1308–1314. [Google Scholar] [CrossRef]
- Anderson, J.W.; Hoie, L.H. Weight loss and lipid changes with low-energy diets: Comparator study of milk-based versus soy-based liquid meal replacement interventions. J. Am. Coll. Nutr. 2005, 24, 210–216. [Google Scholar] [CrossRef]
- Akhlaghi, M.; Zare, M.; Nouripour, F. Effect of Soy and Soy Isoflavones on Obesity-Related Anthropometric Measures: A Systematic Review and Meta-analysis of Randomized Controlled Clinical Trials. Adv. Nutr. 2017, 8, 705–717. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, S.; Li, Y.; Wang, Z.; Sarkar, F.H. Multi-targeted therapy of cancer by genistein. Cancer Lett. 2008, 269, 226–242. [Google Scholar] [CrossRef] [PubMed]
- Mai, Z.; Blackburn, G.L.; Zhou, J.R. Genistein sensitizes inhibitory effect of tamoxifen on the growth of estrogen receptor-positive and HER2-overexpressing human breast cancer cells. Mol. Carcinog. 2007, 46, 534–542. [Google Scholar] [CrossRef]
- Pons, D.G.; Nadal-Serrano, M.; Torrens-Mas, M.; Oliver, J.; Roca, P. The Phytoestrogen Genistein Affects Breast Cancer Cells Treatment Depending on the ERα/ERβ Ratio. J. Cell. Biochem. 2016, 117, 218–229. [Google Scholar] [CrossRef] [PubMed]
- Hilakivi-Clarke, L.; Andrade, J.E.; Helferich, W. Is Soy Consumption Good or Bad for the Breast? J. Nutr. 2010, 140, 2326S–2334S. [Google Scholar] [CrossRef]
- Liu, R.; Yu, X.; Chen, X.; Zhong, H.; Liang, C.; Xu, X.; Xu, W.; Cheng, Y.; Wang, W.; Yu, L.; et al. Individual factors define the overall effects of dietary genistein exposure on breast cancer patients. Nutr. Res. 2019, 67, 1–16. [Google Scholar] [CrossRef]
- Castro, L.; Gao, X.; Moore, A.B.; Yu, L.; Di, X.; Kissling, G.E.; Dixon, D. A High Concentration of Genistein Induces Cell Death in Human Uterine Leiomyoma Cells by Autophagy. Expert Opin. Environ. Biol. 2016, 5 (Suppl. 1), 10.4172/2325-9655.S1-003. [Google Scholar] [CrossRef] [PubMed]
- Di, X.; Andrews, D.M.K.; Tucker, C.J.; Yu, L.; Moore, A.B.; Zheng, X.; Castro, L.; Hermon, T.; Xiao, H.; Dixon, D. A high concentration of genistein down-regulates activin A, Smad3 and other TGF-β pathway genes in human uterine leiomyoma cells. Exp. Mol. Med. 2012, 44, 281–292. [Google Scholar] [CrossRef]
- Miyake, A.; Takeda, T.; Isobe, A.; Wakabayashi, A.; Nishimoto, F.; Morishige, K.-I.; Sakata, M.; Kimura, T. Repressive effect of the phytoestrogen genistein on estradiol-induced uterine leiomyoma cell proliferation. Gynecol. Endocrinol. 2009, 25, 403–409. [Google Scholar] [CrossRef] [PubMed]
- Di, X.; Yu, L.; Moore, A.B.; Castro, L.; Zheng, X.; Hermon, T.; Dixon, D. A low concentration of genistein induces estrogen receptor-alpha and insulin-like growth factor-I receptor interactions and proliferation in uterine leiomyoma cells. Hum. Reprod. 2008, 23, 1873–1883. [Google Scholar] [CrossRef]
