
Metabolite Profile and In Vitro Beneficial Effects of Black Garlic (Allium sativum L.) Polar Extract

 ,
, 
 , and
, and 
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Plant Material and Preparation of the Methanol Extract of Black Garlic
2.3. LC-ESI/HRMS/MS Analysis
2.4. Cell Lines and Culture Conditions
2.5. Cellular Bioactivity
2.5.1. Cell Viability Assay
2.5.2. Cellular Oxidative Stress: Nitrite Levels in the Culture Medium
2.5.3. Cellular Oxidative Stress: Lipid Peroxidation in Cytosol
2.5.4. Metabolic Parameters
2.5.5. Oil Red O Staining
2.6. Microscopic Analysis
2.7. Differentiation Assay
2.8. Western Blot Analysis
3. Results and Discussion
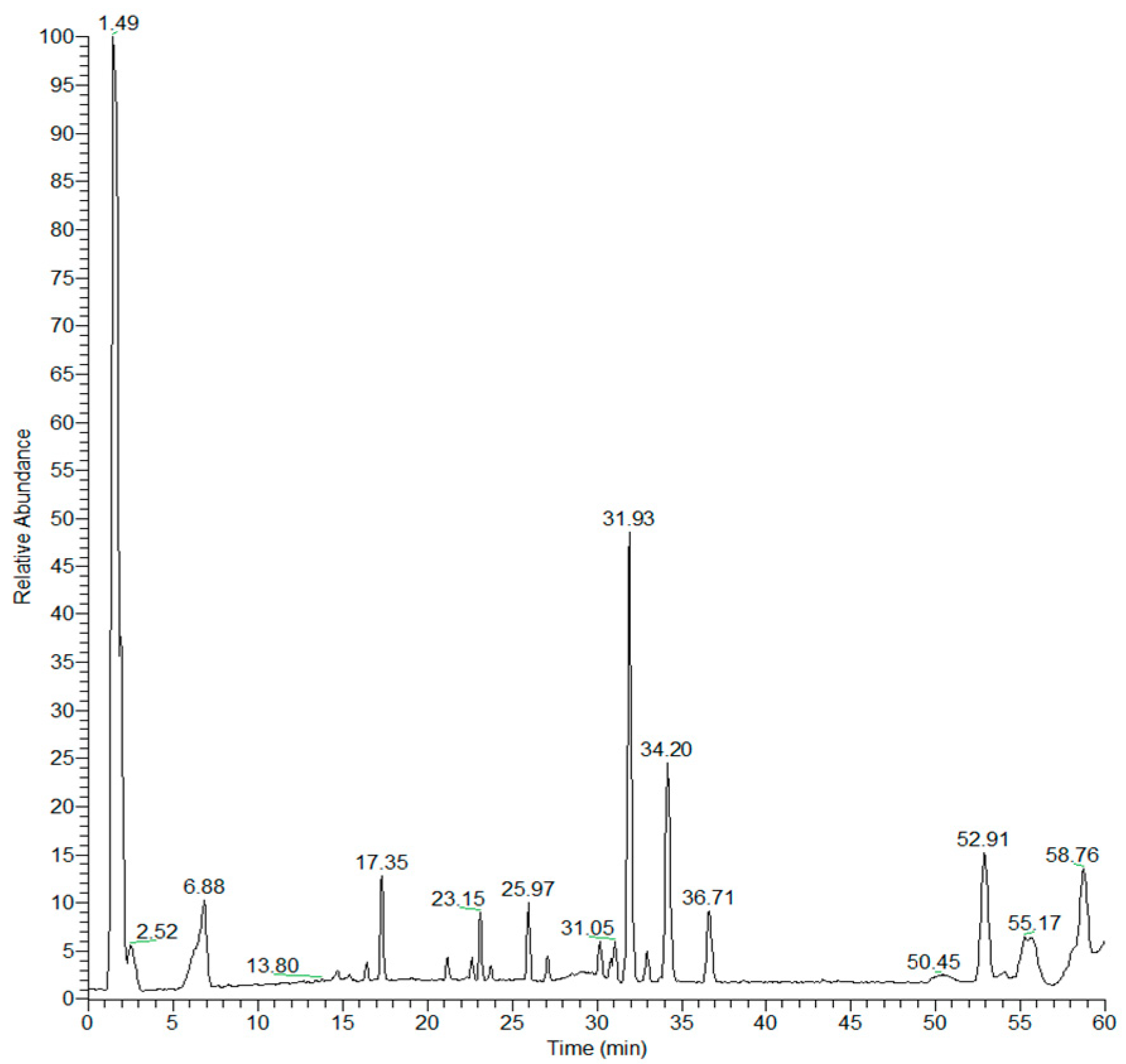
3.1. Characterization of Black Garlic Methanol Extract
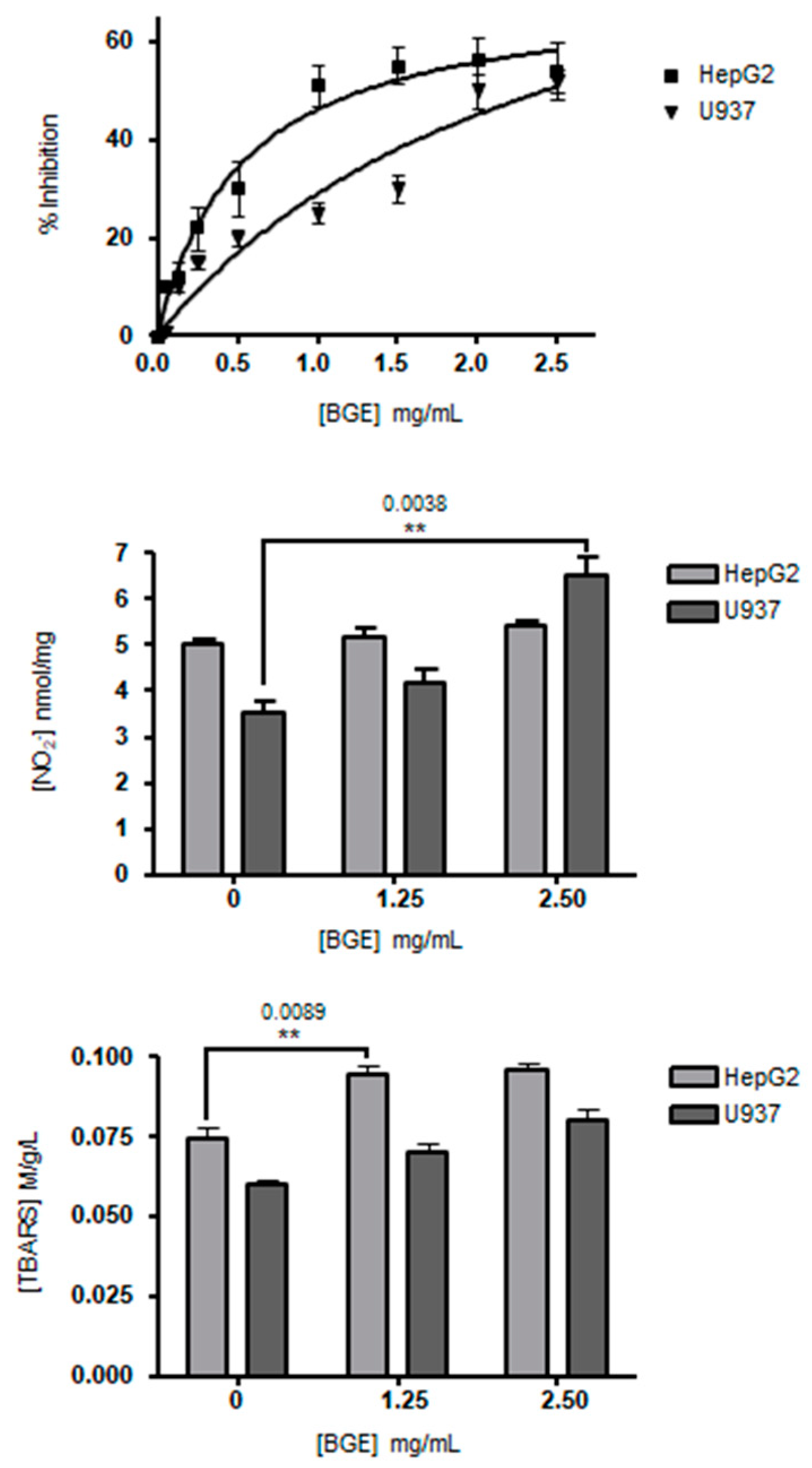
3.2. Effects of Black Garlic Methanol Extract on Cancer Cell Viability and Oxidative Stress
3.3. Black Garlic Methanol Extract Reduces Glucose, Triglycerides and Cholesterol Accumulation in Liver Cells
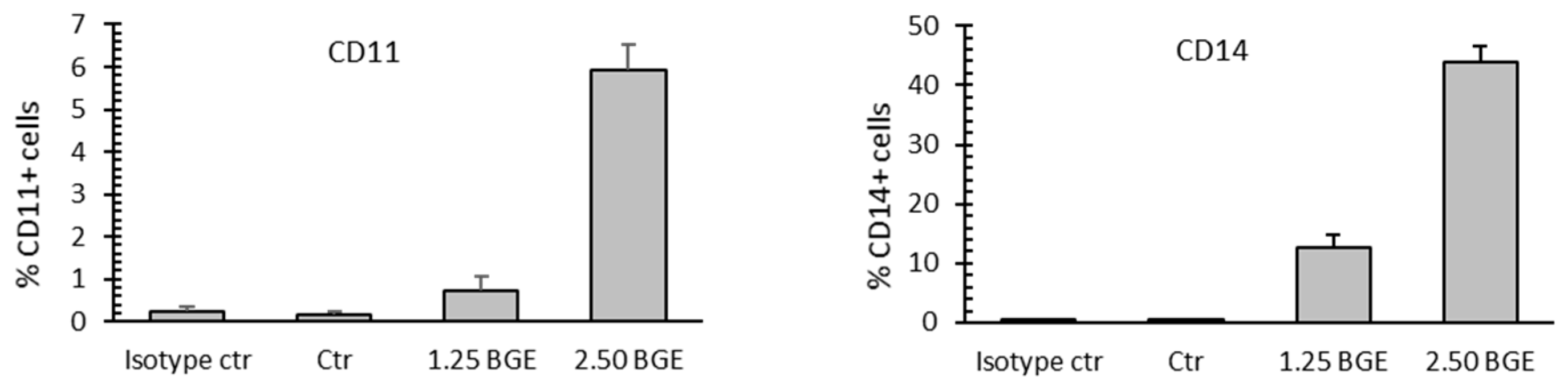
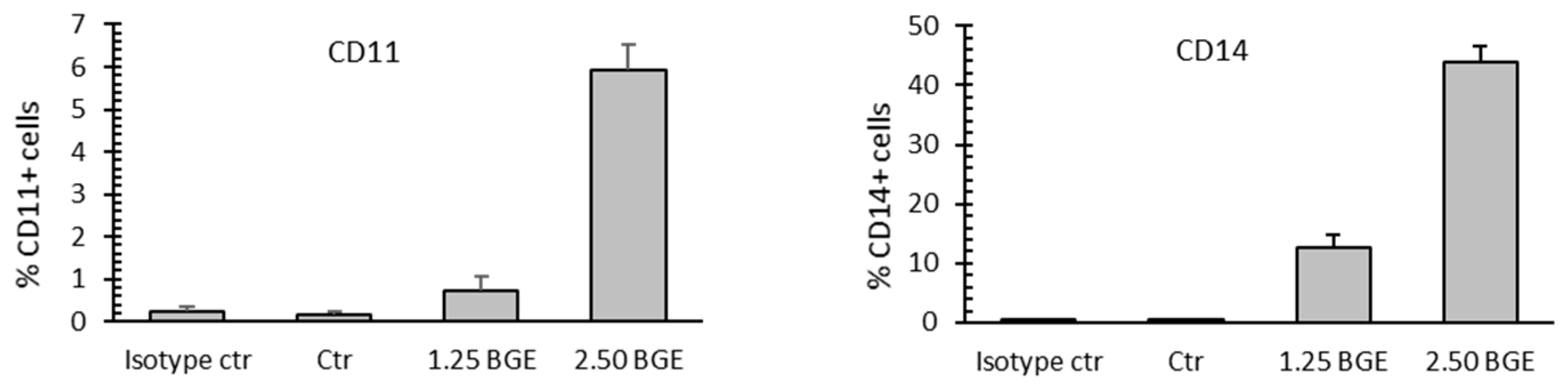
3.4. Black Garlic Mehanol Extract Induces Differentiation in Hematological Cancer Cell Lines
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mushtaq, S.; Abbasi, B.H.; Uzair, B.; Abbasi, R. Natural products as reservoirs of novel therapeutic agents. EXCLI J. 2018, 17, 420–451. [Google Scholar] [CrossRef]
- Nyakudya, T.T.; Tshabalala, T.; Dangarembizi, R.; Erlwanger, K.H.; Ndhlala, A.R. The Potential Therapeutic Value of Medicinal Plants in the Management of Metabolic Disorders. Molecules 2020, 25, 2669. [Google Scholar] [CrossRef]
- Slavin, J.L.; Lloyd, B. Health benefits of fruits and vegetables. Adv. Nutr. 2012, 3, 506–516. [Google Scholar] [CrossRef] [Green Version]
- Bontempo, P.; Carafa, V.; Grassi, R.; Basile, A.; Tenore, G.C.; Formisano, C.; Rigano, D.; Altucci, L. Antioxidant, antimicrobial and antiproliferative activities of Solanum tuberosum L. var. Vitelotte. Food Chem. Toxicol. 2013, 55, 304–312. [Google Scholar] [CrossRef] [PubMed]
- Scafuri, B.; Bontempo, P.; Altucci, L.; De Masi, L.; Facchiano, A. Molecular Docking Simulations on Histone Deacetylases (HDAC)-1 and -2 to Investigate the Flavone Binding. Biomedicines 2020, 8, 568. [Google Scholar] [CrossRef] [PubMed]
