
Effect of Microalgae and Macroalgae Extracts on Non-Alcoholic Fatty Liver Disease

 ,
,  and
and 
Abstract
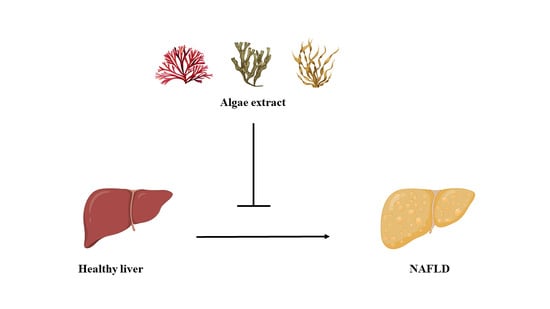
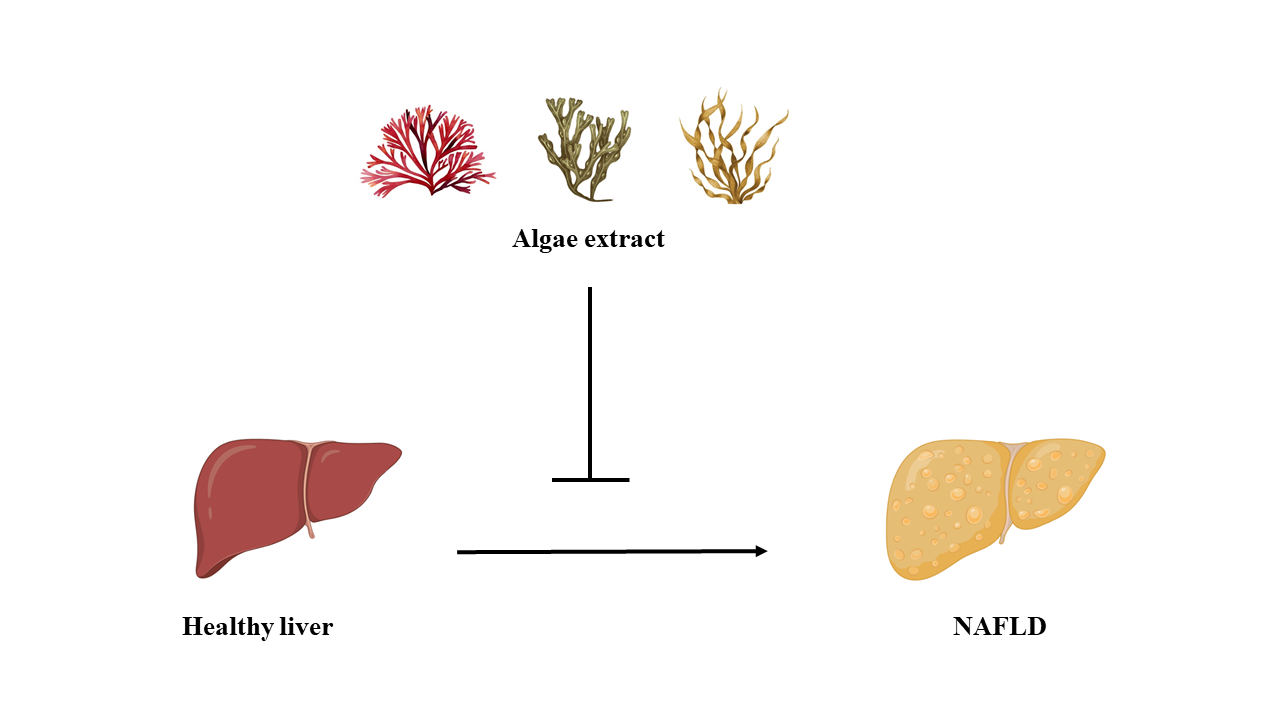
1. Introduction
2. Animal Studies
2.1. Microalgae
2.2. Macroalgae
2.2.1. Green Algae
2.2.2. Red Algae
2.2.3. Brown Algae
3. Human Studies
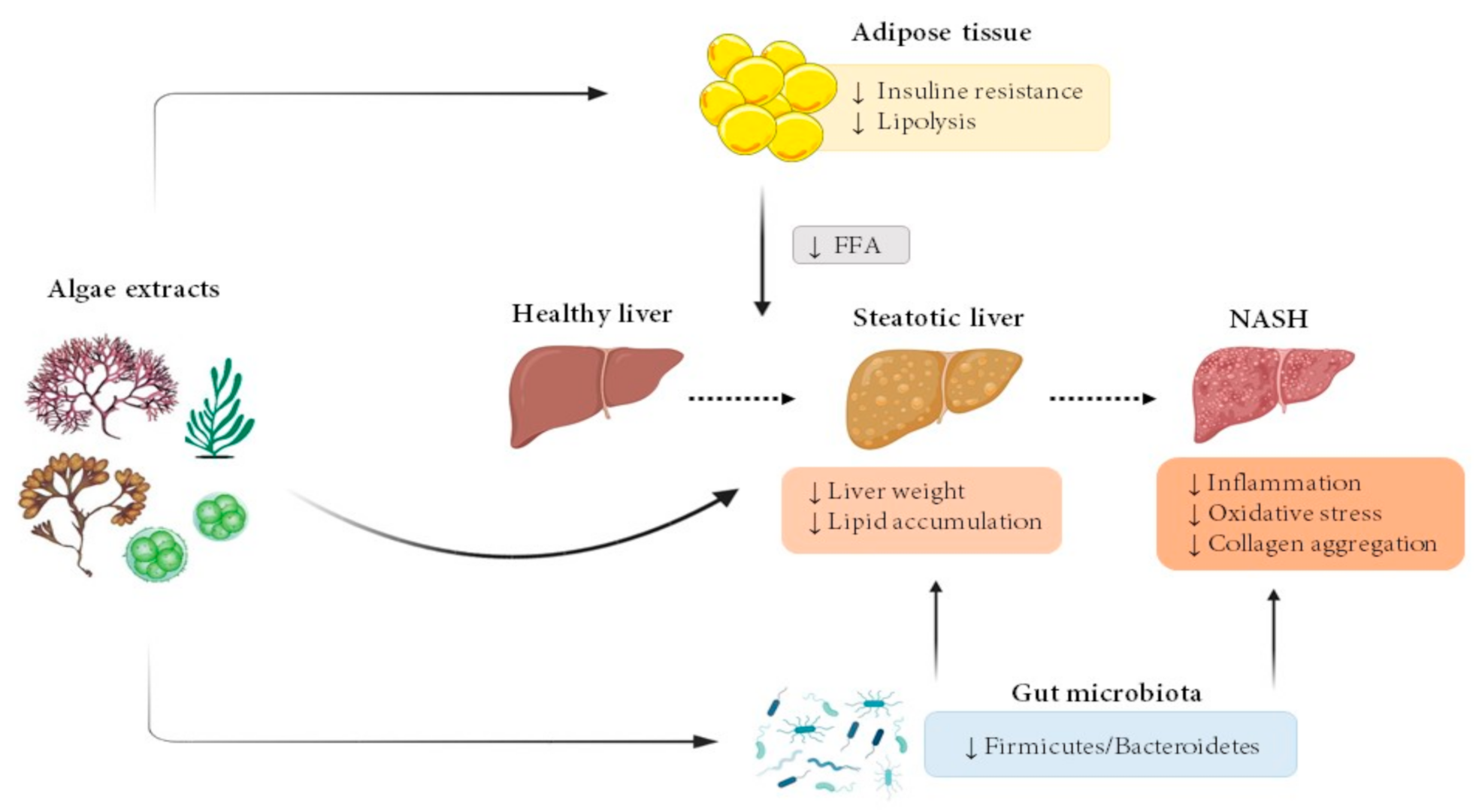
4. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Bessone, F.; Razori, M.V.; Roma, M.G. Molecular pathways of nonalcoholic fatty liver disease development and progression. Cell. Mol. Life Sci. 2019, 76, 99–128. [Google Scholar] [CrossRef] [PubMed]
- Younossi, Z.M. Non-alcoholic fatty liver disease—A global public health perspective. J. Hepatol. 2019, 70, 531–544. [Google Scholar] [CrossRef] [PubMed]
- Than, N.N.; Newsome, P.N. A concise review of non-alcoholic fatty liver disease. Atherosclerosis 2015, 239, 192–202. [Google Scholar] [CrossRef] [PubMed]
- Makri, E.; Goulas, A.; Polyzos, S.A. Epidemiology, Pathogenesis, Diagnosis and Emerging Treatment of Nonalcoholic Fatty Liver Disease. Arch. Med. Res. 2021, 52, 25–37. [Google Scholar] [CrossRef]
- Eslam, M.; Sanyal, A.J.; George, J.; Panel, I.C. MAFLD: A Consensus-Driven Proposed Nomenclature for Metabolic Associated Fatty Liver Disease. Gastroenterology 2020, 158, 1999–2014.e1. [Google Scholar] [CrossRef]
- Younossi, Z.M.; Rinella, M.E.; Sanyal, A.J.; Harrison, S.A.; Brunt, E.M.; Goodman, Z.; Cohen, D.E.; Loomba, R. From NAFLD to MAFLD: Implications of a Premature Change in Terminology. Hepatology 2021, 73, 1194–1198. [Google Scholar] [CrossRef]
- Biris-Dorhoi, E.S.; Michiu, D.; Pop, C.R.; Rotar, A.M.; Tofana, M.; Pop, O.L.; Socaci, S.A.; Farcas, A.C. Macroalgae-A Sustainable Source of Chemical Compounds with Biological Activities. Nutrients 2020, 12, 3085. [Google Scholar] [CrossRef]
- Cherry, P.; O’Hara, C.; Magee, P.J.; McSorley, E.M.; Allsopp, P.J. Risks and benefits of consuming edible seaweeds. Nutr. Rev. 2019, 77, 307–329. [Google Scholar] [CrossRef]
- Maehre, H.K.; Malde, M.K.; Eilertsen, K.E.; Elvevoll, E.O. Characterization of protein, lipid and mineral contents in common Norwegian seaweeds and evaluation of their potential as food and feed. J. Sci. Food Agric. 2014, 94, 3281–3290. [Google Scholar] [CrossRef]
