1. Introduction
Obesity is a worldwide health issue, and awareness of it has greatly increased in recent years. When obesity occurs, comorbidities follow, including hyperlipidemia, hyperglycemia, hypertension, and other metabolic abnormalities. White adipose tissues (WATs) are composed of white adipocytes, and play an important role in the body’s energy balance. Adipocytes were found to be over-expanded in obese individuals, while the expansion occurs in two different ways [
1]. In principle, hypertrophy is the enlargement of adipocyte size, and hyperplasia is an increase in adipocyte numbers. With the size expansion, the function of adipocytes is gradually lost, which is believed to cause metabolic abnormalities in individuals [
2]. For example, dyslipidemia, an abnormal accumulation of one or more kinds of lipoproteins, leads to changes in blood lipid concentrations. Evidence suggests that obesity is caused by excessive energy intake that leads to abdominal accumulation and possibly contributing to peripheral insulin resistance (IR) [
3,
4].
Torng et al. [
5] found that menopause in Taiwanese women had an adverse effect on blood lipids, especially increases in total cholesterol (TC) and low-density lipoprotein cholesterol (LDL-C), and a decrease in high-density lipoprotein cholesterol (HDL-C). In the process of menopause, the concentration of HDL-C changes in a U-shaped curve [
5]. The concentration of HDL-C reaches the highest during menopause and begins to decrease after menopause. At the same time, compared to before menopause, LDL-C, which causes atherosclerosis, appears in large amounts in postmenopausal women, resulting in a three-fold increase in the risk of cardiovascular diseases and coronary artery calcification. Based on past human and animal experiments, estrogen is believed to reduce food intake and increase energy consumption in the body, and is therefore related to preventing obesity [
6,
7]. The reduction in estrogen after menopause affects the location of fat storage in the body, resulting in increases in the overall fat mass and abdominal fat [
8]. Under the menopausal animal model, the concentration of free fatty acids (FAs, FFAs) in the body was significantly reduced, but after 60 days of estrogen supplementation, their concentration rose. At the same time, that study further pointed out that the accumulation of abdominal fat may be the result of abnormal lipolysis caused by low estrogen level [
9]. An ovariectomy (OVX), surgery to remove the ovaries, is a common menopausal animal model. Removal of the ovaries simulates a state of low estrogen and is often used to observe related effects of metabolism in the body in this state. In previous studies, it was found that 1 week after an OVX, OVX rats had a significant increase in body weight compared to rats that had undergone sham surgery. Even with the same food intake, OVX animals still tended to gain more body weight. This was accompanied by a decrease in energy consumption [
10] and accumulation of additional abdominal white fat [
11]. In the OVX animal model, low estrogen can easily cause a large amount of fat to accumulate in the liver. In previous literature, the occurrence of this phenomenon was associated with increases in fat synthesis pathways [
12] and decreases in liver fat oxidation pathways [
13]. At the same time, the loss of the protective effect of estrogen can easily cause an imbalance in the oxidative and antioxidant functions in the liver of OVX animals, resulting in increases in reactive oxygen species (ROS) [
14].
