
Vitamin A Update: Forms, Sources, Kinetics, Detection, Function, Deficiency, Therapeutic Use and Toxicity

 ,
,  , , ,
, , ,  and
and 
Abstract
1. Introduction and Forms of Vitamin A
2. Sources of Vitamin A
3. Vitamin A Pharmacokinetics
3.1. Absorption, Distribution, Metabolism and Elimination
3.1.1. Absorption
3.1.2. Distribution and Cytoplasmic Fate of Vitamin A
3.1.3. Elimination
3.2. Other Factors Influencing Vitamin A Pharmacokinetics
3.3. Pharmacokinetics of Vitamin A during Pregnancy
4. Vitamin A Functions
4.1. Vision
4.2. Interaction with Nuclear Receptors
4.2.1. Vitamin A and Cancer
4.2.2. Vitamin A, Immunity and Inflammation
4.2.3. Other Functions Associated with Gene Transcription
4.3. Other Functions of Retinoids
5. Analytical Approaches for Measuring Vitamin A Levels
5.1. Detection Means
5.2. Chromatographic Considerations
5.3. Other Methods
5.4. Considerations on the Stability of Retinoic Acid
6. Vitamin A Deficiency
6.1. Symptoms
6.2. Causes and Epidemiology
7. Therapeutic Use of Vitamin A
8. Hypervitaminosis and Vitamin A Toxicity
9. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- D’Ambrosio, D.N.; Clugston, R.D.; Blaner, W.S. Vitamin A metabolism: An update. Nutrients 2011, 3, 63–103. [Google Scholar] [CrossRef] [PubMed]
- Kam, R.K.; Deng, Y.; Chen, Y.; Zhao, H. Retinoic acid synthesis and functions in early embryonic development. Cell. Biosci. 2012, 2, 11. [Google Scholar] [CrossRef] [PubMed]
- Chiricozzi, A.; Panduri, S.; Dini, V.; Tonini, A.; Gualtieri, B.; Romanelli, M. Optimizing acitretin use in patients with plaque psoriasis. Dermatol. Ther. 2017, 30. [Google Scholar] [CrossRef]
- Dragnev, K.H.; Petty, W.J.; Shah, S.J.; Lewis, L.D.; Black, C.C.; Memoli, V.; Nugent, W.C.; Hermann, T.; Negro-Vilar, A.; Rigas, J.R.; et al. A proof-of-principle clinical trial of bexarotene in patients with non-small cell lung cancer. Clin. Cancer Res. 2007, 13, 1794–1800. [Google Scholar] [CrossRef] [PubMed]
- Thielitz, A.; Gollnick, H. Topical retinoids in acne vulgaris: Update on efficacy and safety. Am. J. Clin. Dermatol. 2008, 9, 369–381. [Google Scholar] [CrossRef] [PubMed]
- Duvic, M.; Hymes, K.; Heald, P.; Breneman, D.; Martin, A.G.; Myskowski, P.; Crowley, C.; Yocum, R.C.; Bexarotene Worldwide Study, G. Bexarotene is effective and safe for treatment of refractory advanced-stage cutaneous T-cell lymphoma: Multinational phase II-III trial results. J. Clin. Oncol. 2001, 19, 2456–2471. [Google Scholar] [CrossRef]
- Qu, L.; Tang, X. Bexarotene: A promising anticancer agent. Cancer Chemother. Pharmacol. 2010, 65, 201–205. [Google Scholar] [CrossRef] [PubMed]
- FDA. Drug Trials Snapshot: AKLIEF. Available online: https://www.fda.gov/drugs/drug-approvals-and-databases/drug-trials-snapshots-aklief (accessed on 27 January 2021).
- Kelly, M.E.; Ramkumar, S.; Sun, W.; Colon Ortiz, C.; Kiser, P.D.; Golczak, M.; von Lintig, J. The Biochemical Basis of Vitamin A Production from the Asymmetric Carotenoid β-Cryptoxanthin. ACS Chem. Biol. 2018, 13, 2121–2129. [Google Scholar] [CrossRef]
- Barbosa-Filho, J.; Alencar, A.; Nunes, X.P.; Tomaz, A.A.; Sena-Filho, J.G.; Athayde-Filho, P.; Silva, M.E.; Souza, M.D.F.V.; da-Cunha, E.V. Sources of alpha-, beta-, gamma-, delta- and epsilon-carotenes: A twentieth century review. Rev. Bras. Farmacogn. Braz. J. Pharmacogn. 2008, 18, 135–154. [Google Scholar] [CrossRef]
- Beltrán-de-Miguel, B.; Estévez-Santiago, R.; Olmedilla-Alonso, B. Assessment of dietary vitamin A intake (retinol, α-carotene, β-carotene, β-cryptoxanthin) and its sources in the National Survey of Dietary Intake in Spain (2009–2010). Int. J. Food Sci. Nutr. 2015, 66, 706–712. [Google Scholar] [CrossRef]
- Maiani, G.; Castón, M.J.; Catasta, G.; Toti, E.; Cambrodón, I.G.; Bysted, A.; Granado-Lorencio, F.; Olmedilla-Alonso, B.; Knuthsen, P.; Valoti, M.; et al. Carotenoids: Actual knowledge on food sources, intakes, stability and bioavailability and their protective role in humans. Mol. Nutr. Food Res. 2009, 53 (Suppl. 12), S194–S218. [Google Scholar] [CrossRef]
- Ma, G.; Zhang, L.; Iida, K.; Madono, Y.; Yungyuen, W.; Yahata, M.; Yamawaki, K.; Kato, M. Identification and quantitative analysis of β-cryptoxanthin and β-citraurin esters in Satsuma mandarin fruit during the ripening process. Food Chem. 2017, 234, 356–364. [Google Scholar] [CrossRef] [PubMed]
- Jiao, Y.; Reuss, L.; Wang, Y. β-Cryptoxanthin: Chemistry, Occurrence, and Potential Health Benefits. Curr. Pharmacol. Rep. 2019, 5, 20–34. [Google Scholar] [CrossRef]
- Breithaupt, D.E.; Bamedi, A. Carotenoid esters in vegetables and fruits: A screening with emphasis on beta-cryptoxanthin esters. J. Agric. Food Chem. 2001, 49, 2064–2070. [Google Scholar] [CrossRef] [PubMed]
- Schlatterer, J.; Breithaupt, D.E. Cryptoxanthin Structural Isomers in Oranges, Orange Juice, and Other Fruits. J. Agric. Food Chem. 2005, 53, 6355–6361. [Google Scholar] [CrossRef] [PubMed]
- Seroczynska, A.; Korzeniewska, A.; Sztangret-Wisniewska, J.; Niemirowicz-Szczytt, K.; Marek, G. Relationship between carotenoids content and flower or fruit flesh colour of winter squash (Cucurbita maxima Duch.). Folia Hortic. 2006, 18, 51–61. [Google Scholar]
- Teow, C.C.; Truong, V.-D.; McFeeters, R.F.; Thompson, R.L.; Pecota, K.V.; Yencho, G.C. Antioxidant activities, phenolic and β-carotene contents of sweet potato genotypes with varying flesh colours. Food Chem. 2007, 103, 829–838. [Google Scholar] [CrossRef]
- Dragovic-Uzelac, V.; Levaj, B.; Mrkic, V.; Bursac, D.; Boras, M. The content of polyphenols and carotenoids in three apricot cultivars depending on stage of maturity and geographical region. Food Chem. 2007, 102, 966–975. [Google Scholar] [CrossRef]
- Andersson, S.C.; Olsson, M.E.; Johansson, E.; Rumpunen, K. Carotenoids in Sea Buckthorn (Hippophae rhamnoides L.) Berries during Ripening and Use of Pheophytin a as a Maturity Marker. J. Agric. Food Chem. 2009, 57, 250–258. [Google Scholar] [CrossRef]
- Zaccari, F.; Galietta, G. α-Carotene and β-Carotene Content in Raw and Cooked Pulp of Three Mature Stage Winter Squash “Type Butternut”. Foods 2015, 4, 477–486. [Google Scholar] [CrossRef]
- Laurie, S.M.; Faber, M.; van Jaarsveld, P.J.; Laurie, R.N.; du Plooy, C.P.; Modisane, P.C. β-Carotene yield and productivity of orange-fleshed sweet potato (Ipomoea batatas L. Lam.) as influenced by irrigation and fertilizer application treatments. Sci. Hortic. 2012, 142, 180–184. [Google Scholar] [CrossRef]
- Ndawula, J.; Kabasa, J.D.; Byaruhanga, Y.B. Alterations in fruit and vegetable beta-carotene and vitamin C content caused by open-sun drying, visqueen-covered and polyethylene-covered solar-dryers. Afr. Health Sci. 2004, 4, 125–130. [Google Scholar] [PubMed]
- Karabulut, I.; Topcu, A.; Duran, A.; Turan, S.; Ozturk, B. Effect of hot air drying and sun drying on color values and β-carotene content of apricot (Prunus armenica L.). LWT-Food Sci. Technol. 2007, 40, 753–758. [Google Scholar] [CrossRef]
- Negi, P.S.; Roy, S.K. Effect of Blanching and Drying Methods on β -Carotene, Ascorbic acid and Chlorophyll Retention of Leafy Vegetables. LWT-Food Sci. Technol. 2000, 33, 295–298. [Google Scholar] [CrossRef]
- Speek, A.J.; Speek-Saichua, S.; Schreurs, W.H.P. Total carotenoid and β-carotene contents of Thai vegetables and the effect of processing. Food Chem. 1988, 27, 245–257. [Google Scholar] [CrossRef]
- Igual, M.; García-Martínez, E.; Camacho, M.M.; Martínez-Navarrete, N. Jam processing and storage effects on β-carotene and flavonoids content in grapefruit. J. Funct. Foods 2013, 5, 736–744. [Google Scholar] [CrossRef]
- van het Hof, K.H.; West, C.E.; Weststrate, J.A.; Hautvast, J.G.A.J. Dietary Factors That Affect the Bioavailability of Carotenoids. J. Nutr. 2000, 130, 503–506. [Google Scholar] [CrossRef]
- Raja, R.; Hemaiswarya, S.; Rengasamy, R. Exploitation of Dunaliella for β-carotene production. Appl. Microbiol. Biotechnol. 2007, 74, 517–523. [Google Scholar] [CrossRef]
- Bogacz-Radomska, L.; Harasym, J. β-Carotene—Properties and production methods. Food Qual. Saf. 2018, 2, 69–74. [Google Scholar] [CrossRef]
- Seshadri, C.V.; Umesh, B.V.; Manoharan, R. Beta-carotene studies in Spirulina. Bioresour. Technol. 1991, 38, 111–113. [Google Scholar] [CrossRef]
- Christaki, E.; Bonos, E.; Giannenas, I.; Florou-Paneri, P. Functional properties of carotenoids originating from algae. J. Sci. Food Agric. 2013, 93, 5–11. [Google Scholar] [CrossRef] [PubMed]
- Ram, S.; Mitra, M.; Shah, F.; Tirkey, S.R.; Mishra, S. Bacteria as an alternate biofactory for carotenoid production: A review of its applications, opportunities and challenges. J. Funct. Foods 2020, 67, 103867. [Google Scholar] [CrossRef]
- Mitra, R.; Chaudhuri, S.; Dutta, D. Modelling the growth kinetics of Kocuria marina DAGII as a function of single and binary substrate during batch production of β-Cryptoxanthin. Bioprocess. Biosyst. Eng. 2017, 40, 99–113. [Google Scholar] [CrossRef]
- Bhaskarachary, K.; Rao, D.S.S.; Deosthale, Y.G.; Reddy, V. Carotene content of some common and less familiar foods of plant origin. Food Chem. 1995, 54, 189–193. [Google Scholar] [CrossRef]
- Booth, S.L.; Johns, T.; Kuhnlein, H.V. Natural Food Sources of Vitamin A and Provitamin A. Food Nutr. Bull. 1992, 14, 1–15. [Google Scholar] [CrossRef]
- Wall, M.M.; Waddell, C.A.; Bosland, P.W. Variation in β-Carotene and Total Carotenoid Content in Fruits of Capsicum. Hort. Sci. 2001, 36, 746. [Google Scholar] [CrossRef]
- Kandlakunta, B.; Rajendran, A.; Thingnganing, L. Carotene content of some common (cereals, pulses, vegetables, spices and condiments) and unconventional sources of plant origin. Food Chem. 2008, 106, 85–89. [Google Scholar] [CrossRef]
- Laur, L.M.; Tian, L. Provitamin A and vitamin C contents in selected California-grown cantaloupe and honeydew melons and imported melons. J. Food Compos. Anal. 2011, 24, 194–201. [Google Scholar] [CrossRef]
- Kopsell, D.A.; Kopsell, D.E.; Lefsrud, M.G.; Curran-Celentano, J.; Dukach, L.E. Variation in Lutein, β-carotene, and Chlorophyll Concentrations among Brassica oleracea Cultigens and Seasons. Hort. Sci. 2004, 39, 361. [Google Scholar] [CrossRef]
