
Influence of Dietary Polar Lipid Supplementation on Memory and Longitudinal Brain Development

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Housing
2.2. Dietary Groups
2.3. Imaging Acquisition
2.4. Behavioral Testing
2.5. Statistical Analysis
3. Results
3.1. Growth and Intake
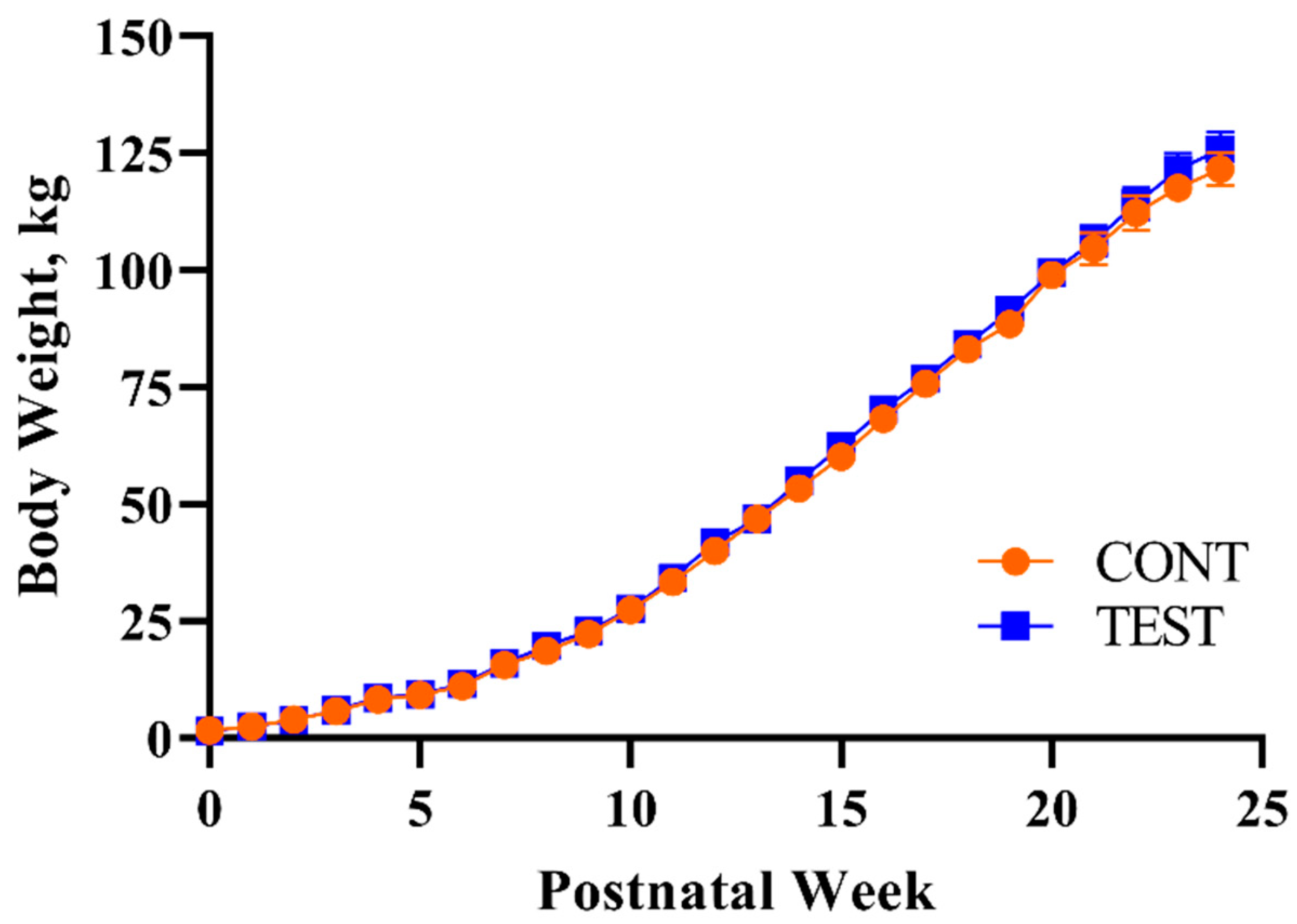
3.1.1. Body Weight
3.1.2. Feed Intake
3.1.3. Growth Performance
3.2. Neuroimaging Outcomes
3.3. Behavioral Outcomes
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Walker, A. Breast milk as the gold standard for protective nutrients. J. Pediatr. 2010, 156, S3–S7. [Google Scholar] [CrossRef] [PubMed]
- Deoni, S.C.L.; Dean, D.C.; Piryatinsky, I.; O’Muircheartaigh, J.; Waskiewicz, N.; Lehman, K.; Han, M.; Dirks, H. Breastfeeding and early white matter development: A cross-sectional study. Neuroimage 2013, 82, 77–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thakkar, S.K.; Giuffrida, F.; Cristina, C.H.; De Castro, C.A.; Mukherjee, R.; Tran, L.A.; Steenhout, P.; Lee, L.Y.; Destaillats, F. Dynamics of human milk nutrient composition of women from singapore with a special focus on lipids. Am. J. Hum. Biol. 2013, 25, 770–779. [Google Scholar] [CrossRef] [PubMed]
- Anto, L.; Warykas, S.W.; Torres-Gonzalez, M.; Blesso, C.N. Milk polar lipids: Underappreciated lipids with emerging health benefits. Nutrients 2020, 12, 1001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rombaut, R.; Dewettinck, K. Properties, analysis and purification of milk polar lipids. Int. Dairy J. 2006, 16, 1362–1373. [Google Scholar] [CrossRef]
- Zheng, L.; Fleith, M.; Giuffrida, F.; O’Neill, B.V.; Schneider, N. Dietary polar lipids and cognitive development: A narrative review. Adv. Nutr. 2019, 10, 1163–1176. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, M.U.; Ali, S.M.; Ahmad, A.; Sheikh, S.; Chen, P.; Ahmad, I. Polar lipids: Phospholipids and glycolipids. Bailey’s Ind. Oil Fat Prod. 2020, 1–27. [Google Scholar] [CrossRef]
- Mathai, J.C.; Tristram-Nagle, S.; Nagle, J.F.; Zeidel, M.L. Structural determinants of water permeability through the lipid membrane. J. Gen. Physiol. 2008, 131, 69–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alberts, B.; Johnson, A.; Lewis, J.; Raff, M.; Roberts, K.; Walter, P. The lipid bilayer. In Molecular Biology of the Cell; Garland Science: New York, NY, USA, 2002. [Google Scholar]
- Exton, J.H.; Taylor, S.J.; Augert, G.; Bocckino, S.B. Cell signalling through phospholipid breakdown. Mollecular Cell. Biol. 1991, 104, 81–86. [Google Scholar] [CrossRef]
- Vance, J.E.; Steenbergen, R. Metabolism and functions of phosphatidylserine. Prog. Lipid Res. 2005, 44, 207–234. [Google Scholar] [CrossRef]
- Zassler, B.; Schermer, C.; Humpel, C. Protein kinase C and phosphoinositol-3-kinase mediate differentiation or proliferation of slice-derived rat microglia. Pharmacology 2003, 67, 211–215. [Google Scholar] [CrossRef] [PubMed]
- Spitsberg, V.L. Invited Review: Bovine milk fat globule membrane as a potential nutraceutical. J. Dairy Sci. 2005, 88, 2289–2294. [Google Scholar] [CrossRef]