- Shang, A.; Cao, S.-Y.; Xu, X.-Y.; Gan, R.-Y.; Tang, G.-Y.; Corke, H.; Mavumengwana, V.; Li, H.-B. Bioactive Compounds and Biological Functions of Garlic (Allium sativum L.). Foods 2019, 8, 246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salehi, B.; Zucca, P.; Orhan, I.E.; Azzini, E.; Adetunji, C.O.; Mohammed, S.A.; Banerjee, S.K.; Sharopov, F.; Rigano, D.; Sharifi-Rad, J.; et al. Allicin and health: A comprehensive review. Trends Food Sci. Tech. 2019, 86, 502–516. [Google Scholar] [CrossRef]
- Kimura, S.; Tung, Y.C.; Pan, M.H.; Su, N.W.; Lai, Y.J.; Cheng, K.C. Black garlic: A critical review of its production, bioactivity, and application. J. Food Drug Anal. 2017, 25, 62–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryu, J.H.; Kang, D. Physicochemical Properties, Biological Activity, Health Benefits, and General Limitations of Aged Black Garlic: A Review. Molecules 2017, 22, 919. [Google Scholar] [CrossRef] [Green Version]
- Nassur, R.C.; Boas, E.V.; Resende, F.V. Black garlic: Transformation effects, characterization and consumer purchase intention. Comun. Sci. 2018, 8, 444–451. [Google Scholar] [CrossRef]
- Lindenmeier, M.; Faist, V.; Hofmann, T. Structural and functional characterization of pronyl-lysine, a novel protein modification in bread crust melanoidins showing in vitro antioxidative and phase I/II enzyme modulating activity. J. Agric. Food Chem. 2002, 50, 6997–7006. [Google Scholar] [CrossRef] [PubMed]
- Kang, O.J. Evaluation of Melanoidins Formed from Black Garlic after Different Thermal Processing Steps. Prev. Nutr. Food Sci. 2016, 21, 398–405. [Google Scholar] [CrossRef] [Green Version]
- Chang, W.C.-W.; Chen, Y.-T.; Chen, H.-J.; Hsieh, C.-W.; Liao, P.-C. Comparative UHPLC-Q-Orbitrap HRMS-Based Metabolomics Unveils Biochemical Changes of Black Garlic during Aging Process. J. Agric. Food Chem. 2020, 68, 47, 14049–14058. [Google Scholar] [CrossRef] [PubMed]
- Amor, S.; González-Hedström, D.; Martín-Carro, B.; Inarejos-García, A.M.; Almodóvar, P.; Prodanov, M.; García-Villalón, A.L.; Granado, M. Beneficial Effects of an Aged Black Garlic Extract in the Metabolic and Vascular Alterations Induced by a High Fat/Sucrose Diet in Male Rats. Nutrients 2019, 11, 153. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.G.; Kang, M.J.; Hong, S.S.; Choi, Y.H.; Shin, J.H. Antiinflammatory Effects of Functionally Active Compounds Isolated from Aged Black Garlic. Phytother. Res. 2017, 31, 53–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, J.H.; Lee, C.W.; Oh, S.J.; Yun, J.; Kang, M.R.; Han, S.B.; Park, H.; Jung, J.C.; Chung, Y.H.; Kang, J.S. Hepatoprotective effect of aged black garlic extract in rodents. Toxicol. Res. 2014, 