There are not many previous studies on the relationship between OVX animal models and the FA composition. Kim et al. [
15] reported that n-3 polyunsaturated FAs (PUFAs) given to OVX rats could reduce triglycerides (TGs). In that study, 48 mice were subjected to an OVX, and then after a 1-week recovery period, they were fed an AIN-93G diet the formula of which was adjusted to provide 0%, 1%, or 2% kcal eicosapentaenoic acid (EPA) + docosahexaenoic acid (DHA), in an experiment that lasted 12 weeks. Results showed that the expressions of sterol regulatory element-binding protein (SREBP)-1, acetyl-CoA carboxylase (ACC), FA synthase (FAS), and diacylglycerol acyltransferase 2 (DGAT2) in the liver were significantly reduced. Expressions of AMP-activated protein kinase (AMPK), phosphorylated AMPK, peroxisomal proliferator-activated receptor α (PPARα), and carnitine palmitoyltransferase (CPT)-1 in the liver and skeletal muscles significantly increased. The authors stated that n-3 PUFAs administered to OVX rats reduced TGs, which may have been due to reductions in TG biosynthesis pathways in the liver and increased oxidation of TGs [
15]. Liao et al. [
16] fed hamsters with different proportions of an FA diet, and found that there were no differences in food intake among the various groups. Hamsters fed experimental oil with a polyunsaturated/saturated FA (P/S) ratio of 5 and a monounsaturated FA (MUFA) ratio of 60% had a lower body weight (BW) and WAT weight, and had lower plasma insulin and free FA concentrations along with liver lipid-synthesizing enzyme activity. Activities of the liver FA oxidation-related enzymes, acyl-CoA oxidase (ACO) and CPT-1, increased, which inhibited body fat accumulation. In 2017, it was further discovered that experimental oil with an FA composition of P/S = 5 and a MUFA ratio of 60% could still effectively reduce lipid synthesis and promote lipid oxidation under a high-dose high-fat diet (HFD) to reduce body fat accumulation. This experimental oil seemed to prevent an increase in BW caused by the intake of medium-fat or HFDs by preventing increases in the insulin concentration, FAS and lipoprotein lipase (LPL) enzyme activities, and peroxisome proliferator-activated receptor (PPAR)-γ and LPL gene expressions [
17].
Although a lot of research has been conducted to study the effect of fat and its fatty acid composition on normal animal body weight, fat and glucose metabolism, the effect of fatty acid composition on OVX rats has not been studied. In this study, OVX rats were fed diets containing different doses of a specific proportion of experimental oil to explore its effects on WAT quality, blood lipid concentrations, liver lipid accumulation, and FA metabolism-related enzyme activities and gene expressions. Therefore, the results of this study can provide a reference for how to choose fatty acid composition during menopause.
2. Materials and Methods
2.1. FA Composition Analysis
Experimental oil was mixed following Liao’s method [
18]. Soybean oil and experimental oil mixture were purchased from a local supermarket and analyzed by gas chromatography (GC). Samples were extracted using a modified Folch method [
19]. Samples was extracted using chloroform/methanol = 2/1 (
v/
v) for 1 h, and then distilled water was added to separate the liquid. The extract was incubated for 10 min at room temperature and centrifuged at 4 °C and 3000 rpm for 10 min. The lower phase was collected. FA methylation was performed by heating the sample to 88 °C with 14% boron trifluoride/methanol (B1252, Sigma, St. Louis, MO, USA) for 1 h to form FA methyl ether (FAME) and then the solvent was removed. FAME was analyzed using a FOCUS™ GC (Thermo Fisher Scientific, Milan, Italy) equipped with a 30-m × 0.32-mm inner diameter (I.D.) × 0.20-μm df Rtx-2330 column (Restek, Bellefonte, PA, USA) and flame ionization detector.
FA compositions of the soybean oil and experimental oil were analyzed by GC. Results were obtained based on the retention time of the appropriate standard (GLC-455; Supelco, St. Louis, MO, USA), and percentages of the FA profile were calculated based on 16 different FAs, as shown in
Table 1.
2.2. Animals and Experimental Design
A total of 40 female Sprague-Darley rats were purchased from BioLasco (Taipei, Taiwan) at 9 weeks of age (ethics number LAC-2017-0443, Taipei, Taiwan). The study was conducted under strict guidelines of the institutional animal care and used committee of Taipei Medical University. Thirty-two rats underwent an OVX, while eight were sham-operated, and serum estradiol level was tested to ensure the effects of the surgery. Animals were housed in an air-conditioned room (22 ± 2 °C and 65% ± 5% relative humidity) with a 12 h light/12 h dark circle. All mice have free access to f water and a basic diet (Rodent Laboratory Chow 5001; PMI Nutrition International, St Louis, MO, USA). Rats were then communally housed in plastic cages (three per cage).