- Žnidarčič, D.; Ban, D.; Šircelj, H. Carotenoid and chlorophyll composition of commonly consumed leafy vegetables in Mediterranean countries. Food Chem. 2011, 129, 1164–1168. [Google Scholar] [CrossRef]
- Veda, S.; Platel, K.; Srinivasan, K. Varietal Differences in the Bioaccessibility of β-Carotene from Mango (Mangifera indica) and Papaya (Carica papaya) Fruits. J. Agric. Food Chem. 2007, 55, 7931–7935. [Google Scholar] [CrossRef] [PubMed]
- Souza, L.M.d.; Ferreira, K.S.; Chaves, J.B.c.P.; Teixeira, S.l.L. L-ascorbic acid, ²-carotene and lycopene content in papaya fruits (Carica papaya) with or without physiological skin freckles. Sci. Agric. 2008, 65, 246–250. [Google Scholar] [CrossRef]
- Khush, G.S.; Lee, S.; Cho, J.-I.; Jeon, J.-S. Biofortification of crops for reducing malnutrition. Plant. Biotechnol. Rep. 2012, 6, 195–202. [Google Scholar] [CrossRef]
- Hornero-Méndez, D.; Mínguez-Mosquera, M.I. Carotenoid pigments in Rosa mosqueta hips, an alternative carotenoid source for foods. J. Agric. Food Chem. 2000, 48, 825–828. [Google Scholar] [CrossRef] [PubMed]
- Piccaglia, R.; Marotti, M.; Chiavari, G.; Gandini, N. Effects of Harvesting Date and Climate on the Flavonoid and Carotenoid Contents of Marigold (Calendula officinalis L.). Flavour Fragr. J. 1997, 12, 85–90. [Google Scholar] [CrossRef]
- Tudor, C.; Bohn, T.; Iddir, M.; Dulf, F.V.; Focşan, M.; Rugină, D.O.; Pintea, A. Sea Buckthorn Oil as a Valuable Source of Bioaccessible Xanthophylls. Nutrients 2019, 12, 76. [Google Scholar] [CrossRef] [PubMed]
- Mba, O.I.; Dumont, M.-J.; Ngadi, M. Palm oil: Processing, characterization and utilization in the food industry—A review. Food Biosci. 2015, 10, 26–41. [Google Scholar] [CrossRef]
- Caprioli, G.; Kamgang Nzekoue, F.; Fiorini, D.; Scocco, P.; Trabalza-Marinucci, M.; Acuti, G.; Tardella, F.M.; Sagratini, G.; Catorci, A. The effects of feeding supplementation on the nutritional quality of milk and cheese from sheep grazing on dry pasture. Int. J. Food Sci. Nutr. 2020, 71, 50–62. [Google Scholar] [CrossRef]
- Ahmad, S. Composition and physico-chemical characteristics of buffalo milk with particular emphasis on lipids, proteins, minerals, enzymes and vitamins. J. Anim. Plant. Sci. 2013, 23, 62–74. [Google Scholar]
- Kondyli, E.; Svarnas, C.; Samelis, J.; Katsiari, M.C. Chemical composition and microbiological quality of ewe and goat milk of native Greek breeds. Small Rumin. Res. 2012, 103, 194–199. [Google Scholar] [CrossRef]
- Barłowska, J.; Szwajkowska, M.; Litwińczuk, Z.; Król, J. Nutritional Value and Technological Suitability of Milk from Various Animal Species Used for Dairy Production. Compr. Rev. Food Sci. Food Saf. 2011, 10, 291–302. [Google Scholar] [CrossRef]
- Fedele, V.; Rubino, R.; Salvatore, C.; Manzi, P.; Marconi, S.; Pizzoferrato, L. Seasonal variation in retinol concentration of goat milk associated with grazing compared to indoor feeding. S. Afr. J. Anim. Sci. 2004, 34, 148–150. [Google Scholar]
- Ramalho, H.M.M.; Santos, J.; Casal, S.; Alves, M.R.; Oliveira, M. Fat-soluble vitamin (A, D, E, and β-carotene) contents from a Portuguese autochthonous cow breed—Minhota. J. Dairy Sci. 2012, 95, 5476–5484. [Google Scholar] [CrossRef] [PubMed]
- Ollilainen, V.; Heinonen, M.; Linkola, E.; Varo, P.; Koivistoinen, P. Carotenoids and Retinoids in Finnish Foods: Dairy Products and Eggs. J. Dairy Sci. 1989, 72, 2257–2265. [Google Scholar] [CrossRef]
- Majchrzak, D.; Fabian, E.; Elmadfa, I. Vitamin A content (retinol and retinyl esters) in livers of different animals. Food Chem. 2006, 98, 704–710. [Google Scholar] [CrossRef]
- Álvarez, R.; Meléndez-Martínez, A.J.; Vicario, I.M.; Alcalde, M.J. Carotenoid and Vitamin A Contents in Biological Fluids and Tissues of Animals as an Effect of the Diet: A Review. Food Rev. Int. 2015, 31, 319–340. [Google Scholar] [CrossRef]
- Ollilainen, V.; Heinonen, M.; Linkola, E.; Varo, P.; Koivistoinen, P. Carotenoids and retinoids in finnish foods: Meat and meat products. J. Food Compos. Anal. 1988, 1, 178–188. [Google Scholar] [CrossRef]
- Darwish, W.S.; Ikenaka, Y.; Morshdy, A.E.; Eldesoky, K.I.; Nakayama, S.; Mizukawa, H.; Ishizuka, M. β-carotene and retinol contents in the meat of herbivorous ungulates with a special reference to their public health importance. J. Vet. Med. Sci. 2016, 78, 351–354. [Google Scholar] [CrossRef]
- Schweigert, F.; Buchholz, I.; Schuhmacher, A. Effect of dietary β-carotene on the accumulation of β-carotene and vitamin A in plasma and tissues of gilts. Reprod. Nutr. Dev. Reprod. Nutr. Dev. 2001, 41, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Domínguez, R.; Barba, F.J.; Centeno, J.A.; Putnik, P.; Alpas, H.; Lorenzo, J.M. Simple and Rapid Method for the Simultaneous Determination of Cholesterol and Retinol in Meat Using Normal-Phase HPLC Technique. Food Anal. Methods 2018, 11, 319–326. [Google Scholar] [CrossRef]
- La Frano, M.R.; Burri, B.J. Analysis of retinol, 3-hydroxyretinol and 3,4-didehydroretinol in North American farm-raised freshwater fish liver, muscle and feed. Aquac. Nutr. 2014, 20, 722–730. [Google Scholar] [CrossRef]
- Dewailly, E.; Rouja, P.; Schultz, E.; Julien, P.; Tucker, T. Vitamin A intoxication from reef fish liver consumption in Bermuda. J. Food Prot. 2011, 74, 1581–1583. [Google Scholar] [CrossRef]
- Li, H.; Tyndale, S.T.; Heath, D.D.; Letcher, R.J. Determination of carotenoids and all-trans-retinol in fish eggs by liquid chromatography-electrospray ionization-tandem mass spectrometry. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2005, 816, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Sunarya; Hole, M.; Taylor, K.D.A. Methods of extraction composition and stability of vitamin A and other components in dogfish (Squalus acanthias) liver oil. Food Chem. 1996, 55, 215–220. [Google Scholar] [CrossRef]
- Irie, T.; Sugimoto, T.; Ueki, N.; Senoo, H.; Seki, T. Retinoid storage in the egg of reptiles and birds. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2010, 157, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Anderson, K.E. Comparison of fatty acid, cholesterol, vitamin A and E composition, and trans fats in eggs from brown and white egg strains that were molted or nonmolted. Poult. Sci. 2013, 92, 3259–3265. [Google Scholar] [CrossRef]
- Jiang, Y.H.; McGeachin, R.B.; Bailey, C.A. α-Tocopherol, β-Carotene, and Retinol Enrichment of Chicken Eggs. Poult. Sci. 1994, 73, 1137–1143. [Google Scholar] [CrossRef]
- Heying, E.K.; Tanumihardjo, J.P.; Vasic, V.; Cook, M.; Palacios-Rojas, N.; Tanumihardjo, S.A. Biofortified Orange Maize Enhances β-Cryptoxanthin Concentrations in Egg Yolks of Laying Hens Better than Tangerine Peel Fortificant. J. Agric. Food Chem. 2014, 62, 11892–11900. [Google Scholar] [CrossRef] [PubMed]
- Kang, K.R.; Cherian, G.; Sim, J.S. Tocopherols, Retinol and Carotenes in Chicken Egg and Tissues as Influenced by Dietary Palm Oil. J. Food Sci. 1998, 63, 592–596. [Google Scholar] [CrossRef]
- Harrison, E.H. Mechanisms involved in the intestinal absorption of dietary vitamin A and provitamin A carotenoids. Biochim. Biophys. Acta 2012, 1821, 70–77. [Google Scholar] [CrossRef]
- Quick, T.C.; Ong, D.E. Vitamin A metabolism in the human intestinal Caco-2 cell line. Biochemistry 1990, 29, 11116–11123. [Google Scholar] [CrossRef]
- Christian, P.; West, K.P., Jr. Interactions between zinc and vitamin A: An update. Am. J. Clin. Nutr. 1998, 68, 435S–441S. [Google Scholar] [CrossRef]
- Ong, D.E.; Chytil, F. Cellular retinol-binding protein from rat liver. Purification and characterization. J. Biol. Chem. 1978, 253, 828–832. [Google Scholar] [CrossRef]
- Chytil, F.; Ong, D.E. Cellular vitamin A binding proteins. Vitam. Horm. 1978, 36, 1–32. [Google Scholar] [CrossRef]
- Kanai, M.; Raz, A.; Goodman, D.S. Retinol-binding protein: The transport protein for vitamin A in human plasma. J. Clin. Investig. 1968, 47, 2025–2044. [Google Scholar] [CrossRef] [PubMed]
- Borel, P.; Lietz, G.; Goncalves, A.; Szabo de Edelenyi, F.; Lecompte, S.; Curtis, P.; Goumidi, L.; Caslake, M.J.; Miles, E.A.; Packard, C.; et al. CD36 and SR-BI are involved in cellular uptake of provitamin A carotenoids by Caco-2 and HEK cells, and some of their genetic variants are associated with plasma concentrations of these micronutrients in humans. J. Nutr. 2013, 143, 448–456. [Google Scholar] [CrossRef] [PubMed]
- Blomstrand, R.; Werner, B. Studies on the intestinal absorption of radioactive beta-carotene and vitamin A in man. Conversion of beta-carotene into vitamin A. Scand. J. Clin. Lab. Investig. 1967, 19, 339–345. [Google Scholar] [CrossRef]
- Moran, N.E.; Mohn, E.S.; Hason, N.; Erdman, J.W., Jr.; Johnson, E.J. Intrinsic and Extrinsic Factors Impacting Absorption, Metabolism, and Health Effects of Dietary Carotenoids. Adv. Nutr. 2018, 9, 465–492. [Google Scholar] [CrossRef]
- Glover, J.; Redfearn, E.R. The mechanism of the transformation of beta-carotene into vitamin A in vivo. Biochem. J. 1954, 58, xv–xvi. [Google Scholar]
- Napoli, J.L.; Race, K.R. Biogenesis of retinoic acid from beta-carotene. Differences between the metabolism of beta-carotene and retinal. J. Biol. Chem. 1988, 263, 17372–17377. [Google Scholar] [CrossRef]
- Lobo, G.P.; Amengual, J.; Baus, D.; Shivdasani, R.A.; Taylor, D.; von Lintig, J. Genetics and diet regulate vitamin A production via the homeobox transcription factor ISX. J. Biol. Chem. 2013, 288, 9017–9027. [Google Scholar] [CrossRef] [PubMed]
- van Bennekum, A.; Werder, M.; Thuahnai, S.T.; Han, C.H.; Duong, P.; Williams, D.L.; Wettstein, P.; Schulthess, G.; Phillips, M.C.; Hauser, H. Class B scavenger receptor-mediated intestinal absorption of dietary beta-carotene and cholesterol. Biochemistry 2005, 44, 4517–4525. [Google Scholar] [CrossRef] [PubMed]
- Lobo, G.P.; Hessel, S.; Eichinger, A.; Noy, N.; Moise, A.R.; Wyss, A.; Palczewski, K.; von Lintig, J. ISX is a retinoic acid-sensitive gatekeeper that controls intestinal beta, beta-carotene absorption and vitamin A production. FASEB J. 2010, 24, 1656–1666. [Google Scholar] [CrossRef] [PubMed]
- Hebuterne, X.; Wang, X.D.; Smith, D.E.; Tang, G.; Russell, R.M. In vivo biosynthesis of retinoic acid from beta-carotene involves and excentric cleavage pathway in ferret intestine. J. Lipid Res. 1996, 37, 482–492. [Google Scholar] [CrossRef]