- Timby, N.; Domellöf, E.; Hernell, O.; Lönnerdal, B.; Domellöf, M. Neurodevelopment, nutrition, and growth until 12 mo of age in infants fed a low-energy, low-protein formula supplemented with bovine milk fat globule membranes: A randomized controlled trial. Am. J. Clin. Nutr. 2014, 99, 860–868. [Google Scholar] [CrossRef] [Green Version]
- Brink, L.R.; Lönnerdal, B. The role of milk fat globule membranes in behavior and cognitive function using a suckling rat pup supplementation model. J. Nutr. Biochem. 2018, 58, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Gurnida, D.A.; Rowan, A.M.; Idjradinata, P.; Muchtadi, D.; Sekarwana, N. Association of complex lipids containing gangliosides with cognitive development of 6-month-old infants. Early Hum. Dev. 2012, 88, 595–601. [Google Scholar] [CrossRef]
- Meck, W.H.; Williams, C.L. Perinatal choline supplementation increases the threshold for chunking in spatial memory. Neuroreport 1997, 8, 3053–3059. [Google Scholar] [CrossRef]
- Meck, W.H.; Smith, R.A.; Williams, C.L. Organizational changes in cholinergic activity and enhanced visuospatial memory as a function of choline administered prenatally or postnatally or both. Behav. Neurosci. 1989, 103, 1234–1241. [Google Scholar] [CrossRef]
- Oshida, K.; Shimizu, T.; Takase, M.; Tamura, Y.; Shimizu, T.; Yamashiro, Y. Effects of dietary sphingomyelin on central nervous system myelination in developing rats. Pediatr. Res. 2003, 53, 589–593. [Google Scholar] [CrossRef] [Green Version]
- Deoni, S.; Dean, D.; Joelson, S.; O’Regan, J.; Schneider, N. Early nutrition influences developmental myelination and cognition in infants and young children. Neuroimage 2018, 1–52. [Google Scholar] [CrossRef]
- Sciascia, Q.; Daş, G.; Metges, C.C. Review: The pig as a model for humans: Effects of nutritional factors on intestinal function and health. J. Anim. Sci. 2016, 94, 441–452. [Google Scholar] [CrossRef] [Green Version]
- Pond, W.G.; Mersmann, H.J. Biology of the Domestic Pig; Comstock Pub. Associates, Cornell University Press: Ithaca, NY, USA, 2001. [Google Scholar]
- Miller, E.R.; Ullrey, D.E. The pig as a model for human nutrition. Annu. Rev. Nutr. 1987, 7, 361–382. [Google Scholar] [CrossRef]
- Lind, N.M.; Moustgaard, A.; Jelsing, J.; Vajta, G.; Cumming, P.; Hansen, A.K. The use of pigs in neuroscience: Modeling brain disorders. Neurosci. Biobehav. Rev. 2007, 31, 728–751. [Google Scholar] [CrossRef]
- Dickerson, J.W.T.; Dobbing, J. Prenatal and postnatal growth and development of the central nervous system of the pig. Proc. R. Soc. Lond. Ser. B. Biol. Sci. 1967, 166, 1–12. [Google Scholar] [CrossRef]
- Dobbing, J.; Sands, J. Comparative aspects of the brain growth spurt. Early Hum. Dev. 1979, 311, 79–83. [Google Scholar] [CrossRef]
- Mayhew, T.M.; Mwamengele, G.L.; Dantzer, V.; Williams, S. The gyrification of mammalian cerebral cortex: Quantitative evidence of anisomorphic surface expansion during phylogenetic and ontogenetic development. J. Anat. 1996, 188, 53–58. [Google Scholar]
- Centers for Disease Control and Prevention. Available online: https://www.cdc.gov/breastfeeding/data/reportcard (accessed on 4 July 2021).
