
Reduced Liver-Specific PGC1a Increases Susceptibility for Short-Term Diet-Induced Weight Gain in Male Mice

 , , ,
, , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Genotyping and RT-PCR
2.3. Liver Isolated Mitochondria Fatty Acid and Pyruvate Oxidation
2.4. Mitochondrial Respiration
2.5. Indirect Calorimetry Experiments
2.6. Common Hepatic Branch Vagotomy (HBV) Experiments
2.6.1. Surgery
2.6.2. HFHS-Induced Weight Gain and Body Composition
2.7. Statistical Analysis
3. Results
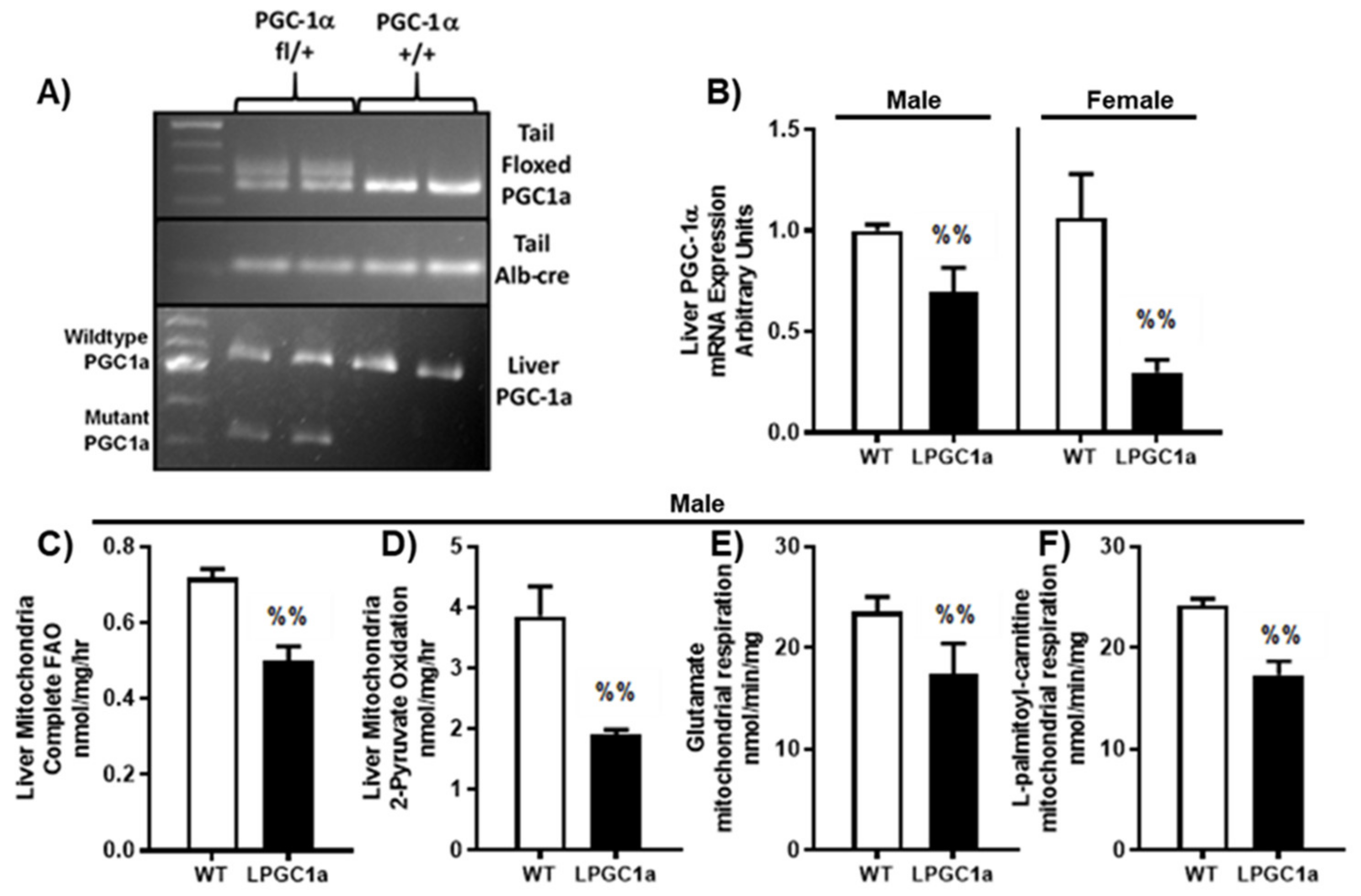
3.1. Liver-Specific PGC1a Heterozygous Mice Have Reduced Mitochondrial Fatty Acid Oxidation and Respiratory Capacity
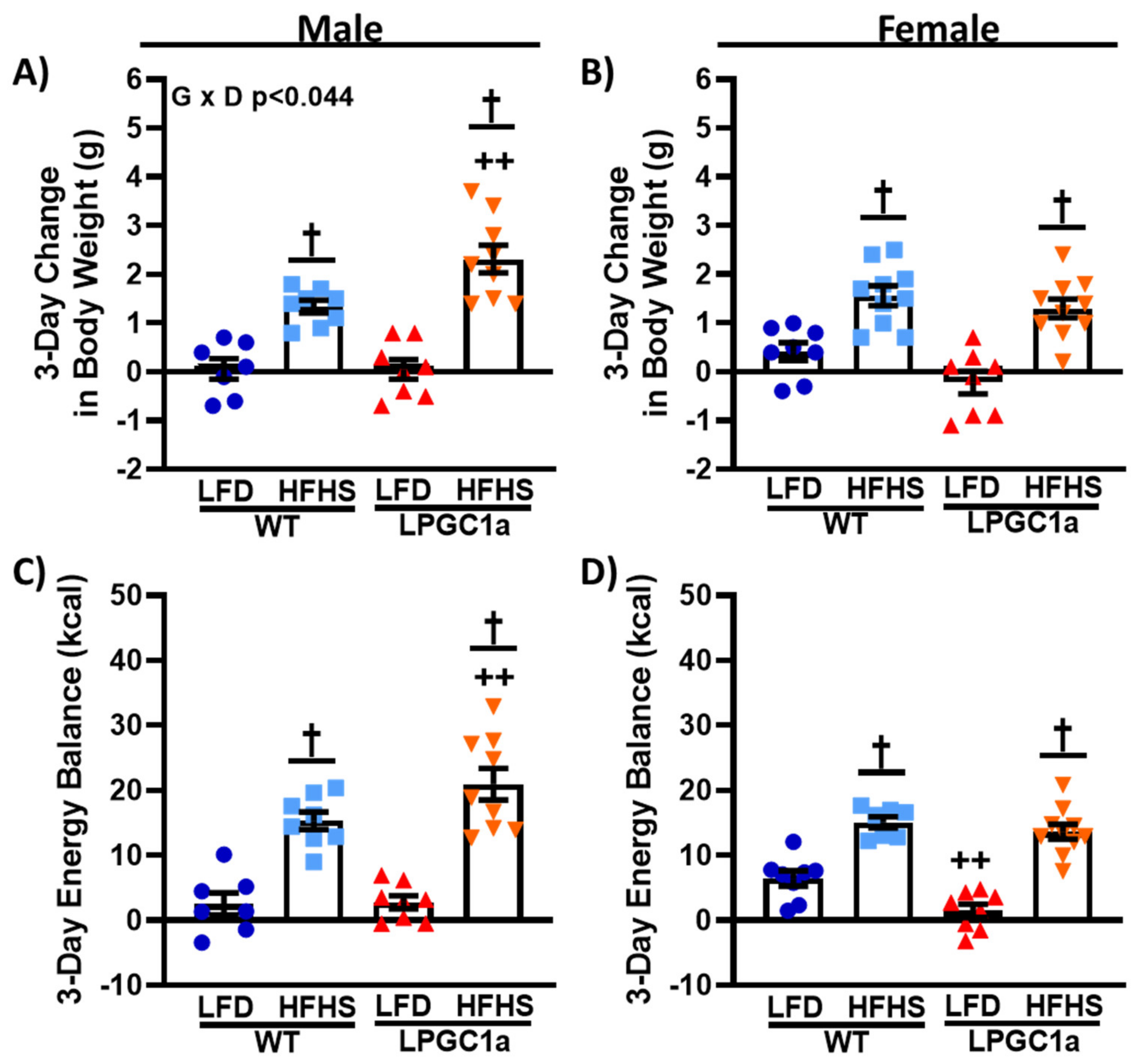
3.2. Reduced Liver PGC1a Increases Susceptibility to Short-Term HFHS Weight Gain in Male, but Not Female, Mice
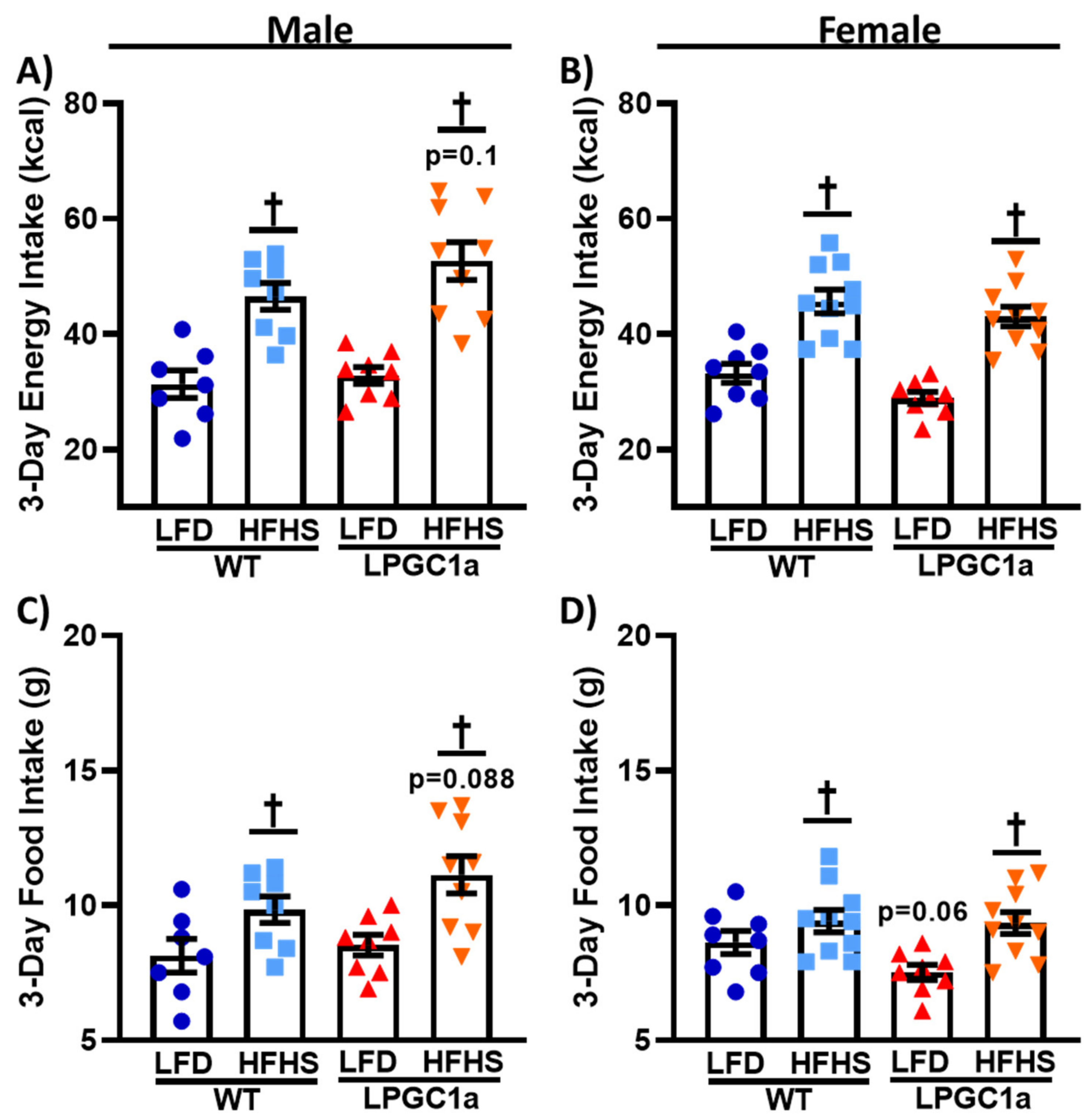
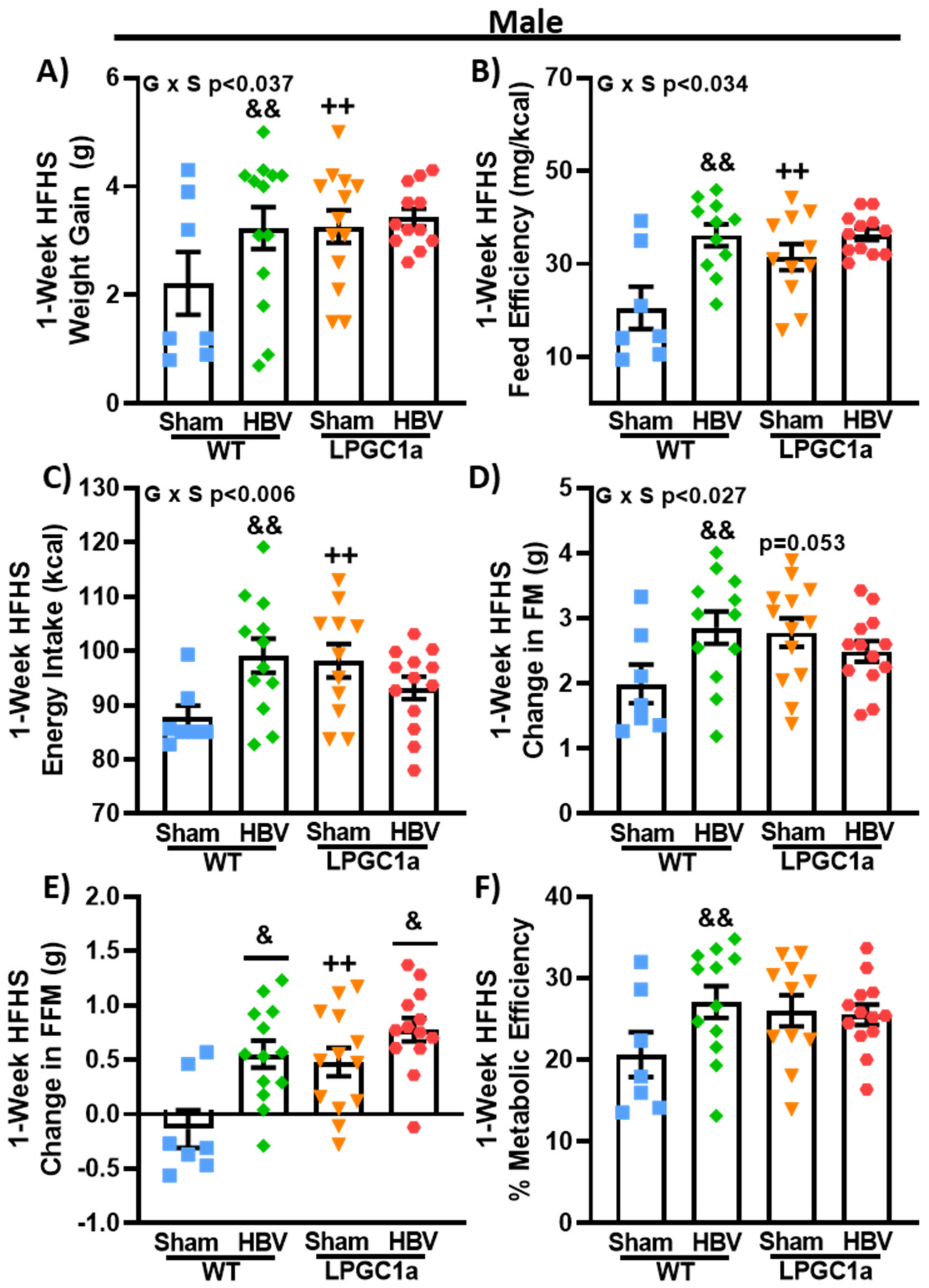
3.3. Male LPGC1a Mice Have Slight Increases in HFHS Intake and Altered Feeding Behaviors
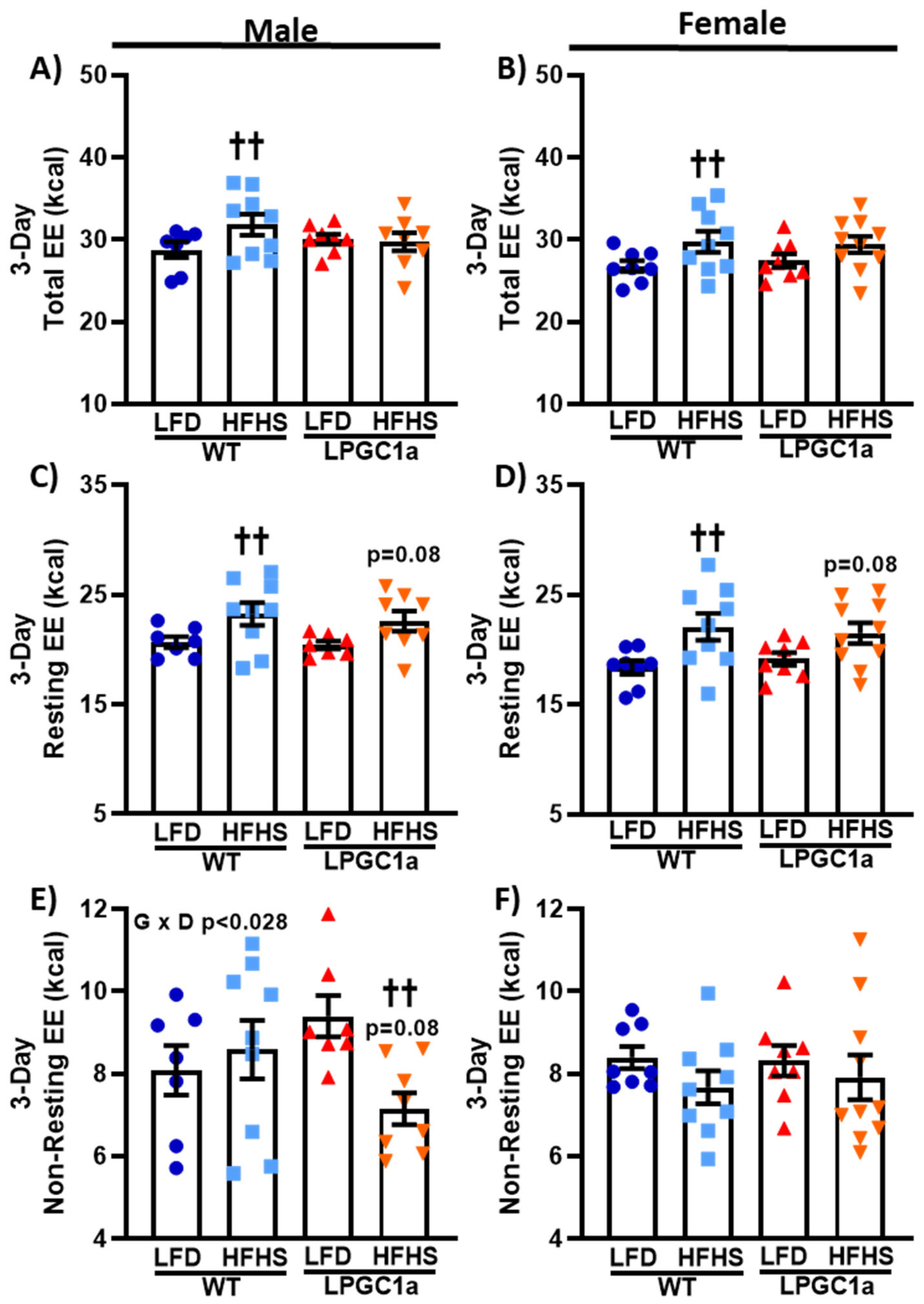
3.4. Lower Non-Resting EE in HFHS-Fed LPGC1a Male Mice Prevents HFHS-Induced Total EE Increase
3.5. Male LPGC1a Mice Fed a HFHS Diet Have Reduced Activity EE and Home Cage Activity
3.6. Common Hepatic Branch Vagotomy Increases Short-Term HFHS-Induced Weight Gain in WT Mice, but Not in LPGC1a Mice
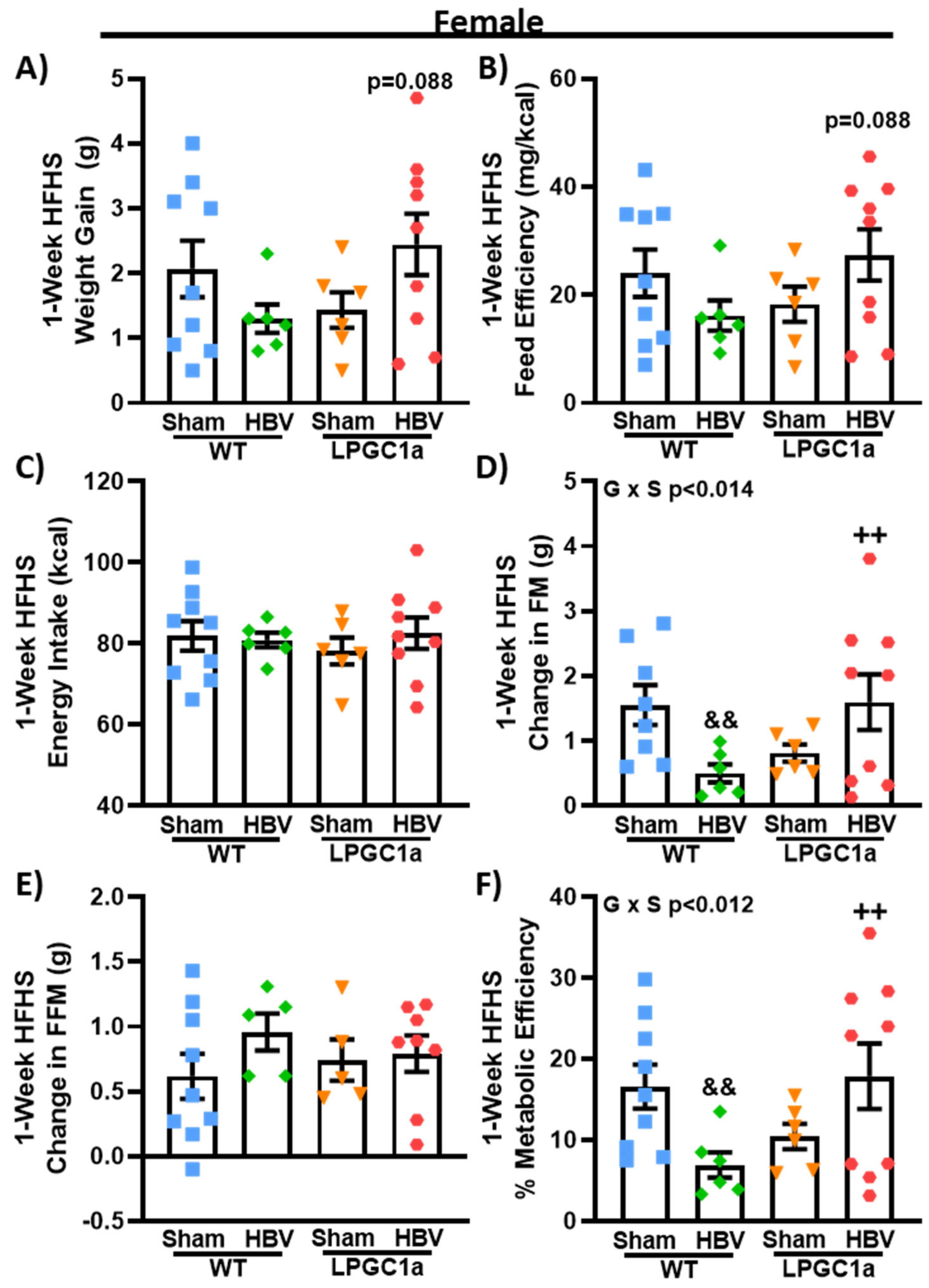
3.7. Common Hepatic Branch Vagotomy Divergently Impacts Short-Term HFHS-Induced Fat Mass Gains in Female WT and LPGC1a Mice
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ogden, C.L.; Fryar, C.D.; Martin, C.B.; Freedman, D.S.; Carroll, M.D.; Gu, Q.; Hales, C.M. Trends in Obesity Prevalence by Race and Hispanic Origin—1999–2000 to 2017–2018. JAMA 2020, 324, 1208. [Google Scholar] [CrossRef]
- Hill, J.O.; Wyatt, H.R.; Peters, J.C. Energy balance and obesity. Circulation 2012, 126, 126–132. [Google Scholar] [CrossRef]
- Racette, S.B.; Weiss, E.P.; Schechtman, K.B.; Steger-May, K.; Villareal, D.T.; Obert, K.A.; Holloszy, J.O. Influence of Weekend Lifestyle Patterns on Body Weight. Obesity 2008, 16, 1826–1830. [Google Scholar] [CrossRef]
- Hull, H.R.; Radley, D.; Dinger, M.K.; Fields, D.A. The effect of the Thanksgiving Holiday on weight gain. Nutr. J. 2006, 5, 29. [Google Scholar] [CrossRef] [PubMed]
- Yanovski, J.A.; Yanovski, S.Z.; Sovik, K.N.; Nguyen, T.T.; O’Neil, P.M.; Sebring, N.G. A Prospective Study of Holiday Weight Gain. N. Engl. J. Med. 2000, 342, 861–867. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Olendzki, B.C.; Li, W.; Hafner, A.R.; Chiriboga, D.; Hebert, J.R.; Campbell, M.; Sarnie, M.; Ockene, I.S. Seasonal variation in food intake, physical activity, and body weight in a predominantly overweight population. Eur. J. Clin. Nutr. 2005, 60, 519–528. [Google Scholar] [CrossRef]