30, 49–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Botas, J.; Fernandes, Â.; Barros, L.M.; Alves, J.; Carvalho, A.M.; Ferreira, I.C.F.R. A Comparative Study of Black and White Allium sativum L.: Nutritional Composition and Bioactive Properties. Molecules 2019, 11, 2194–2205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, X.; Li, N.; Qiao, X.; Qiu, Z.; Liu, P. Composition analysis and antioxidant properties of black garlic extract. J. Food Drug Anal. 2017, 25, 340–349. [Google Scholar] [CrossRef] [Green Version]
- De Maria, S.; Scognamiglio, I.; Lombardi, A.; Amodio, N.; Caraglia, M.; Cartenì, M.; Ravagnan, G.; Stiuso, P. Polydatin, a natural precursor of resveratrol, induces cell cycle arrest and differentiation of human colorectal Caco-2 cell. J. Transl. Med. 2013, 11. [Google Scholar] [CrossRef] [Green Version]
- Lama, S.; Vanacore, D.; Diano, N.; Nicolucci, C.; Errico, S.; Dallio, M.; Federico, A.; Loguercio, C.; Stiuso, P. Ameliorative effect of Silybin on bisphenol A induced oxidative stress, cell proliferation and steroid hormones oxidation in HepG2 cell cultures. Sci. Rep. 2019, 9. [Google Scholar] [CrossRef]
- Stiuso, P.; Scognamiglio, I.; Murolo, M.; Ferranti, P.; de Simone, C.; Rizzo, M.R.; Tuccillo, C.; Caraglia, M.; Loguercio, C.; Federico, A. Serum oxidative stress markers and lipidomic profile to detect NASH patients responsive to an antioxidant treatment: A pilot study. Oxid. Med. Cell. Longev. 2014. [Google Scholar] [CrossRef] [PubMed]
- Bontempo, P.; De Masi, L.; Carafa, V.; Rigano, D.; Scisciola, L.; Iside, C.; Grassi, R.; Molinari, A.M.; Aversano, R.; Nebbioso, A.; et al. Anticancer activities of anthocyanin extract from genotyped Solanum tuberosum L. “Vitelotte”. J. Funct. Foods 2015, 19, 584–593. [Google Scholar] [CrossRef] [Green Version]
- De Masi, L.; Bontempo, P.; Rigano, D.; Stiuso, P.; Carafa, V.; Nebbioso, A.; Piacente, S.; Montoro, P.; Aversano, R.; D’Amelia, V.; et al. Comparative Phytochemical Characterization, Genetic Profile, and Antiproliferative Activity of Polyphenol-Rich Extracts from Pigmented Tubers of Different Solanum tuberosum Varieties. Molecules 2020, 25, 233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bontempo, P.; Doto, A.; Miceli, M.; Mita, L.; Benedetti, R.; Nebbioso, A.; Veglione, M.; Rigano, D.; Cioffi, M.; Sica, V.; et al. Psidium guajava L. anti-neoplastic effects: Induction of apoptosis and cell differentiation. Cell Prolif. 2012, 45, 22–31. [Google Scholar] [CrossRef]
