
Consequences of Paternal Nutrition on Offspring Health and Disease



Abstract
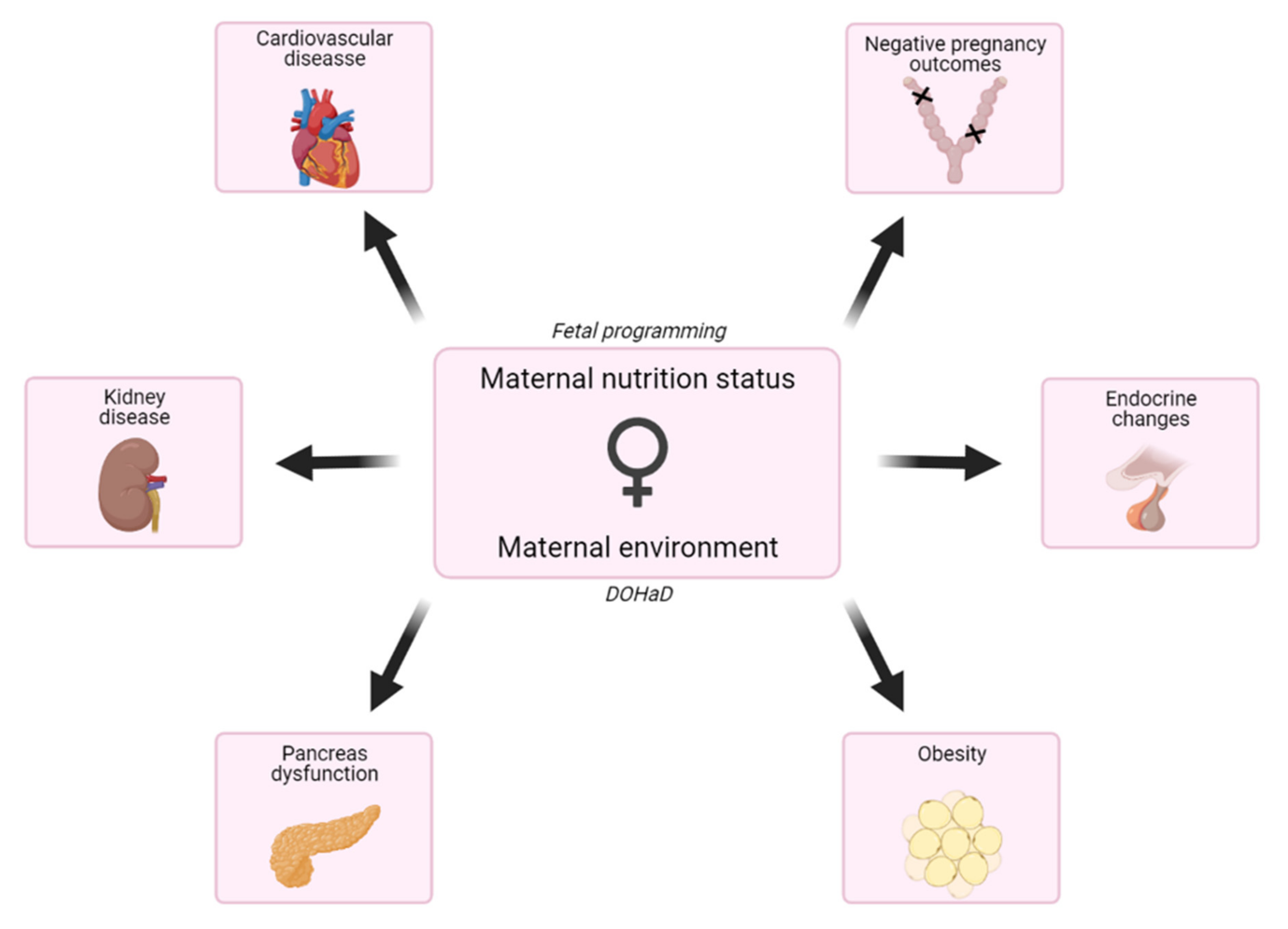
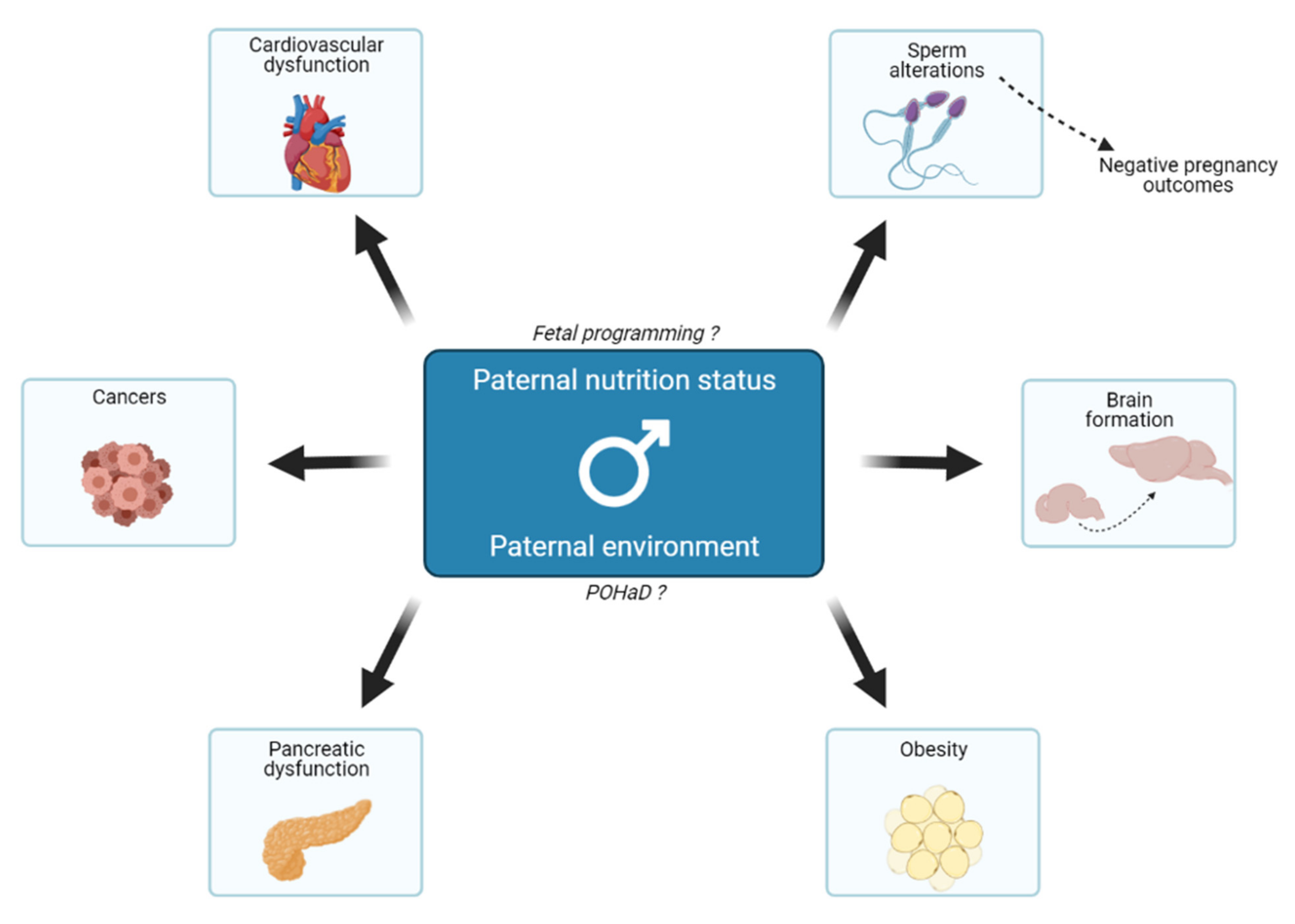
:1. Introduction
2. Epigenetic Mechanisms Involved in the Transgenerational Transmission
3. Impact of Paternal Folic Acid Intake on Progeny’s Health
3.1. Paternal Folate Deficiency
3.2. Paternal Folate Supplementation
4. Depleted Paternal Diets and Their Effects on the Progeny
4.1. Impact of a Paternal Low-Protein Diet
4.2. What about Paternal Food Restrictions?
5. Effects of Enriched Paternal Diets on Offspring
5.1. Consequences of a Paternal High-Fat Diet
5.2. Impacts of a Paternal High-Sugar Diet
6. Conclusions and Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| DOHaD | developmental origins of health and diseases |
| HFD | high-fat diet |
| LPD | low protein diet |
| NTD | neural tube defect |
| PND | postnatal day |
| POHaD | paternal origins of health and disease |
| SAM | S-adenosyl methionine |
| sncRNAs | small non-coding RNAs |
| WHO | world health organization |
References
- Hales, C.N.; Barker, D.J.P. The thrifty phenotype hypothesis. Br. Med. Bull. 2001, 60, 5–20. [Google Scholar] [CrossRef] [Green Version]
- Barouki, R.; Gluckman, P.D.; Grandjean, P.; Hanson, M.; Heindel, J.J. Developmental origins of non-communicable disease: Implications for research and public health. Environ. Health 2012, 11, 42. [Google Scholar] [CrossRef] [Green Version]
- Marangoni, F.; Cetin, I.; Verduci, E.; Canzone, G.; Giovannini, M.; Scollo, P.; Corsello, G.; Poli, A. Maternal Diet and Nutrient Requirements in Pregnancy and Breastfeeding. An Italian Consensus Document. Nutrients 2016, 8, 629. [Google Scholar] [CrossRef] [PubMed]
- De-Regil, L.M.; Fernández-Gaxiola, A.C.; Dowswell, T.; Peña-Rosas, J.P. Effects and safety of periconceptional folate supplementation for preventing birth defects. In Cochrane Database of Systematic Reviews; The Cochrane Collaboration, Ed.; John Wiley & Sons, Ltd.: Chichester, UK, 2010; p. CD007950.pub2. [Google Scholar]
- Crott, J.W. Effects of altered parental folate and one-carbon nutrient status on offspring growth and metabolism. Mol. Asp. Med. 2017, 53, 28–35. [Google Scholar] [CrossRef] [Green Version]
- El Hajj Chehadeh, S.E.; Pourié, G.; Martin, N.; Alberto, J.-M.; Daval, J.-L.; Guéant, J.-L.; Leininger-Muller, B. Gestational methyl donor deficiency alters key proteins involved in neurosteroidogenesis in the olfactory bulbs of newborn female rats and is associated with impaired olfactory performance. Br. J. Nutr. 2014, 111, 1021–1031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El Hajj Chehadeh, S.; Dreumont, N.; Willekens, J.; Canabady-Rochelle, L.; Jeannesson, E.; Alberto, J.-M.; Daval, J.-L.; Guéant, J.-L.; Leininger-Muller, B. Early methyl donor deficiency alters cAMP signaling pathway and neurosteroidogenesis in the cerebellum of female rat pups. Am. J. Physiol. Metab. 2014, 307, E1009–E1019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willekens, J.; Hergalant, S.; Pourié, G.; Marin, F.; Alberto, J.-M.; Georges, L.; Paoli, J.; Nemos, C.; Daval, J.-L.; Guéant, J.-L.; et al. Wnt Signaling Pathways Are Dysregulated in Rat Female Cerebellum Following Early Methyl Donor Deficiency. Mol. Neurobiol. 2019, 56, 892–906. [Google Scholar] [CrossRef]
- Kramer, M.S.; Kakuma, R. Energy and protein intake in pregnancy. In Cochrane Database of Systematic Reviews; The Cochrane Collaboration, Ed.; John Wiley & Sons, Ltd.: Chichester, UK, 2003; p. CD000032. [Google Scholar]
- Koletzko, B.; Cetin, I.; Thomas Brenna, J.; for the Perinatal Lipid Intake Working Group. Dietary fat intakes for pregnant and lactating women. Br. J. Nutr. 2007, 98, 873–877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koletzko, B.; Bauer, C.P.; Bung, P.; Cremer, M.; Flothkötter, M.; Hellmers, C.; Kersting, M.; Krawinkel, M.; Przyrembel, H.; Rasenack, R.; et al. German National Consensus Recommendations on Nutrition and Lifestyle in Pregnancy by the ‘Healthy Start–Young Family Network’. Ann. Nutr. Metab. 2013, 63, 311–322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trumbo, P.; Schlicker, S.; Yates, A.A.; Poos, M. Dietary Reference Intakes for Energy, Carbohydrate, Fiber, Fat, Fatty Acids, Cholesterol, Protein and Amino Acids. J. Am. Diet. Assoc. 2002, 102, 1621–1630. [Google Scholar] [CrossRef]