- Rajapakse, N.; Kim, S.K. Nutritional and digestive health benefits of seaweed. Adv. Food Nutr. Res. 2011, 64, 17–28. [Google Scholar] [CrossRef]
- Khan, M.I.; Shin, J.H.; Kim, J.D. The promising future of microalgae: Current status, challenges, and optimization of a sustainable and renewable industry for biofuels, feed, and other products. Microb. Cell Fact. 2018, 17, 36. [Google Scholar] [CrossRef]
- Roohinejad, S.; Koubaa, M.; Barba, F.J.; Saljoughian, S.; Amid, M.; Greiner, R. Application of seaweeds to develop new food products with enhanced shelf-life, quality and health-related beneficial properties. Food Res. Int. 2017, 99, 1066–1083. [Google Scholar] [CrossRef]
- Cofrades, S.; Benedí, J.; Garcimartin, A.; Sánchez-Muniz, F.J.; Jimenez-Colmenero, F. A comprehensive approach to formulation of seaweed-enriched meat products: From technological development to assessment of healthy properties. Food Res. Int. 2017, 99, 1084–1094. [Google Scholar] [CrossRef]
- Kumar, S.A.; Magnusson, M.; Ward, L.C.; Paul, N.A.; Brown, L. A green algae mixture of Scenedesmus and Schroederiella attenuates obesity-linked metabolic syndrome in rats. Nutrients 2015, 7, 2771–2787. [Google Scholar] [CrossRef]
- Nakashima, A.; Sugimoto, R.; Suzuki, K.; Shirakata, Y.; Hashiguchi, T.; Yoshida, C.; Nakano, Y. Anti-fibrotic activity of Euglena gracilis and paramylon in a mouse model of non-alcoholic steatohepatitis. Food Sci. Nutr. 2018, 7, 139–147. [Google Scholar] [CrossRef]
- Pham, T.X.; Lee, Y.; Bae, M.; Hu, S.; Kang, H.; Kim, M.B.; Park, Y.K.; Lee, J.Y. Spirulina supplementation in a mouse model of diet-induced liver fibrosis reduced the pro-inflammatory response of splenocytes. Br. J. Nutr. 2019, 121, 748–755. [Google Scholar] [CrossRef]
- Mayer, C.; Richard, L.; Côme, M.; Ulmann, L.; Nazih, H.; Chénais, B.; Ouguerram, K.; Mimouni, V. The Marine Microalga, Tisochrysis lutea, protects agains metabolic disorders associated with metabolic syndrome and obesity. Nutrients 2021, 13, 430. [Google Scholar] [CrossRef]
- Sharma, B.R.; Kim, H.J.; Kim, M.S.; Park, C.M.; Rhyu, D.Y. Caulerpa okamurae extract inhibits adipogenesis in 3T3-L1 adipocytes and prevents high-fat diet-induced obesity in C57BL/6 mice. Nutr. Res. 2017, 47, 44–52. [Google Scholar] [CrossRef]