- Harrison, E.H.; Hussain, M.M. Mechanisms involved in the intestinal digestion and absorption of dietary vitamin A. J. Nutr. 2001, 131, 1405–1408. [Google Scholar] [CrossRef]
- Blomhoff, R.; Blomhoff, H.K. Overview of retinoid metabolism and function. J. Neurobiol. 2006, 66, 606–630. [Google Scholar] [CrossRef]
- Blomhoff, R.; Green, M.H.; Berg, T.; Norum, K.R. Transport and storage of vitamin A. Science 1990, 250, 399–404. [Google Scholar] [CrossRef]
- Parker, R.S. Absorption, metabolism, and transport of carotenoids. FASEB J. 1996, 10, 542–551. [Google Scholar] [CrossRef]
- Jing, J.; Isoherranen, N.; Robinson-Cohen, C.; Petrie, I.; Kestenbaum, B.R.; Yeung, C.K. Chronic Kidney Disease Alters Vitamin A Homeostasis via Effects on Hepatic RBP4 Protein Expression and Metabolic Enzymes. Clin. Transl. Sci. 2016, 9, 207–215. [Google Scholar] [CrossRef]
- Sedjo, R.L.; Ranger-Moore, J.; Foote, J.; Craft, N.E.; Alberts, D.S.; Xu, M.J.; Giuliano, A.R. Circulating endogenous retinoic acid concentrations among participants enrolled in a randomized placebo-controlled clinical trial of retinyl palmitate. Cancer Epidemiol. Biomark. Prev. 2004, 13, 1687–1692. [Google Scholar]
- Harrison, E.H. Mechanisms of digestion and absorption of dietary vitamin A. Annu. Rev. Nutr. 2005, 25, 87–103. [Google Scholar] [CrossRef] [PubMed]
- Blomhoff, R.; Helgerud, P.; Rasmussen, M.; Berg, T.; Norum, K.R. In vivo uptake of chylomicron [3H]retinyl ester by rat liver: Evidence for retinol transfer from parenchymal to nonparenchymal cells. Proc. Natl. Acad. Sci. USA 1982, 79, 7326–7330. [Google Scholar] [CrossRef]
- Newcomer, M.E.; Ong, D.E. Plasma retinol binding protein: Structure and function of the prototypic lipocalin. Biochim. Biophys. Acta 2000, 1482, 57–64. [Google Scholar] [CrossRef]
- Soprano, D.R.; Soprano, K.J.; Goodman, D.S. Retinol-binding protein messenger RNA levels in the liver and in extrahepatic tissues of the rat. J. Lipid. Res. 1986, 27, 166–171. [Google Scholar] [CrossRef]
- Raz, A.; Goodman, D.S. The interaction of thyroxine with human plasma prealbumin and with the prealbumin-retinol-binding protein complex. J. Biol. Chem. 1969, 244, 3230–3237. [Google Scholar] [CrossRef]
- Goodman, D.S. Vitamin A and retinoids in health and disease. N. Engl. J. Med. 1984, 310, 1023–1031. [Google Scholar] [CrossRef] [PubMed]
- Peterson, P.A.; Berggard, I. Isolation and properties of a human retinol-transporting protein. J. Biol. Chem. 1971, 246, 25–33. [Google Scholar] [CrossRef]
- Vieira, M.; Saraiva, M.J. Transthyretin: A multifaceted protein. Biomol. Concepts 2014, 5, 45–54. [Google Scholar] [CrossRef]
- Episkopou, V.; Maeda, S.; Nishiguchi, S.; Shimada, K.; Gaitanaris, G.A.; Gottesman, M.E.; Robertson, E.J. Disruption of the transthyretin gene results in mice with depressed levels of plasma retinol and thyroid hormone. Proc. Natl. Acad. Sci. USA 1993, 90, 2375–2379. [Google Scholar] [CrossRef]
- Kawaguchi, R.; Yu, J.; Honda, J.; Hu, J.; Whitelegge, J.; Ping, P.; Wiita, P.; Bok, D.; Sun, H. A membrane receptor for retinol binding protein mediates cellular uptake of vitamin A. Science 2007, 315, 820–825. [Google Scholar] [CrossRef]
- Alapatt, P.; Guo, F.; Komanetsky, S.M.; Wang, S.; Cai, J.; Sargsyan, A.; Rodriguez Diaz, E.; Bacon, B.T.; Aryal, P.; Graham, T.E. Liver retinol transporter and receptor for serum retinol-binding protein (RBP4). J. Biol. Chem. 2013, 288, 1250–1265. [Google Scholar] [CrossRef] [PubMed]
- Berry, D.C.; O’Byrne, S.M.; Vreeland, A.C.; Blaner, W.S.; Noy, N. Cross talk between signaling and vitamin A transport by the retinol-binding protein receptor STRA6. Mol. Cell. Biol. 2012, 32, 3164–3175. [Google Scholar] [CrossRef] [PubMed]
- Paik, J.; Vogel, S.; Quadro, L.; Piantedosi, R.; Gottesman, M.; Lai, K.; Hamberger, L.; Vieira Mde, M.; Blaner, W.S. Vitamin A: Overlapping delivery pathways to tissues from the circulation. J. Nutr. 2004, 134, 276S–280S. [Google Scholar] [CrossRef] [PubMed]
- Senoo, H. Structure and function of hepatic stellate cells. Med. Electron. Microsc. 2004, 37, 3–15. [Google Scholar] [CrossRef] [PubMed]
- Futterman, S.; Andrews, J.S. The Composition of Liver Vitamin a Ester and the Synthesis of Vitamin a Ester by Liver Microsomes. J. Biol. Chem. 1964, 239, 4077–4080. [Google Scholar] [CrossRef]
- Wyss, R.; Bucheli, F. Determination of endogenous levels of 13-cis-retinoic acid (isotretinoin), all-trans-retinoic acid (tretinoin) and their 4-oxo metabolites in human and animal plasma by high-performance liquid chromatography with automated column switching and ultraviolet detection. J. Chromatogr. B Biomed. Sci. Appl. 1997, 700, 31–47. [Google Scholar] [CrossRef]
- Barua, A.B.; Sidell, N. Retinoyl beta-glucuronide: A biologically active interesting retinoid. J. Nutr. 2004, 134, 286S–289S. [Google Scholar] [CrossRef]
- Kurlandsky, S.B.; Gamble, M.V.; Ramakrishnan, R.; Blaner, W.S. Plasma delivery of retinoic acid to tissues in the rat. J. Biol. Chem. 1995, 270, 17850–17857. [Google Scholar] [CrossRef]
- Barua, A.B.; Olson, J.A. Retinoyl beta-glucuronide: An endogenous compound of human blood. Am. J. Clin. Nutr. 1986, 43, 481–485. [Google Scholar] [CrossRef]
- Jung, U.S.; Jeong, K.J.; Kang, J.K.; Yi, K.; Shin, J.H.; Seo, H.S.; Kim, T.; Kim, S.H.; Hur, J.Y. Effects of estrogen receptor alpha and beta on the expression of visfatin and retinol-binding protein 4 in 3T3-L1 adipocytes. Int. J. Mol. Med. 2013, 32, 723–728. [Google Scholar] [CrossRef][Green Version]
- Clugston, R.D.; Blaner, W.S. The adverse effects of alcohol on vitamin A metabolism. Nutrients 2012, 4, 356–371. [Google Scholar] [CrossRef]
- Berggren Soderlund, M.; Fex, G.A.; Nilsson-Ehle, P. Concentrations of retinoids in early pregnancy and in newborns and their mothers. Am. J. Clin. Nutr. 2005, 81, 633–636. [Google Scholar] [CrossRef][Green Version]
- Dancis, J.; Levitz, M.; Katz, J.; Wilson, D.; Blaner, W.S.; Piantedosi, R.; Goodman, D.S. Transfer and metabolism of retinol by the perfused human placenta. Pediatr. Res. 1992, 32, 195–199. [Google Scholar] [CrossRef]
- Spiegler, E.; Kim, Y.K.; Wassef, L.; Shete, V.; Quadro, L. Maternal-fetal transfer and metabolism of vitamin A and its precursor beta-carotene in the developing tissues. Biochim. Biophys. Acta 2012, 1821, 88–98. [Google Scholar] [CrossRef]
- Quadro, L.; Hamberger, L.; Gottesman, M.E.; Colantuoni, V.; Ramakrishnan, R.; Blaner, W.S. Transplacental delivery of retinoid: The role of retinol-binding protein and lipoprotein retinyl ester. Am. J. Physiol. Endocrinol. Metab. 2004, 286, E844–E851. [Google Scholar] [CrossRef] [PubMed]
- Bastos Maia, S.; Rolland Souza, A.; Costa Caminha, M.; Lins Da Silva, S.; Callou Cruz, R.; Carvalho Dos Santos, C.; Batista Filho, M. Vitamin A and Pregnancy: A Narrative Review. Nutrients 2019, 11, 681. [Google Scholar] [CrossRef]
- Rothman, K.J.; Moore, L.L.; Singer, M.R.; Nguyen, U.S.; Mannino, S.; Milunsky, A. Teratogenicity of high vitamin A intake. N. Engl. J. Med. 1995, 333, 1369–1373. [Google Scholar] [CrossRef] [PubMed]
- Hunt, J.R. Teratogenicity of high vitamin A intake. N. Engl. J. Med. 1996, 334, 1197. [Google Scholar] [CrossRef]
- Maden, M. Retinoid signalling in the development of the central nervous system. Nat. Rev. Neurosci. 2002, 3, 843–853. [Google Scholar] [CrossRef] [PubMed]
- Maden, M. Retinoids in lung development and regeneration. Curr. Top. Dev. Biol. 2004, 61, 153–189. [Google Scholar] [CrossRef]
- Clagett-Dame, M.; DeLuca, H.F. The role of vitamin A in mammalian reproduction and embryonic development. Annu. Rev. Nutr. 2002, 22, 347–381. [Google Scholar] [CrossRef] [PubMed]
- Niederreither, K.; Dolle, P. Retinoic acid in development: Towards an integrated view. Nat. Rev. Genet. 2008, 9, 541–553. [Google Scholar] [CrossRef] [PubMed]
- Niles, R.M. Vitamin A (retinoids) regulation of mouse melanoma growth and differentiation. J. Nutr. 2003, 133, 282S–286S. [Google Scholar] [CrossRef] [PubMed]
- Dawson, M.I. The importance of vitamin A in nutrition. Curr. Pharm. Des. 2000, 6, 311–325. [Google Scholar] [CrossRef]
- Morriss-Kay, G.M.; Sokolova, N. Embryonic development and pattern formation. FASEB J. 1996, 10, 961–968. [Google Scholar] [CrossRef]
- Gudas, L.J.; Wagner, J.A. Retinoids regulate stem cell differentiation. J. Cell. Physiol. 2011, 226, 322–330. [Google Scholar] [CrossRef]
- Eskild, W.; Simard, J.; Hansson, V.; Guerin, S.L. Binding of a member of the NF1 family of transcription factors to two distinct cis-acting elements in the promoter and 5’-flanking region of the human cellular retinol binding protein 1 gene. Mol. Endocrinol. 1994, 8, 732–745. [Google Scholar] [CrossRef][Green Version]
- Bono, M.R.; Tejon, G.; Flores-Santibanez, F.; Fernandez, D.; Rosemblatt, M.; Sauma, D. Retinoic Acid as a Modulator of T Cell Immunity. Nutrients 2016, 8, 349. [Google Scholar] [CrossRef]
- Shearer, K.D.; Stoney, P.N.; Morgan, P.J.; McCaffery, P.J. A vitamin for the brain. Trends Neurosci. 2012, 35, 733–741. [Google Scholar] [CrossRef]
- Cocco, S.; Diaz, G.; Stancampiano, R.; Diana, A.; Carta, M.; Curreli, R.; Sarais, L.; Fadda, F. Vitamin A deficiency produces spatial learning and memory impairment in rats. Neuroscience 2002, 115, 475–482. [Google Scholar] [CrossRef]
- Sommer, A. Uses and misuses of vitamin A. Curr. Issues Public Health 1996, 2, 161–164. [Google Scholar]
- Bonet, M.L.; Ribot, J.; Felipe, F.; Palou, A. Vitamin A and the regulation of fat reserves. Cell Mol. Life Sci. 2003, 60, 1311–1321. [Google Scholar] [CrossRef]
- Villarroya, F.; Iglesias, R.; Giralt, M. Retinoids and retinoid receptors in the control of energy balance: Novel pharmacological strategies in obesity and diabetes. Curr. Med. Chem. 2004, 11, 795–805. [Google Scholar] [CrossRef]
- Niles, R.M. Signaling pathways in retinoid chemoprevention and treatment of cancer. Mutat. Res. 2004, 555, 81–96. [Google Scholar] [CrossRef]
- Soprano, D.R.; Qin, P.; Soprano, K.J. Retinoic acid receptors and cancers. Annu. Rev. Nutr. 2004, 24, 201–221. [Google Scholar] [CrossRef]
- Yamauchi, P.S.; Rizk, D.; Lowe, N.J. Retinoid therapy for psoriasis. Dermatol. Clin. 2004, 22, 467–476. [Google Scholar] [CrossRef]