- Yu, L.; Ham, K.; Gao, X.; Castro, L.; Yan, Y.; Kissling, G.E.; Tucker, C.J.; Flagler, N.; Dong, R.; Archer, T.K.; et al. Epigenetic regulation of transcription factor promoter regions by low-dose genistein through mitogen-activated protein kinase and mitogen-and-stress activated kinase 1 nongenomic signaling. Cell Commun. Signal. 2016, 14, 18. [Google Scholar] [CrossRef]
- Simon, G.A.; Fletcher, H.M.; Golden, K.; McFarlane-Anderson, N.D. Urinary isoflavone and lignan phytoestrogen levels and risk of uterine fibroid in Jamaican women. Maturitas 2015, 82, 170–175. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, S.; Bergqvist, A.; Chapron, C.; D’Hooghe, T.; Dunselman, G.; Greb, R.; Hummelshoj, L.; Prentice, A.; Saridogan, E. ESHRE guideline for the diagnosis and treatment of endometriosis. Hum. Reprod. 2005, 20, 2698–2704. [Google Scholar] [CrossRef]
- Bitto, A.; Granese, R.; Polito, F.; Triolo, O.; Giordano, D.; Squadrito, F.; D’Anna, R.; Santamaria, A. Genistein reduces angiogenesis and apoptosis in women with endometrial hyperplasia. Botanics 2015, 5, 27–32. [Google Scholar] [CrossRef][Green Version]
- Mumford, S.L.; Weck, J.; Kannan, K.; Louis, G.M.B. Urinary phytoestrogen concentrations are not associated with incident endometriosis in premenopausal women. J. Nutr. 2017, 147, 227–234. [Google Scholar] [CrossRef]
- Tsuchiya, M.; Miura, T.; Hanaoka, T.; Iwasaki, M.; Sasaki, H.; Tanaka, T.; Nakao, H.; Katoh, T.; Ikenoue, T.; Kabuto, M.; et al. Effect of soy isoflavones on endometriosis: Interaction with estrogen receptor 2 gene polymorphism. Epidemiology 2007, 18, 402–408. [Google Scholar] [CrossRef]
- Yen, C.F.; Kim, M.R.; Lee, C.L. Epidemiologic Factors Associated with Endometriosis in East Asia. Gynecol. Minim. Invasive 2019, 8, 4–11. [Google Scholar] [CrossRef]
- Cotronero, M.S.; Lamartiniere, C.A. Pharmacologic, but not dietary, genistein supports endometriosis in a rat model. Toxicol. Sci. 2001, 61, 68–75. [Google Scholar] [CrossRef]
- Yavuz, E.M.D.; Oktem, M.M.D.; Esinler, I.M.D.; Toru, S.A.M.D.; Zeyneloglu, H.B.M.D. Genistein causes regression of endometriotic implants in the rat model. Fertil. Steril. 2007, 88, 1129–1134. [Google Scholar] [CrossRef] [PubMed]
- Sutrisno, S.; Wulandari, R.R.C.L.; Sulistyowati, D.W.W.; Wulandari, R.F.; Wahyuni, E.S.; Yueniwati, Y.; Santoso, S. Effect of genistein on proinflammatory cytokines and estrogen receptor-β in mice model of endometriosis. Asian Pac. J. Reprod. 2015, 4, 96–99. [Google Scholar] [CrossRef]
- Sutrisno, S.; Sulistyorini, C.; Manungkalit, E.M.; Winarsih, L.; Noorhamdani, N.; Winarsih, S. The effect of genistein on TGF-β signal, dysregulation of apoptosis, cyclooxygenase-2 pathway, and NF-kB pathway in mice peritoneum of endometriosis model. Middle East. Fertil. Soc. J. 2017, 22, 295–299. [Google Scholar] [CrossRef]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed]
- Morice, P.D.P.; Leary, A.M.D.; Creutzberg, C.P.; Abu-Rustum, N.P.; Darai, E.P. Endometrial cancer. Lancet 2015, 387, 1094–1108. [Google Scholar] [CrossRef]