- du Preez, R.; Majzoub, M.E.; Thomas, T.; Panchal, S.K.; Brown, L. Caulerpa lentillifera (Sea Grapes) Improves Cardiovascular and Metabolic Health of Rats with Diet-Induced Metabolic Syndrome. Metabolites 2020, 10, 500. [Google Scholar] [CrossRef]
- Song, W.; Wang, Z.; Zhang, X.; Li, Y. Ethanol Extract from Ulva prolifera prevents High-fat diet-induced insulin resistance, oxidative stress, and inflammation response in mice. Biomed. Res. Int. 2018, 2018, 1374565. [Google Scholar] [CrossRef]
- Kang, M.C.; Kang, N.; Ko, S.C.; Kim, Y.B.; Jeon, Y.J. Anti-obesity effects of seaweeds of Jeju Island on the differentiation of 3T3-L1 preadipocytes and obese mice fed a high-fat diet. Food Chem. Toxicol. 2016, 90, 36–44. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.A.; Lee, H.G.; Li, X.; Hyun, J.M.; Kim, H.S.; Kim, T.H.; Kim, H.M.; Lee, J.J.; Kang, M.C.; Jeon, Y.J. Anti-obesity effects of red seaweed, Plocamium telfairiae, in C57BL/6 mice fed a high-fat diet. Food Funct. 2020, 11, 2299–2308. [Google Scholar] [CrossRef] [PubMed]
- Nakayama, H.; Shimada, Y.; Zang, L.; Terasawa, M.; Nishiura, K.; Matsuda, K.; Toombs, C.; Langdon, C.; Nishimura, N. Novel Anti-Obesity Properties of Palmaria mollis in Zebrafish and Mouse Models. Nutrients 2018, 10, 1401. [Google Scholar] [CrossRef] [PubMed]
- du Preez, R.; Paul, N.; Mouatt, P.; Majzoub, M.E.; Thomas, T.; Panchal, S.K.; Brown, L. Carrageenans from the red seaweed Sarconema filiforme attenuate symptoms of diet-induced metabolic syndrome in rats. Mar. Drugs 2020, 18, 97. [Google Scholar] [CrossRef]
- Lee, H.G.; Lu, Y.A.; Li, X.; Hyun, J.M.; Kim, H.S.; Lee, J.J.; Kim, T.H.; Kim, H.M.; Kang, M.C.; Jeon, A.Y. Anti-Obesity effects of Grateloupia elliptica, a red seaweed, in mice with high-fat diet-induced obesity via suppression of adipogenic factors in white adipose tissue and increased thermogenic factors in brown adipose tissue. Nutrients 2020, 12, 308. [Google Scholar] [CrossRef]
- Nabil-Adam, A.; Shreadah, M.A. Red algae natural products for prevention of lipopolysaccharides (LPS)-induced liver and kidney inflammation and injuries. Biosci. Rep. 2021, 41. [Google Scholar] [CrossRef]
- Murata, M.; Ishihara, K.; Saito, H. Hepatic fatty acid oxidation enzyme activities are stimulated in rats fed the brown seaweed, Undaria pinnatifida (wakame). J. Nutr. 1999, 129, 146–151. [Google Scholar] [CrossRef]