- Genaro Pde, S.; Martini, L.A. Vitamin A supplementation and risk of skeletal fracture. Nutr. Rev. 2004, 62, 65–67. [Google Scholar] [CrossRef]
- Gudas, L.J. Emerging roles for retinoids in regeneration and differentiation in normal and disease states. Biochim. Biophys. Acta 2012, 1821, 213–221. [Google Scholar] [CrossRef]
- von Lintig, J. Metabolism of carotenoids and retinoids related to vision. J. Biol. Chem. 2012, 287, 1627–1634. [Google Scholar] [CrossRef]
- Zhong, M.; Kawaguchi, R.; Kassai, M.; Sun, H. Retina, retinol, retinal and the natural history of vitamin A as a light sensor. Nutrients 2012, 4, 2069–2096. [Google Scholar] [CrossRef]
- Perusek, L.; Maeda, T. Vitamin A derivatives as treatment options for retinal degenerative diseases. Nutrients 2013, 5, 2646–2666. [Google Scholar] [CrossRef] [PubMed]
- McCaffery, P.; Mey, J.; Drager, U.C. Light-mediated retinoic acid production. Proc. Natl. Acad. Sci. USA 1996, 93, 12570–12574. [Google Scholar] [CrossRef]
- Wolf, G. The discovery of the visual function of vitamin A. J. Nutr. 2001, 131, 1647–1650. [Google Scholar] [CrossRef] [PubMed]
- Balmer, J.E.; Blomhoff, R. Gene expression regulation by retinoic acid. J. Lipid Res. 2002, 43, 1773–1808. [Google Scholar] [CrossRef]
- Bohn, T. Carotenoids, Chronic Disease Prevention and Dietary Recommendations. Int. J. Vitam. Nutr. Res. 2017, 87, 121–130. [Google Scholar] [CrossRef]
- di Masi, A.; De Marinis, E.; Ascenzi, P.; Marino, M. Nuclear receptors CAR and PXR: Molecular, functional, and biomedical aspects. Mol. Aspects Med. 2009, 30, 297–343. [Google Scholar] [CrossRef]
- Giguere, V.; Ong, E.S.; Segui, P.; Evans, R.M. Identification of a receptor for the morphogen retinoic acid. Nature 1987, 330, 624–629. [Google Scholar] [CrossRef]
- Petkovich, M.; Brand, N.J.; Krust, A.; Chambon, P. A human retinoic acid receptor which belongs to the family of nuclear receptors. Nature 1987, 330, 444–450. [Google Scholar] [CrossRef]
- Krust, A.; Kastner, P.; Petkovich, M.; Zelent, A.; Chambon, P. A third human retinoic acid receptor, hRAR-gamma. Proc. Natl. Acad. Sci. USA 1989, 86, 5310–5314. [Google Scholar] [CrossRef]
- Brand, N.; Petkovich, M.; Krust, A.; Chambon, P.; de The, H.; Marchio, A.; Tiollais, P.; Dejean, A. Identification of a second human retinoic acid receptor. Nature 1988, 332, 850–853. [Google Scholar] [CrossRef]
- Germain, P.; Chambon, P.; Eichele, G.; Evans, R.M.; Lazar, M.A.; Leid, M.; De Lera, A.R.; Lotan, R.; Mangelsdorf, D.J.; Gronemeyer, H. International Union of Pharmacology. LXIII. Retinoid X receptors. Pharmacol. Rev. 2006, 58, 760–772. [Google Scholar] [CrossRef]
- Canon, E.; Cosgaya, J.M.; Scsucova, S.; Aranda, A. Rapid effects of retinoic acid on CREB and ERK phosphorylation in neuronal cells. Mol. Biol. Cell. 2004, 15, 5583–5592. [Google Scholar] [CrossRef]
- Poon, M.M.; Chen, L. Retinoic acid-gated sequence-specific translational control by RARalpha. Proc. Natl. Acad. Sci. USA 2008, 105, 20303–20308. [Google Scholar] [CrossRef]
- Mangelsdorf, D.J.; Ong, E.S.; Dyck, J.A.; Evans, R.M. Nuclear receptor that identifies a novel retinoic acid response pathway. Nature 1990, 345, 224–229. [Google Scholar] [CrossRef]
- Yu, V.C.; Delsert, C.; Andersen, B.; Holloway, J.M.; Devary, O.V.; Naar, A.M.; Kim, S.Y.; Boutin, J.M.; Glass, C.K.; Rosenfeld, M.G. RXR beta: A coregulator that enhances binding of retinoic acid, thyroid hormone, and vitamin D receptors to their cognate response elements. Cell 1991, 67, 1251–1266. [Google Scholar] [CrossRef]
- Mangelsdorf, D.J.; Borgmeyer, U.; Heyman, R.A.; Zhou, J.Y.; Ong, E.S.; Oro, A.E.; Kakizuka, A.; Evans, R.M. Characterization of three RXR genes that mediate the action of 9-cis retinoic acid. Genes Dev. 1992, 6, 329–344. [Google Scholar] [CrossRef]
- Heyman, R.A.; Mangelsdorf, D.J.; Dyck, J.A.; Stein, R.B.; Eichele, G.; Evans, R.M.; Thaller, C. 9-cis retinoic acid is a high affinity ligand for the retinoid X receptor. Cell 1992, 68, 397–406. [Google Scholar] [CrossRef]
- Chebaro, Y.; Sirigu, S.; Amal, I.; Lutzing, R.; Stote, R.H.; Rochette-Egly, C.; Rochel, N.; Dejaegere, A. Allosteric Regulation in the Ligand Binding Domain of Retinoic Acid Receptorgamma. PLoS ONE 2017, 12, e0171043. [Google Scholar] [CrossRef]
- Le Douarin, B.; Zechel, C.; Garnier, J.M.; Lutz, Y.; Tora, L.; Pierrat, P.; Heery, D.; Gronemeyer, H.; Chambon, P.; Losson, R. The N-terminal part of TIF1, a putative mediator of the ligand-dependent activation function (AF-2) of nuclear receptors, is fused to B-raf in the oncogenic protein T18. EMBO J. 1995, 14, 2020–2033. [Google Scholar] [CrossRef]
- Durand, B.; Saunders, M.; Gaudon, C.; Roy, B.; Losson, R.; Chambon, P. Activation function 2 (AF-2) of retinoic acid receptor and 9-cis retinoic acid receptor: Presence of a conserved autonomous constitutive activating domain and influence of the nature of the response element on AF-2 activity. EMBO J. 1994, 13, 5370–5382. [Google Scholar] [CrossRef]
- Lindh, J.D.; Bjorkhem-Bergman, L.; Eliasson, E. Vitamin D and drug-metabolising enzymes. Photochem. Photobiol. Sci. 2012, 11, 1797–1801. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Luo, X.Y.; Wu, D.H.; Xu, Y. ROR nuclear receptors: Structures, related diseases, and drug discovery. Acta Pharmacol. Sin. 2015, 36, 71–87. [Google Scholar] [CrossRef]
- Jetten, A.M.; Kurebayashi, S.; Ueda, E. The ROR nuclear orphan receptor subfamily: Critical regulators of multiple biological processes. Prog. Nucleic. Acid Res. Mol. Biol. 2001, 69, 205–247. [Google Scholar] [CrossRef]
- Solt, L.A.; Burris, T.P. Action of RORs and their ligands in (patho)physiology. Trends Endocrinol. Metab. 2012, 23, 619–627. [Google Scholar] [CrossRef]
- Stehlin-Gaon, C.; Willmann, D.; Zeyer, D.; Sanglier, S.; Van Dorsselaer, A.; Renaud, J.P.; Moras, D.; Schule, R. All-trans retinoic acid is a ligand for the orphan nuclear receptor ROR beta. Nat. Struct. Biol. 2003, 10, 820–825. [Google Scholar] [CrossRef]
- Napoli, J.L. Retinoic acid biosynthesis and metabolism. FASEB J. 1996, 10, 993–1001. [Google Scholar] [CrossRef]
- Schug, T.T.; Berry, D.C.; Shaw, N.S.; Travis, S.N.; Noy, N. Opposing effects of retinoic acid on cell growth result from alternate activation of two different nuclear receptors. Cell 2007, 129, 723–733. [Google Scholar] [CrossRef] [PubMed]
- Hihi, A.K.; Michalik, L.; Wahli, W. PPARs: Transcriptional effectors of fatty acids and their derivatives. Cell. Mol. Life Sci. 2002, 59, 790–798. [Google Scholar] [CrossRef]
- Chandra, V.; Huang, P.; Hamuro, Y.; Raghuram, S.; Wang, Y.; Burris, T.P.; Rastinejad, F. Structure of the intact PPAR-gamma-RXR- nuclear receptor complex on DNA. Nature 2008, 456, 350–356. [Google Scholar] [CrossRef]
- Michalik, L.; Auwerx, J.; Berger, J.P.; Chatterjee, V.K.; Glass, C.K.; Gonzalez, F.J.; Grimaldi, P.A.; Kadowaki, T.; Lazar, M.A.; O’Rahilly, S.; et al. International Union of Pharmacology. LXI. Peroxisome proliferator-activated receptors. Pharmacol. Rev. 2006, 58, 726–741. [Google Scholar] [CrossRef]
- Barish, G.D.; Narkar, V.A.; Evans, R.M. PPAR delta: A dagger in the heart of the metabolic syndrome. J. Clin. Investig. 2006, 116, 590–597. [Google Scholar] [CrossRef] [PubMed]
- Krey, G.; Braissant, O.; L’Horset, F.; Kalkhoven, E.; Perroud, M.; Parker, M.G.; Wahli, W. Fatty acids, eicosanoids, and hypolipidemic agents identified as ligands of peroxisome proliferator-activated receptors by coactivator-dependent receptor ligand assay. Mol. Endocrinol. 1997, 11, 779–791. [Google Scholar] [CrossRef]
- Kliewer, S.A.; Forman, B.M.; Blumberg, B.; Ong, E.S.; Borgmeyer, U.; Mangelsdorf, D.J.; Umesono, K.; Evans, R.M. Differential expression and activation of a family of murine peroxisome proliferator-activated receptors. Proc. Natl. Acad. Sci. USA 1994, 91, 7355–7359. [Google Scholar] [CrossRef] [PubMed]
- Glatz, J.F.; Lagarde, M. Lipid sensing and lipid sensors. Cell. Mol. Life Sci. 2007, 64, 2449–2451. [Google Scholar] [CrossRef]
- Xu, H.E.; Lambert, M.H.; Montana, V.G.; Parks, D.J.; Blanchard, S.G.; Brown, P.J.; Sternbach, D.D.; Lehmann, J.M.; Wisely, G.B.; Willson, T.M.; et al. Molecular recognition of fatty acids by peroxisome proliferator-activated receptors. Mol. Cell. 1999, 3, 397–403. [Google Scholar] [CrossRef]
- Berry, D.C.; Noy, N. Is PPARbeta/delta a Retinoid Receptor? PPAR Res. 2007, 2007, 73256. [Google Scholar] [CrossRef] [PubMed]
- Berry, D.C.; Noy, N. All-trans-retinoic acid represses obesity and insulin resistance by activating both peroxisome proliferation-activated receptor beta/delta and retinoic acid receptor. Mol. Cell. Biol 2009, 29, 3286–3296. [Google Scholar] [CrossRef]
- Lampen, A.; Meyer, S.; Nau, H. Effects of receptor-selective retinoids on CYP26 gene expression and metabolism of all-trans-retinoic acid in intestinal cells. Drug Metab. Dispos. 2001, 29, 742–747. [Google Scholar]
- White, J.A.; Ramshaw, H.; Taimi, M.; Stangle, W.; Zhang, A.; Everingham, S.; Creighton, S.; Tam, S.P.; Jones, G.; Petkovich, M. Identification of the human cytochrome P450, P450RAI-2, which is predominantly expressed in the adult cerebellum and is responsible for all-trans-retinoic acid metabolism. Proc. Natl. Acad. Sci. USA 2000, 97, 6403–6408. [Google Scholar] [CrossRef]
- Taimi, M.; Helvig, C.; Wisniewski, J.; Ramshaw, H.; White, J.; Amad, M.; Korczak, B.; Petkovich, M. A novel human cytochrome P450, CYP26C1, involved in metabolism of 9-cis and all-trans isomers of retinoic acid. J. Biol. Chem. 2004, 279, 77–85. [Google Scholar] [CrossRef]
- Reijntjes, S.; Gale, E.; Maden, M. Generating gradients of retinoic acid in the chick embryo: Cyp26C1 expression and a comparative analysis of the Cyp26 enzymes. Dev. Dyn. 2004, 230, 509–517. [Google Scholar] [CrossRef]
- Cawley, S.; Bekiranov, S.; Ng, H.H.; Kapranov, P.; Sekinger, E.A.; Kampa, D.; Piccolboni, A.; Sementchenko, V.; Cheng, J.; Williams, A.J.; et al. Unbiased mapping of transcription factor binding sites along human chromosomes 21 and 22 points to widespread regulation of noncoding RNAs. Cell 2004, 116, 499–509. [Google Scholar] [CrossRef]
- Chen, J.Y.; Penco, S.; Ostrowski, J.; Balaguer, P.; Pons, M.; Starrett, J.E.; Reczek, P.; Chambon, P.; Gronemeyer, H. RAR-specific agonist/antagonists which dissociate transactivation and AP1 transrepression inhibit anchorage-independent cell proliferation. EMBO J. 1995, 14, 1187–1197. [Google Scholar] [CrossRef]