- Reiland, S. Growth and skeletal development of the pig. Acta Radiol. Suppl. 1978, 358, 15–22. [Google Scholar] [PubMed]
- National Research Council. Nutrient Requirements of Swine; National Academies Press: Valencia, CA, USA, 2012. [Google Scholar]
- Fil, J.E.; Joung, S.; Hayes, C.A.; Dilger, R.N. Influence of rearing environment on longitudinal brain development, object recognition memory, and exploratory behaviors in the domestic Pig (Sus scrofa). Front. Neurosci. 2021, 1–16. [Google Scholar] [CrossRef]
- Fil, J.E.; Joung, S.; Zimmerman, B.J.; Sutton, B.P.; Dilger, R.N. High-resolution magnetic resonance imaging-based atlases for the young and adolescent domesticated pig ( Sus scrofa ). J. Neurosci. Methods 2021, 354, 109107. [Google Scholar] [CrossRef]
- Jenkinson, M.; Beckmann, C.F.; Behrens, T.E.J.; Woolrich, M.W.; Smith, S.M. FSL. Neuroimage 2012, 62, 782–790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deoni, S.C.L.; Mercure, E.; Blasi, A.; Gasston, D.; Thomson, A.; Johnson, M.; Williams, S.C.R.; Murphy, D.G.M. Mapping infant brain myelination with magnetic resonance imaging. J. Neurosci. 2011, 31, 784–791. [Google Scholar] [CrossRef] [Green Version]
- Fleming, S.A.; Dilger, R.N. Young pigs exhibit differential exploratory behavior during novelty preference tasks in response to age, sex, and delay. Behav. Brain Res. 2017, 321, 50–60. [Google Scholar] [CrossRef] [PubMed]
- Gompertz, B. On the nature of the function expressive of the law of human mortality, and on a new mode of determining the value of life contingencies. Philos. Trans. R. Soc. Lond. 1825, 115, 513–583. [Google Scholar] [CrossRef]
- Bleasdale, J.K.A.; Nelder, J.A. Plant population and crop yield. Nature 1960, 188, 342. [Google Scholar] [CrossRef]
- Richards, F.J. A flexible growth function for empirical use. J. Exp. Bot. 1959, 10, 290–301. [Google Scholar] [CrossRef]
- Stannard, C.J.; Williams, A.P.; Gibbs, P.A. Temperature/growth relationships for psychrotrophic food-spoilage bacteria. Food Microbiol. 1985, 2, 115–122. [Google Scholar] [CrossRef]
- Dean, D.C.I.; O’Muircheartaigh, J.; Dirks, H.; Waskiewicz, N.; Lehman, K.; Walker, L.; Han, M.; Deoni, S.C.L. Modeling healthy male white matter and myelin development: 3 through 60 months of age. Neuroimage 2014, 84, 742–752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conrad, M.S.; Dilger, R.N.; Johnson, R.W. Brain growth of the domestic pig (sus scrofa) from 2 to 24 weeks of age: A longitudinal MRI study. Dev. Neurosci. 2012, 34, 291–298. [Google Scholar] [CrossRef] [Green Version]
- Schwarz, G. Estimating the dimension of a model. Ann. Stat. 1978, 6, 461–464. [Google Scholar] [CrossRef]
- Bhinder, G.; Allaire, J.M.; Garcia, C.; Lau, J.T.; Chan, J.M.; Ryz, N.R.; Bosman, E.S.; Graef, F.A.; Crowley, S.M.; Celiberto, L.S.; et al. Milk fat globule membrane supplementation in formula modulates the neonatal gut microbiome and normalizes intestinal development. Sci. Rep. 2017, 7, 1–15. [Google Scholar] [CrossRef]
- Snow, D.R.; Ward, R.E.; Olsen, A.; Jimenez-Flores, R.; Hintze, K.J. Membrane-rich milk fat diet provides protection against gastrointestinal leakiness in mice treated with lipopolysaccharide. J. Dairy Sci. 2011, 94, 2201–2212. [Google Scholar] [CrossRef] [Green Version]
- Zhou, A.L.; Ward, R.E. Milk polar lipids modulate lipid metabolism, gut permeability, and systemic inflammation in high-fat-fed C57BL/6J ob/ob mice, a model of severe obesity. J. Dairy Sci. 2019, 102, 4816–4831. [Google Scholar] [CrossRef]