- Murata, M.; Sano, Y.; Ishihara, K.; Uchida, M. Dietary fish oil and Undaria pinnatifida (wakame) synergistically decrease rat serum and liver triacylglycerol. J. Nutr. 2002, 132, 742–747. [Google Scholar] [CrossRef]
- Li, L.; Wang, Y.; Yuan, J.; Liu, Z.; Ye, C.; Qin, S. Undaria pinnatifida improves obesity-related outcomes in association with gut microbiota and metabolomics modulation in high-fat diet-fed mice. Appl. Microbiol. Biotechnol. 2020, 104, 10217–10231. [Google Scholar] [CrossRef]
- Gabbia, D.; Saponaro, M.; Sarcognato, S.; Guido, M.; Ferri, N.; Carrara, M.; De Martin, S. Fucus vesiculosus and Aescophyllum nodosum ameliorate liver function by reducing diet-induced steatosis in rats. Mar. Drugs 2020, 18, 62. [Google Scholar] [CrossRef]
- Kang, M.C.; Lee, H.G.; Kim, H.S.; Song, K.M.; Chun, Y.G.; Lee, M.H.; Kim, B.K.; Jeon, Y.J. Anti-Obesity effects of Sargassum thunbergii via downregulation of adipogenesis gene and upregulation of thermogenic genes in high-fat diet-induced obese mice. Nutrients 2020, 12, 3325. [Google Scholar] [CrossRef]
- Murakami, S.; Hirazawa, C.; Ohya, T.; Yoshikawa, R.; Mizutani, T.; Ma, N.; Moriyama, M.; Ito, T.; Matsuzaki, C. The edible brown seaweed Sargassum horneri (Turneri) C. Agardh ameliorated high-fat diet-induced obesity, diabetes, and hepatic steatosis in mice. Nutrients 2021, 13, 551. [Google Scholar] [CrossRef]
- Ebrahimi-Mameghani, M.; Aliashrafi, S.; Javadzadeh, Y.; AsghariJafarabadi, M. The Effect of Chlorella vulgaris Supplementation on Liver En-zymes, Serum Glucose and Lipid Profile in Patients with Non-Alcoholic Fatty Liver Disease. Health Promot. Perspect. 2014, 4, 107–115. [Google Scholar] [CrossRef][Green Version]
- Li, H.; Gu, Y.; Wu, X.; Rayamajhi, S.; Bian, S.; Zhang, Q.; Meng, G.; Liu, L.; Wu, H.; Zhang, S.; et al. Association between consumption of edible seaweeds and newly diagnosed non-alcohol fatty liver disease: The TCLSIH Cohort Study. Liver Int. 2021, 41, 311–320. [Google Scholar] [CrossRef]
- Meroni, M.; Longo, M.; Rustichelli, A.; Dongiovanni, P. Nutrition and Genetics in NAFLD: The Perfect Binomium. Int. J. Mol. Sci. 2020, 21, 2986. [Google Scholar] [CrossRef]
- Meroni, M.; Longo, M.; Dongiovanni, P. The Role of Probiotics in Nonalcoholic Fatty Liver Disease: A New Insight into Therapeutic Strategies. Nutrients 2019, 11, 2642. [Google Scholar] [CrossRef]