- Lokman, N.A.; Ho, R.; Gunasegaran, K.; Bonner, W.M.; Oehler, M.K.; Ricciardelli, C. Anti-tumour effects of all-trans retinoid acid on serous ovarian cancer. J. Exp. Clin. Cancer Res. 2019, 38, 10. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Liu, Y.; Qi, G.; Brand, D.; Zheng, S.G. Role of Vitamin A in the Immune System. J. Clin. Med. 2018, 7, 258. [Google Scholar] [CrossRef]
- Altucci, L.; Gronemeyer, H. The promise of retinoids to fight against cancer. Nat. Rev. Cancer 2001, 1, 181–193. [Google Scholar] [CrossRef]
- Nagy, L.; Thomazy, V.A.; Heyman, R.A.; Davies, P.J. Retinoid-induced apoptosis in normal and neoplastic tissues. Cell Death Differ. 1998, 5, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Mrass, P.; Rendl, M.; Mildner, M.; Gruber, F.; Lengauer, B.; Ballaun, C.; Eckhart, L.; Tschachler, E. Retinoic acid increases the expression of p53 and proapoptotic caspases and sensitizes keratinocytes to apoptosis: A possible explanation for tumor preventive action of retinoids. Cancer Res. 2004, 64, 6542–6548. [Google Scholar] [CrossRef]
- Maalmi, H.; Walter, V.; Jansen, L.; Owen, R.W.; Ulrich, A.; Schottker, B.; Chang-Claude, J.; Hoffmeister, M.; Brenner, H. Dose-Response Relationship between Serum Retinol Levels and Survival in Patients with Colorectal Cancer: Results from the DACHS Study. Nutrients 2018, 10, 510. [Google Scholar] [CrossRef]
- Huang, X.; Gao, Y.; Zhi, X.; Ta, N.; Jiang, H.; Zheng, J. Association between vitamin A, retinol and carotenoid intake and pancreatic cancer risk: Evidence from epidemiologic studies. Sci. Rep. 2016, 6, 38936. [Google Scholar] [CrossRef]
- Costantini, L.; Molinari, R.; Farinon, B.; Merendino, N. Retinoic Acids in the Treatment of Most Lethal Solid Cancers. J. Clin. Med. 2020, 9, 360. [Google Scholar] [CrossRef] [PubMed]
- Bama, E.S.; Grace, V.M.B.; Sundaram, V.; Jesubatham, P.D. Synergistic effect of co-treatment with all-trans retinoic acid and 9-cis retinoic acid on human lung cancer cell line at molecular level. 3 Biotech 2019, 9, 159. [Google Scholar] [CrossRef]
- Esteva, F.J.; Glaspy, J.; Baidas, S.; Laufman, L.; Hutchins, L.; Dickler, M.; Tripathy, D.; Cohen, R.; DeMichele, A.; Yocum, R.C.; et al. Multicenter phase II study of oral bexarotene for patients with metastatic breast cancer. J. Clin. Oncol. 2003, 21, 999–1006. [Google Scholar] [CrossRef]
- Rousseau, E.J.; Davison, A.J.; Dunn, B. Protection by beta-carotene and related compounds against oxygen-mediated cytotoxicity and genotoxicity: Implications for carcinogenesis and anticarcinogenesis. Free Radic. Biol. Med. 1992, 13, 407–433. [Google Scholar] [CrossRef]
- Monsen, E.R. Dietary reference intakes for the antioxidant nutrients: Vitamin C, vitamin E, selenium, and carotenoids. J. Am. Diet. Assoc. 2000, 100, 637–640. [Google Scholar] [CrossRef]
- Blumberg, J.; Block, G. The Alpha-Tocopherol, Beta-Carotene Cancer Prevention Study in Finland. Nutr. Rev. 1994, 52, 242–245. [Google Scholar] [CrossRef] [PubMed]
- Omenn, G.S.; Goodman, G.E.; Thornquist, M.D.; Balmes, J.; Cullen, M.R.; Glass, A.; Keogh, J.P.; Meyskens, F.L.; Valanis, B.; Williams, J.H.; et al. Effects of a combination of beta carotene and vitamin A on lung cancer and cardiovascular disease. N. Engl. J. Med. 1996, 334, 1150–1155. [Google Scholar] [CrossRef]
- Buring, J.E.; Hebert, P.; Hennekens, C.H. The alpha-tocopherol, beta-carotene lung cancer prevention trial of vitamin E and beta-carotene: The beginning of the answers. Ann. Epidemiol. 1994, 4, 75. [Google Scholar] [CrossRef]
- Middha, P.; Weinstein, S.J.; Mannisto, S.; Albanes, D.; Mondul, A.M. beta-Carotene Supplementation and Lung Cancer Incidence in the Alpha-Tocopherol, Beta-Carotene Cancer Prevention Study: The Role of Tar and Nicotine. Nicotine Tob. Res. 2019, 21, 1045–1050. [Google Scholar] [CrossRef] [PubMed]
- Nishino, H.; Murakosh, M.; Ii, T.; Takemura, M.; Kuchide, M.; Kanazawa, M.; Mou, X.Y.; Wada, S.; Masuda, M.; Ohsaka, Y.; et al. Carotenoids in cancer chemoprevention. Cancer Metastasis Rev. 2002, 21, 257–264. [Google Scholar] [CrossRef]
- Koklesova, L.; Liskova, A.; Samec, M.; Buhrmann, C.; Samuel, S.M.; Varghese, E.; Ashrafizadeh, M.; Najafi, M.; Shakibaei, M.; Busselberg, D.; et al. Carotenoids in Cancer Apoptosis-The Road from Bench to Bedside and Back. Cancers 2020, 12, 2425. [Google Scholar] [CrossRef] [PubMed]
- Park, H.A.; Hayden, M.M.; Bannerman, S.; Jansen, J.; Crowe-White, K.M. Anti-Apoptotic Effects of Carotenoids in Neurodegeneration. Molecules 2020, 25, 3453. [Google Scholar] [CrossRef] [PubMed]
- Iwata, M.; Hirakiyama, A.; Eshima, Y.; Kagechika, H.; Kato, C.; Song, S.Y. Retinoic acid imprints gut-homing specificity on T cells. Immunity 2004, 21, 527–538. [Google Scholar] [CrossRef]
- Benson, M.J.; Pino-Lagos, K.; Rosemblatt, M.; Noelle, R.J. All-trans retinoic acid mediates enhanced T reg cell growth, differentiation, and gut homing in the face of high levels of co-stimulation. J. Exp. Med. 2007, 204, 1765–1774. [Google Scholar] [CrossRef] [PubMed]
- Pino-Lagos, K.; Benson, M.J.; Noelle, R.J. Retinoic acid in the immune system. Ann. N. Y. Acad. Sci. 2008, 1143, 170–187. [Google Scholar] [CrossRef] [PubMed]
- Moore, C.; Sauma, D.; Reyes, P.A.; Morales, J.; Rosemblatt, M.; Bono, M.R.; Fierro, J.A. Dendritic cells and B cells cooperate in the generation of CD4(+)CD25(+)FOXP3(+) allogeneic T cells. Transplant. Proc. 2010, 42, 371–375. [Google Scholar] [CrossRef]
- Ross, A.C. Vitamin A and retinoic acid in T cell-related immunity. Am. J. Clin. Nutr. 2012, 96, 1166S–1172S. [Google Scholar] [CrossRef]
- Bayon, Y.; Ortiz, M.A.; Lopez-Hernandez, F.J.; Gao, F.; Karin, M.; Pfahl, M.; Piedrafita, F.J. Inhibition of IkappaB kinase by a new class of retinoid-related anticancer agents that induce apoptosis. Mol. Cell. Biol. 2003, 23, 1061–1074. [Google Scholar] [CrossRef]
- Adams, J.; Kiss, E.; Arroyo, A.B.; Bonrouhi, M.; Sun, Q.; Li, Z.; Gretz, N.; Schnitger, A.; Zouboulis, C.C.; Wiesel, M.; et al. 13-cis retinoic acid inhibits development and progression of chronic allograft nephropathy. Am. J. Pathol. 2005, 167, 285–298. [Google Scholar] [CrossRef]
- Shibakura, M.; Niiya, K.; Niiya, M.; Asaumi, N.; Yoshida, C.; Nakata, Y.; Tanimoto, M. Induction of CXC and CC chemokines by all-trans retinoic acid in acute promyelocytic leukemia cells. Leuk. Res. 2005, 29, 755–759. [Google Scholar] [CrossRef]
- Buck, J.; Derguini, F.; Levi, E.; Nakanishi, K.; Hammerling, U. Intracellular signaling by 14-hydroxy-4,14-retro-retinol. Science 1991, 254, 1654–1656. [Google Scholar] [CrossRef]
- Buck, J.; Grun, F.; Derguini, F.; Chen, Y.; Kimura, S.; Noy, N.; Hammerling, U. Anhydroretinol: A naturally occurring inhibitor of lymphocyte physiology. J. Exp. Med. 1993, 178, 675–680. [Google Scholar] [CrossRef]
- Mora, J.R.; Iwata, M.; von Andrian, U.H. Vitamin effects on the immune system: Vitamins A and D take centre stage. Nat. Rev. Immunol. 2008, 8, 685–698. [Google Scholar] [CrossRef]
- De Luca, L.; Wolf, G. Mechanism of action of vitamin A in differentiation of mucus-secreting epithelia. J. Agric. Food Chem. 1972, 20, 474–476. [Google Scholar] [CrossRef]
- Aydelotte, M.B. Vitamin A deficiency in chickens. Br. J. Nutr. 1963, 17, 205–210. [Google Scholar] [CrossRef]
- McCullough, F.S.; Northrop-Clewes, C.A.; Thurnham, D.I. The effect of vitamin A on epithelial integrity. Proc. Nutr. Soc. 1999, 58, 289–293. [Google Scholar] [CrossRef]
- Wolbach, S.B.; Howe, P.R. Tissue Changes Following Deprivation of Fat-Soluble a Vitamin. J. Exp. Med. 1925, 42, 753–777. [Google Scholar] [CrossRef]
- Fu, P.P.; Xia, Q.; Boudreau, M.D.; Howard, P.C.; Tolleson, W.H.; Wamer, W.G. Physiological role of retinyl palmitate in the skin. Vitam. Horm. 2007, 75, 223–256. [Google Scholar] [CrossRef]
- Chung, S.S.; Wolgemuth, D.J. Role of retinoid signaling in the regulation of spermatogenesis. Cytogenet. Genome Res. 2004, 105, 189–202. [Google Scholar] [CrossRef]
- Clagett-Dame, M.; Knutson, D. Vitamin A in reproduction and development. Nutrients 2011, 3, 385–428. [Google Scholar] [CrossRef]
- Baume, L.J.; Franquin, J.C.; Korner, W.W. The prenatal effects of maternal vitamin A deficiency on the cranial and dental development of the progeny. Am. J. Orthod. 1972, 62, 447–460. [Google Scholar] [CrossRef]
- Stoltzfus, R.J.; Underwood, B.A. Breast-milk vitamin A as an indicator of the vitamin A status of women and infants. Bull. World Health Organ. 1995, 73, 703–711. [Google Scholar]
- Dror, D.K.; Allen, L.H. Retinol-to-Fat Ratio and Retinol Concentration in Human Milk Show Similar Time Trends and Associations with Maternal Factors at the Population Level: A Systematic Review and Meta-Analysis. Adv. Nutr. 2018, 9, 332S–346S. [Google Scholar] [CrossRef] [PubMed]
- Varani, J.; Fisher, G.J.; Kang, S.; Voorhees, J.J. Molecular mechanisms of intrinsic skin aging and retinoid-induced repair and reversal. J. Investig. Dermatol. Symp. Proc. 1998, 3, 57–60. [Google Scholar] [PubMed]
- Wang, Y.X.; Lee, C.H.; Tiep, S.; Yu, R.T.; Ham, J.; Kang, H.; Evans, R.M. Peroxisome-proliferator-activated receptor delta activates fat metabolism to prevent obesity. Cell 2003, 113, 159–170. [Google Scholar] [CrossRef]
- Schwarz, E.J.; Reginato, M.J.; Shao, D.; Krakow, S.L.; Lazar, M.A. Retinoic acid blocks adipogenesis by inhibiting C/EBPbeta-mediated transcription. Mol. Cell Biol. 1997, 17, 1552–1561. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Graham, T.E.; Mody, N.; Preitner, F.; Peroni, O.D.; Zabolotny, J.M.; Kotani, K.; Quadro, L.; Kahn, B.B. Serum retinol binding protein 4 contributes to insulin resistance in obesity and type 2 diabetes. Nature 2005, 436, 356–362. [Google Scholar] [CrossRef]
- Ziouzenkova, O.; Orasanu, G.; Sharlach, M.; Akiyama, T.E.; Berger, J.P.; Viereck, J.; Hamilton, J.A.; Tang, G.; Dolnikowski, G.G.; Vogel, S.; et al. Retinaldehyde represses adipogenesis and diet-induced obesity. Nat. Med. 2007, 13, 695–702. [Google Scholar] [CrossRef]