- Galgani, J.; Ravussin, E. Energy metabolism, fuel selection and body weight regulation. Int. J. Obes. 2008, 32, S109–S119. [Google Scholar] [CrossRef]
- Hopkins, M.; Jeukendrup, A.; King, N.A.; Blundell, J.E. The relationship between substrate metabolism, exercise and appetite control: Does glycogen availability influence the motivation to eat, energy intake or food choice? Sports Med. 2011, 41, 507–521. [Google Scholar] [CrossRef]
- Hariri, N.; Thibault, L. High-fat diet-induced obesity in animal models. Nutr. Res. Rev. 2010, 23, 270–299. [Google Scholar] [CrossRef] [PubMed]
- Saper, C.B.; Chou, T.C.; Elmquist, J.K. The Need to Feed: Homeostatic and Hedonic Control of Eating. Neuron 2002, 36, 199–211. [Google Scholar] [CrossRef]
- Kim, K.-S.; Seeley, R.J.; Sandoval, D.A. Signalling from the periphery to the brain that regulates energy homeostasis. Nat. Rev. Neurosci. 2018, 19, 185–196. [Google Scholar] [CrossRef] [PubMed]
- Hand, G.A.; Shook, R.P.; Hill, J.O.; Giacobbi, P.R.; Blair, S.N. Energy flux: Staying in energy balance at a high level is necessary to prevent weight gain for most people. Expert Rev. Endocrinol. Metab. 2015, 10, 599–605. [Google Scholar] [CrossRef] [PubMed]
- Friedman, M.I.; Harris, R.B.; Ji, H.; Ramirez, I.; Tordoff, M.G. Fatty acid oxidation affects food intake by altering hepatic energy status. Am. J. Physiol. 1999, 276, R1046–R1053. [Google Scholar] [CrossRef]
- Horn, C.C.; Ji, H.; Friedman, M.I. Etomoxir, A fatty acid oxidation inhibitor, increases food intake and reduces hepatic energy status in rats. Physiol. Behav. 2004, 81, 157–162. [Google Scholar] [CrossRef]
- Horn, C.C.; Tordoff, M.G.; Friedman, M.I. Role of vagal afferent innervation in feeding and brain Fos expression produced by metabolic inhibitors. Brain Res. 2001, 919, 198–206. [Google Scholar] [CrossRef]
- Langhans, W.; Scharrer, E. Evidence for a vagally mediated satiety signal derived from hepatic fatty acid oxidation. J. Auton. Nerv. Syst. 1987, 18, 13–18. [Google Scholar] [CrossRef]
- Friedman, M.I.; Sawchenko, P.E. Evidence for hepatic involvement in control of ad libitum food intake in rats. Am. J. Physiol. Integr. Comp. Physiol. 1984, 247, R106–R113. [Google Scholar] [CrossRef]
- Estall, J.L.; Kahn, M.; Cooper, M.P.; Fisher, F.M.; Wu, M.K.; Laznik, D.; Qu, L.; Cohen, D.E.; Shulman, G.I.; Spiegelman, B.M. Sensitivity of Lipid Metabolism and Insulin Signaling to Genetic Alterations in Hepatic Peroxisome Proliferator-Activated Receptor- Coactivator-1 Expression. Diabetes 2009, 58, 1499–1508. [Google Scholar] [CrossRef]