- Cian, R.E.; Drago, S.R.; de Medina, F.S.; Martínez-Augustin, O. Proteins and Carbohydrates from Red Seaweeds: Evidence for Beneficial Effects on Gut Function and Microbiota. Mar. Drugs 2015, 13, 5358–5383. [Google Scholar] [CrossRef]
- de Jesus Raposo, M.F.; de Morais, A.M.; de Morais, R.M. Emergent Sources of Prebiotics: Seaweeds and Microalgae. Mar. Drugs 2016, 14, 27. [Google Scholar] [CrossRef]
- Zheng, L.X.; Chen, X.Q.; Cheong, K.L. Current trends in marine algae polysaccharides: The digestive tract, microbial catabolism, and prebiotic potential. Int. J. Biol. Macromol. 2020, 151, 344–354. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Author | Algae Species | Animal Model and Experimental Period Length | Experimental Groups | Effects | Mechanisms |
|---|---|---|---|---|---|
| Kumar et al., 2015 [14] | Scenedesmus dimorphus + Schroederiella apiculate mixture (green microalgae) | Male Wistar rats 8 weeks | Corn starch diet (C) High-carbohydrate high-fat diet (H) Corn starch diet + 5% microalgae mixture (CSC) High-carbohydrate high-fat diet + 5% microalgae mixture (HSC) | ↓ Liver weight ↓ Enlargement of fat vacuoles in hepatocytes (HC vs. H) ↓ Inflammation ↓ ALT and AST activities (HC vs. H) Improved glucose tolerance and insulin sensitivity (HC vs. H) | ↓ Infiltration of inflammatory cells |
| Nakashima et al., 2018 [15] | Euglena gracilis (green microalgae) | Male STAM mice 27 days | High-fat diet High-fat diet + 3 g/kg BW/day E. gracilis High-fat diet + 3 g/kg BW/day Paramylon High-fat diet + 10 mg/kg BW/day Telmisartan | Liver weight: NS Liver TG: NS ↓ Collagen aggregation (Euglena vs. vehicle) NAS Score: NS Serum ALT: NS | ↓ Immunostaining of F4/80, α-SMA (trend) Inflammation-related genes: NS Fibrosis-related genes: NS |
| Pham et al., 2019 [16] | Spirulina platensis (blue-green microalgae) | Male C57BL/6J mice 20 weeks | Low-fat (LF) High-fat/high-sucrose/high-cholesterol (HF) HF + 2.5% S. platensis (HF/SP) | Liver weight: NS Liver TG and cholesterol: NS ↓ Plasma ALT level Improvement of glucose tolerance Hepatic collagen accumulation: NS | mRNA levels of Col1a1 in liver: NS ↓ Il-1β mRNA levels in splenocytes |