- Moukarzel, S.; Dyer, R.A.; Garcia, C.; Wiedeman, A.M.; Boyce, G.; Weinberg, J.; Keller, B.O.; Elango, R.; Innis, S.M. Milk fat globule membrane supplementation in formula-fed rat pups improves reflex development and may alter brain lipid composition. Sci. Rep. 2018, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, K.; Hosozawa, M.; Kudo, N.; Yoshikawa, N.; Hisata, K.; Shoji, H.; Shinohara, K.; Shimizu, T. The pilot study: Sphingomyelin-fortified milk has a positive association with the neurobehavioural development of very low birth weight infants during infancy, randomized control trial. Brain Dev. 2013, 35, 45–52. [Google Scholar] [CrossRef]
- Timby, N.; Domellöf, M.; Lönnerdal, B.; Hernell, O. Supplementation of infant formula with bovine milk fat globule membranes. Adv. Nutr. An. Int. Rev. J. 2017, 8, 351–355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Billeaud, C.; Puccio, G.; Saliba, E.; Guillois, B.; Vaysse, C.; Pecquet, S.; Steenhout, P. Safety and tolerance evaluation of milk fat globule membrane-enriched infant formulas: A randomized controlled multicenter non-inferiority trial in healthy term infants. Clin. Med. Insights Pediatr. 2014, 51–60. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Radlowski, E.C.; Conrad, M.S.; Li, Y.; Dilger, R.N.; Johnson, R.W. Early supplementation of phospholipids and gangliosides affects brain and cognitive development in neonatal piglets. J. Nutr. 2014, 144, 1903–1909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basser, P.J.; Pierpaoli, C. Microstructrual and physiological features of tissues elucidated by quantitative-diffusion-tensor MRI. J. Magn. Reson. 2011, 213, 560–570. [Google Scholar] [CrossRef] [PubMed]
- Melhem, E.R.; Mori, S.; Mukundan, G.; Kraut, M.A.; Pomper, M.G.; Van Zijl, P.C.M. Diffusion tensor MR imaging of the brain and white matter tractography. Am. J. Roentgenol. 2002, 178, 3–16. [Google Scholar] [CrossRef] [PubMed]
- Alonso-Ortiz, E.; Levesque, I.R.; Pike, G.B. MRI-based myelin water imaging: A technical review. Magn. Reson. Med. 2015, 73, 70–81. [Google Scholar] [CrossRef] [PubMed]
- MacKay, A.L.; Laule, C. Magnetic resonance of myelin water: An in vivo marker for myelin. Brain Plast. 2016, 2, 71–91. [Google Scholar] [CrossRef] [Green Version]
- Joung, S.; Fil, J.E.; Heckmann, A.B.; Kvistgaard, A.S.; Dilger, R.N. Early-life supplementation of bovine milk osteopontin supports neurodevelopment and influences exploratory behavior. Nutrients 2020, 12, 2206. [Google Scholar] [CrossRef]
- Vickers, M.H.; Guan, J.; Gustavsson, M.; Krägeloh, C.U.; Breier, B.H.; Davison, M.; Fong, B.; Norris, C.; McJarrow, P.; Hodgkinson, S.C. Supplementation with a mixture of complex lipids derived from milk to growing rats results in improvements in parameters related to growth and cognition. Nutr. Res. 2009, 29, 426–435. [Google Scholar] [CrossRef] [PubMed]
- Mika, A.; Gaffney, M.; Roller, R.; Hills, A.; Bouchet, C.A.; Hulen, K.A.; Thompson, R.S.; Chichlowski, M.; Berg, B.M.; Fleshner, M.; et al. Feeding the developing brain: Juvenile rats fed diet rich in prebiotics and bioactive milk fractions exhibit reduced anxiety-related behavior and modified gene expression in emotion circuits. Neurosci. Lett. 2018, 677, 103–109. [Google Scholar] [CrossRef] [PubMed]
- Garcia, C.; Antona, C.; Robert, B.; Lopez, C.; Armand, M. The size and interfacial composition of milk fat globules are key factors controlling triglycerides bioavailability in simulated human gastro-duodenal digestion. Food Hydrocoll. 2014, 35, 494–504. [Google Scholar] [CrossRef]
- Michalski, M.C.; Genot, C.; Gayet, C.; Lopez, C.; Fine, F.; Joffre, F.; Vendeuvre, J.L.; Bouvier, J.; Chardigny, J.M.; Raynal-Ljutovac, K. Multiscale structures of lipids in foods as parameters affecting fatty acid bioavailability and lipid metabolism. Prog. Lipid Res. 2013, 52, 354–373. [Google Scholar] [CrossRef] [Green Version]
- Michalski, M.-C.; Soares, A.F.; Lopez, C.; Leconte, N.; Briard, V.; Geloen, A. The supramolecular structure of milk fat influences plasma triacylglycerols and fatty acid profile in the rat. Eur. J. Nutr. 2006, 45, 215–224. [Google Scholar] [CrossRef]