- Morris, E.M.; Meers, G.M.E.; Booth, F.W.; Fritsche, K.L.; Hardin, C.D.; Thyfault, J.P.; Ibdah, J.A. PGC-1α overexpression results in increased hepatic fatty acid oxidation with reduced triacylglycerol accumulation and secretion. Am. J. Physiol. Gastrointest. Liver Physiol. 2012, 303, G979–G992. [Google Scholar] [CrossRef]
- Handschin, C.; Spiegelman, B.M. Peroxisome Proliferator-Activated Receptor γ Coactivator 1 Coactivators, Energy Homeostasis, and Metabolism. Endocr. Rev. 2006, 27, 728–735. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Marcos, P.J.; Auwerx, J. Regulation of PGC-1α, a nodal regulator of mitochondrial biogenesis. Am. J. Clin. Nutr. 2011, 93, 884S–890S. [Google Scholar] [CrossRef]
- Fletcher, J.A.; Linden, M.A.; Sheldon, R.D.; Meers, G.M.; Morris, E.M.; Butterfield, A.; Perfield, J.W., 2nd; Rector, R.S.; Thyfault, J.P. Fibroblast growth factor 21 increases hepatic oxidative capacity but not physical activity or energy expenditure in hepatic peroxisome proliferator-activated receptor γ coactivator-1α-deficient mice. Exp. Physiol. 2018, 103, 408–418. [Google Scholar] [CrossRef]
- Von Schulze, A.; McCoin, C.S.; Onyekere, C.; Allen, J.; Geiger, P.; Dorn, G.W., 2nd; Morris, E.M.; Thyfault, J.P. Hepatic mitochondrial adaptations to physical activity: Impact of sexual dimorphism, PGC1α and BNIP3-mediated mitophagy. J. Physiol. 2018, 596, 6157–6171. [Google Scholar] [CrossRef]
- Lin, J.; Wu, P.-H.; Tarr, P.T.; Lindenberg, K.S.; St-Pierre, J.; Zhang, C.-Y.; Mootha, V.K.; Jager, S.; Vianna, C.R.; Reznick, R.M.; et al. Defects in Adaptive Energy Metabolism with CNS-Linked Hyperactivity in PGC-1α Null Mice. Cell 2004, 119, 121–135. [Google Scholar] [CrossRef] [PubMed]
- Morris, E.M.; Jackman, M.R.; Meers, G.M.E.; Johnson, G.C.; Lopez, J.L.; MacLean, P.S.; Thyfault, J.P. Reduced hepatic mitochondrial respiration following acute high-fat diet is prevented by PGC-1α overexpression. Am. J. Physiol. Liver Physiol. 2013, 305, G868–G880. [Google Scholar] [CrossRef] [PubMed]
- Kaiyala, K.J.; Wisse, B.E.; Lighton, J.R.B. Validation of an equation for energy expenditure that does not require the respiratory quotient. PLoS ONE 2019, 14, e0211585. [Google Scholar] [CrossRef] [PubMed]
- Abreu-Vieira, G.; Xiao, C.; Gavrilova, O.; Reitman, M.L. Integration of body temperature into the analysis of energy expenditure in the mouse. Mol. Metab. 2015, 4, 461–470. [Google Scholar] [CrossRef]