After a 4-week surgical recovery period, OVX rats were randomly divided into four different dietary groups (
n = 8 in each group). Hamsters were assigned to low-fat diet group (5%
w/
w soybean oil, 3.85 kcal/g) and high-fat diet group (35%
w/
w soybean oil, 5 kcal/g, 52% of energy) according to the AIN-93M formulation [
20] and modification. Therefore, we use 5% soybean oil as the control. One of the groups (C group) was fed the same diet as the sham group (S group) which contained 5% (
w/
w) fat from soybean oil. The other three groups were fed with 5%, 15%, and 20% (
w/
w) of the experimental oil mixture, namely L, M, and H groups, respectively. The diet consisted of a modified AIN-93 M formula, and the experimental oil mixture consisted of 60% MUFAs and P/S = 5 of a mixture of soybean and canola oils, as shown in
Table 2. All animals were fed ad libitum for 8 weeks. Body weight and food intake were measured weekly, and caloric intake and feed efficiency ratio (FER, %; food intake/body weight increment) were calculated. Each group (8 rats) was assigned to three cages, and total food intake of each rat was calculated as the total food amount of three cages divided by eight as average food intake. After 8 weeks of the experiment, the rats were starved for 12 h, and blood was collected under anesthesia with rompun (Bayer, Leverkusen, German) and zoletil (Virbac, Carros, France). Serum was collected centrifuged at 3500×
g for 20 min at 4 °C. The liver, kidneys, spleen, uterus, and WATs from gonadal, retroperitoneal, and perirenal locations were removed and weighed, and all samples were stored at −80 °C until analyzed.
2.3. Serum Measurements
Serum TC, TGs, HDL-C, LDL-C, nonesterified FAs, and glucose levels were analyzed by enzymatic colorimetric analysis using commercial enzyme kits (Randox Laboratory, Crumlin, Northland, UK); serum insulin concentration was analyzed using commercially available enzyme-linked immunosorbent assay (ELISA) kits (Mercodia, Uppsala, Sweden); serum leptin concentration was analyzed using a Mouse and Rat Leptin ELISA Kit (BioVender, Brno, Czech Republic); serum adiponectin concentration was analyzed using a Rat Adiponectin (ACRP30) ELISA Kit (AssayPro, St. Charles, MI, USA); serum estradiol concentration was analyzed using a Rat Estradiol ELISA Kit (Wuhan Fine Biotech, Wuhan, China); and serum follicular-stimulating hormone (FSH) concentration was analyzed using an FSH (Rodent) ELISA Kit (Wuhan Fine Biotech, Wuhan, China) using a VERSAmax microplate reader (Abnova, Taipei, Taiwan). The IR index was estimated by a homoeostasis model assessment (HOMA).
2.4. Hepatic Lipid Measurements
Total lipids were extracted from the liver, as previous method [
21]. Hepatic cholesterol and TG levels were estimated by commercial enzyme kits (Randox Laboratory); and hepatic nonesterified FA level was estimated by commercial enzyme kits (Randox Laboratory).
2.5. Real-Time Euantitative Polymerase Chain Reaction (qPCR)
Total RNA was extracted using Trizol reagent (Life Technologies, Carlsbad, CA, USA) from rat liver and gonadal adipose tissues. The mRNA levels of ACC, ACO, FAS, CPT, AMPK, PPARα, LPL, HSL and PPARγ were quantified by quantitative real-time PCR following Yang’s method [
17]. GAPDH mRNA was the internal control. The primers were shown in
Table 3.
2.6. Hepatic Enzyme Assay and Adipose Tissue Lipoprotein Lipase (LPL) Enzyme Assay
The activities of hepatic lipogenic enzymes (FAS, ACC, ACO, and CPT-I) were measured according to previous protocols [
22,
23,
24,
25]. The activity of adipose LPL was measured following Yang’s method [
17].
2.7. Statistical Analysis
Data are presented as the mean ± standard deviation (SD). Significant differences between the S and C groups were analyzed statistically by Student’s t-test to determine changes after an OVX. Significant changes between OVX groups were analyzed by a one-way analysis of variance (ANOVA). When significant changes were observed in ANOVA tests, Tukey’s tests were applied to locate the source of the significant difference. The significance level was set to p < 0.05. These calculations were performed using SAS version 9.4 (SAS Institute, Cary, NC, USA).