- Soubry, A. POHaD: Why we should study future fathers. Environ. Epigenet. 2018, 4, dvy007. [Google Scholar] [CrossRef]
- Tirpák, F.; Greifová, H.; Lukáč, N.; Stawarz, R.; Massányi, P. Exogenous Factors Affecting the Functional Integrity of Male Reproduction. Life 2021, 11, 213. [Google Scholar] [CrossRef] [PubMed]
- Zegers-Hochschild, F.; Adamson, G.D.; Dyer, S.; Racowsky, C.; de Mouzon, J.; Sokol, R.; Rienzi, L.; Sunde, A.; Schmidt, L.; Cooke, I.D.; et al. The International Glossary on Infertility and Fertility Care. Fertil. Steril. 2017, 108, 393–406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marcho, C.; Oluwayiose, O.A.; Pilsner, J.R. The preconception environment and sperm epigenetics. Andrology 2020, 8, 924–942. [Google Scholar] [CrossRef]
- Nan, Y.; Yi, S.-J.; Zhu, H.-L.; Xiong, Y.-W.; Shi, X.-T.; Cao, X.-L.; Zhang, C.; Gao, L.; Zhao, L.-L.; Zhang, J.; et al. Paternal cadmium exposure increases the susceptibility to diet-induced testicular injury and spermatogenic disorders in mouse offspring. Chemosphere 2020, 246, 125776. [Google Scholar] [CrossRef]
- Messerlian, C.; Bellinger, D.; Mínguez-Alarcón, L.; Romano, M.E.; Ford, J.B.; Williams, P.L.; Calafat, A.M.; Hauser, R.; Braun, J.M. Paternal and maternal preconception urinary phthalate metabolite concentrations and child behavior. Environ. Res. 2017, 158, 720–728. [Google Scholar] [CrossRef]
- Barakat, R.; Lin, P.-C.; Park, C.J.; Zeineldin, M.; Zhou, S.; Rattan, S.; Brehm, E.; Flaws, J.A.; Ko, C.J. Germline-dependent transmission of male reproductive traits induced by an endocrine disruptor, di-2-ethylhexyl phthalate, in future generations. Sci. Rep. 2020, 10, 5705–5718. [Google Scholar] [CrossRef]
- Silva, P.V.; Dos Santos Borges, C.; Rosa, J.D.L.; Pacheco, T.L.; Figueiredo, T.M.; Leite, G.A.A.; Guerra, M.T.; Anselmo-Franci, J.A.; Klinefelter, G.R.; Kempinas, W.D.G. Effects of isolated or combined exposure to sibutramine and rosuvastatin on reproductive parameters of adult male rats. J. Appl. Toxicol. 2020, 40, 947–964. [Google Scholar] [CrossRef] [PubMed]
- McPherson, N.O.; Fullston, T.; Kang, W.X.; Sandeman, L.Y.; Corbett, M.A.; Owens, J.A.; Lane, M. Paternal under-nutrition programs metabolic syndrome in offspring which can be reversed by antioxidant/vitamin food fortification in fathers. Sci. Rep. 2016, 6, 27010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sansone, A.; Di Dato, C.; De Angelis, C.; Menafra, D.; Pozza, C.; Pivonello, R.; Isidori, A.; Gianfrilli, D. Smoke, alcohol and drug addiction and male fertility. Reprod. Biol. Endocrinol. 2018, 16, 3. [Google Scholar] [CrossRef] [Green Version]
- Day, J.; Savani, S.; Krempley, B.D.; Nguyen, M.; Kitlinska, J.B. Influence of paternal preconception exposures on their offspring: Through epigenetics to phenotype. Am. J. Stem Cells 2016, 5, 11–18. [Google Scholar]
- Carone, B.R.; Fauquier, L.; Habib, N.; Shea, J.M.; Hart, C.E.; Li, R.; Bock, C.; Li, C.; Gu, H.; Zamore, P.D.; et al. Paternally Induced Transgenerational Environmental Reprogramming of Metabolic Gene Expression in Mammals. Cell 2010, 143, 1084–1096. [Google Scholar] [CrossRef] [Green Version]
- Sharma, U.; Conine, C.C.; Shea, J.M.; Boskovic, A.; Derr, A.G.; Bing, X.Y.; Belleannee, C.; Kucukural, A.; Serra, R.W.; Sun, F.; et al. Biogenesis and function of tRNA fragments during sperm maturation and fertilization in mammals. Science 2016, 351, 391–396. [Google Scholar] [CrossRef] [Green Version]
- Sadler-Riggleman, I.; Klukovich, R.; Nilsson, E.; Beck, D.; Xie, Y.; Yan, W.; Skinner, M.K. Epigenetic transgenerational inheritance of testis pathology and Sertoli cell epimutations: Generational origins of male infertility. Environ. Epigenet. 2019, 5, dvz013. [Google Scholar] [CrossRef] [PubMed]
- Dupont, C.; Kappeler, L.; Saget, S.; Grandjean, V.; Lévy, R. Role of miRNA in the Transmission of Metabolic Diseases Associated With Paternal Diet-Induced Obesity. Front. Genet. 2019, 10, 337. [Google Scholar] [CrossRef] [PubMed]
- Klastrup, L.K.; Bak, S.T.; Nielsen, A.L. The influence of paternal diet on sncRNA-mediated epigenetic inheritance. Mol. Genet. Genom. 2019, 294, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Sharma, U. Paternal Contributions to Offspring Health: Role of Sperm Small RNAs in Intergenerational Transmission of Epigenetic Information. Front. Cell Dev. Biol. 2019, 7, 215. [Google Scholar] [CrossRef] [PubMed]