- Bourlieu, C.; Michalski, M.-C. Structure-function relationship of the milk fat globule. Curr. Opin. Clin. Nutr. Metab. Care 2015, 18, 118–127. [Google Scholar] [CrossRef] [PubMed]
- Rahmann, H. Brain gangliosides and memory formation. Behav. Brain Res. 1995, 66, 105–116. [Google Scholar] [CrossRef]
- Ohmi, Y.; Tajima, O.; Ohkawa, Y.; Mori, A.; Sugiura, Y.; Furukawa, K.; Furukawa, K. Gangliosides play pivotal roles in the regulation of complement systems and in the maintenance of integrity in nerve tissues. Proc. Natl. Acad. Sci. USA 2009, 106, 22405–22410. [Google Scholar] [CrossRef] [Green Version]
- Yu, R.K.; Tsai, Y.-T.; Ariga, T. Functional roles of gangliosides in neurodevelopment—An overview of recent advances. Neurochem. Res. 2012, 37, 1230–1244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, E.J.; Suh, M.; Ramanujam, K.; Steiner, K.; Begg, D.; Clandinin, M.T. Diet-induced changes in membrane gangliosides in rat intestinal mucosa, plasma and brain. J. Pediatr. Gastroenterol. Nutr. 2005, 40, 487–495. [Google Scholar] [CrossRef] [PubMed]
- Ryan, J.M.; Rice, G.E.; Mitchell, M.D. The role of gangliosides in brain development and the potential benefits of perinatal supplementation. Nutr. Res. 2013, 33, 877–887. [Google Scholar] [CrossRef] [PubMed]
- Gardner, J.M.M.; Powell, C.A.; Baker-Henningham, H.; Walker, S.P.; Cole, T.J.; Grantham-McGregor, S.M. Zinc supplementation and psychosocial stimulation: Effects on the development of undernourished Jamaican children. Am. J. Clin. Nutr. 2005, 82, 399–405. [Google Scholar] [CrossRef]
- Timby, N.; Adamsson, M.; Domellöf, E.; Grip, T.; Hernell, O.; Lönnerdal, B.; Domellöf, M. Neurodevelopment and growth until 6.5 years of infants who consumed a low-energy, low-protein formula supplemented with bovine milk fat globule membranes: A randomized controlled trial. Am. J. Clin. Nutr. 2021, 113, 586–592. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
| Diet | Pooled | |||
|---|---|---|---|---|
| Measure | CONT | TEST | SEM | p-Value 2 |
| PNW 1–4 | ||||
| ADG, kg/day | 0.245 | 0.260 | 0.0108 | 0.233 |
| ADFI, kg solids/day | 0.260 | 0.280 | 0.0801 | 0.031 |
| G:F, kg BW:kg solids | 0.188 | 0.184 | 0.0049 | 0.556 |
| PNW 5–24 | ||||
| ADG, kg/day | 0.818 | 0.845 | 0.0361 | 0.355 |
| ADFI, kg solids/day | 1.904 | 2.000 | 0.1054 | 0.431 |
| G:F, kg BW:kg solids | 0.483 | 0.424 | 0.0637 | 0.455 |
| CONT | TEST | Pooled SEM | p-Value 2 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ROI Parameter 3 | a | b | g | a | b | g | a | b | g | a | b | g |
| Whole brain | 147,805.08 | 0.34 | 0.11 | 151,390.00 | 0.33 | 0.11 | 7221.096 | 0.021 | 0.010 | 0.634 | 0.722 | 0.987 |
| Grey matter | 60,903.83 | 0.20 | 0.11 | 62,357.33 | −0.07 | 0.13 | 5604.781 | 0.210 | 0.041 | 0.798 | 0.180 | 0.680 |
| White matter | 58,858.29 | 0.56 | 0.13 | 58,095.33 | 0.59 | 0.13 | 3168.560 | 0.027 | 0.010 | 0.812 | 0.329 | 0.515 |
| Cerebrospinal fluid | 29,378.14 | 1.13 | 0.17 | 26,629.08 | 1.09 | 0.17 | 2519.084 | 0.052 | 0.021 | 0.264 | 0.486 | 0.904 |
| Cerebral aqueduct | 82.46 | 0.04 | 0.09 | 64.92 | −0.14 | 0.15 | 14.475 | 0.173 | 0.080 | 0.238 | 0.321 | 0.488 |
| Corpus callosum | 1500.20 | 0.38 | 0.08 | 1711.90 | 0.47 | 0.07 | 391.500 | 0.081 | 0.018 | 0.594 | 0.265 | 0.707 |
| Cerebellum | 24,387.70 | 0.42 | 0.08 | 34,678.80 | 0.56 | 0.08 | 9305.100 | 0.107 | 0.018 | 0.281 | 0.215 | 0.874 |
| Fourth ventricle | 227.80 | −0.05 | 0.06 | 187.80 | −0.19 | 0.07 | 65.106 | 0.234 | 0.011 | 0.546 | 0.557 | 0.355 |
| Hypothalamus | 388.70 | 0.36 | 0.09 | 400.80 | 0.36 | 0.09 | 49.367 | 0.052 | 0.011 | 0.809 | 0.950 | 0.908 |
| Left caudate | 1102.40 | 0.32 | 0.06 | 1235.90 | 0.29 | 0.07 | 277.300 | 0.122 | 0.016 | 0.646 | 0.824 | 0.563 |