- Bontempo, P.; Rigano, D.; Doto, A.; Formisano, C.; Conte, M.; Nebbioso, A.; Carafa, V.; Caserta, G.; Sica, V.; Molinari, A.M.; et al. Genista sessilifolia DC. extracts induce apoptosis across a range of cancer cell lines. Cell Prolif. 2013, 46, 183–192. [Google Scholar] [CrossRef] [PubMed]
- Molina-Calle, M.; Sánchez de Medina, V.; Calderón-Santiago, M.; Priego-Capote, F.; Luque de Castro, M.D. Untargeted analysis to monitor metabolic changes of garlic along heat treatment by LC-QTOF MS/MS. Electrophoresis 2017, 38, 2349–2360. [Google Scholar] [CrossRef]
- Sun, H.; Liu, M.; Lin, Z.; Jiang, H.; Niu, Y.; Wang, H.; Chen, S. Comprehensive identification of 125 multifarious constituents in Shuang-huang-lian powder injection by HPLC-DAD-ESI-IT-TOF-MS. J. Pharm. Biomed. Anal. 2015, 115, 86–106. [Google Scholar] [CrossRef] [PubMed]
- Marzouki, S.M.; Almagro, L.; Sabater-Jara, A.B.; Barcelo, A.R.; Pedreno, M.A. Kinetic characterization of a basic peroxidase from garlic (Allium sativum L.) cloves. J. Food Sci. 2010, 75, C740–C746. [Google Scholar] [CrossRef]
- Woo, K.W.; Lee, K.R. Phytochemical constituents of Allium victorialis var. platyphyllum. Nat. Prod. Sci. 2013, 19, 221–226. [Google Scholar]
- Matsuura, H.; Ushiroguchi, T.; Itakura, Y.; Fuwa, T. Further studies on steroidal glycosides from bulbs, roots and leaves of Allium sativum L. Chem. Pharm. Bull. 1989, 37, 2741–2743. [Google Scholar] [CrossRef] [Green Version]
- Lanzotti, V. The analysis of onion and garlic. J. Chromatogr. A 2006, 1112, 3–22. [Google Scholar] [CrossRef] [Green Version]
- Lanzotti, V.; Barile, E.; Antignani, V.; Bonanomi, G.; Scala, F. Antifungal saponins from bulbs of garlic, Allium sativum L. var. Voghiera. Phytochemistry 2012, 78, 126–134. [Google Scholar] [CrossRef] [PubMed]
- Diretto, G.; Rubio-Moraga, A.; Argandoña, J.; Castillo, P.; Gómez-Gómez, L.; Ahrazem, O. Tissue-Specific Accumulation of Sulfur Compounds and Saponins in Different Parts of Garlic Cloves from Purple and White Ecotypes. Molecules 2017, 22, 1359–1372. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues Adão, C.; Pereira da Silva, B.; Wanderley Tinoco, L.; Parente, J.P. Haemolytic Activity and Immunological Adjuvant Effect of a New Steroidal Saponin from Allium ampeloprasum var. porrum. Chem. Biodivers. 2012, 9, 58–67. [Google Scholar] [CrossRef]
- Patras, M.A.; Milev, B.P.; Vrancken, G.; Kuhnert, N. Identification of novel cocoa flavonoids from raw fermented cocoa beans by HPLC-MSn. Food Res. Int. 2014, 63, 353–359. [Google Scholar] [CrossRef]