- Painter, R.; Osmond, C.; Gluckman, P.; Hanson, M.; Phillips, D.; Roseboom, T. Transgenerational effects of prenatal exposure to the Dutch famine on neonatal adiposity and health in later life. BJOG Int. J. Obstet. Gynaecol. 2008, 115, 1243–1249. [Google Scholar] [CrossRef] [PubMed]
- Veenendaal, M.; Painter, R.; De Rooij, S.R.; Bossuyt, P.; van der Post, J.; Gluckman, P.; Hanson, M.; Roseboom, T. Transgenerational effects of prenatal exposure to the 1944-45 Dutch famine. BJOG: Int. J. Obstet. Gynaecol. 2013, 120, 548–554. [Google Scholar] [CrossRef] [PubMed]
- Kaati, G.; Bygren, L.; Edvinsson, S. Cardiovascular and diabetes mortality determined by nutrition during parents’ and grandparents’ slow growth period. Eur. J. Hum. Genet. 2002, 10, 682–688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- The ALSPAC Study Team; Pembrey, M.; Bygren, L.O.; Kaati, G.; Edvinsson, S.; Northstone, K.; Sjöström, M.; Golding, J. Sex-specific, male-line transgenerational responses in humans. Eur. J. Hum. Genet. 2005, 14, 159–166. [Google Scholar] [CrossRef]
- Vågerö, D.; Pinger, P.R.; Aronsson, V.; Berg, G.J.V.D. Paternal grandfather’s access to food predicts all-cause and cancer mortality in grandsons. Nat. Commun. 2018, 9, 5124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bygren, L.O.; Kaati, G.; Edvinsson, S. Longevity Determined by Paternal Ancestors’ Nutrition during Their Slow Growth Period. Acta Biotheor. 2001, 49, 53–59. [Google Scholar] [CrossRef] [PubMed]
- McCoy, C.R.; Jackson, N.L.; Brewer, R.L.; Moughnyeh, M.M.; Smith, D.L.; Clinton, S.M. A paternal methyl donor depleted diet leads to increased anxiety- and depression-like behavior in adult rat offspring. Biosci. Rep. 2018, 38, BSR20180730. [Google Scholar] [CrossRef] [Green Version]
- Watkins, A.J.; Sinclair, K.D. Paternal low protein diet affects adult offspring cardiovascular and metabolic function in mice. Am. J. Physiol. Circ. Physiol. 2014, 306, H1444–H1452. [Google Scholar] [CrossRef] [Green Version]
- Watkins, A.J.; Dias, I.; Tsuro, H.; Allen, D.; Emes, R.D.; Moreton, J.; Wilson, R.; Ingram, R.J.M.; Sinclair, K.D. Paternal diet programs offspring health through sperm- and seminal plasma-specific pathways in mice. Proc. Natl. Acad. Sci. USA 2018, 115, 10064–10069. [Google Scholar] [CrossRef] [Green Version]
- de Castro Barbosa, T.; Ingerslev, L.R.; Alm, P.S.; Versteyhe, S.; Massart, J.; Rasmussen, M.; Donkin, I.; Sjögren, R.; Mudry, J.M.; Vetterli, L.; et al. High-Fat Diet Reprograms the Epigenome of Rat Spermatozoa and Transgenerationally Affects Metabolism of the Offspring. Mol. Metab. 2016, 5, 184–197. [Google Scholar] [CrossRef]
- Grandjean, V.; Fourré, S.; De Abreu, D.A.F.; Derieppe, M.-A.; Remy, J.-J.; Rassoulzadegan, M. RNA-mediated paternal heredity of diet-induced obesity and metabolic disorders. Sci. Rep. 2016, 5, 18193. [Google Scholar] [CrossRef] [Green Version]
- Ghanayem, B.I.; Bai, R.; Kissling, G.E.; Travlos, G.; Hoffler, U. Diet-Induced Obesity in Male Mice Is Associated with Reduced Fertility and Potentiation of Acrylamide-Induced Reproductive Toxicity. Biol. Reprod. 2010, 82, 96–104. [Google Scholar] [CrossRef] [Green Version]
- Binder, N.K.; Mitchell, M.; Gardner, D.K. Parental diet-induced obesity leads to retarded early mouse embryo development and altered carbohydrate utilisation by the blastocyst. Reprod. Fertil. Dev. 2012, 24, 804–812. [Google Scholar] [CrossRef]
- Lin, W.-T.; Huang, H.-L.; Huang, M.-C.; Chan, T.-F.; Ciou, S.-Y.; Lee, C.-Y.; Chiu, Y.-W.; Duh, T.-H.; Lin, P.-L.; Wang, T.-N.; et al. Effects on uric acid, body mass index and blood pressure in adolescents of consuming beverages sweetened with high-fructose corn syrup. Int. J. Obes. (Lond.) 2013, 37, 532–539. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.; Guo, W.; Yan, T.; Zhou, J.; Li, Y.; Zhu, Y.; Duan, Y.; Yang, X. Spermatozoal mRNAs expression implicated in embryonic development were influenced by dietary folate supplementation of breeder roosters by altering spermatozoal piRNA expression profiles. Theriogenology 2019, 138, 102–110. [Google Scholar] [CrossRef]
- Ng, S.-F.; Lin, R.C.Y.; Laybutt, D.R.; Barres, R.; Owens, J.A.; Morris, M.J. Chronic high-fat diet in fathers programs β-cell dysfunction in female rat offspring. Nature 2010, 467, 963–966. [Google Scholar] [CrossRef] [PubMed]
- Carapeto, P.V.; Ornellas, F.; Mandarim-De-Lacerda, C.A.; Aguila, M.B. Liver metabolism in adult male mice offspring: Consequences of a maternal, paternal or both maternal and paternal high-fructose diet. J. Dev. Orig. Health Dis. 2018, 9, 450–459. [Google Scholar] [CrossRef] [PubMed]