| Left hippocampus | 1388.50 | 0.41 | 0.08 | 1718.10 | 0.49 | 0.07 | 332.000 | 0.087 | 0.017 | 0.331 | 0.350 | 0.552 |
| Left inferior colliculus | 435.00 | 0.58 | 0.05 | 389.00 | 0.46 | 0.06 | 71.070 | 0.129 | 0.015 | 0.525 | 0.369 | 0.477 |
| Left internal capsule | 2021.30 | 0.26 | 0.08 | 2759.70 | 0.43 | 0.06 | 395.000 | 0.102 | 0.010 | 0.107 | 0.129 | 0.095 |
| Left olfactory bulb | 3432.50 | 0.36 | 0.09 | 3567.80 | 0.42 | 0.11 | 459.800 | 0.151 | 0.018 | 0.773 | 0.700 | 0.509 |
| Left putamen globus pallidus | 831.80 | 0.32 | 0.06 | 845.60 | 0.43 | 0.04 | 229.800 | 0.147 | 0.012 | 0.952 | 0.473 | 0.302 |
| Left superior colliculi | 1085.80 | 0.54 | 0.06 | 863.80 | 0.45 | 0.06 | 196.400 | 0.096 | 0.011 | 0.270 | 0.327 | 0.685 |
| Lateral ventricle | 2068.70 | 0.36 | 0.06 | 2330.30 | 0.43 | 0.06 | 439.900 | 0.101 | 0.019 | 0.558 | 0.471 | 0.972 |
| Medulla | 5210.30 | 0.60 | 0.08 | 8241.60 | 0.67 | 0.08 | 1995.800 | 0.149 | 0.018 | 0.d180 | 0.665 | 0.852 |
| Midbrain | 6179.30 | 0.33 | 0.09 | 6044.50 | 0.35 | 0.09 | 319.300 | 0.053 | 0.011 | 0.677 | 0.710 | 0.642 |
| Pons | 4237.10 | 0.53 | 0.08 | 3985.80 | 0.51 | 0.08 | 354.700 | 0.149 | 0.015 | 0.487 | 0.891 | 0.928 |
| Right caudate | 1256.10 | 0.39 | 0.06 | 2094.30 | 0.49 | 0.06 | 620.000 | 0.136 | 0.021 | 0.216 | 0.461 | 0.957 |
| Right hippocampus | 1152.20 | 0.37 | 0.08 | 1863.40 | 0.72 | 0.05 | 380.000 | 0.156 | 0.011 | 0.135 | 0.053 | 0.009 |
| Right inferior colliculus | 489.60 | 0.60 | 0.05 | 524.60 | 0.53 | 0.05 | 91.486 | 0.127 | 0.018 | 0.706 | 0.552 | 0.722 |
| Right internal capsule | 2134.60 | 0.34 | 0.07 | 2335.30 | 0.35 | 0.07 | 294.700 | 0.069 | 0.012 | 0.503 | 0.797 | 0.603 |
| Right olfactory bulb | 3687.60 | 0.32 | 0.20 | 3491.80 | 0.33 | 0.08 | 715.400 | 0.135 | 0.101 | 0.781 | 0.941 | 0.268 |
| Right putamen globus pallidus | 576.00 | 0.29 | 0.05 | 657.40 | 0.31 | 0.06 | 90.235 | 0.094 | 0.012 | 0.396 | 0.817 | 0.486 |
| Right superior colliculi | 811.10 | 0.41 | 0.07 | 1002.30 | 0.47 | 0.07 | 184.600 | 0.096 | 0.018 | 0.311 | 0.544 | 0.716 |
| Thalamus | 3563.40 | 0.36 | 0.06 | 3639.10 | 0.41 | 0.05 | 381.100 | 0.194 | 0.011 | 0.845 | 0.801 | 0.640 |
| CONT | TEST | Pooled SEM | p-Value 2 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ROI Parameter 3 | a | b | g | a | b | g | a | b | g | a | b | g |
| Corpus callosum | 0.33 | −1.01 | 1.02 | 0.33 | −1.01 | 1.01 | 0.006 | 0.005 | 0.007 | 0.554 | 0.105 | 0.128 |
| Cerebellum | 0.31 | −1.00 | 1.00 | 0.31 | −1.00 | 1.00 | 0.002 | 0.002 | 0.002 | 0.585 | 0.514 | 0.546 |
| Left caudate | 0.37 | −1.04 | 1.00 | 0.37 | −1.05 | 1.04 | 0.004 | 0.044 | 0.043 | 0.713 | 0.807 | 0.361 |
| Left hippocampus | 0.34 | −1.01 | 1.01 | 0.35 | −1.01 | 1.00 | 0.005 | 0.005 | 0.007 | 0.014 | 0.271 | 0.337 |
| Left internal capsule | 0.43 | −1.46 | 1.67 | 0.43 | −1.60 | 1.90 | 0.005 | 0.104 | 0.151 | 0.454 | 0.169 | 0.137 |
| Left side | 0.36 | −1.10 | 1.12 | 0.36 | −1.16 | 1.17 | 0.004 | 0.057 | 0.066 | 0.591 | 0.270 | 0.449 |
| Right caudate | 0.37 | −1.01 | 0.98 | 0.36 | −1.04 | 1.00 | 0.006 | 0.041 | 0.026 | 0.283 | 0.409 | 0.562 |
| Right hippocampus | 0.34 | −1.03 | 1.04 | 0.35 | −1.03 | 1.02 | 0.006 | 0.021 | 0.026 | 0.287 | 0.822 | 0.471 |
| Right internal capsule | 0.43 | −1.49 | 1.73 | 0.43 | −1.63 | 1.96 | 0.006 | 0.098 | 0.137 | 0.965 | 0.171 | 0.114 |
| Right side | 0.36 | −1.10 | 1.09 | 0.36 | −1.15 | 1.10 | 0.005 | 0.067 | 0.061 | 0.891 | 0.468 | 0.855 |
| Thalamus | 0.37 | −0.99 | 0.95 | 0.38 | −1.00 | 0.98 | 0.004 | 0.012 | 0.021 | 0.100 | 0.155 | 0.170 |
| FA mask | 0.34 | −1.00 | 1.00 | 0.34 | −1.00 | 1.00 | 0.003 | 0.001 | 0.001 | 0.861 | 0.530 | 0.442 |
| White matter mask | 0.36 | −1.15 | 1.19 | 0.36 | −1.17 | 1.18 | 0.005 | 0.049 | 0.058 | 0.746 | 0.818 | 0.917 |