- Hayes, M.R.; Kanoski, S.E.; De Jonghe, B.C.; Leichner, T.M.; Alhadeff, A.L.; Fortin, S.M.; Arnold, M.; Langhans, W.; Grill, H.J. The common hepatic branch of the vagus is not required to mediate the glycemic and food intake suppressive effects of glucagon-like-peptide-1. Am. J. Physiol. Integr. Comp. Physiol. 2011, 301, R1479–R1485. [Google Scholar] [CrossRef]
- Luijten, I.H.; Brooks, K.; Boulet, N.; Shabalina, I.G.; Jaiprakash, A.; Carlsson, B.; Fischer, A.; Cannon, B.; Nedergaard, J. Glucocorticoid-Induced Obesity Develops Independently of UCP1. Cell Rep. 2019, 27, 1686–1698.e5. [Google Scholar] [CrossRef]
- Langhans, W.; Egli, G.; Scharrer, E. Selective hepatic vagotomy eliminates the hypophagic effect of different metabolites. J. Auton. Nerv. Syst. 1985, 13, 255–262. [Google Scholar] [CrossRef]
- Lee, K.C.; Miller, R.E. The Hepatic Vagus Nerve and the Neural Regulation of Insulin Secretion. Endocrinology 1985, 117, 307–314. [Google Scholar] [CrossRef]
- Nagase, H.; Inoue, S.; Tanaka, K.; Takamura, Y.; Niijima, A. Hepatic glucose-sensitive unit regulation of glucose-induced insulin secretion in rats. Physiol. Behav. 1993, 53, 139–143. [Google Scholar] [CrossRef]
- Dean, J.T.; Tran, L.; Beaven, S.; Tontonoz, P.; Reue, K.; Dipple, K.M.; Liao, J.C. Resistance to Diet-Induced Obesity in Mice with Synthetic Glyoxylate Shunt. Cell Metab. 2009, 9, 525–536. [Google Scholar] [CrossRef]
- Orellana-Gavaldà, J.M.; Herrero, L.; Malandrino, M.I.; Pañeda, A.; Rodríguez-Peña, M.S.; Petry, H.; Asins, G.; Van Deventer, S.; Hegardt, F.G.; Serra, D. Molecular therapy for obesity and diabetes based on a long-term increase in hepatic fatty-acid oxidation. Hepatol. 2010, 53, 821–832. [Google Scholar] [CrossRef]
- Seok, S.; Kim, Y.-C.; Byun, S.; Choi, S.; Xiao, Z.; Iwamori, N.; Zhang, Y.; Wang, C.; Ma, J.; Ge, K.; et al. Fasting-induced JMJD3 histone demethylase epigenetically activates mitochondrial fatty acid β-oxidation. J. Clin. Investig. 2018, 128, 3144–3159. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wong, K.; Giles, A.; Jiang, J.; Lee, J.W.; Adams, A.C.; Kharitonenkov, A.; Yang, Q.; Gao, B.; Guarente, L.; et al. Hepatic SIRT1 Attenuates Hepatic Steatosis and Controls Energy Balance in Mice by Inducing Fibroblast Growth Factor 21. Gastroenterology 2014, 146, 539–549.e7. [Google Scholar] [CrossRef]
- Lee, H.; Ahn, J.; Shin, S.S.; Yoon, M. Ascorbic acid inhibits visceral obesity and nonalcoholic fatty liver disease by activating peroxisome proliferator-activated receptor α in high-fat-diet-fed C57BL/6J mice. Int. J. Obes. 2018, 43, 1620–1630. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Godlewski, G.; Jourdan, T.; Liu, Z.; Cinar, R.; Xiong, K.; Kunos, G. Cannabinoid-1 Receptor Antagonism Improves Glycemic Control and Increases Energy Expenditure Through Sirtuin-1/Mechanistic Target of Rapamycin Complex 2 and 5′Adenosine Monophosphate–Activated Protein Kinase Signaling. Hepatology 2019, 69, 1535–1548. [Google Scholar] [CrossRef] [PubMed]