| Mayer et al., 2021 [17] | Tisochrysis lutea (brown-golden microalgae) | Male Wistar rats 8 weeks | Standard diet (CTRL) High-fat high-fructose diet (HF) High-fat high-fructose + 12% T. lutea (HF-Tiso) | ↓ Liver TG and cholesterol ↓ Plasma AST, AST/ALT ↓ Plasma glucose, insulin, leptin ↓ Plasma TNF-α ↓ HOMAR-IR | No information provided |
| Author | Algae Species | Animal Model and Experimental Period Length | Experimental Groups | Effects | Mechanisms |
|---|---|---|---|---|---|
| Sharma et al., 2017 [18] | Caulerpa lentillifera | Male C57BL/6J mice 10 weeks | Standard diet High-fat diet High-fat diet + 250 mg/kg BW/day of C. lentillifera | ↓ Liver weight ↓ Liver TG, TC and FFA ↓ Plasma FFA, glucose and insulin | No information provided |
| du Preez et al., 2020 [19] | Caulerpa lentillifera | Male Wistar rats 16 weeks | Corn starch (C) Standard diet (C) High-carbohydrate high-fat (H) Standard diet + 5% C. lentillifera (CCL) High-carbohydrate high-fat + 5% C. lentillifera (HCL) | ↓ Liver TG content (H vs. HCL) Inflammatory cell infiltration: NS Plasma ALT, AST: NS ↓Firmicutes/Bacteroidetes ratio (H vs. HCL) | No information provided |
| Song et al., 2018 [20] | Ulva prolifera | Male C57BL/6 mice 8 weeks | Standard diet High-fat diet High-fat diet + 2% ethanol extract of U. prolifera in drinking water High-fat diet + 5% ethanol extract of U. prolifera in drinking water | ↓ Liver weight ↓ Liver TG content ↓ Serum insulin ↓ Oxidative stress | ↓ Dgat1 and Dgat2 liver mRNA levels ↑ Cpt-1a, Acadm and Acox1 liver mRNA levels ↓ Serum IL-1β, IL-6 and TNF-α ↓ Il-1β, Il-6 and Tnf-α liver mRNA levels ↓ Liver ROS ↑ GSH content and GSHPx activity |
| Author | Algae Species | Animal Model and Experimental Period Length | Experimental Groups | Effects | Mechanisms |
|---|---|---|---|---|---|
| Kang et al., 2016 [21] | Plocamium telfairiae | Male C57BL/6 mice 14 weeks | Standard diet High-fat diet High-fat diet +100 mg/kg BW/day of P. telfairiae | ↓ Liver steatosis ↓ Serum glucose | No information provided |
| Lu et al., 2020 [22] | Plocamium telfairiae | Male C57BL/6 mice 7 weeks | Standard diet High-fat diet High-fat diet +100 mg/kg BW/day of P. telfairiae High-fat diet +165 mg/kg BW/day of P. telfairiae High-fat diet + 300 mg/kg BW/day of P. telfairiae | ↓ Hepatic steatosis (all doses) | No information provided |