- Han, B.; Xin, Z.; Ma, S.; Liu, W.; Zhang, B.; Ran, L.; Yi, L.; Ren, D. Comprehensive characterization and identification of antioxidants in Folium Artemisiae Argyi using high-resolution tandem mass spectrometry. J. Chromatogr. B 2017, 1063, 84–92. [Google Scholar] [CrossRef]
- Napolitano, A.; Cerulli, A.; Pizza, C.; Piacente, S. Multi-class polar lipid profiling in fresh and roasted hazelnut (Corylus avellana cultivar “Tonda di Giffoni”) by LC-ESI/LTQOrbitrap/MS/MSn. Food Chem. 2018, 269, 125–135. [Google Scholar] [CrossRef]
- Tsiaganis, M.C.; Laskari, K.; Melissari, E. Fatty acid composition of Allium species lipids. J. Food Compos. Anal. 2006, 19, 620–627. [Google Scholar] [CrossRef]
- García-Villalón, A.L.; Amor, S.; Monge, L.; Fernández, N.; Prodanov, M.; Muñoz, M.; Inarejos-García, A.M.; Granado, M. In vitro studies of an aged black garlic extract enriched in S-allylcysteine and polyphenols with cardioprotective effects. J. Funct. Foods 2016, 27, 189–200. [Google Scholar] [CrossRef] [Green Version]
- Magrinat, G.; Mason, S.N.; Shami, P.J.; Weinberg, J.B. Nitric oxide modulation of human leukemia cell differentiation and gene expression. Blood 1992, 80, 1880–1884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shami, P.J.; Weinberg, J.B. Differential Effects of Nitric Oxide on Erythroid and Myeloid Colony Growth From CD34+ Human Bone Marrow Cells. Blood 1996, 87, 977–982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shami, P.J.; Kanai, N.; Wang, L.Y.; Vreeke, T.M.; Parker, C.J. Identification and characterization of a novel gene that is upregulated in leukaemia cells by nitric oxide. Br. J. Haematol. 2001, 112, 138–147. [Google Scholar] [CrossRef] [PubMed]
- Rinaldi, M.; Barrera, G.; Aquino, A.; Spinsanti, P.; Pizzimenti, S.; Farace, M.G.; Dianzani, M.U.; Fazio, V.M. 4-Hydroxynonenal-induced MEL cell differentiation involves PKC activity translocation. Biochem. Biophys. Res. Commun. 2000, 272, 75–80. [Google Scholar] [CrossRef]
- Wang, X.; Jiao, F.; Wang, Q.-W.; Wang, J.; Yang, K.; Hu, R.-R.; Liu, H.-C.; Wang, H.-Y.; Wang, Y.-S. Aged black garlic extract induces inhibition of gastric cancer cell growth in vitro and in vivo. Mol. Med. Rep. 2012, 5, 66–72. [Google Scholar] [CrossRef]
- Netala, V.R.; Ghosh, S.B.; Bobbu, P.; Anitha, D.; Tartte, V. Triterpenoid saponins: A review on biosynthesis, applications and mechanism of their action. Int. J. Pharm. Pharm. Sci. 2014, 7, 24–28. Available online: https://innovareacademics.in/journals/index.php/ijpps/article/view/2807 (accessed on 11 August 2021).