- Mansouri, A.; Pacheco-López, G.; Ramachandran, D.; Arnold, M.; Leitner, C.; Prip-Buus, C.; Langhans, W.; Morral, N. Enhancing hepatic mitochondrial fatty acid oxidation stimulates eating in food-deprived mice. Am. J. Physiol. Integr. Comp. Physiol. 2015, 308, R131–R137. [Google Scholar] [CrossRef]
- Morris, E.M.; Jackman, M.R.; Johnson, G.C.; Liu, T.-W.; Lopez, J.L.; Kearney, M.L.; Fletcher, J.; Meers, G.M.E.; Koch, L.G.; Britton, S.L.; et al. Intrinsic aerobic capacity impacts susceptibility to acute high-fat diet-induced hepatic steatosis. Am. J. Physiol. Metab. 2014, 307, E355–E364. [Google Scholar] [CrossRef]
- Morris, E.M.; Meers, G.M.; Koch, L.G.; Britton, S.L.; MacLean, P.S.; Thyfault, J.P. Increased aerobic capacity reduces susceptibility to acute high-fat diet-induced weight gain. Obesity 2016, 24, 1929–1937. [Google Scholar] [CrossRef]
- Thyfault, J.P.; Morris, E.M. Intrinsic (Genetic) Aerobic Fitness Impacts Susceptibility for Metabolic Disease. Exerc. Sport Sci. Rev. 2017, 45, 7–15. [Google Scholar] [CrossRef]
- Friedman, M.I. Obesity and the hepatic control of feeding behavior. Drug News Perspect. 2007, 20, 573–578. [Google Scholar] [CrossRef]
- Thupari, J.N.; Landree, L.E.; Ronnett, G.V.; Kuhajda, F.P. C75 increases peripheral energy utilization and fatty acid oxidation in diet-induced obesity. Proc. Natl. Acad. Sci. USA 2002, 99, 9498–9502. [Google Scholar] [CrossRef] [PubMed]
- Thupari, J.N.; Kim, E.-K.; Moran, T.H.; Ronnett, G.V.; Kuhajda, F.P. Chronic C75 treatment of diet-induced obese mice increases fat oxidation and reduces food intake to reduce adipose mass. Am. J. Physiol. Metab. 2004, 287, E97–E104. [Google Scholar] [CrossRef] [PubMed]
- Rawson, N.E.; Blum, H.; Osbakken, M.D.; Friedman, M.I. Hepatic phosphate trapping, decreased ATP, and increased feeding after 2,5-anhydro-D-mannitol. Am. J. Physiol. Integr. Comp. Physiol. 1994, 266, R112–R117. [Google Scholar] [CrossRef]
- Cork, S.C. The role of the vagus nerve in appetite control: Implications for the pathogenesis of obesity. J. Neuroendocr. 2018, 30, e12643. [Google Scholar] [CrossRef] [PubMed]
- Lowell, B.B.; Spiegelman, B.M. Towards a molecular understanding of adaptive thermogenesis. Nat. Cell Biol. 2000, 404, 652–660. [Google Scholar] [CrossRef] [PubMed]
- Bachman, E.S.; Dhillon, H.; Zhang, C.-Y.; Cinti, S.; Bianco, A.C.; Kobilka, B.K.; Lowell, B.B. beta AR Signaling Required for Diet-Induced Thermogenesis and Obesity Resistance. Science 2002, 297, 843–845. [Google Scholar] [CrossRef] [PubMed]