4. Discussion
In this study, OVX rats in the OVX control group (C group) had significantly lower estrogen level and lower uterine weight, accompanied by uterine atrophy, compared to the sham-operated (S) group, indicating that the operation successfully simulated low estrogen level in menopausal women. Past studies determined that ovarian-related hormones play important roles in regulating energy, appetite, and BW. The low estrogen status of postmenopausal women can easily lead to increased risks of obesity and metabolic-related abnormalities [
26]. Similar to results of human experiments, Iwasa et al. [
27] found that food intake and BW significantly increased after short-term suppression of ovarian hormone secretion in rats. However, the long-term effects of an OVX on energy metabolism in rats are still inconclusive [
28]. In our study, although there were no significant differences in food or energy intake between the OVX and sham (S) group rats, OVX rats still showed a significant increase in BW. This result is similar to experimental results of Jones et al. [
29]. Previous results showed that mice with low estrogen level caused by aromatase knockout had significantly heavier BWs and fat weight than wild-type mice, but no significant differences in food intake were seen. In another study, after OVX mice were fed an HFD, it was found that compared to sham-operated mice given the same HFD, their BWs increased by about 27.2% [
30], indicating that changes in appetite and food intake were not the necessary factors for body weight gain, as increased food utilization and changes in physical activities may also participate in energy metabolism in OVX animals, which in turn affects body weight changes [
31].
Compared to non-menopausal women, postmenopausal women are considered to be more prone to a high body fat percentage and abdominal fat accumulation [
8]. In results of past longitudinal studies, postmenopausal women had a 2.88-fold higher risk of developing abdominal obesity than non-menopausal women [
32]. Different sex hormones in the body affect the location of fat accumulation in the body. Estrogen in the ovaries is believed to increase the accumulation of peripheral fat in the buttocks and legs, while male hormones are likely to increase accumulation of fat in the abdomen. The decrease of estrogen in the body after menopause causes a relatively high concentration of male hormones in a woman’s body, causing changes in the location of body fat accumulation, and ultimately leading to abdominal fat accumulation [
33]. In results of this study, it was found that OVX rats had significant body fat accumulation, which is consistent with excessive fat accumulation in the abdomen under the OVX animal model in past experiments [
34]. Past studies found that even under different dietary conditions, OVX animals had increased total fat and abdominal fat [
35], reflecting the status of body fat accumulation in postmenopausal women, and the reduction of estrogen in the body after an OVX significantly increased the BW and occurrence of body fat accumulation [
36].
It is generally believed that under similar food intake level, HFDs have a higher calorie density per unit of food, which easily causes excessive caloric intake, which then accumulates in the body in the form of TGs, resulting in body weight gain and obesity. However, in this experiment, there were no significant differences in caloric intake among OVX rats given different FA ratios, and there was no higher caloric intake due to the high calorie density of the HFD. After 8 weeks of the intervention with high-dose (20%
w/
w) experimental oil (H group) given to OVX rats, BWs did not appear to differ with OVX rats fed a low-dose (5%
w/
w) of the experimental oil (L group), indicating that even if diets with different fat-to-total calorie ratios were given, on the basis of isocaloric intake, the 60% MUFA experimental oil with a P/S ratio of 5 did not produce a significant increase in BW. When observing the relative fat weight between groups of OVX rats given different diets, it was found that the relative gonad fat weight of M increased body fat than L and H groups. This phenomenon is consistent with previous studies. In a previous study by Díaz-Rúa et al., 8-week-old Wistar male rats were given a control group diet with fat making up 10% of total calories and an HFD with fat making up 60% of total calories. The daily food intake was controlled so that the two groups reached the same calorie intake. After 4 months of the experiment, the isocaloric HFD group did not a significant increase in body weight, while a higher body fat ratio and a state of obesity, along with increases in fasting plasma glucose and insulin concentrations were seen [
37]. According to previous studies, Yang et al. [
17] fed DIO hamsters 5%, 15%, and 20% (
w/
w) experimental oils for 8 weeks, and found that regardless of the fat percentages in the diet, the experimental oil mixture with 60% MUFAs and P/S = 5 had the effect of preventing body fat accumulation and balancing blood lipids in obese hamsters. Though HFD tends to increase greater BW, due to the results of previous studies, we may believe that regardless of the high percentage of fat content, experimental oil with designed percentage of MUFA may improves the body fat gaining process, which implied that experimental oil may interfere with the fatty acid metabolism pathways.