- Malloy, K.M.; Wang, J.; Clark, L.H.; Fang, Z.; Sun, W.; Yin, Y.; Kong, W.; Zhou, C.X.; Bae-Jump, V.L. Novasoy and genistein inhibit endometrial cancer cell proliferation through disruption of the AKT/mTOR and MAPK signaling pathways. Am. J. Transl. Res. 2018, 10, 784–795. [Google Scholar] [PubMed]
- Konstantakopoulos, N.; Montgomery, K.G.; Chamberlain, N.; Quinn, M.A.; Baker, M.S.; Rice, G.E.; Georgiou, H.M.; Campbell, I.G. Changes in gene expressions elicited by physiological concentrations of genistein on human endometrial cancer cells. Mol. Carcinog. 2006, 45, 752–763. [Google Scholar] [CrossRef]
- Hu, Y.-C.; Wu, C.-T.; Lai, J.-N.; Tsai, Y.-T. Detection of a negative correlation between prescription of Chinese herbal products containing coumestrol, genistein or daidzein and risk of subsequent endometrial cancer among tamoxifen-treated female breast cancer survivors in Taiwan between 1998 and 2008: A population-based study. J. Ethnopharmacol. 2015, 169, 356–362. [Google Scholar] [CrossRef]
- Sampey, B.P.; Lewis, T.D.; Barbier, C.S.; Makowski, L.; Kaufman, D.G. Genistein effects on stromal cells determines epithelial proliferation in endometrial co-cultures. Exp. Mol. Pathol. 2011, 90, 257–263. [Google Scholar] [CrossRef]
- Zhang, G.Q.; Chen, J.L.; Liu, Q.; Zhang, Y.; Zeng, H.; Zhao, Y. Soy Intake Is Associated With Lower Endometrial Cancer Risk: A Systematic Review and Meta-Analysis of Observational Studies. Medicine 2015, 94, e2281. [Google Scholar] [CrossRef]
- Lee, J.Y.; Kim, H.S.; Song, Y.S. Genistein as a Potential Anticancer Agent against Ovarian Cancer. J. Tradit. Complement. Med. 2012, 2, 96–104. [Google Scholar] [CrossRef]
- Sirmans, S.M.; Pate, K.A. Epidemiology, diagnosis, and management of polycystic ovary syndrome. Clin. Epidemiol. 2013, 6, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Khani, B.; Mehrabian, F.; Khalesi, E.; Eshraghi, A. Effect of soy phytoestrogen on metabolic and hormonal disturbance of women with polycystic ovary syndrome. J. Res. Med. Sci. 2011, 16, 297–302. [Google Scholar] [PubMed]
- Jamilian, M.; Asemi, Z. The Effects of Soy Isoflavones on Metabolic Status of Patients With Polycystic Ovary Syndrome. J. Clin. Endocrinol. Metab. 2016, 101, 3386–3394. [Google Scholar] [CrossRef] [PubMed]
- Karamali, M.; Kashanian, M.; Alaeinasab, S.; Asemi, Z. The effect of dietary soy intake on weight loss, glycaemic control, lipid profiles and biomarkers of inflammation and oxidative stress in women with polycystic ovary syndrome: A randomised clinical trial. J. Hum. Nutr. Diet. 2018, 31, 533–543. [Google Scholar] [CrossRef]
- Romualdi, D.; Costantini, B.; Campagna, G.; Lanzone, A.; Guido, M. Is there a role for soy isoflavones in the therapeutic approach to polycystic ovary syndrome? Results from a pilot study. Fertil. Steril. 2008, 90, 1826–1833. [Google Scholar] [CrossRef]