- Ha, A.W.; Ying, T.; Kim, W.K. The effects of black garlic (Allium satvium) extracts on lipid metabolism in rats fed a high fat diet. Nutr. Res. Pract. 2015, 9, 30–36. [Google Scholar] [CrossRef] [Green Version]
- Chandrasekaran, K.; Swaminathan, K.; Chatterjee, S.; Dey, A. Apoptosis in HepG2 cells exposed to high glucose. Toxicol. Vitr. 2010, 24, 387–396. [Google Scholar] [CrossRef]
- Valtueña, S.; Pellegrini, N.; Ardigò, D.; Del Rio, D.; Numeroso, F.; Scazzina, F.; Monti, L.; Zavaroni, I.; Brighenti, F. Dietary glycemic index and liver steatosis. Am. J. Clin. Nutr. 2006, 84, 136–142. [Google Scholar] [CrossRef]
- Kim, M.H.; Kim, M.J.; Lee, J.H.; Han, J.I.; Kim, J.H.; Sok, D.E.; Kim, M.R. Hepatoprotective effect of aged black garlic on chronic alcohol-induced liver injury in rats. J. Med. Food 2011, 14, 732–738. [Google Scholar] [CrossRef]
- Joost, H.G.; Bell, G.I.; Best, J.D.; Birnbaum, M.J.; Charron, M.J.; Chen, Y.T.; Doege, H.; James, D.E.; Lodish, H.F.; Moley, K.H.; et al. Nomenclature of the GLUT/SLC2A family of sugar/polyol transport facilitators. Am. J. Physiol. Endocrinol. Metab. 2002, 282, 974–976. [Google Scholar] [CrossRef] [Green Version]
- Marrelli, M.; Conforti, F.; Araniti, F.; Statti, G.A. Effects of Saponins on Lipid Metabolism: A Review of Potential Health Benefits in the Treatment of Obesity. Molecules 2016, 21, 1404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, M.; Yang, G.; Liu, H.; Liu, X.; Lin, S.; Sun, D.; Wang, Y. Aged black garlic extract inhibits HT29 colon cancer cell growth via the PI3K/Akt signaling pathway. Biomed. Rep. 2014, 2, 250–254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, C.; Park, S.; Chung, Y.H.; Kim, G.Y.; Choi, Y.W.; Kim, B.W.; Choi, Y.H. Induction of apoptosis by a hexane extract of aged black garlic in the human leukemic U937 cells. Nutr. Res. Pract. 2014, 8, 132–137. [Google Scholar] [CrossRef] [PubMed]
- Toledano Medina, M.Á.; Merinas-Amo, T.; Fernández-Bedmar, Z.; Font, R.; del Río-Celestino, M.; Pérez-Aparicio, J.; Moreno-Ortega, A.; Alonso-Moraga, Á.; Moreno-Rojas, R. Physicochemical Characterization and Biological Activities of Black and White Garlic: In Vivo and In Vitro Assays. Foods 2019, 8, 220. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| n° | Compound | Rt (min) | Molecular Formula | [M-H]− | [(M + H2CO2)-H]− | Delta (ppm) | Characteristic Product Ions (m/z) |