| CONT | TEST | Pooled SEM | p-Value 2 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ROI Parameter 3 | a | b | g | a | b | g | a | b | g | a | b | g |
| Corpus callosum | 0.26 | −0.50 | 0.24 | 0.25 | −0.17 | 0.63 | 0.113 | 0.337 | 0.258 | 0.970 | 0.381 | 0.262 |
| Cerebellum | 0.16 | 1.64 | 4.01 | 6.36 | 15.64 | 2.46 | 4.484 | 13.335 | 1.352 | 0.321 | 0.460 | 0.368 |
| Combined cortex | 0.08 | −1.22 | 0.36 | 0.25 | −0.90 | 0.66 | 0.118 | 0.274 | 0.265 | 0.336 | 0.367 | 0.372 |
| Combined hippocampus | 0.12 | −0.71 | 0.33 | 0.10 | −0.70 | 0.29 | 0.032 | 0.142 | 0.066 | 0.581 | 0.944 | 0.659 |
| Combined internal capsule | 0.24 | −0.83 | 0.18 | 0.21 | −0.41 | 0.36 | 0.108 | 0.291 | 0.111 | 0.728 | 0.154 | 0.133 |
| Hypothalamus | 0.16 | 1.35 | 2.64 | 4.74 | −0.24 | 0.39 | 3.194 | 1.516 | 1.612 | 0.327 | 0.445 | 0.297 |
| Left cortex | 0.10 | −1.47 | 0.23 | 0.26 | 0.42 | 1.66 | 0.126 | 0.823 | 0.821 | 0.381 | 0.124 | 0.221 |
| Left hemisphere | 0.12 | −0.55 | 2.08 | 0.12 | −14.14 | 0.81 | 0.041 | 9.913 | 1.458 | 0.973 | 0.315 | 0.513 |
| Left hippocampus | 0.47 | −0.49 | 0.33 | 0.23 | 0.07 | 0.57 | 0.176 | 0.317 | 0.184 | 0.361 | 0.223 | 0.337 |
| Left inferior colliculus | 0.47 | 0.97 | 4.30 | 4.89 | 36.92 | 3.93 | 3.569 | 27.766 | 1.239 | 0.352 | 0.355 | 0.832 |
| Left internal capsule | 0.18 | −0.81 | 0.31 | 0.17 | −0.44 | 0.45 | 0.070 | 0.312 | 0.209 | 0.913 | 0.255 | 0.516 |
| Left olfactory bulb | 0.09 | −6.37 | 8.97 | 0.12 | −3.63 | 9.75 | 0.059 | 7.601 | 3.627 | 0.691 | 0.682 | 0.881 |
| Left putamen globus-pallidus | 0.21 | −0.37 | 0.30 | 6.28 | 16.77 | 0.49 | 4.497 | 12.274 | 0.136 | 0.330 | 0.317 | 0.262 |
| Left superior colliculus | 0.23 | 40.74 | 0.87 | 0.17 | 1.74 | 2.57 | 0.103 | 28.316 | 1.018 | 0.689 | 0.286 | 0.220 |
| Medulla | 0.18 | −2.52 | 9.56 | 5.80 | 40.49 | 6.00 | 4.162 | 32.027 | 1.665 | 0.331 | 0.334 | 0.144 |
| Midbrain | 0.16 | 0.99 | 2.58 | 0.10 | −0.37 | 1.49 | 0.038 | 1.215 | 1.296 | 0.247 | 0.355 | 0.482 |
| Pons | 0.09 | 5.99 | 10.57 | 6.41 | 39.20 | 12.26 | 3.789 | 29.250 | 5.239 | 0.254 | 0.359 | 0.752 |
| Right cortex | 0.08 | −1.26 | 0.37 | 0.05 | −8.54 | 0.36 | 0.043 | 5.427 | 0.059 | 0.642 | 0.333 | 0.910 |
| Right hemisphere | 0.09 | −1.29 | 0.49 | 0.09 | −0.96 | 0.55 | 0.015 | 0.203 | 0.118 | 0.780 | 0.178 | 0.698 |
| Right hippocampus | 0.09 | −0.80 | 0.35 | 0.11 | −0.71 | 0.25 | 0.024 | 0.120 | 0.060 | 0.533 | 0.621 | 0.274 |
| Right inferior colliculus | 0.18 | 1.46 | 4.96 | 0.21 | 0.60 | 6.21 | 0.102 | 3.420 | 2.973 | 0.866 | 0.862 | 0.771 |
| Right internal capsule | 0.47 | 0.05 | 0.13 | 0.24 | −0.42 | 0.33 | 0.153 | 0.302 | 0.120 | 0.131 | 0.134 | 0.104 |
| Right olfactory bulb | 0.11 | −5.24 | 7.32 | 0.11 | −1.91 | 6.96 | 0.048 | 4.758 | 2.221 | 0.997 | 0.465 | 0.900 |
| Right putamen globus-pallidus | 0.32 | −0.22 | 0.22 | 0.39 | −0.10 | 0.23 | 0.168 | 0.227 | 0.089 | 0.664 | 0.669 | 0.954 |
| Right superior colliculus | 0.09 | −0.26 | 1.13 | 0.13 | 0.02 | 1.42 | 0.040 | 0.475 | 0.628 | 0.498 | 0.403 | 0.385 |
| Thalamus | 0.25 | −0.66 | 0.23 | 0.11 | −0.58 | 0.33 | 0.101 | 0.234 | 0.083 | 0.329 | 0.678 | 0.389 |
| Whole brain | 0.11 | 1.53 | 3.57 | 0.13 | −0.92 | 0.82 | 0.053 | 2.120 | 2.144 | 0.820 | 0.425 | 0.379 |
| Diet | n | Mean | SEM | p-Value 2 |
|---|---|---|---|---|
| Week 4 | ||||
| CONT | 12 | 0.51 | 0.083 | 0.471 |
| TEST | 8 | 0.59 | 0.077 | 0.132 |
| Week 8 | ||||
| CONT | 15 | 0.53 | 0.068 | 0.357 |
| TEST | 10 | 0.53 | 0.087 | 0.355 |
| Measurements | RI | Total Object Visit Time, s | Number of All Object Visits, n | Mean Object Visit Time, s/Visit | Latency to First Object Visit, s * | Latency to Last Object Visit, s |
|---|---|---|---|---|---|---|
| Effect of diet | ||||||
| CONT | 0.56 | 47.6 | 11.5 | 3.8 | 4.7 | 229.6 |
| TEST | 0.53 | 62.0 | 11.7 | 4.6 | 10.4 | 257.9 |