- Ornellas, F.; Carapeto, P.V.; Aguila, M.B.; Mandarim-De-Lacerda, C.A. Sex-linked changes and high cardiovascular risk markers in the mature progeny of father, mother, or both father and mother consuming a high-fructose diet. Nutrition 2020, 71, 110612. [Google Scholar] [CrossRef] [PubMed]
- Binder, N.K.; Hannan, N.J.; Gardner, D.K. Paternal Diet-Induced Obesity Retards Early Mouse Embryo Development, Mitochondrial Activity and Pregnancy Health. PLoS ONE 2012, 7, e52304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lambrot, R.; Xu, C.; Saintphar, S.; Chountalos, G.; Cohen, T.; Paquet, M.; Suderman, M.; Hallett, M.; Kimmins, S. Low paternal dietary folate alters the mouse sperm epigenome and is associated with negative pregnancy outcomes. Nat. Commun. 2013, 4, 2889. [Google Scholar] [CrossRef] [PubMed]
- Watkins, A.J.; Sirovica, S.; Stokes, B.; Isaacs, M.; Addison, O.; Martin, R. Paternal low protein diet programs preimplantation embryo gene expression, fetal growth and skeletal development in mice. Biochim. Biophys. Acta (BBA)—Mol. Basis Dis. 2017, 1863, 1371–1381. [Google Scholar] [CrossRef]
- Ly, L.; Chan, D.; Aarabi, M.; Landry, M.; Behan, N.A.; Macfarlane, A.J.; Trasler, J. Intergenerational impact of paternal lifetime exposures to both folic acid deficiency and supplementation on reproductive outcomes and imprinted gene methylation. MHR Basic Sci. Reprod. Med. 2017, 23, 461–477. [Google Scholar] [CrossRef]
- Martín-Calvo, N.; Mínguez-Alarcón, L.; Gaskins, A.J.; Nassan, F.L.; Williams, P.L.; Souter, I.; Hauser, R.; Chavarro, J.E. Paternal preconception folate intake in relation to gestational age at delivery and birthweight of newborns conceived through assisted reproduction. Reprod. Biomed. Online 2019, 39, 835–843. [Google Scholar] [CrossRef]
- Kim, H.W.; Choi, Y.J.; Kim, K.N.; Tamura, T.; Chang, N. Effect of paternal folate deficiency on placental folate content and folate receptor α expression in rats. Nutr. Res. Pract. 2011, 5, 112–116. [Google Scholar] [CrossRef] [Green Version]
- Da Cruz, R.S.; Carney, E.J.; Clarke, J.; Cao, H.; Cruz, M.I.; Benitez, C.; Jin, L.; Fu, Y.; Cheng, Z.; Wang, Y.; et al. Paternal malnutrition programs breast cancer risk and tumor metabolism in offspring. Breast Cancer Res. 2018, 20, 99. [Google Scholar] [CrossRef] [Green Version]
- Bjornstad, P.; Laffel, L.; Lynch, J.; El Ghormli, L.; Weinstock, R.S.; Tollefsen, S.E.; Nadeau, K.J. Elevated Serum Uric Acid Is Associated With Greater Risk for Hypertension and Diabetic Kidney Diseases in Obese Adolescents with Type 2 Diabetes: An Observational Analysis From the Treatment Options for Type 2 Diabetes in Adolescents and Youth (TODAY) Study. Diabetes Care 2019, 42, 1120–1128. [Google Scholar] [CrossRef]
- Yuan, H.-F.; Zhao, K.; Zang, Y.; Liu, C.-Y.; Hu, Z.-Y.; Wei, J.-J.; Zhou, T.; Li, Y.; Zhang, H.-P. Effect of folate deficiency on promoter methylation and gene expression of Esr1, Cav1, and Elavl1, and its influence on spermatogenesis. Oncotarget 2017, 8, 24130–24141. [Google Scholar] [CrossRef] [Green Version]
- Swayne, B.G.; Kawata, A.; Behan, N.A.; Williams, A.; Wade, M.G.; MacFarlane, A.J.; Yauk, C.L. Investigating the effects of dietary folic acid on sperm count, DNA damage and mutation in Balb/c mice. Mutat. Res. Mol. Mech. Mutagen. 2012, 737, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Tomizawa, H.; Matsuzawa, D.; Ishii, D.; Matsuda, S.; Kawai, K.; Mashimo, Y.; Sutoh, C.; Shimizu, E. Methyl-Donor Deficiency in Adolescence Affects Memory and Epigenetic Status in the Mouse Hippocampus: Methyl-Donor Deficiency in Adolescence. Genes Brain Behav. 2015, 14, 301–309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, H.; Hauser, R.; Krawetz, S.A.; Pilsner, J.R. Environmental Susceptibility of the Sperm Epigenome During Windows of Male Germ Cell Development. Curr. Environ. Health Rep. 2015, 2, 356–366. [Google Scholar] [CrossRef] [Green Version]