Under the same food and energy intake conditions, the C group had significantly greater levels of serum TC, LDL-C, fasting blood glucose, insulin, HOMA-IR, and leptin than the S group. There were no significant differences in lipid concentrations or enzyme activities in the liver between the S and the C groups, while lipid oxidation-related mRNA expressions of CPT-1 and PPARα significantly decreased, and AMPK significantly increased. PPAR-α is related to mitochondrial β-oxidation of fatty acids and CPT-1 is a key enzyme that enables fatty acids to go through the inner mitochondrial membrane and reach the mitochondrial matrix to be metabolized [
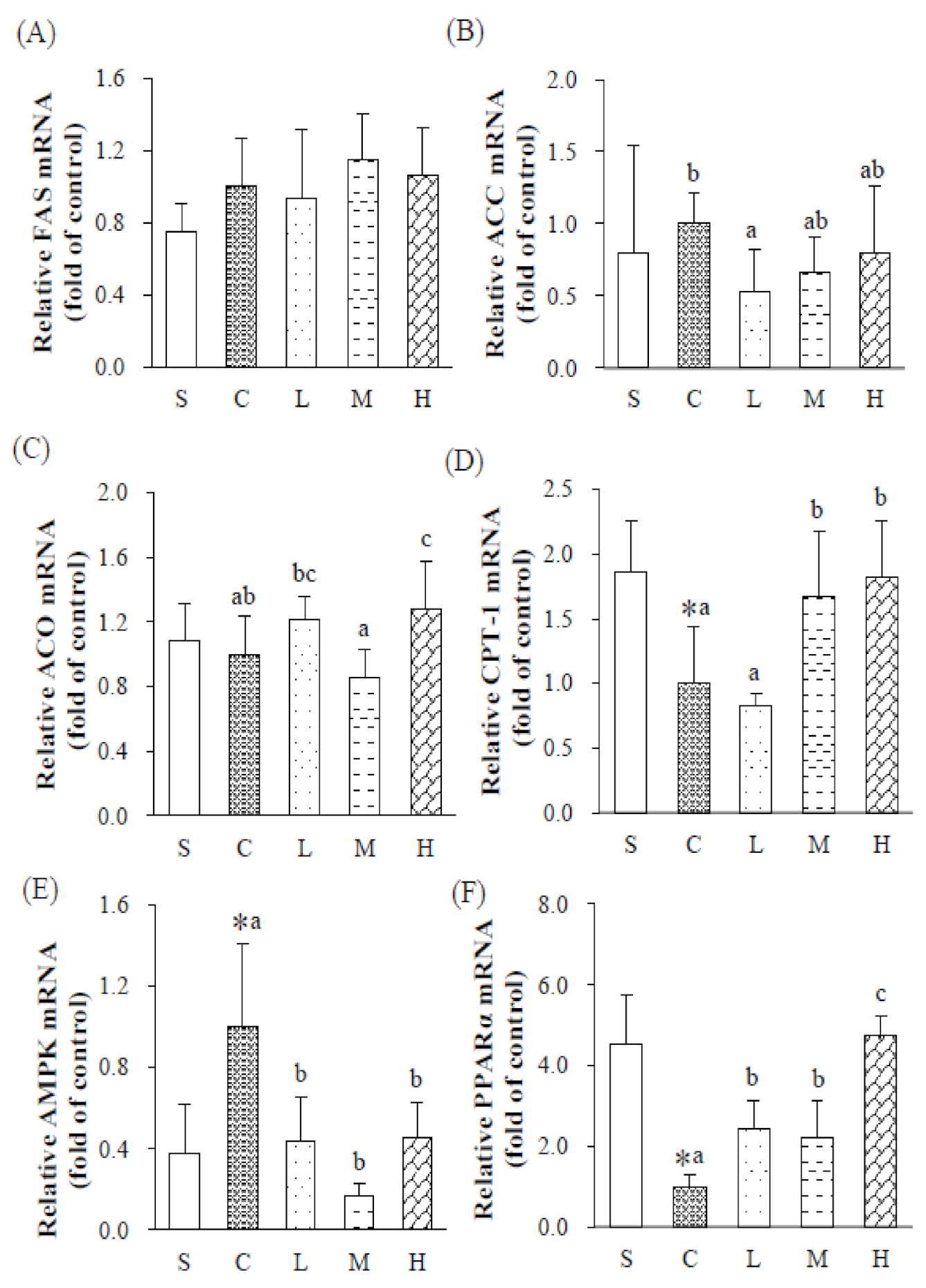
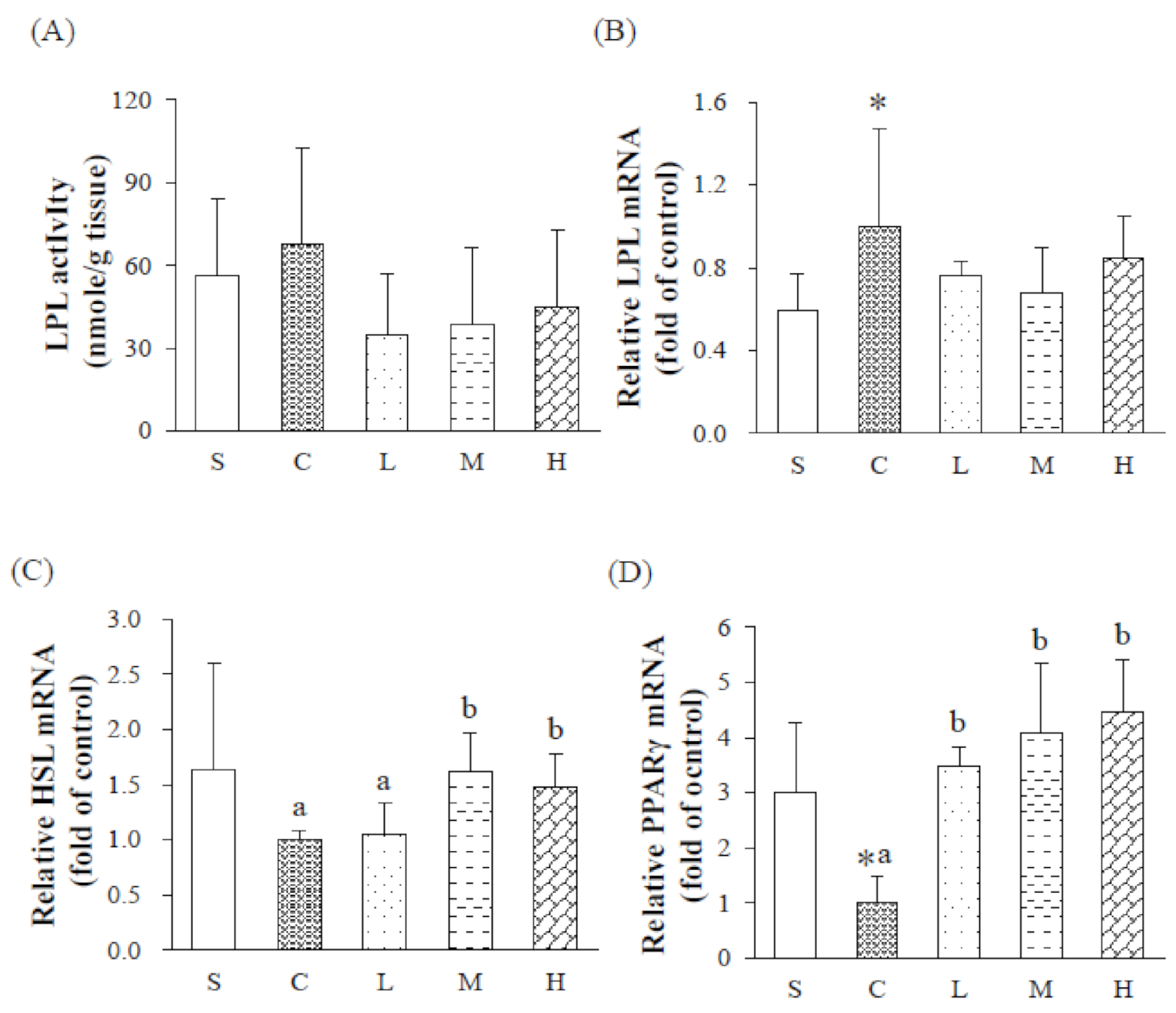
38]. LPL mRNA expression of WATs increased, and mRNA expressions of PPARγ significantly decreased. In summary, an OVX significantly reduced estrogen level and increased insulin concentrations in rats. High concentrations of insulin tend to inhibit the phosphorylation of AMPK, which indirectly inhibits downstream CPT-1 and reduces lipid oxidation. At the same time, it also increases the expression of LPL mRNA and increases adipogenesis in WATs, and ultimately alters lipid metabolism pathways, causing abnormal lipid metabolism in rats.