- Lima, I.O.L.; Peres, W.A.F.; Cruz, S.; Ramalho, A. Association of Ischemic Cardiovascular Disease with Inadequacy of Liver Store of Retinol in Elderly Individuals. Oxid. Med. Cell Longev. 2018, 2018, 9785231. [Google Scholar] [CrossRef]
- Brazionis, L.; Walker, K.Z.; Itsiopoulos, C.; O’Dea, K. Plasma retinol: A novel marker for cardiovascular disease mortality in Australian adults. Nutr. Metab. Cardiovasc. Dis. 2012, 22, 914–920. [Google Scholar] [CrossRef]
- Beydoun, M.A.; Chen, X.; Jha, K.; Beydoun, H.A.; Zonderman, A.B.; Canas, J.A. Carotenoids, vitamin A, and their association with the metabolic syndrome: A systematic review and meta-analysis. Nutr. Rev. 2019, 77, 32–45. [Google Scholar] [CrossRef]
- Garcia, O.P.; Long, K.Z.; Rosado, J.L. Impact of micronutrient deficiencies on obesity. Nutr. Rev. 2009, 67, 559–572. [Google Scholar] [CrossRef]
- Mounien, L.; Tourniaire, F.; Landrier, J.F. Anti-Obesity Effect of Carotenoids: Direct Impact on Adipose Tissue and Adipose Tissue-Driven Indirect Effects. Nutrients 2019, 11, 1562. [Google Scholar] [CrossRef]
- Kane, M.A.; Folias, A.E.; Pingitore, A.; Perri, M.; Obrochta, K.M.; Krois, C.R.; Cione, E.; Ryu, J.Y.; Napoli, J.L. Identification of 9-cis-retinoic acid as a pancreas-specific autacoid that attenuates glucose-stimulated insulin secretion. Proc. Natl. Acad. Sci. USA 2010, 107, 21884–21889. [Google Scholar] [CrossRef]
- Brun, P.J.; Yang, K.J.; Lee, S.A.; Yuen, J.J.; Blaner, W.S. Retinoids: Potent regulators of metabolism. Biofactors 2013, 39, 151–163. [Google Scholar] [CrossRef]
- Trasino, S.E.; Gudas, L.J. Vitamin A: A missing link in diabetes? Diabetes Manag. 2015, 5, 359–367. [Google Scholar] [CrossRef]
- Bowles, J.; Knight, D.; Smith, C.; Wilhelm, D.; Richman, J.; Mamiya, S.; Yashiro, K.; Chawengsaksophak, K.; Wilson, M.J.; Rossant, J.; et al. Retinoid signaling determines germ cell fate in mice. Science 2006, 312, 596–600. [Google Scholar] [CrossRef]
- Niederreither, K.; Fraulob, V.; Garnier, J.M.; Chambon, P.; Dolle, P. Differential expression of retinoic acid-synthesizing (RALDH) enzymes during fetal development and organ differentiation in the mouse. Mech. Dev. 2002, 110, 165–171. [Google Scholar] [CrossRef]
- Koubova, J.; Menke, D.B.; Zhou, Q.; Capel, B.; Griswold, M.D.; Page, D.C. Retinoic acid regulates sex-specific timing of meiotic initiation in mice. Proc. Natl. Acad. Sci. USA 2006, 103, 2474–2479. [Google Scholar] [CrossRef]
- Childs, A.J.; Cowan, G.; Kinnell, H.L.; Anderson, R.A.; Saunders, P.T. Retinoic Acid signalling and the control of meiotic entry in the human fetal gonad. PLoS ONE 2011, 6, e20249. [Google Scholar] [CrossRef] [PubMed]
- Jorgensen, A.; Rajpert-De Meyts, E. Regulation of meiotic entry and gonadal sex differentiation in the human: Normal and disrupted signaling. Biomol. Concepts 2014, 5, 331–341. [Google Scholar] [CrossRef]
- Stillwell, W.; Nahmias, S. Effect of retinol and retinoic acid on P/O ratios of coupled mitochondria. Biochem. Int. 1983, 6, 385–392. [Google Scholar]
- Kambhampati, S.; Verma, A.; Li, Y.; Parmar, S.; Sassano, A.; Platanias, L.C. Signalling pathways activated by all-trans-retinoic acid in acute promyelocytic leukemia cells. Leuk. Lymphoma 2004, 45, 2175–2185. [Google Scholar] [CrossRef] [PubMed]
- Kuenzli, S.; Saurat, J.H. Retinoids for the treatment of psoriasis: Outlook for the future. Curr. Opin. Investig. Drugs 2001, 2, 625–630. [Google Scholar]
- Aggarwal, S.; Kim, S.W.; Cheon, K.; Tabassam, F.H.; Yoon, J.H.; Koo, J.S. Nonclassical action of retinoic acid on the activation of the cAMP response element-binding protein in normal human bronchial epithelial cells. Mol. Biol. Cell. 2006, 17, 566–575. [Google Scholar] [CrossRef]
- Lerner, A.J.; Gustaw-Rothenberg, K.; Smyth, S.; Casadesus, G. Retinoids for treatment of Alzheimer’s disease. Biofactors 2012, 38, 84–89. [Google Scholar] [CrossRef] [PubMed]
- Acin-Perez, R.; Hoyos, B.; Zhao, F.; Vinogradov, V.; Fischman, D.A.; Harris, R.A.; Leitges, M.; Wongsiriroj, N.; Blaner, W.S.; Manfredi, G.; et al. Control of oxidative phosphorylation by vitamin A illuminates a fundamental role in mitochondrial energy homoeostasis. FASEB J. 2010, 24, 627–636. [Google Scholar] [CrossRef]
- Hoyos, B.; Imam, A.; Korichneva, I.; Levi, E.; Chua, R.; Hammerling, U. Activation of c-Raf kinase by ultraviolet light. Regulation by retinoids. J. Biol. Chem. 2002, 277, 23949–23957. [Google Scholar] [CrossRef]
- Hurley, J.H.; Newton, A.C.; Parker, P.J.; Blumberg, P.M.; Nishizuka, Y. Taxonomy and function of C1 protein kinase C homology domains. Protein Sci. 1997, 6, 477–480. [Google Scholar] [CrossRef]
- Imam, A.; Hoyos, B.; Swenson, C.; Levi, E.; Chua, R.; Viriya, E.; Hammerling, U. Retinoids as ligands and coactivators of protein kinase C alpha. FASEB J. 2001, 15, 28–30. [Google Scholar] [CrossRef]
- Ochoa, W.F.; Torrecillas, A.; Fita, I.; Verdaguer, N.; Corbalan-Garcia, S.; Gomez-Fernandez, J.C. Retinoic acid binds to the C2-domain of protein kinase C(alpha). Biochemistry 2003, 42, 8774–8779. [Google Scholar] [CrossRef]
- Tapiero, H.; Townsend, D.M.; Tew, K.D. The role of carotenoids in the prevention of human pathologies. Biomed. Pharm. 2004, 58, 100–110. [Google Scholar] [CrossRef]
- Palace, V.P.; Khaper, N.; Qin, Q.; Singal, P.K. Antioxidant potentials of vitamin A and carotenoids and their relevance to heart disease. Free Radic. Biol. Med. 1999, 26, 746–761. [Google Scholar] [CrossRef]
- Ribeiro, D.; Freitas, M.; Silva, A.M.S.; Carvalho, F.; Fernandes, E. Antioxidant and pro-oxidant activities of carotenoids and their oxidation products. Food Chem. Toxicol. 2018, 120, 681–699. [Google Scholar] [CrossRef]
- Dao, D.Q.; Ngo, T.C.; Thong, N.M.; Nam, P.C. Is Vitamin A an Antioxidant or a Pro-oxidant? J. Phys. Chem. B 2017, 121, 9348–9357. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Dou, H.L.; Wu, Y.Q.; Huang, Y.M.; Huang, Y.B.; Xu, X.R.; Zou, Z.Y.; Lin, X.M. Lutein and zeaxanthin intake and the risk of age-related macular degeneration: A systematic review and meta-analysis. Br. J. Nutr. 2012, 107, 350–359. [Google Scholar] [CrossRef]
- Christoforidis, J.B.; Tecce, N.; Dell’Omo, R.; Mastropasqua, R.; Verolino, M.; Costagliola, C. Age related macular degeneration and visual disability. Curr. Drug Targets 2011, 12, 221–233. [Google Scholar] [CrossRef] [PubMed]
- Murillo, A.G.; Fernandez, M.L. Potential of Dietary Non-Provitamin A Carotenoids in the Prevention and Treatment of Diabetic Microvascular Complications. Adv. Nutr. 2016, 7, 14–24. [Google Scholar] [CrossRef]
- Brazionis, L.; Rowley, K.; Jenkins, A.; Itsiopoulos, C.; O’Dea, K. Plasminogen activator inhibitor-1 activity in type 2 diabetes: A different relationship with coronary heart disease and diabetic retinopathy. Arter. Thromb. Vasc. Biol. 2008, 28, 786–791. [Google Scholar] [CrossRef]
- Nongenomic interactions of retinoic acid and thyroid hormone at the plasma membrane. Nutr. Rev. 1989, 47, 213–215. [CrossRef]
- Denisenko-Nehrbass, N.I.; Mello, C.V. Molecular targets of disulfiram action on song maturation in zebra finches. Brain Res. Mol. Brain Res. 2001, 87, 246–250. [Google Scholar] [CrossRef]
- Tournier, S.; Raynaud, F.; Gerbaud, P.; Lohmann, S.M.; Anderson, W.B.; Evain-Brion, D. Retinoylation of the type II cAMP-binding regulatory subunit of cAMP-dependent protein kinase is increased in psoriatic human fibroblasts. J. Cell Physiol. 1996, 167, 196–203. [Google Scholar] [CrossRef]
- Genchi, G.; Olson, J.A. Retinoylation of proteins in cell-free fractions of rat tissues in vitro. Biochim. Biophys. Acta 2001, 1530, 146–154. [Google Scholar] [CrossRef]
- Tanumihardjo, S.A. Vitamin A and bone health: The balancing act. J. Clin. Densitom. 2013, 16, 414–419. [Google Scholar] [CrossRef] [PubMed]
- de Jonge, E.A.; Kiefte-de Jong, J.C.; Campos-Obando, N.; Booij, L.; Franco, O.H.; Hofman, A.; Uitterlinden, A.G.; Rivadeneira, F.; Zillikens, M.C. Dietary vitamin A intake and bone health in the elderly: The Rotterdam Study. Eur. J. Clin. Nutr. 2015, 69, 1375. [Google Scholar] [CrossRef] [PubMed]
- Johansson, S.; Lind, P.M.; Hakansson, H.; Oxlund, H.; Orberg, J.; Melhus, H. Subclinical hypervitaminosis A causes fragile bones in rats. Bone 2002, 31, 685–689. [Google Scholar] [CrossRef]
- Kneissel, M.; Studer, A.; Cortesi, R.; Susa, M. Retinoid-induced bone thinning is caused by subperiosteal osteoclast activity in adult rodents. Bone 2005, 36, 202–214. [Google Scholar] [CrossRef]
- Wu, A.M.; Huang, C.Q.; Lin, Z.K.; Tian, N.F.; Ni, W.F.; Wang, X.Y.; Xu, H.Z.; Chi, Y.L. The relationship between vitamin A and risk of fracture: Meta-analysis of prospective studies. J. Bone Miner. Res. 2014, 29, 2032–2039. [Google Scholar] [CrossRef]
- Goncalves, A.; Roi, S.; Nowicki, M.; Dhaussy, A.; Huertas, A.; Amiot, M.J.; Reboul, E. Fat-soluble vitamin intestinal absorption: Absorption sites in the intestine and interactions for absorption. Food Chem. 2015, 172, 155–160. [Google Scholar] [CrossRef]
- Caire-Juvera, G.; Ritenbaugh, C.; Wactawski-Wende, J.; Snetselaar, L.G.; Chen, Z. Vitamin A and retinol intakes and the risk of fractures among participants of the Women’s Health Initiative Observational Study. Am. J. Clin. Nutr. 2009, 89, 323–330. [Google Scholar] [CrossRef]
- Mata-Granados, J.M.; Cuenca-Acevedo, R.; Luque de Castro, M.D.; Sosa, M.; Quesada-Gomez, J.M. Vitamin D deficiency and high serum levels of vitamin A increase the risk of osteoporosis evaluated by Quantitative Ultrasound Measurements (QUS) in postmenopausal Spanish women. Clin. Biochem. 2010, 43, 1064–1068. [Google Scholar] [CrossRef] [PubMed]
- Joo, N.S.; Yang, S.W.; Song, B.C.; Yeum, K.J. Vitamin A intake, serum vitamin D and bone mineral density: Analysis of the Korea National Health and Nutrition Examination Survey (KNHANES, 2008–2011). Nutrients 2015, 7, 1716–1727. [Google Scholar] [CrossRef] [PubMed]
- Johansson, S.; Melhus, H. Vitamin A antagonizes calcium response to vitamin D in man. J. Bone Miner. Res. 2001, 16, 1899–1905. [Google Scholar] [CrossRef]
- Rohde, C.M.; DeLuca, H.F. All-trans retinoic acid antagonizes the action of calciferol and its active metabolite, 1,25-dihydroxycholecalciferol, in rats. J. Nutr. 2005, 135, 1647–1652. [Google Scholar] [CrossRef][Green Version]