- Haudum, C.; Lindheim, L.; Ascani, A.; Trummer, C.; Horvath, A.; Muenzker, J.; Obermayer-Pietsch, B. Impact of Short-Term Isoflavone Intervention in Polycystic Ovary Syndrome (PCOS) Patients on Microbiota Composition and Metagenomics. Nutrients 2020, 12, 1622. [Google Scholar] [CrossRef] [PubMed]
- Fontham, E.T.H.; Wolf, A.M.D.; Church, T.R.; Etzioni, R.; Flowers, C.R.; Herzig, A.; Guerra, C.E.; Oeffinger, K.C.; Shih, Y.C.T.; Walter, L.C.; et al. Cervical cancer screening for individuals at average risk: 2020 guideline update from the American Cancer Society. CA Cancer J. Clin. 2020, 70, 321–346. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Liu, J.; Pan, J.; Han, S.; Yin, X.; Wang, B.; Hu, G. Combined Treatment of Ionizing Radiation With Genistein on Cervical Cancer HeLa Cells. J. Pharmacol. Sci. 2006, 102, 129–135. [Google Scholar] [CrossRef] [PubMed]
- Yashar, C.M.; Spanos, W.J.; Taylor, D.D.; Gercel-Taylor, C. Potentiation of the radiation effect with genistein in cervical cancer cells. Gynecol. Oncol. 2005, 99, 199–205. [Google Scholar] [CrossRef] [PubMed]
- Sahin, K.; Tuzcu, M.; Basak, N.; Caglayan, B.; Kilic, U.; Sahin, F.; Kucuk, O. Sensitization of Cervical Cancer Cells to Cisplatin by Genistein: The Role of NFκB and Akt/mTOR Signaling Pathways. J. Oncol. 2012, 2012, 461562. [Google Scholar] [CrossRef]
- Nelson, H.D. Menopause. Lancet 2008, 371, 760–770. [Google Scholar] [CrossRef]
- Melby, M.K.; Lock, M.; Kaufert, P. Culture and symptom reporting at menopause. Hum. Reprod. Update 2005, 11, 495–512. [Google Scholar] [CrossRef]
- Gold, E.B. Factors Associated with Age at Natural Menopause in a Multiethnic Sample of Midlife Women. Am. J. Epidemiol. 2001, 153, 865–874. [Google Scholar] [CrossRef]
- Boulet, M.J.; Oddens, B.J.; Lehert, P.; Vemer, H.M.; Visser, A. Climacteric and menopause in seven south-east Asian countries. Maturitas 2008, 61, 34–53. [Google Scholar] [CrossRef]
- Mintziori, G.; Lambrinoudaki, I.; Goulis, D.G.; Ceausu, I.; Depypere, H.; Erel, C.T.; Perez-Lopez, F.R.; Schenck-Gustafsson, K.; Simoncini, T.; Tremollieres, F.; et al. EMAS position statement: Non-hormonal management of menopausal vasomotor symptoms. Maturitas 2015, 81, 410–413. [Google Scholar] [CrossRef]
- Crisafulli, A.; Marini, H.; Bitto, A.; Altavilla, D.; Squadrito, G.; Romeo, A.; Adamo, E.B.; Marini, R.; D’Anna, R.; Corrado, F.; et al. Effects of genistein on hot flushes in early postmenopausal women: A randomized, double-blind EPT- and placebo-controlled study. Menopause 2004, 11, 400–404. [Google Scholar] [CrossRef]
- Morabito, N.; Crisafulli, A.; Vergara, C.; Gaudio, A.; Lasco, A.; Frisina, N.; D’Anna, R.; Corrado, F.; Pizzoleo, M.A.; Cincotta, M.; et al. Effects of Genistein and Hormone-Replacement Therapy on Bone Loss in Early Postmenopausal Women: A Randomized Double-Blind Placebo-Controlled Study. J. Bone Miner. Res. 2002, 17, 1904–1912. [Google Scholar] [CrossRef] [PubMed]