- Rando, O.J. Daddy Issues: Paternal Effects on Phenotype. Cell 2012, 151, 702–708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nilsson, E.E.; Sadler-Riggleman, I.; Skinner, M.K. Environmentally induced epigenetic transgenerational inheritance of disease. Environ. Epigenet. 2018, 4, dvy016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guéant, J.-L.; Namour, F.; Guéant-Rodriguez, R.-M.; Daval, J.-L. Folate and fetal programming: A play in epigenomics? Trends Endocrinol. Metab. 2013, 24, 279–289. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, K.; Maekawa, T.; Ly, N.H.; Fujita, S.-I.; Muratani, M.; Ando, M.; Katou, Y.; Araki, H.; Miura, F.; Shirahige, K.; et al. ATF7-Dependent Epigenetic Changes Are Required for the Intergenerational Effect of a Paternal Low-Protein Diet. Mol. Cell 2020, 78, 445–458.e6. [Google Scholar] [CrossRef] [PubMed]
- Fullston, T.; Teague, E.M.C.O.; Palmer, N.O.; DeBlasio, M.J.; Mitchell, M.; Corbett, M.; Print, C.G.; Owens, J.A.; Lane, M. Paternal obesity initiates metabolic disturbances in two generations of mice with incomplete penetrance to the F 2 generation and alters the transcriptional profile of testis and sperm microRNA content. FASEB J. 2013, 27, 4226–4243. [Google Scholar] [CrossRef]
- Fullston, T.; Ohlsson-Teague, E.M.C.; Print, C.G.; Sandeman, L.Y.; Lane, M. Sperm microRNA Content Is Altered in a Mouse Model of Male Obesity, but the Same Suite of microRNAs Are Not Altered in Offspring’s Sperm. PLoS ONE 2016, 11, e0166076. [Google Scholar] [CrossRef] [Green Version]
- Claycombe-Larson, K.G.; Bundy, A.N.; Roemmich, J.N. Paternal high-fat diet and exercise regulate sperm miRNA and histone methylation to modify placental inflammation, nutrient transporter mRNA expression and fetal weight in a sex-dependent manner. J. Nutr. Biochem. 2020, 81, 108373. [Google Scholar] [CrossRef]
- Teltumbade, M.; Bhalla, A.; Sharma, A. Paternal inheritance of diet induced metabolic traits correlates with germline regulation of diet induced coding gene expression. Genomics 2020, 112, 567–573. [Google Scholar] [CrossRef]
- Wu, S.; Guo, W.; Li, X.; Liu, Y.; Li, Y.; Lei, X.; Yao, J.; Yang, X. Paternal chronic folate supplementation induced the transgenerational inheritance of acquired developmental and metabolic changes in chickens. Proc. R. Soc. B 2019, 286, 20191653. [Google Scholar] [CrossRef]
- Nätt, D.; Kugelberg, U.; Casas, E.; Nedstrand, E.; Zalavary, S.; Henriksson, P.; Nijm, C.; Jäderquist, J.; Sandborg, J.; Flinke, E.; et al. Human sperm displays rapid responses to diet. PLoS Biol. 2019, 17, e3000559. [Google Scholar] [CrossRef] [Green Version]
- Shea, J.M.; Serra, R.W.; Carone, B.R.; Shulha, H.P.; Kucukural, A.; Ziller, M.J.; Vallaster, M.P.; Gu, H.; Tapper, A.R.; Gardner, P.D.; et al. Genetic and Epigenetic Variation, but Not Diet, Shape the Sperm Methylome. Dev. Cell 2015, 35, 750–758. [Google Scholar] [CrossRef] [Green Version]
- Radford, E.J.; Ito, M.; Shi, H.; Corish, J.A.; Yamazawa, K.; Isganaitis, E.; Seisenberger, S.; Hore, T.A.; Reik, W.; Erkek, S.; et al. In utero undernourishment perturbs the adult sperm methylome and intergenerational metabolism. Science 2014, 345, 1255903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holland, M.L.; Lowe, R.; Caton, P.W.; Gemma, C.; Carbajosa, G.; Danson, A.F.; Carpenter, A.A.M.; Loche, E.; Ozanne, S.E.; Rakyan, V.K. Early-life nutrition modulates the epigenetic state of specific rDNA genetic variants in mice. Science 2016, 353, 495–498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, U.; Rando, O.J. Metabolic Inputs into the Epigenome. Cell Metab. 2017, 25, 544–558. [Google Scholar] [CrossRef]
- Chen, Q.; Yan, M.; Cao, Z.; Li, X.; Zhang, Y.; Shi, J.; Feng, G.-H.; Peng, H.; Zhang, X.; Qian, J.; et al. Sperm tsRNAs contribute to intergenerational inheritance of an acquired metabolic disorder. Science 2016, 351, 397–400. [Google Scholar] [CrossRef] [Green Version]
- Gatewood, J.M.; Cook, G.R.; Balhorn, R.; Bradbury, E.M.; Schmid, C.W.; Sugimura, T. Sequence-specific packaging of DNA in human sperm chromatin. Science 1987, 236, 962–964. [Google Scholar] [CrossRef] [PubMed]
- Bagherzadeh Khodashahri, S.; Siadat, S.D.; Rahbar, M.; Abdollahpour-Alitappeh, M.; Vaziri, F.; Rahna-maye-Farzami, M.; Mohammadzadeh, M.; Davari, M.; Fateh, A.; Masoumi, M. Genotyping of Haemophilus Influenzae Type b Strains and Their Incidence in the Clinical Samples Isolated from Iranian Patients. Iran. J. Microbiol. 2015, 7, 136–143. [Google Scholar] [PubMed]