- Rohde, C.M.; Manatt, M.; Clagett-Dame, M.; DeLuca, H.F. Vitamin A antagonizes the action of vitamin D in rats. J. Nutr. 1999, 129, 2246–2250. [Google Scholar] [CrossRef] [PubMed]
- Sha, J.; Pan, J.; Ping, P.; Xuan, H.; Li, D.; Bo, J.; Liu, D.; Huang, Y. Synergistic effect and mechanism of vitamin A and vitamin D on inducing apoptosis of prostate cancer cells. Mol. Biol. Rep. 2013, 40, 2763–2768. [Google Scholar] [CrossRef]
- Cheng, T.Y.; Goodman, G.E.; Thornquist, M.D.; Barnett, M.J.; Beresford, S.A.; LaCroix, A.Z.; Zheng, Y.; Neuhouser, M.L. Estimated intake of vitamin D and its interaction with vitamin A on lung cancer risk among smokers. Int. J. Cancer 2014, 135, 2135–2145. [Google Scholar] [CrossRef]
- Ferreiro-Vera, C.; Mata-Granados, J.M.; Quesada Gomez, J.M.; Luque de Castro, M.D. On-line coupling of automatic solid-phase extraction and HPLC for determination of carotenoids in serum. Talanta 2011, 85, 1842–1847. [Google Scholar] [CrossRef]
- Boulet, L.; Alex, B.; Clavey, N.; Martinez, J.; Ducros, V. Simultaneous analysis of retinol, six carotenoids, two tocopherols, and coenzyme Q10 from human plasma by HPLC. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2020, 1151, 122158. [Google Scholar] [CrossRef]
- Lazzarino, G.; Longo, S.; Amorini, A.M.; Di Pietro, V.; D’Urso, S.; Lazzarino, G.; Belli, A.; Tavazzi, B. Single-step preparation of selected biological fluids for the high performance liquid chromatographic analysis of fat-soluble vitamins and antioxidants. J. Chromatogr. A 2017, 1527, 43–52. [Google Scholar] [CrossRef]
- Hartmann, S.; Froescheis, O.; Ringenbach, F.; Wyss, R.; Bucheli, F.; Bischof, S.; Bausch, J.; Wiegand, U.W. Determination of retinol and retinyl esters in human plasma by high-performance liquid chromatography with automated column switching and ultraviolet detection. J. Chromatogr. B Biomed. Sci. Appl. 2001, 751, 265–275. [Google Scholar] [CrossRef]
- Ruhl, R.; Schweigert, F.J. Automated solid-phase extraction and liquid chromatographic method for retinoid determination in biological samples. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2003, 798, 309–316. [Google Scholar] [CrossRef] [PubMed]
- Mata-Granados, J.M.; Luque De Castro, M.D.; Quesada, J.M. Fully automated method for the determination of 24,25(OH)2 and 25(OH) D3 hydroxyvitamins, and vitamins A and E in human serum by HPLC. J. Pharm. Biomed. Anal. 2004, 35, 575–582. [Google Scholar] [CrossRef]
- Xuan, R.; Wang, T.; Hou, C.; Li, X.; Li, Y.; Chen, Y.; Gao, Y.; Qiu, D.; Xiao, X.; Zhang, L.; et al. Determination of vitamin A in blood serum based on solid-phase extraction using cetyltrimethyl ammonium bromide-modified attapulgite. J. Sep. Sci. 2019, 42, 3521–3527. [Google Scholar] [CrossRef] [PubMed]
- Pan, Q.; Shen, M.; Yu, T.; Yang, X.; Li, Q.; Zhao, B.; Zou, J.; Zhang, M. Liquid chromatography as candidate reference method for the determination of vitamins A and E in human serum. J. Clin. Lab. Anal. 2020, 34, e23528. [Google Scholar] [CrossRef] [PubMed]
- Erhardt, J.G.; Craft, N.E.; Heinrich, F.; Biesalski, H.K. Rapid and simple measurement of retinol in human dried whole blood spots. J. Nutr. 2002, 132, 318–321. [Google Scholar] [CrossRef] [PubMed]
- Plisek, J.; Kasparova, M.; Solichova, D.; Krcmova, L.; Kucerova, B.; Sobotka, L.; Solich, P. Application of core-shell technology for determination of retinol and alpha-tocopherol in breast milk. Talanta 2013, 107, 382–388. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, K.; Kiko, T.; Hatade, K.; Asai, A.; Kimura, F.; Sookwong, P.; Tsuduki, T.; Arai, H.; Miyazawa, T. Development of a high-performance liquid chromatography-based assay for carotenoids in human red blood cells: Application to clinical studies. Anal. Biochem. 2008, 381, 129–134. [Google Scholar] [CrossRef]
- Thibeault, D.; Su, H.; MacNamara, E.; Schipper, H.M. Isocratic rapid liquid chromatographic method for simultaneous determination of carotenoids, retinol, and tocopherols in human serum. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2009, 877, 1077–1083. [Google Scholar] [CrossRef]
- Hsu, B.Y.; Pu, Y.S.; Inbaraj, B.S.; Chen, B.H. An improved high performance liquid chromatography-diode array detection-mass spectrometry method for determination of carotenoids and their precursors phytoene and phytofluene in human serum. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2012, 899, 36–45. [Google Scholar] [CrossRef]
- Bell, E.C.; John, M.; Hughes, R.J.; Pham, T. Ultra-performance liquid chromatographic determination of tocopherols and retinol in human plasma. J. Chromatogr. Sci. 2014, 52, 1065–1070. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Yuan, C.; Burgyan, M.; Bunch, D.R.; Reineks, E.; Jackson, R.; Steinle, R.; Wang, S. Fast, simple, and sensitive high-performance liquid chromatography method for measuring vitamins A and E in human blood plasma. J. Sep. Sci. 2014, 37, 2293–2299. [Google Scholar] [CrossRef]
- Kandar, R.; Drabkova, P.; Myslikova, K.; Hampl, R. Determination of retinol and alpha-tocopherol in human seminal plasma using an HPLC with UV detection. Andrologia 2014, 46, 472–478. [Google Scholar] [CrossRef] [PubMed]
- Gleize, B.; Steib, M.; Andre, M.; Reboul, E. Simple and fast HPLC method for simultaneous determination of retinol, tocopherols, coenzyme Q(10) and carotenoids in complex samples. Food Chem. 2012, 134, 2560–2564. [Google Scholar] [CrossRef]
- Liu, Z.E.A. Solid phase extraction with electrospun nanofibers for determination of retinol and α-tocopherol in plasma. Micro. Chim. Acta 2009, 2010, 59–64. [Google Scholar] [CrossRef] [PubMed]
- Kucerova, B.; Krcmova, L.; Solichova, D.; Plisek, J.; Solich, P. Comparison of a new high-resolution monolithic column with core-shell and fully porous columns for the analysis of retinol and alpha-tocopherol in human serum and breast milk by ultra-high-performance liquid chromatography. J. Sep. Sci. 2013, 36, 2223–2230. [Google Scholar] [CrossRef]
- Zhang, H.E.A. Simultaneous determination of Vitamin A, 25-hydroxyl vitamin D 3 α-tocopherol in small biological fluids by liquid chromatography-tandem mass spectrometry. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2018, 15, 1–8. [Google Scholar] [CrossRef]
- Le, J.; Yuan, T.F.; Zhang, Y.; Wang, S.T.; Li, Y. New LC-MS/MS method with single-step pretreatment analyzes fat-soluble vitamins in plasma and amniotic fluid. J. Lipid Res. 2018, 59, 1783–1790. [Google Scholar] [CrossRef] [PubMed]
- Peersman, N.; Elslande, J.V.; Lepage, Y.; De Amicis, S.; Desmet, K.; Vermeersch, P. UPLC-MS/MS method for determination of retinol and alpha-tocopherol in serum using a simple sample pretreatment and UniSpray as ionization technique to reduce matrix effects. Clin. Chem. Lab. Med. 2020, 58, 769–779. [Google Scholar] [CrossRef]
- Arnold, S.L.; Amory, J.K.; Walsh, T.J.; Isoherranen, N. A sensitive and specific method for measurement of multiple retinoids in human serum with UHPLC-MS/MS. J. Lipid Res. 2012, 53, 587–598. [Google Scholar] [CrossRef]
- Khaksari, M.; Mazzoleni, L.R.; Ruan, C.; Kennedy, R.T.; Minerick, A.R. Determination of water-soluble and fat-soluble vitamins in tears and blood serum of infants and parents by liquid chromatography/mass spectrometry. Exp. Eye Res. 2017, 155, 54–63. [Google Scholar] [CrossRef] [PubMed]
- Kucerova, K.e.a. Determination of urinary retinol and creatinine as an early sensitive marker of renal dysfunction. J. Chromatogr. A 2019, 1607. [Google Scholar] [CrossRef] [PubMed]
- Zoccali, M.; Giuffrida, D.; Granese, R.; Salafia, F.; Dugo, P.; Mondello, L. Determination of free apocarotenoids and apocarotenoid esters in human colostrum. Anal. Bioanal. Chem. 2020, 412, 1335–1342. [Google Scholar] [CrossRef] [PubMed]
- Zoccali, M.; Giuffrida, D.; Salafia, F.; Giofre, S.V.; Mondello, L. Carotenoids and apocarotenoids determination in intact human blood samples by online supercritical fluid extraction-supercritical fluid chromatography-tandem mass spectrometry. Anal. Chim. Acta 2018, 1032, 40–47. [Google Scholar] [CrossRef]
- Petruzziello, F.; Grand-Guillaume Perrenoud, A.; Thorimbert, A.; Fogwill, M.; Rezzi, S. Quantitative Profiling of Endogenous Fat-Soluble Vitamins and Carotenoids in Human Plasma Using an Improved UHPSFC-ESI-MS Interface. Anal. Chem. 2017, 89, 7615–7622. [Google Scholar] [CrossRef] [PubMed]
- Matsubara, A.; Uchikata, T.; Shinohara, M.; Nishiumi, S.; Yoshida, M.; Fukusaki, E.; Bamba, T. Highly sensitive and rapid profiling method for carotenoids and their epoxidized products using supercritical fluid chromatography coupled with electrospray ionization-triple quadrupole mass spectrometry. J. Biosci. Bioeng. 2012, 113, 782–787. [Google Scholar] [CrossRef]
- Wang, L.H.e.a. Determination of retinoids in human serum, tocopherol and retinyl acetate in pharmaceuticals by RP-LC with electrochemical detection. J. Pharm. Biomed. Anal. 2001, 25, 785–793. [Google Scholar] [CrossRef]
- Hermans, N.; Cos, P.; Berghe, D.V.; Vlietinck, A.J.; de Bruyne, T. Method development and validation for monitoring in vivo oxidative stress: Evaluation of lipid peroxidation and fat-soluble vitamin status by HPLC in rat plasma. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2005, 822, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Ferruzzi, M.G.; Sander, L.C.; Rock, C.L.; Schwartz, S.J. Carotenoid determination in biological microsamples using liquid chromatography with a coulometric electrochemical array detector. Anal. Biochem. 1998, 256, 74–81. [Google Scholar] [CrossRef]
- My BioSource. Human Vitamin A Elisa Kit. Available online: https://cdn.mybiosource.com/tds/protocol_manuals/000000-799999/MBS729269.pdf (accessed on 27 January 2021).
- Abbexa. Vitamin A Elisa Kit. Available online: https://www.abbexa.com/vitamin-a-elisa-kit (accessed on 27 January 2021).
- Aviva Systems Biology. Vitamin A Elisa Kit. Available online: https://www.avivasysbio.com/pub/media/pdf/products/OKEH02573.pdf (accessed on 27 January 2021).
- Kamiya Biomedical Company. Human Vitamin A Elisa K-Assay. Available online: https://www.kamiyabiomedical.com/pdf/KT-31968.pdf (accessed on 27 January 2021).
- LifeSpan BioSciences. Human Vitamin A Elisa Kit. Available online: https://www.lsbio.com/elisakits/manualpdf/ls-f10051.pdf (accessed on 27 January 2021).