- Squadrito, F.; Altavilla, D.; Morabito, N.; Crisafulli, A.; D’Anna, R.; Corrado, F.; Ruggeri, P.; Campo, G.M.; Calapai, G.; Caputi, A.P.; et al. The effect of the phytoestrogen genistein on plasma nitric oxide concentrations, endothelin-1 levels and endothelium dependent vasodilation in postmenopausal women. Atherosclerosis 2002, 163, 339–347. [Google Scholar] [CrossRef]
- Squadrito, F.; Altavilla, D.; Crisafulli, A.; Saitta, A.; Cucinotta, D.; Morabito, N.; D’Anna, R.; Corrado, F.; Ruggeri, P.; Frisina, N.; et al. Effect of genistein on endothelial function in postmenopausal women: A randomized, double-blind, controlled study. Am. J. Med. 2003, 114, 470–476. [Google Scholar] [CrossRef]
- Crisafulli, A.; Altavilla, D.; Marini, H.; Bitto, A.; Cucinotta, D.; Frisina, N.; Corrado, F.; D’Anna, R.; Squadrito, G.; Adamo, E.B.; et al. Effects of the phytoestrogen genistein on cardiovascular risk factors in postmenopausal women. Menopause 2005, 12, 186–192. [Google Scholar] [CrossRef] [PubMed]
- Franco, O.H.; Chowdhury, R.; Troup, J.; Voortman, T.; Kunutsor, S.; Kavousi, M.; Oliver-Williams, C.; Muka, T. Use of Plant-Based Therapies and Menopausal Symptoms: A Systematic Review and Meta-analysis. JAMA J. Am. Med. Assoc. 2016, 315, 2554–2563. [Google Scholar] [CrossRef] [PubMed]
- Hillman, G.G.; Wang, Y.; Che, M.; Raffoul, J.J.; Yudelev, M.; Kucuk, O.; Sarkar, F.H. Progression of renal cell carcinoma is inhibited by genistein and radiation in an orthotopic model. BMC Cancer 2007, 7, 4. [Google Scholar] [CrossRef] [PubMed]
- Akimoto, T.; Nonaka, T.; Ishikawa, H.; Sakurai, H.; Saitoh, J.-I.; Takahashi, T.; Mitsuhashi, N. Genistein, a tyrosine kinase inhibitor, enhanced radiosensitivity in human esophageal cancer cell lines in vitro: Possible involvement of inhibition of survival signal transduction pathways. Int. J. Radiat. Oncol. Biol. Phys. 2001, 50, 195–201. [Google Scholar] [CrossRef]
- Takaoka, O.; Mori, T.; Ito, F.; Okimura, H.; Kataoka, H.; Tanaka, Y.; Koshiba, A.; Kusuki, I.; Shigehiro, S.; Amami, T.; et al. Daidzein-rich isoflavone aglycones inhibit cell growth and inflammation in endometriosis. J. Steroid Biochem. Mol. Biol. 2018, 181, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Brahmbhatt, S.; Brahmbhatt, R.M.; Boyages, S.C. Thyroid ultrasound is the best prevalence indicator for assessment of iodine deficiency disorders: A study in rural/tribal schoolchildren from Gujarat (Western India). Eur. J. Endocrinol. 2000, 143, 37–46. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Amazon. Search: “Genistein Supplements”. Available online: https://www.amazon.com/s?k=genistein+supplements&crid=27YEIAKQM5YCZ&sprefix=Genistein+supple%2Caps%2C185&ref=nb_sb_ss_ts-da-p_1_16 (accessed on 17 December 2020).
- Vital Nutrients. Genistein 125 mg Supplement. Available online: https://www.vitalnutrients.net/genistein.html (accessed on 20 December 2020).
- WebMD. Genistein Combined Polysaccharide. Available online: https://www.webmd.com/vitamins/ai/ingredientmono-1088/genistein-combined-polysaccharide (accessed on 17 December 2020).