- Terashima, M.; Barbour, S.; Ren, J.; Yu, W.; Han, Y.; Muegge, K. Effect of high fat diet on paternal sperm histone distribution and male offspring liver gene expression. Epigenetics 2015, 10, 861–871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Öst, A.; Lempradl, A.; Casas, E.; Weigert, M.; Tiko, T.; Deniz, M.; Pantano, L.; Boenisch, U.; Itskov, P.M.; Stoeckius, M.; et al. Paternal Diet Defines Offspring Chromatin State and Intergenerational Obesity. Cell 2014, 159, 1352–1364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siklenka, K.; Erkek, S.; Godmann, M.; Lambrot, R.; McGraw, S.; Lafleur, C.; Cohen, T.; Xia, J.; Suderman, M.; Hallett, M.; et al. Disruption of histone methylation in developing sperm impairs offspring health transgenerationally. Science 2015, 350, aab2006. [Google Scholar] [CrossRef]
- Chambers, T.J.G.; Morgan, M.D.; Heger, A.H.; Sharpe, R.M.; Drake, A.J. High-fat diet disrupts metabolism in two generations of rats in a parent-of-origin specific manner. Sci. Rep. 2016, 6, 31857. [Google Scholar] [CrossRef] [Green Version]
- Fullston, T.; Palmer, N.O.; Owens, J.A.; Mitchell, M.; Bakos, H.W.; Lane, M. Diet-induced paternal obesity in the absence of diabetes diminishes the reproductive health of two subsequent generations of mice. Hum. Reprod. 2012, 27, 1391–1400. [Google Scholar] [CrossRef] [Green Version]
- Forges, T.; Monnier-Barbarino, P.; Alberto, J.M.; Guéant-Rodriguez, R.M.; Daval, J.L.; Guéant, J.L. Impact of folate and homocysteine metabolism on human reproductive health. Hum. Reprod. Updat. 2007, 13, 225–238. [Google Scholar] [CrossRef]
- Kim, H.W.; Kim, K.N.; Choi, Y.J.; Chang, N. Effects of paternal folate deficiency on the expression of insulin-like growth factor-2 and global DNA methylation in the fetal brain. Mol. Nutr. Food Res. 2013, 57, 671–676. [Google Scholar] [CrossRef]
- Mejos, K.K.; Kim, H.W.; Lim, E.M.; Chang, N. Effects of parental folate deficiency on the folate content, global DNA methylation, and expressions of FRα, IGF-2 and IGF-1R in the postnatal rat liver. Nutr. Res. Pract. 2013, 7, 281–286. [Google Scholar] [CrossRef]
- Ryan, D.P.; Henzel, K.S.; Pearson, B.L.; Siwek, M.E.; Papazoglou, A.; Guo, L.; Paesler, K.; Yu, M.; Müller, R.; Xie, K.; et al. A paternal methyl donor-rich diet altered cognitive and neural functions in offspring mice. Mol. Psychiatry 2018, 23, 1345–1355. [Google Scholar] [CrossRef] [Green Version]
- Roseboom, T.J.; Painter, R.C.; van Abeelen, A.F.M.; Veenendaal, M.V.E.; de Rooij, S.R. Hungry in the womb: What are the consequences? Lessons from the Dutch famine. Maturitas 2011, 70, 141–145. [Google Scholar] [CrossRef] [PubMed]
- Pigeyre, M.; Yazdi, F.T.; Kaur, Y.; Meyre, D. Recent progress in genetics, epigenetics and metagenomics unveils the pathophysiology of human obesity. Clin. Sci. 2016, 130, 943–986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reddon, H.; Guéant, J.-L.; Meyre, D. The importance of gene–environment interactions in human obesity. Clin. Sci. 2016, 130, 1571–1597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morgan, H.L.; Ampong, I.; Eid, N.; Rouillon, C.; Griffiths, H.R.; Watkins, A.J. Low protein diet and methyl-donor supplements modify testicular physiology in mice. Reproduction 2020, 159, 627–641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ly, N.H.; Maekawa, T.; Yoshida, K.; Liu, Y.; Muratani, M.; Ishii, S. RNA-Sequencing Analysis of Paternal Low-Protein Diet-Induced Gene Expression Change in Mouse Offspring Adipocytes. G3 Genes|Genomes|Genet. 2019, 9, 2161–2170. [Google Scholar] [CrossRef] [Green Version]
- Anderson, L.M.; Riffle, L.; Wilson, R.; Travlos, G.S.; Lubomirski, M.S.; Alvord, W.G. Preconceptional fasting of fathers alters serum glucose in offspring of mice. Nutrition 2006, 22, 327–331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miersch, C.; Döring, F. Paternal dietary restriction affects progeny fat content in Caenorhabditis elegans. IUBMB Life 2012, 64, 644–648. [Google Scholar] [CrossRef]
- Corley, B.T.; Carroll, R.W.; Hall, R.M.; Weatherall, M.; Parry-Strong, A.; Krebs, J.D. Intermittent fasting in Type 2 diabetes mellitus and the risk of hypoglycaemia: A randomized controlled trial. Diabet. Med. 2018, 35, 588–594. [Google Scholar] [CrossRef] [PubMed]
- Baumeier, C.; Kaiser, D.; Heeren, J.; Scheja, L.; John, C.; Weise, C.; Eravci, M.; Lagerpusch, M.; Schulze, G.; Joost, H.-G.; et al. Caloric restriction and intermittent fasting alter hepatic lipid droplet proteome and diacylglycerol species and prevent diabetes in NZO mice. Biochim. Biophys. Acta (BBA)—Mol. Cell Biol. Lipids 2015, 1851, 566–576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Souza Marinho, T.D.; Ornellas, F.; Barbosa-Da-Silva, S.; Mandarim-De-Lacerda, C.A.; Aguila, M.B. Beneficial effects of intermittent fasting on steatosis and inflammation of the liver in mice fed a high-fat or a high-fructose diet. Nutrition 2019, 65, 103–112. [Google Scholar] [CrossRef]