|---|---|---|---|---|---|---|---|
| 1 | S-Allyl-cysteine | 1.08 | C6H11NO2S | 160.0430 | 2.09 | 73, 88 | |
| 2 | Alliin | 1.44 | C6H11NO3S | 176.0383 | 3.92 | 88, 89 | |
| 3 | disaccharide | 1.49 | C12H22O11 | 341.1090 | 387.1141 | 3.44 | 323, 297, 281, 251, 179, 161, 143, 113 |
| 4 | β-D-fructofuranose 2′,1:2,1′-dianhydride | 1.54 | C12H20O10 | 323.0984 | 369.1038 | 3.36 | 233, 161 |
| 5 | Allicin | 1.64 | C6H10OS2 | 161.0092 | 1.47 | 73, 89 | |
| 6 | hexose-phosphate | 1.75 | C6H13O9P | 259.0220 | 2.53 | 241, 217, 199, 169, 97, 79 | |
| 7 | L-pyroglutamic acid-O- hexose ester | 1.94 | C11H17O8N | 290.0876 | 1.88 | 272, 230, 200, 170, 128, 113 | |
| 8 | Citric acid | 2.30 | C6H8O7 | 191.0193 | 3.51 | 173, 147, 111, 87 | |
| 9 | Methyl citric acid | 2.52 | C7H10O7 | 205.0350 | 3.66 | 173, 161, 143, 111 | |
| 10 | γ-Glutamylphenylalanine | 6.21 | C14H18O5N2 | 293.1136 | 1.24 | 275, 249, 164, 128 | |
| 11 | γ-Glutamylphenylalanine-O-hexose ester | 6.88 | C20H28O10N2 | 455.1659 | -0.07 | 437, 393, 365, 335, 293, 290, 275, 200, 170, 164, 128 | |
| 12 | Syringin rhamnoside | 15.09 | C23H34O13 | 517.1916 | 517.1916 | 0.04 | 499, 307, 163 |
| 13 | Porrigenin B 3-O-pentasaccharide | 17.55 | C57H92O30 | 1255.5562 | 1301.5618 | -2.24 | 1093, 931, 769 |
| 14 | 9,12,13-Trihydroxy octadecenoic acid | 21.30 | C18H34O5 | 329.2329 | 1.91 | 311, 293, 229, 211, 199, 197, 171, 141, 127 | |
| 15 | gitogenin 3-O-tetrasaccharide or β-chlorogenin 3-O-tetrasaccharide | 23.75 | C51H84O24 | 1079.5248 | 1125.5314 | -1.94 | 917, 899, 755, 593 |
| 16 | gitogenin 3-O-trisaccharide or β-chlorogenin-3-O-trisaccharide | 24.18 | C45H74O19 | 917.4766 | 963.4778 | 2.73 | 755, 737, 593 |
| 17 | Tigogenin | 27.10 | C27H44O3 | 415.3192 | -3.49 | 371, 369, 355, 347, 311, 253 | |
| 18 | Hydroxyjasmonic acid sulfate | 27.31 | C12H18O7S | 305.0694 | 1.38 | 287, 261, 225, 207, 175, 97 | |
| 19 | DGMG (18:3) | 28.54 | C33H56O14 | 675.3573 | 721.3636 | -1.97 | 397, 277 |
| 20 | l-PE (18:2) | 30.80 | C23H44O7NP | 476.2775 | 0.64 | 279, 214, 196, 153 | |
| 21 | DGMG (18:2) | 30.82 | C33H58O14 | 677.3737 | 723.3794 | -0.83 | 415, 397 |
| 22 | l-PC (18:2) | 31.05 | C26H50O7NP | 564.3296 | 0.09 | 504, 279 | |
| 23 | l-PE (18:2) | 31.62 | C23H44O7NP | 476.2769 | -0.64 | 279, 214, 196, 153 | |
| 24 | l-PC (18:2) | 31.92 | C26H50O7NP | 564.3294 | -0.33 | 504, 279 | |
| 25 | l-PE (16:0) | 32.55 | C21H44O7NP | 452.2779 | 1.53 | 255, 214, 196 | |
| 26 | l-PE (18:2) | 32.60 | C23H44O7NP | 476.2767 | -1.04 | 279, 214, 196, 153 | |