- Susan, R.; Loretta, P.M.; Patricia, B.H.; Charles, H.T.; Stephen, B.; Connie, M.W. A Longitudinal Study of the Effect of Genistein on Bone in Two Different Murine Models of Diminished Estrogen-Producing Capacity. J. Osteoporos. 2010, 2010, 145170. [Google Scholar] [CrossRef]
- Gilda, G.H.; Jeffrey, D.F.; Omer, K.; Mark, Y.; Richard, L.M.; Johanna, R.; Andrey, L.; Samuel, T.-M.; Judith, A.; Fazlul, H.S. Genistein Potentiates the Radiation Effect on Prostate Carcinoma Cells. Clin. Cancer Res. 2001, 7, 382–390. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Food | Mean Genistein Concentration a (mg Genistein/ 100 g Food) | Standard Deviation | References |
|---|---|---|---|
| Textured Soy Flour | 89.42 | 26.96 | [7,8,9,10,11,12,13,14,15] |
| Instant Beverage Soy Powder | 62.18 | 3.69 | [14,16,17,18,19,20] |
| Soy Protein Isolate | 57.28 | 14.17 | [7,14,16,19,21,22,23,24,25,26,27,28,29] |
| Meatless Bacon Bits | 45.77 | 0.11 | [13] |
| Kellog’s Smart-Start Soy Protein Cereal | 41.90 | N/A b (n < 3) | [13] |
| Natto | 37.66 | 7.85 | [30,31,32,33,34,35,36] |
| Uncooked Tempeh | 36.15 | 17.64 | [11,14,16,29,31,37,38,39] |
| Miso | 23.24 | 8.37 | [14,16,17,30,31,33,35,36,40,41,42,43] |
| Sprouted Raw Soybeans | 18.77 | 11.22 | [23,32,40,44,45,46,47,48,49] |
| Cooked Firm Tofu | 10.83 | 3.98 | [30,40,50] |
| Red Clovers | 10.00 | 0.00 | [51] |
| Worthington FriChik canned meatless chicken nuggets (prepared) | 9.35 | N/A (n < 3) | [31] |
| American Soy Cheese | 8.70 | N/A (n < 3) | [30] |
| Kellog’s Kashi Go-Lean Cereal | 7.70 | N/A (n < 3) | [13] |
| Chocolate Power Bar | 3.27 | N/A (n < 3) | [44] |
| Hoisin Sauce | 3.25 | N/A (n < 3) | [13] |
| Cake-Type Plain Doughnuts | 2.44 | 1.11 | [13,40] |
| Raw Pistachios | 1.75 | N/A (n < 3) | [40,52] |
| Reconstituted Infant Formula (Abbot Nutrition) | 1.37 | 0.37 | [53,54] |
| Cooked USDA Commodity Beef Patties | 1.09 | 0.42 | [31] |
| Fat Free Frankfurter Beef | 1.00 | N/A (n < 3) | [13] |
| Raw Chicken Breast Tenders | 0.25 | N/A (n < 3) | [13] |
| Raw White Grapefruit | 0.03 | N/A (n < 3) | [44] |
| Whole Raw Eggs | 0.02 | N/A (n < 3) | [44,45] |
| Mature Raw Black Beans | 0.00 | 0.00 | [44,55,56] |
| Population | Number of Subjects | Sample Type | Quantified Genistein | References | Year |
|---|---|---|---|---|---|
| Healthy infants in Pennsylvania, collected at the Children’s Hospital of Philadelphia and its affiliated clinics | Blood, urine, and saliva samples from cow- and breast-milk-fed infants | Large majority, except for cow’s milk-formula-fed infants, below LOD (<27 ng/mL in blood, <1.4 ng/mL in saliva, and <0.8 ng/mL in urine) | [57] | 2009 | |
| 165 | Urine (cow-formula-fed infants) | 13.6 ng/mL | |||
| Blood (soy-formula-fed infants) | 890.7 ng/mL (median) | ||||
| Urine (soy-formula-fed infants) | 7220 ng/mL (median) | ||||
| Saliva (soy-formula-fed infants) | 10.9 ng/mL (median) | ||||
| Cohort of women in Philadelphia, PA, USA | 451 | Daily consumption | 2.4–3.9 mg (average) | [58] | 2008 |
| Subgroup of larger cohort of women in Philadelphia, PA, USA | 27 | Daily urine excretion | 136.4 ng genistein/mg creatine (average) | ||
| Adult participants from Ireland, Italy, the Netherlands, and the UK | Daily consumption (Ireland) | 0.368 mg/day (average) | [59] | 2003 | |
| 7312 | Daily consumption (Italy) | 0.302 mg/day (average) | |||
| Daily consumption (the Netherlands) | 0.516 mg/day (average) | ||||