| Nakayama et al., 2018 [23] | Palmaria mollis | Male NSY/HOS mice 4 weeks | Standard diet High-fat diet High-fat diet + 2.5% of P. mollis | ↓ Liver TG content | ↑ Pparα, C/ebpα and Acox1 mRNA levels ↓ Pparɣ mRNA levels Acadm and Srebf1 mRNA levels: NS |
| du Preez et al., 2020 [24] | Sarconema filiforme | Male Wistar rats 16 weeks | Corn starch diet Corn starch diet + 5% S. filiforme High-carbohydrate high-fat diet High-carbohydrate high-fat diet + 5% S. filiforme (drinking water of rats fed the steatotic diet was supplemented with 25% fructose) | ↓ Liver steatosis and infiltration of inflammatory cells (high-carbohydrate high-fat diet 5% S. filiforme vs. high-carbohydrate high-fat diet) Serum glucose: NS ↓ Serum ALT and AST (high-carbohydrate high-fat diet 5% S. filiforme vs. high-carbohydrate high-fat diet) Firmicutes/Bacteroidetes: NS | No information provided |
| Lee et al., 2020 [25] | Grateloupia elliptica | Male C57BL/6 mice 7 weeks | Standard diet High-fat high-sucrose diet High-fat high-sucrose diet +125 mg/kg BW/day of G. elliptica High-fat high-sucrose diet +250 mg/kg BW/day of G. elliptica | ↓ Hepatic steatosis | No information provided |
| Nabil-Adam et al., 2021 [26] | Gromphadorhina oblongata | BALB/C mice 1 week | Negative control: saline solution Induction control: 5 mg/kg BW/day of LPS Protected group: 200 mg/kg BW/day of G. oblongata 2 h before LPS Positive control: 200 mg/kg BW/day of G. oblongata without LPS | ↓ Liver injury (inflammation and oxidative stress) ↑ Liver apoptosis ↓ Serum ALT, AST | No information provided |
| Author | Algae Species | Animal Model and Experimental Period Length | Experimental Groups | Effects | Mechanisms |
|---|---|---|---|---|---|
| Murata et al., 1999 [27] | Undaria pinnatifida | Male Sprague–Dawley rats 3 weeks | Standard diet Standard diet + 0.5% U. pinnatifida Standard diet + 1% U. pinnatifida Standard diet + 2% U.pinnatifida Standard diet + 5% U. pinnatifida Standard diet + 10% U.pinnatifida | ↓ Liver TG content in 1, 2, 5 and 10% groups ↓ Liver TC content in 10% group | ↓G6PD activity in 5 and 10% groups ↑ CPT activity in 10% group ↑ ACADs activity in 5 and 10% groups ↑ ACO in 10% group ↑ DECR1 in 5 and 10% groups |