- Kazak, L.; Chouchani, E.T.; Lu, G.Z.; Jedrychowski, M.P.; Bare, C.J.; Mina, A.I.; Kumari, M.; Zhang, S.; Vuckovic, I.; Laznik-Bogoslavski, D.; et al. Genetic Depletion of Adipocyte Creatine Metabolism Inhibits Diet-Induced Thermogenesis and Drives Obesity. Cell Metab. 2017, 26, 693. [Google Scholar] [CrossRef] [PubMed]
- Jung, R.T.; Shetty, P.S.; James, W.P.T.; Barrand, M.A.; Callingham, B.A. Reduced thermogenesis in obesity. Nat. Cell Biol. 1979, 279, 322–323. [Google Scholar] [CrossRef]
- Shetty, P.S.; Jung, R.T.; James, W.P. Reduced dietary-induced thermogenesis in obese subjects before and after weight loss. Proc. Nutr. Soc. 1979, 38, 87A. [Google Scholar] [PubMed]
- Nagata, N.; Matsuo, K.; Bettaieb, A.; Bakke, J.; Matsuo, I.; Graham, J.; Xi, Y.; Liu, S.; Tomilov, A.; Tomilova, N.; et al. Hepatic Src Homology Phosphatase 2 Regulates Energy Balance in Mice. Endocrinology 2012, 153, 3158–3169. [Google Scholar] [CrossRef][Green Version]
- De Lartigue, G. Role of the vagus nerve in the development and treatment of diet-induced obesity. J. Physiol. 2016, 594, 5791–5815. [Google Scholar] [CrossRef]
- Banni, S.; Carta, G.; Murru, E.; Cordeddu, L.; Giordano, E.; Marrosu, F.; Puligheddu, M.; Floris, G.; Asuni, G.P.; Cappai, A.L.; et al. Vagus Nerve Stimulation Reduces Body Weight and Fat Mass in Rats. PLoS ONE 2012, 7, e44813. [Google Scholar] [CrossRef] [PubMed]
- Yao, G.; Kang, L.; Li, J.; Long, Y.; Wei, H.; Ferreira, C.A.; Jeffery, J.J.; Lin, Y.; Cai, W.; Wang, X. Effective weight control via an implanted self-powered vagus nerve stimulation device. Nat. Commun. 2018, 9, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Shikora, S.A.; Wolfe, B.M.; Apovian, C.M.; Anvari, M.; Sarwer, D.B.; Gibbons, R.D.; Ikramuddin, S.; Miller, C.J.; Knudson, M.B.; Tweden, K.S.; et al. Sustained Weight Loss with Vagal Nerve Blockade but Not with Sham: 18-Month Results of the ReCharge Trial. J. Obes. 2015, 2015, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Morton, J.M.; Shah, S.N.; Wolfe, B.M.; Apovian, C.M.; Miller, C.J.; Tweden, K.S.; Billington, C.J.; Shikora, S.A. Effect of Vagal Nerve Blockade on Moderate Obesity with an Obesity-Related Comorbid Condition: The ReCharge Study. Obes. Surg. 2016, 26, 983–989. [Google Scholar] [CrossRef]
- Geisler, C.E.; Ghimire, S.; Hepler, C.; Miller, K.E.; Bruggink, S.M.; Kentch, K.P.; Higgins, M.R.; Banek, C.T.; Yoshino, J.; Klein, S.; et al. Hepatocyte membrane potential regulates serum insulin and insulin sensitivity by altering hepatic GABA release. Cell Rep. 2021, 35, 109298. [Google Scholar] [CrossRef] [PubMed]