| Daily consumption (the UK) | 0.389 mg/day (average) | ||||
| Women of various racial and ethnic groups across the US | 1550 | Daily consumption (White women) | 3.6 μg genistein/day (average) | [60] | 2006 |
| 935 | Daily consumption (African American women) | 1.7 μg genistein/day (Average) | |||
| 286 | Daily consumption (Hispanic women) | 0 μg genistein/day (average) | |||
| 185 | Daily consumption (Chinese women) | 3534 μg genistein/day (average) | |||
| 195 | Daily consumption (Japanese women) | 6788 μg genistein/day (average) | |||
| Adults from various regions of Japan | 215 | Daily consumption | 14.5–18.3 mg genistein/day | [61] | 2001 |
| Serum level | 475.3 nmol genistein/liter of serum | ||||
| Daily excretion in urine | 14.2 μmol genistein/day | ||||
| Chinese men | 48 | Daily consumption | 19.4 ± 12.36 mg/day | [62] | 2007 |
| Adult (20–39 years old) women from the UK | 20 | Plasma genistein concentration of women that rarely consumed soy products | 14.3 nmol/L (geometric mean) | [63] | 2001 |
| 20 | Plasma genistein concentration of women that drank no soy milk but ate some solid soya foods | 16.5 nmol/L (Geometric mean) | |||
| 20 | Plasma genistein concentration of women that drank 0.25 pints of soy milk daily and ate some solid soya foods | 119 nmol/L (geometric mean) | |||
| 20 | Plasma genistein concentration of women that drank 0.5+ pints of soy milk daily and ate solid soya foods regularly | 378 nmol/L (geometric mean) |
| Category of Studies’ Conclusions Regarding Genistein/Whole Isoflavones/Genistein Metabolites | Total Number of Studies (Number Included in the Exposure Testing Range/Daily Dosage Column In Vivo) a | Exposure Testing Range In Vitro (µM) | Daily Dosage Testing Range In Vivo (mg) | References |
|---|---|---|---|---|
| Evidence suggests effects are primarily beneficial | 42 (27) | Genistein: 2.0–370 | Genistein: 36–600 (all doses above 54 were in one study) Genistein Mode: 54 (7 studies) Whole Isoflavones: 40–165 Soy Intakes/Week: 0.76–12.0 | [67,68,95,108,109,110,111,112,120,121,125,132,143,144,145,146,147,149,150,152,161,171,172,173,175,176,178,179,180,182,184,185,186,192,193,194,195,196,197,198,199,200] |
| Evidence suggests effects are debated/inconclusive, but does suggest potential benefits | 11 (9) | Genistein: 0.0037–185 | Genistein: 30–54 Whole Isoflavones: 45 | [108,114,119,128,129,151,154,155,163,174,181] |
| Evidence suggests effects are debated/inconclusive, and does not show any potential for benefits | 4 (2) | Genistein: 1–10 | Whole Isoflavones: 33.3–300 | [148,153,159,162] |
| Evidence suggests effects are primarily detrimental | 5 (5) | Genistein: 0.001–3.7 | N/A b | [127,133,157,158,201] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, L.; Rios, E.; Castro, L.; Liu, J.; Yan, Y.; Dixon, D. Genistein: Dual Role in Women’s Health. Nutrients 2021, 13, 3048. https://doi.org/10.3390/nu13093048
Yu L, Rios E, Castro L, Liu J, Yan Y, Dixon D. Genistein: Dual Role in Women’s Health. Nutrients. 2021; 13(9):3048. https://doi.org/10.3390/nu13093048
Chicago/Turabian StyleYu, Linda, Eddy Rios, Lysandra Castro, Jingli Liu, Yitang Yan, and Darlene Dixon. 2021. "Genistein: Dual Role in Women’s Health" Nutrients 13, no. 9: 3048. https://doi.org/10.3390/nu13093048
APA StyleYu, L., Rios, E., Castro, L., Liu, J., Yan, Y., & Dixon, D. (2021). Genistein: Dual Role in Women’s Health. Nutrients, 13(9), 3048. https://doi.org/10.3390/nu13093048