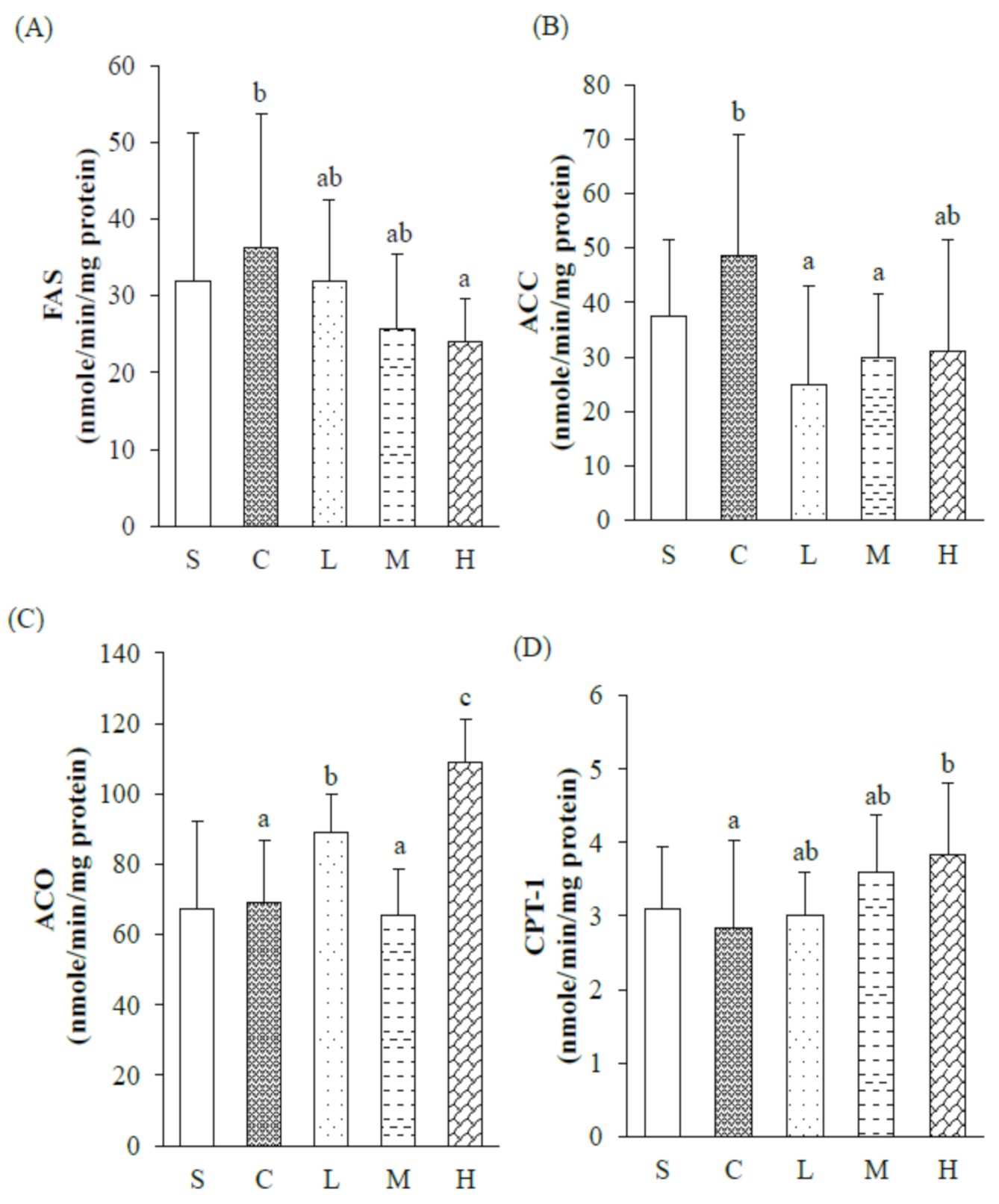
Under the same percentage of fat sources (5%
w/
w), the L group was given experimental oil with 60% MUFAs and P/S = 5. It exhibited significantly reduced serum insulin and HOMA-IR, ACC activity, and mRNA expression in the liver, and increased ACO activity and PPARα mRNA expression in the liver and PPARγ mRNA expression in WATs, compared to the C group. In a previous study, after 4 weeks of diet-induced obesity (DIO), low-fat diets with the same percentage of fat calories (9.5% kcal) were given, and experimental oil consisting of 60% MUFAs and P/S = 5 was found to be more effective in reducing the body fat ratio and plasma TC and leptin concentrations, increasing ACO and CPT-1 activities and mRNA expressions and lipid oxidation in the liver, and reducing body fat accumulation in DIO hamsters compared to those fed soybean oil [
18]. The experimental oil with 60% MUFAs and P/S = 5 was confirmed to have the ability to change lipid metabolism pathways in OVX rats by affecting blood glucose-related values, reducing insulin concentrations and lipid synthesis, and increasing lipid oxidation. Without affecting the calorie intake, increasing the lipid-to-calorie ratio in the diet did not increase insulin level or HOMA-IR in the H group of OVX rats fed the HFD. We speculated that the experimental oil mixture of 60% MUFAs and P/S = 5, even under circumstances of an HFD, did not increase the release of FFAs due to excessive intake of fat, nor accumulate in the liver in the form of TGs, resulting in hyperinsulinemia or the occurrence of IR, which is consistent with past studies [
16].
In a past study, Yang et al. [
17] fed DIO hamsters 5%, 15%, and 20% (