| 27 | DGMG (16:0) | 32.61 | C31H58O14 | 653.3752 | 699.3796 | 1.37 | 397 |
| 28 | l-PC (16:0) | 32.91 | C24H50O7NP | 540.3299 | 0.67 | 480, 255 | |
| 29 | l-PC (18:2) | 32.96 | C26H50O7NP | 564.3296 | 0.09 | 504, 279 | |
| 30 | l-PE (16:0) | 33.73 | C21H44O7NP | 452.2769 | -0.54 | 255, 214, 196 | |
| 31 | l-PC (16:0) | 34.20 | C24H50O7NP | 540.3296 | 0.10 | 480, 255 | |
| 32 | l-PC (18:1) | 35.17 | C26H52O7NP | 566.3451 | -0.06 | 506, 281 | |
| 33 | l-PC (18:1) | 36.71 | C26H52O7NP | 566.3450 | 0.06 | 506, 281 | |
| 34 | l-PI (18:2) | 50.45 | C27H49O12P | 595.2869 | -1.34 | 415, 315, 279, 241 | |
| 35 | l-PI (18:2) | 52.91 | C27H49O12P | 595.2877 | -0.12 | 415, 333, 315, 279, 241, 223 | |
| 36 | l-PI (16:0) | 55.17 | C27H49O12P | 571.2873 | -0.77 | 409, 391, 315, 255, 241, 223, 171 | |
| 37 | l-PI (18:2) | 55.73 | C27H49O12P | 595.2874 | -0.64 | 415, 315, 279, 241, 171 | |
| 38 | l-PI (16:0) | 58.76 | C25H49O12P | 571.2880 | 0.40 | 409, 391, 315, 255, 241, 223, 171 |
| Glucose (mg/dL/Cell n.) | CHO (mg/dL/Cell n.) | TG (mg/dL/Cell n.) | TBARS (M/g/L) | |
|---|---|---|---|---|
| HG-HepG2 (CTR) | 638 ± 50 | 10 ± 1 | 15 ± 3 | 0.15 ± 0.03 |
| HG-HepG2 + BGE | 1530 ± 200 | 38 ± 6 | 74 ± 7 | 0.098 ± 0.005 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bontempo, P.; Stiuso, P.; Lama, S.; Napolitano, A.; Piacente, S.; Altucci, L.; Molinari, A.M.; De Masi, L.; Rigano, D. Metabolite Profile and In Vitro Beneficial Effects of Black Garlic (Allium sativum L.) Polar Extract. Nutrients 2021, 13, 2771. https://doi.org/10.3390/nu13082771
Bontempo P, Stiuso P, Lama S, Napolitano A, Piacente S, Altucci L, Molinari AM, De Masi L, Rigano D. Metabolite Profile and In Vitro Beneficial Effects of Black Garlic (Allium sativum L.) Polar Extract. Nutrients. 2021; 13(8):2771. https://doi.org/10.3390/nu13082771
Chicago/Turabian StyleBontempo, Paola, Paola Stiuso, Stefania Lama, Assunta Napolitano, Sonia Piacente, Lucia Altucci, Anna Maria Molinari, Luigi De Masi, and Daniela Rigano. 2021. "Metabolite Profile and In Vitro Beneficial Effects of Black Garlic (Allium sativum L.) Polar Extract" Nutrients 13, no. 8: 2771. https://doi.org/10.3390/nu13082771
APA StyleBontempo, P., Stiuso, P., Lama, S., Napolitano, A., Piacente, S., Altucci, L., Molinari, A. M., De Masi, L., & Rigano, D. (2021). Metabolite Profile and In Vitro Beneficial Effects of Black Garlic (Allium sativum L.) Polar Extract. Nutrients, 13(8), 2771. https://doi.org/10.3390/nu13082771