- La Fleur, S.E.; Ji, H.; Manalo, S.L.; Friedman, M.I.; Dallman, M.F. The hepatic vagus mediates fat-induced inhibition of diabetic hyperphagia. Diabetes 2003, 52, 2321–2330. [Google Scholar] [CrossRef]
- McDougle, M.; Quinn, D.; Diepenbroek, C.; Singh, A.; de la Serre, C.; de Lartigue, G. Intact vagal gut-brain signalling prevents hyperphagia and excessive weight gain in response to high-fat high-sugar diet. Acta Physiol. 2021, 231, e13530. [Google Scholar] [CrossRef] [PubMed]
- Langhans, W.; Leitner, C.; Arnold, M. Dietary fat sensing via fatty acid oxidation in enterocytes: Possible role in the control of eating. Am. J. Physiol. Integr. Comp. Physiol. 2011, 300, R554–R565. [Google Scholar] [CrossRef] [PubMed]
- Clara, R.; Schumacher, M.; Ramachandran, D.; Fedele, S.; Krieger, J.-P.; Langhans, W.; Mansouri, A. Metabolic Adaptation of the Small Intestine to Short- and Medium-Term High-Fat Diet Exposure. J. Cell. Physiol. 2017, 232, 167–175. [Google Scholar] [CrossRef]
- McCoin, C.S.; Von Schulze, A.; Allen, J.; Fuller, K.N.Z.; Xia, Q.; Koestler, D.C.; Houchen, C.J.; Maurer, A.; Dorn, G.W.; Shankar, K.; et al. Sex modulates hepatic mitochondrial adaptations to high-fat diet and physical activity. Am. J. Physiol. Metab. 2019, 317, E298–E311. [Google Scholar] [CrossRef]
- Berthoud, H.-R. Anatomy and function of sensory hepatic nerves. Anat. Rec. Adv. Integr. Anat. Evol. Biol. 2004, 280, 827–835. [Google Scholar] [CrossRef] [PubMed]
- Jensen-Cody, S.O.; Potthoff, M.J. Hepatokines and metabolism: Deciphering communication from the liver. Mol. Metab. 2021, 44, 101138. [Google Scholar] [CrossRef]
- Nahon, J.-L.; Tratner, I.; Poliard, A.; Presse, F.; Poiret, M.; Gal, A.; Sala-Trepat, J.M.; Legrès, L.; Feldmann, G.; Bernuau, D. Albumin and alpha-fetoprotein gene expression in various nonhepatic rat tissues. J. Biol. Chem. 1988, 263, 11436–11442. [Google Scholar] [CrossRef]
- Martin, S.L.; Epperson, L.E.; Rose, J.C.; Kurtz, C.C.; Ané, C.; Carey, H.V. Proteomic analysis of the winter-protected phenotype of hibernating ground squirrel intestine. Am. J. Physiol. Integr. Comp. Physiol. 2008, 295, R316–R328. [Google Scholar] [CrossRef] [PubMed]
- Randich, A.; Spraggins, D.S.; Cox, J.E.; Meller, S.T.; Kelm, G.R. Jejunal or portal vein infusions of lipids increase hepatic vagal afferent activity. NeuroReport 2001, 12, 3101–3105. [Google Scholar] [CrossRef]
- Yan, Z.; Yan, H.; Ou, H. Human thyroxine binding globulin (TBG) promoter directs efficient and sustaining transgene expression in liver-specific pattern. Gene 2012, 506, 289–294. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genotyping | Forward Primer | Reverse Primer | |