| SEM | 0.059 | 11.28 | 1.50 | 0.68 | 2.29 | 18.46 |
| Effect of age | ||||||
| Week 4 | 0.51 | 59.0 | 12.0 | 4.6 | 10.7 | 239.6 |
| Week 8 | 0.56 | 50.7 | 11.2 | 3.8 | 4.4 | 247.9 |
| SEM | 0.065 | 10.57 | 1.21 | 0.71 | 2.20 | 20.18 |
| Interaction means | ||||||
| CONT:Week 4 | 0.50 | 56.0 | 12.3 | 4.2 | 4.0 a | 241.7 |
| CONT:Week 8 | 0.59 | 39.3 | 10.8 | 3.4 | 5.4 a | 217.4 |
| TEST:Week 4 | 0.53 | 61.9 | 11.7 | 5.0 | 17.4 b | 237.5 |
| TEST:Week 8 | 0.54 | 62.1 | 11.7 | 4.2 | 3.4 a | 278.4 |
| SEM | 0.592 | 15.36 | 1.77 | 1.03 | 3.28 | 26.67 |
| p-Value 2 | ||||||
| Diet | 0.860 | 0.354 | 0.922 | 0.406 | 0.450 | 0.120 |
| Age | 0.546 | 0.501 | 0.490 | 0.396 | 0.001 | 0.720 |
| Diet × Age | 0.633 | 0.491 | 0.465 | 0.993 | 0.004 | 0.165 |
| Measurements | Total Novel Object Visit Time, s | Number of Novel Object Visits, n | Mean Novel Object Visit Time, s/Visit | Latency to First Novel Object Visit, s * | Latency to Last Novel Object Visit, s |
|---|---|---|---|---|---|
| Effect of diet | |||||
| CONT | 24.9 | 5.8 | 3.7 | 11.1 | 193.9 |
| TEST | 37.2 | 5.7 | 6.0 | 26.1 | 221.4 |
| SEM | 7.45 | 0.82 | 1.16 | 7.92 | 25.28 |
| Effect of age | |||||
| Week 4 | 29.9 | 5.9 | 4.7 | 21.9 | 200.2 |
| Week 8 | 32.2 | 5.6 | 5.0 | 15.3 | 215.0 |
| SEM | 7.91 | 0.71 | 1.25 | 8.44 | 23.97 |
| Interaction means | |||||
| CONT:Week 4 | 29.1 | 6.2 | 4.0 | 13.6 | 184.8 |
| CONT:Week 8 | 20.7 | 5.5 | 3.5 | 8.7 | 202.9 |
| TEST:Week 4 | 30.7 | 5.6 | 5.4 | 30.3 | 215.6 |
| TEST:Week 8 | 43.6 | 5.8 | 6.6 | 21.8 | 227.1 |
| SEM | 11.07 | 1.03 | 1.68 | 11.08 | 32.47 |
| p-value 2 | |||||
| Diet | 0.193 | 0.899 | 0.073 | 0.294 | 0.345 |
| Age | 0.818 | 0.683 | 0.810 | 0.006 | 0.516 |
| Diet × Age | 0.291 | 0.539 | 0.552 | 0.308 | 0.886 |
| Measurements | Total Sample Object Visit Time, s | Number of Sample Object Visits, n | Mean Sample Object Visit Time, s/visit | Latency to First Sample Object Visit, s 3 | Latency to Last Sample Object Visit, s |
|---|---|---|---|---|---|
| Effect of diet | |||||
| CONT | 21.1 | 5.4 | 3.6 | 11.1 | 212.3 |
| TEST | 25.3 | 6.5 | 3.7 | 20.9 | 235.4 |
| SEM | 5.80 | 0.89 | 0.61 | 5.93 | 24.67 |
| Effect of age | |||||
| Week 4 | 26.3 | 6.3 | 4.2 | 22.8 | 223.9 |
| Week 8 | 20.1 | 5.5 | 3.1 | 9.3 | 223.8 |
| SEM | 5.33 | 0.97 | 0.59 | 5.43 | 25.74 |
| Interaction means | |||||
| CONT:Week 4 | 26.9 | 6.5 | 4.1 | 12.0 ab | 231.3 |
| CONT:Week 8 | 15.4 | 4.2 | 3.1 | 10.3 ab | 193.3 |
| TEST:Week 4 | 25.8 | 6.1 | 4.3 | 33.6 a | 216.4 |
| TEST:Week 8 | 24.9 | 6.9 | 3.2 | 8.3 b | 254.3 |
| SEM | 7.75 | 1.31 | 0.81 | 8.15 | 32.07 |
| p-Value 2 | |||||
| Diet | 0.599 | 0.266 | 0.888 | 0.415 | 0.269 |
| Age | 0.307 | 0.525 | 0.094 | 0.185 | 0.998 |
| Diet × Age | 0.378 | 0.211 | 0.948 | 0.024 | 0.134 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fil, J.E.; Joung, S.; Hauser, J.; Rytz, A.; Hayes, C.A.; Dilger, R.N. Influence of Dietary Polar Lipid Supplementation on Memory and Longitudinal Brain Development. Nutrients 2021, 13, 2486. https://doi.org/10.3390/nu13082486
Fil JE, Joung S, Hauser J, Rytz A, Hayes CA, Dilger RN. Influence of Dietary Polar Lipid Supplementation on Memory and Longitudinal Brain Development. Nutrients. 2021; 13(8):2486. https://doi.org/10.3390/nu13082486
Chicago/Turabian StyleFil, Joanne E., Sangyun Joung, Jonas Hauser, Andreas Rytz, Courtney A. Hayes, and Ryan N. Dilger. 2021. "Influence of Dietary Polar Lipid Supplementation on Memory and Longitudinal Brain Development" Nutrients 13, no. 8: 2486. https://doi.org/10.3390/nu13082486
APA StyleFil, J. E., Joung, S., Hauser, J., Rytz, A., Hayes, C. A., & Dilger, R. N. (2021). Influence of Dietary Polar Lipid Supplementation on Memory and Longitudinal Brain Development. Nutrients, 13(8), 2486. https://doi.org/10.3390/nu13082486