- Mattson, M.P.; Longo, V.D.; Harvie, M. Impact of intermittent fasting on health and disease processes. Ageing Res. Rev. 2017, 39, 46–58. [Google Scholar] [CrossRef] [PubMed]
- Mashoodh, R.; Habrylo, I.B.; Gudsnuk, K.M.; Pelle, G.; Champagne, F.A. Maternal modulation of paternal effects on offspring development. Proc. Biol. Sci. 2018, 285, 20180118. [Google Scholar] [CrossRef]
- Govic, A.; Penman, J.; Tammer, A.H.; Paolini, A.G. Paternal calorie restriction prior to conception alters anxiety-like behavior of the adult rat progeny. Psychoneuroendocrinology 2016, 64, 1–11. [Google Scholar] [CrossRef]
- Power, C. Combination of low birth weight and high adult body mass index: At what age is it established and what are its determinants? J. Epidemiol. Community Health 2003, 57, 969–973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kasturi, S.S.; Tannir, J.; Brannigan, R.E. The Metabolic Syndrome and Male Infertility. J. Androl. 2008, 29, 251–259. [Google Scholar] [CrossRef]
- Portha, B.; Grandjean, V.; Movassat, J. Mother or Father: Who Is in the Front Line? Mechanisms Underlying the Non-Genomic Transmission of Obesity/Diabetes via the Maternal or the Paternal Line. Nutrients 2019, 11, 233. [Google Scholar] [CrossRef] [Green Version]
- Chowdhury, S.; LeComte, V.; Erlich, J.; Maloney, C.; Morris, M. Paternal High Fat Diet in Rats Leads to Renal Accumulation of Lipid and Tubular Changes in Adult Offspring. Nutrients 2016, 8, 521. [Google Scholar] [CrossRef] [Green Version]
- Masuyama, H.; Mitsui, T.; Eguchi, T.; Tamada, S.; Hiramatsu, Y. The effects of paternal high-fat diet exposure on offspring metabolism with epigenetic changes in the mouse adiponectin and leptin gene promoters. Am. J. Physiol. Metab. 2016, 311, E236–E245. [Google Scholar] [CrossRef] [Green Version]
- Ng, S.; Lin, R.C.Y.; Maloney, C.A.; Youngson, N.A.; Owens, J.A.; Morris, M.J. Paternal high-fat diet consumption induces common changes in the transcriptomes of retroperitoneal adipose and pancreatic islet tissues in female rat offspring. FASEB J. 2014, 28, 1830–1841. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Zhu, H.; Wu, H.; Jin, L.; Chen, B.; Pang, H.; Ming, Z.; Cheng, Y.; Zhou, C.; Guo, M.; et al. Diet-Induced Paternal Obesity Impairs Cognitive Function in Offspring by Mediating Epigenetic Modifications in Spermatozoa. Obesity 2018, 26, 1749–1757. [Google Scholar] [CrossRef]
- Choo, V.L.; Viguiliouk, E.; Blanco Mejia, S.; Cozma, A.I.; Khan, T.A.; Ha, V.; Wolever, T.M.S.; A Leiter, L.; Vuksan, V.; Kendall, C.W.C.; et al. Food sources of fructose-containing sugars and glycaemic control: Systematic review and meta-analysis of controlled intervention studies. BMJ 2018, 363, k4644. [Google Scholar] [CrossRef] [Green Version]
- Ornellas, F.; Carapeto, P.V.; Mandarim-De-Lacerda, C.A.; Aguila, M.B. Obese fathers lead to an altered metabolism and obesity in their children in adulthood: Review of experimental and human studies. J. Pediatr. 2017, 93, 551–559. [Google Scholar] [CrossRef]
- Emborski, C.; Mikheyev, A.S. Ancestral diet transgenerationally influences offspring in a parent-of-origin and sex-specific manner. Philos. Trans. R. Soc. B 2019, 374, 20180181. [Google Scholar] [CrossRef] [Green Version]
- Oshio, L.T.; Andreazzi, A.E.; Ferraz Lopes, J.; Pereira de Sá, J.; Bolotari, M.; Moreira Gonçalves Costa, V.; de Oliveira Guerra, M.; Peters, V.M. A paternal hypercaloric diet affects the metabolism and fertility of F1 and F2 Wistar rat generations. J. Dev. Orig. Health Dis. 2020, 11, 653–663. [Google Scholar] [CrossRef] [PubMed]
- Rando, O.J.; Simmons, R.A. I’m Eating for Two: Parental Dietary Effects on Offspring Metabolism. Cell 2015, 161, 93–105. [Google Scholar] [CrossRef] [Green Version]
- Mima, M.; Greenwald, D.; Ohlander, S. Environmental Toxins and Male Fertility. Curr. Urol. Rep. 2018, 19, 50. [Google Scholar] [CrossRef] [PubMed]
- Schellong, K.; Melchior, K.; Ziska, T.; Rancourt, R.C.; Henrich, W.; Plagemann, A. Maternal but Not Paternal High-Fat Diet (HFD) Exposure at Conception Predisposes for ‘Diabesity’ in Offspring Generations. Int. J. Environ. Res. Public Health 2020, 17, 4229. [Google Scholar] [CrossRef] [PubMed]