| Murata et al., 2002 [28] | Undaria pinnatifida | Male Sprague–Dawley rats 4 weeks | Standard diet Standard diet + 19.1% U. pinnatifida | ↓ Liver weight ↓ Hepatic TG, TC and phospholipids levels | ↓ G6PD activity ↑ ACO and 3-hydroxiacil-CoA dehydrogenase activities CPT activity: NS |
| Li et al., 2020 [29] | Undaria pinnatifida | Male C57BL/6 mice 10 weeks | Standard diet Standard diet + 10% U. pinnatifida High-fat diet High-fat diet + 10% U. pinnatifida | ↓ Liver steatosis ↓ Glucose levels | No information provided |
| Gabbia et al., 2020 [30] | Fucus vesiculosus + Ascophyllum nodosum | Male Wistar rats 5 weeks | High-fat diet (HFD) High-fat diet + 7.5 mg/kg BW/day of F. vesiculosus and A. nodosum | ↓ Liver weight ↓ Microvesicular steatosis ↓ Plasma ALT and AST levels Lower and delayed glucose peak | No information provided |
| Kang et al., 2020 [31] | Sargassum thunbergii | Male C57BL/6 mice 7 weeks | Standard diet High-fat diet High-fat diet + 100 mg/kg BW/day of S. thunbergii High-fat diet + 300 mg/kg BW/day of S. thunbergii | ↓ Lipid steatosis | No information provided |
| Murakami et al., 2021 [32] | Sargassum horneri | Male C57BL/6J mice 13 weeks | Standard diet High-fat diet (HF) High-fat diet + 2% S. horneri (HF + ShL) High-fat diet + 6% S. horneri (HF + ShH) | ↓ Liver weight ↓ Liver TG content ↓ Serum glucose, insulin, ALT, AST, ALP and LAP levels ↑ Serum adiponectin ↓ Serum TNF-α | ↓ Pancreatic lipase activity |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
González-Arceo, M.; Gómez-Zorita, S.; Aguirre, L.; Portillo, M.P. Effect of Microalgae and Macroalgae Extracts on Non-Alcoholic Fatty Liver Disease. Nutrients 2021, 13, 2017. https://doi.org/10.3390/nu13062017
González-Arceo M, Gómez-Zorita S, Aguirre L, Portillo MP. Effect of Microalgae and Macroalgae Extracts on Non-Alcoholic Fatty Liver Disease. Nutrients. 2021; 13(6):2017. https://doi.org/10.3390/nu13062017
Chicago/Turabian StyleGonzález-Arceo, Maitane, Saioa Gómez-Zorita, Leixuri Aguirre, and María P. Portillo. 2021. "Effect of Microalgae and Macroalgae Extracts on Non-Alcoholic Fatty Liver Disease" Nutrients 13, no. 6: 2017. https://doi.org/10.3390/nu13062017
APA StyleGonzález-Arceo, M., Gómez-Zorita, S., Aguirre, L., & Portillo, M. P. (2021). Effect of Microalgae and Macroalgae Extracts on Non-Alcoholic Fatty Liver Disease. Nutrients, 13(6), 2017. https://doi.org/10.3390/nu13062017