|---|---|---|---|
| Tail | LoxP sites | TCC AGT AGG CAG AGA TTT ATG AC | TGT CTG GTT TGA CAA TCT GCT AGG TC |
| Alb-cre | TTA GAG GGG AAC AGC TCC AGA TGG | GTG AAA CAG CAT TGC TGT CAC TT | |
| Liver | WT PGC1a | CCA GTT TCT TCA TTG GTG TG | ACC TGT CTT TGC CTA TGA TTC |
| Mutant PGC1a | TCC AGT AGG CAG AGA TTT ATG AC | CCA ACT GTC TAT AAT TCC AGT TC | |
| RT-PCR | |||
| PGC1a (exon 3–5) | AGC CGT GAC CAC TGA CAA CGA G | GCT GCA TGG TTC TGA GTG CTA AG | |
| WT | LPGC1a | |||
|---|---|---|---|---|
| 3-Day Weight Gain Experiments | LFD | HFHS | LFD | HFHS |
| Body Weight | ||||
| Male | 30.1 ± 0.8 | 28.9 ± 1.1 | 31.8 ± 1.4 | 31.2 ± 1.0 |
| Female | 22.7 ± 1.0 | 23.9 ± 1.0 | 23.3 ± 0.9 | 22.7 ± 0.8 |
| HBV Experiments 7-Days HFHS | Sham | HBV | Sham | HBV |
| Male | ||||
| Body Weight | 25.5 ± 0.6 | 24.9 ± 0.3 | 27.0 ± 0.6 | 24.5 ± 0.4 && |
| Fat Mass | 2.54 ± 0.33 | 2.40 ± 0.24 | 2.88 ± 0.54 | 2.19 ± 0.10 |
| Fat-Free Mass | 22.96 ± 0.69 | 22.44 ± 0.33 | 22.91 ± 0.44 | 22.25 ± 0.37 |
| Female | ||||
| Body Weight | 20.2 ± 0.3 | 19.7 ± 0.1 | 20.1 ± 0.1 | 20.3 ± 0.6 |
| Fat Mass | 1.52 ± 0.09 | 1.39 ± 0.08 | 1.32 ± 0.12 | 1.79 ± 0.24 |
| Fat-Free Mass | 18.70 ± 0.27 | 18.35 ± 0.11 | 18.80 ± 0.24 | 18.48 ± 0.43 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morris, E.M.; Noland, R.D.; Ponte, M.E.; Montonye, M.L.; Christianson, J.A.; Stanford, J.A.; Miles, J.M.; Hayes, M.R.; Thyfault, J.P. Reduced Liver-Specific PGC1a Increases Susceptibility for Short-Term Diet-Induced Weight Gain in Male Mice. Nutrients 2021, 13, 2596. https://doi.org/10.3390/nu13082596
Morris EM, Noland RD, Ponte ME, Montonye ML, Christianson JA, Stanford JA, Miles JM, Hayes MR, Thyfault JP. Reduced Liver-Specific PGC1a Increases Susceptibility for Short-Term Diet-Induced Weight Gain in Male Mice. Nutrients. 2021; 13(8):2596. https://doi.org/10.3390/nu13082596
Chicago/Turabian StyleMorris, E. Matthew, Roberto D. Noland, Michael E. Ponte, Michelle L. Montonye, Julie A. Christianson, John A. Stanford, John M. Miles, Matthew R. Hayes, and John P. Thyfault. 2021. "Reduced Liver-Specific PGC1a Increases Susceptibility for Short-Term Diet-Induced Weight Gain in Male Mice" Nutrients 13, no. 8: 2596. https://doi.org/10.3390/nu13082596
APA StyleMorris, E. M., Noland, R. D., Ponte, M. E., Montonye, M. L., Christianson, J. A., Stanford, J. A., Miles, J. M., Hayes, M. R., & Thyfault, J. P. (2021). Reduced Liver-Specific PGC1a Increases Susceptibility for Short-Term Diet-Induced Weight Gain in Male Mice. Nutrients, 13(8), 2596. https://doi.org/10.3390/nu13082596