- Kong, A.; Frigge, M.L.; Masson, G.; Besenbacher, S.; Sulem, P.; Magnusson, G.; Gudjonsson, S.A.; Sigurdsson, A.; Jonasdottir, A.; Jonasdottir, A.; et al. Rate of de novo mutations and the importance of father’s age to disease risk. Nature 2012, 488, 471–475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jónsson, H.; Sulem, P.; Kehr, B.; Kristmundsdottir, S.; Zink, F.; Hjartarson, E.; Hardarson, M.T.; Hjorleifsson, K.E.; Eggertsson, H.P.; Gudjonsson, S.A.; et al. Parental influence on human germline de novo mutations in 1548 trios from Iceland. Nature 2017, 549, 519–522. [Google Scholar] [CrossRef] [PubMed]
- Jónsson, H.; Sulem, P.; Arnadottir, G.A.; Pálsson, G.; Eggertsson, H.P.; Kristmundsdottir, S.; Zink, F.; Kehr, B.; Hjorleifsson, K.E.; Jensson, B.Ö.; et al. Multiple transmissions of de novo mutations in families. Nat. Genet. 2018, 50, 1674–1680. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Effects | Paternal Diet | References | |
|---|---|---|---|
| Metabolism | Body weight and adiposity | Low protein HFD Food restriction FA deficient High sugar | [21,36,37,38,39,40,41,42,43] |
| Glucose intolerance or hyperglycemia | Low protein HFD FA supplemented High sugar | [37,38,40,44,45,46,47] | |
| Insulin resistance or hyperinsulinemia | HFD High sugar | [40,45,46,47] | |
| Carbohydrate metabolism | HFD | [42,48] | |
| Lipid metabolism | High sugar FA deficient or supplemented Food restriction | [21,44,46,47] | |
| Uricemia | High sugar | [43,46,47] | |
| Development | Skeleton | Low protein FA deficient | [49,50] |
| Fetal or post-natal growth | Food restriction | [21] | |
| Spermatogenesis | FA deficient | [49] | |
| Embryo or placenta abnormalities | FA supplemented HFD | [42,48,51,52,53] | |
| Pregnancy outcomes | Number of plugs/pregnancies | HFD FA deficient | [41,42,48,52] |
| Pre or Post implantation development | HFD FA deficient Food restriction | [21,42,48,52] | |
| Morphological retardation | HFD FA deficient | [42,48,49] | |
| Postnatal mortality | FA deficient or supplemented | [51] | |
| Organ and system dysfunction | Breast cancer | Low protein | [54] |
| Diabetic kidney disease | High sugar | [55] | |
| Cardiovascular system | Low protein High sugar | [37,38,43,46,47,55] | |
| Sperm | HFD High sugar FA deficient or supplemented | [41,46,51,56,57] | |
| Endocrine system | High sugar | [46] | |
| Central nervous system | FA deficient | [36,58] | |
| Effects | Paternal Diet | References | Comments | |
|---|---|---|---|---|
| Epigenetic alterations | Sperm DMRs 1 | Low protein FA deficient or supplemented | [51,54] | Paternally imprinted gene H19 |
| DNA methylation | Low protein HFD FA deficient Food restriction | [38,39,49,56] | Genes involved in neurological and autoimmune disease, diabetes, cancer, autism and schizophrenia | |
| Gene expression and transcriptome | High sugar | [63,67] | H3K9-me3 regulators and Polycomb | |
| Sperm mutations | FA deficient or supplemented | [57] | DNA fragmentation and tandem repetitions | |
| RNA changes | Sperm mRNA content and/or expression levels | Low protein HFD High sugar FA sufficient | [24,25,39,40,64,65,67,68,69] | Metabolic disorders, lipid metabolism, embryonic development and spermatogenesis |
| sncRNA | Low protein High sugar | [25,69] | tRNA and tsRNA | |
| HFD FA supplemented | [39,68] | piRNA | ||
| Low protein HFD FA supplemented | [25,39,40,44,64,65,66] | miRNA let-7 species miR-19 species miR-182 miR-183 miR-340 miR-193b miR-204 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dimofski, P.; Meyre, D.; Dreumont, N.; Leininger-Muller, B. Consequences of Paternal Nutrition on Offspring Health and Disease. Nutrients 2021, 13, 2818. https://doi.org/10.3390/nu13082818
Dimofski P, Meyre D, Dreumont N, Leininger-Muller B. Consequences of Paternal Nutrition on Offspring Health and Disease. Nutrients. 2021; 13(8):2818. https://doi.org/10.3390/nu13082818
Chicago/Turabian StyleDimofski, Pauline, David Meyre, Natacha Dreumont, and Brigitte Leininger-Muller. 2021. "Consequences of Paternal Nutrition on Offspring Health and Disease" Nutrients 13, no. 8: 2818. https://doi.org/10.3390/nu13082818
APA StyleDimofski, P., Meyre, D., Dreumont, N., & Leininger-Muller, B. (2021). Consequences of Paternal Nutrition on Offspring Health and Disease. Nutrients, 13(8), 2818. https://doi.org/10.3390/nu13082818