w/
w) experimental oils for 8 weeks, and found that regardless of the fat percentages in the diet, the experimental oil mixture with 60% MUFAs and P/S = 5 had the effect of preventing body fat accumulation and balancing blood lipids in obese hamsters. In the results of this experiment, it was also found that BW of rats in the H group that were given an HFD with the experimental oil did not significantly differ from those in the C group, and TG level in the liver and serum TG and FFA concentrations were significantly lower. It was speculated that administration of this HFD, consisting of experimental oil with 60% MUFAs and P/S = 5 did not cause an increase in hepatic insulin level of H group rats. Thus, there was no inhibitory effect of high insulin on AMPK phosphorylation, thereby reducing the inhibitory effect on ACC phosphorylation, and preserving ACC in an inactive state, which would reduce the carboxylation of acetyl CoA and also the production of downstream malonyl-CoA. As a result, the inhibitory effects of malonyl-CoA on CPT-1 were reduced, and the expression of ACO mRNA in the liver increased, consequently reducing the synthesis of hepatic TGs and attenuating hepatic FFA release caused by excessive TG accumulation in the liver [
20]. Under an HFD consisting of experimental oil with 60% MUFAs and P/S = 5, it was found that expressions of hepatic PPARα mRNA and PPARγ along with HSL mRNA in WATs significantly increased in the H group, suggesting a trend of the hepatic lipid oxidation pathway, and an increase in lipolysis in adipose tissues, thereby reducing the accumulation of TGs in adipose tissues of OVX rats fed an HFD.








{kind=link}
{kind=link}
{kind=link}