- Novus Biologicals. Human Vitamin A Elisa Kit. Available online: https://resources.novusbio.com/manual/Manual-NBP2-60191-53627008.pdf (accessed on 27 January 2021).
- Cusabio. Human Vitamin A Elisa Kit. Available online: https://cdn.mybiosource.com/tds/protocol_manuals/000000-799999/MBS729269.pdf (accessed on 27 January 2021).
- Laboratory, B.T. Human Beta-Caroten Elisa Kit. Available online: http://www.bt-laboratory.com/product/human-beta-carotene-elisa-kit-2/ (accessed on 27 January 2021).
- BioAnalyt. iCheck Karotene. Available online: https://www.bioanalyt.com/wp-content/uploads/2019/05/iCheck-Carotene_product-information_EN_2019.pdf (accessed on 27 January 2021).
- Recipe. UHPLC Complete Kit Vitamin A and E In Serum/Plasma. Available online: https://recipe.de/products/vitamin-a-e-serum-uhplc/ (accessed on 27 January 2021).
- Recipe. HPLC Complete Kit Vitamin AE In Plasma/Serum. Available online: https://recipe.de/products/vitamin-a-e-serum/ (accessed on 27 January 2021).
- Recipe. HPLC Complete Kit Beta-Carotene in Plasma/Serum. Available online: https://recipe.de/products/%ce%b2-carotene-serum/ (accessed on 27 January 2021).
- Immundiagnostic AG. Vitamin A/E HPLC Kit. Available online: http://www.immundiagnostik.com/en/home/products/kits-assays/hplc-applications.html?tx_mokom01immunprodukte_pi1%5Ban%5D=KC1600&tx_mokom01immunprodukte_pi1%5Bag%5D=409&cHash=639cc97481 (accessed on 27 January 2021).
- Chromsystems. Vitamin A and E in Serum/Plasma. Available online: https://chromsystems.com/en/vitamins-a-and-e-in-serum-plasma-hplc-34000.html (accessed on 27 January 2021).
- Trumbo, P.; Yates, A.A.; Schlicker, S.; Poos, M. Dietary reference intakes: Vitamin A, vitamin K, arsenic, boron, chromium, copper, iodine, iron, manganese, molybdenum, nickel, silicon, vanadium, and zinc. J. Am. Diet. Assoc. 2001, 101, 294–301. [Google Scholar] [CrossRef]
- Gannon, B.M.; Jones, C.; Mehta, S. Vitamin A Requirements in Pregnancy and Lactation. Curr. Dev. Nutr. 2020, 4, nzaa142. [Google Scholar] [CrossRef] [PubMed]
- Bernard, R.A.; Halpern, B.P. Taste changes in vitamin A deficiency. J. Gen. Physiol. 1968, 52, 444–464. [Google Scholar] [CrossRef]
- Biesalski, H.K.; Wellner, U.; Stofft, E.; Bassler, K.H. Vitamin A deficiency and sensory function. Acta Vitam. Enzym. 1985, 7, 45–54. [Google Scholar]
- Bromley, S.M. Smell and taste disorders: A primary care approach. Am. Fam. Physician. 2000, 61, 427–438. [Google Scholar] [PubMed]
- Reifen, R.; Zaiger, G.; Uni, Z. Effect of vitamin A on small intestinal brush border enzymes in a rat. Int. J. Vitam. Nutr. Res. 1998, 68, 281–286. [Google Scholar]
- Wiedermann, U.; Chen, X.J.; Enerback, L.; Hanson, L.A.; Kahu, H.; Dahlgren, U.I. Vitamin A deficiency increases inflammatory responses. Scand J. Immunol. 1996, 44, 578–584. [Google Scholar] [CrossRef]
- Sirisinha, S. The pleiotropic role of vitamin A in regulating mucosal immunity. Asian Pac. J. Allergy. Immunol. 2015, 33, 71–89. [Google Scholar]
- Goetghebuer, T.; Brasseur, D.; Dramaix, M.; DeMol, P.; Donnen, P.; Bahwere, P.; Duchateau, J.; Hennart, P. Significance of very low retinol levels during severe protein-energy malnutrition. J. Trop. Pediatr. 1996, 42, 158–161. [Google Scholar] [CrossRef] [PubMed]
- West, K.P., Jr. Vitamin A deficiency disorders in children and women. Food Nutr. Bull. 2003, 24, S78–S90. [Google Scholar] [CrossRef]
- Huiming, Y.; Chaomin, W.; Meng, M. Vitamin A for treating measles in children. Cochrane Database Syst. Rev. 2005. [Google Scholar] [CrossRef]
- Stevens, G.A.; Bennett, J.E.; Hennocq, Q.; Lu, Y.; De-Regil, L.M.; Rogers, L.; Danaei, G.; Li, G.; White, R.A.; Flaxman, S.R.; et al. Trends and mortality effects of vitamin A deficiency in children in 138 low-income and middle-income countries between 1991 and 2013: A pooled analysis of population-based surveys. Lancet Glob. Health 2015, 3, e528–e536. [Google Scholar] [CrossRef]
- Bailey, R.L.; West, K.P., Jr.; Black, R.E. The epidemiology of global micronutrient deficiencies. Ann. Nutr. Metab. 2015, 66 (Suppl. 2), 22–33. [Google Scholar] [CrossRef]
- De Sole, G.; Belay, Y.; Zegeye, B. Vitamin A deficiency in southern Ethiopia. Am. J. Clin. Nutr. 1987, 45, 780–784. [Google Scholar] [CrossRef]
- Young, J.K.; Giesbrecht, H.E.; Eskin, M.N.; Aliani, M.; Suh, M. Nutrition implications for fetal alcohol spectrum disorder. Adv. Nutr. 2014, 5, 675–692. [Google Scholar] [CrossRef] [PubMed]
- Garg, M.; Sharma, N.; Sharma, S.; Kapoor, P.; Kumar, A.; Chunduri, V.; Arora, P. Biofortified Crops Generated by Breeding, Agronomy, and Transgenic Approaches Are Improving Lives of Millions of People around the World. Front. Nutr. 2018, 5. [Google Scholar] [CrossRef] [PubMed]
- Tallman, M.S.; Andersen, J.W.; Schiffer, C.A.; Appelbaum, F.R.; Feusner, J.H.; Ogden, A.; Shepherd, L.; Willman, C.; Bloomfield, C.D.; Rowe, J.M.; et al. All-trans-retinoic acid in acute promyelocytic leukemia. N. Engl. J. Med. 1997, 337, 1021–1028. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.C.; Hsu, S.L.; Lin, H.; Yang, T.Y. Retinoic acid and cancer treatment. Biomedicine 2014, 4, 22. [Google Scholar] [CrossRef] [PubMed]
- Barclay, A.J.; Foster, A.; Sommer, A. Vitamin A supplements and mortality related to measles: A randomised clinical trial. Br. Med. J. 1987, 294, 294–296. [Google Scholar] [CrossRef] [PubMed]
- Hussey, G.D.; Klein, M. A randomized, controlled trial of vitamin A in children with severe measles. N. Engl. J. Med. 1990, 323, 160–164. [Google Scholar] [CrossRef]
- Bello, S.; Meremikwu, M.M.; Ejemot-Nwadiaro, R.I.; Oduwole, O. Routine vitamin A supplementation for the prevention of blindness due to measles infection in children. Cochrane Database Syst. Rev. 2011. [Google Scholar] [CrossRef]
- Barbul, A.; Thysen, B.; Rettura, G.; Levenson, S.M.; Seifter, E. White cell involvement in the inflammatory, wound healing, and immune actions of vitamin A. JPEN J. Parenter. Enter. Nutr. 1978, 2, 129–138. [Google Scholar] [CrossRef] [PubMed]
- Damery, E.; Solimando, D.A., Jr.; Waddell, J.A. Arsenic Trioxide and Tretinoin (AsO/ATRA) for Acute Promyelocytic Leukemia (APL). Hosp. Pharm. 2016, 51, 628–632. [Google Scholar] [CrossRef]
- Breitman, T.R.; Selonick, S.E.; Collins, S.J. Induction of differentiation of the human promyelocytic leukemia cell line (HL-60) by retinoic acid. Proc. Natl. Acad. Sci. USA 1980, 77, 2936–2940. [Google Scholar] [CrossRef] [PubMed]
- Freemantle, S.J.; Spinella, M.J.; Dmitrovsky, E. Retinoids in cancer therapy and chemoprevention: Promise meets resistance. Oncogene 2003, 22, 7305–7315. [Google Scholar] [CrossRef] [PubMed]
- Duvic, M.; Friedman-Kien, A.E.; Looney, D.J.; Miles, S.A.; Myskowski, P.L.; Scadden, D.T.; Von Roenn, J.; Galpin, J.E.; Groopman, J.; Loewen, G.; et al. Topical treatment of cutaneous lesions of acquired immunodeficiency syndrome-related Kaposi sarcoma using alitretinoin gel: Results of phase 1 and 2 trials. Arch. Dermatol. 2000, 136, 1461–1469. [Google Scholar] [CrossRef]
- Rathi, S.K. Acne vulgaris treatment: The current scenario. Indian J. Dermatol. 2011, 56, 7–13. [Google Scholar] [CrossRef]
- Mukherjee, S.; Date, A.; Patravale, V.; Korting, H.C.; Roeder, A.; Weindl, G. Retinoids in the treatment of skin aging: An overview of clinical efficacy and safety. Clin. Interv. Aging 2006, 1, 327–348. [Google Scholar] [CrossRef]
- Karkeni, E.; Bonnet, L.; Astier, J.; Couturier, C.; Dalifard, J.; Tourniaire, F.; Landrier, J.F. All-trans-retinoic acid represses chemokine expression in adipocytes and adipose tissue by inhibiting NF-kappaB signaling. J. Nutr. Biochem. 2017, 42, 101–107. [Google Scholar] [CrossRef]
- Weiss, J.S.; Ellis, C.N.; Headington, J.T.; Tincoff, T.; Hamilton, T.A.; Voorhees, J.J. Topical tretinoin improves photoaged skin. A double-blind vehicle-controlled study. JAMA 1988, 259, 527–532. [Google Scholar] [CrossRef]
- Peck, G.L.; Olsen, T.G.; Yoder, F.W.; Strauss, J.S.; Downing, D.T.; Pandya, M.; Butkus, D.; Arnaud-Battandier, J. Prolonged remissions of cystic and conglobate acne with 13-cis-retinoic acid. N. Engl. J. Med. 1979, 300, 329–333. [Google Scholar] [CrossRef]
- Orfanos, C.E.; Zouboulis, C.C. Oral retinoids in the treatment of seborrhoea and acne. Dermatology 1998, 196, 140–147. [Google Scholar] [CrossRef]
- Layton, A. The use of isotretinoin in acne. Dermatoendocrinology 2009, 1, 162–169. [Google Scholar] [CrossRef] [PubMed]
- Booij, M.T.; Van De Kerkhof, P.C. Acitretin revisited in the era of biologics. J. Dermatolog. Treat. 2011, 22, 86–89. [Google Scholar] [CrossRef] [PubMed]
- Zasada, M.; Budzisz, E. Retinoids: Active molecules influencing skin structure formation in cosmetic and dermatological treatments. Postepy Dermatol. Alergol. 2019, 36, 392–397. [Google Scholar] [CrossRef] [PubMed]
- Murray, J.S.; White, J. Vitamin A supplementation in infants and children. J. Spec. Pediatr. Nurs. 2016, 21, 212–217. [Google Scholar] [CrossRef] [PubMed]
- Melendez-Martinez, A.J.; Stinco, C.M.; Mapelli-Brahm, P. Skin Carotenoids in Public Health and Nutricosmetics: The Emerging Roles and Applications of the UV Radiation-Absorbing Colourless Carotenoids Phytoene and Phytofluene. Nutrients 2019, 11, 1093. [Google Scholar] [CrossRef] [PubMed]
- Kopcke, W.; Krutmann, J. Protection from sunburn with beta-Carotene--a meta-analysis. Photochem. Photobiol. 2008, 84, 284–288. [Google Scholar] [CrossRef]
- Sies, H.; Stahl, W. Nutritional protection against skin damage from sunlight. Annu. Rev. Nutr. 2004, 24, 173–200. [Google Scholar] [CrossRef]
- Biesalski, H.K.; Hemmes, C.; Hopfenmuller, W.; Schmid, C.; Gollnick, H.P. Effects of controlled exposure of sunlight on plasma and skin levels of beta-carotene. Free Radic. Res. 1996, 24, 215–224. [Google Scholar] [CrossRef]
- Lee, J.; Jiang, S.; Levine, N.; Watson, R.R. Carotenoid supplementation reduces erythema in human skin after simulated solar radiation exposure. Proc. Soc. Exp. Biol. Med. 2000, 223, 170–174. [Google Scholar] [CrossRef]
- Ono, K.; Yamada, M. Vitamin A and Alzheimer’s disease. Geriatr. Gerontol. Int. 2012, 12, 180–188. [Google Scholar] [CrossRef] [PubMed]
- Moise, A.R.; Noy, N.; Palczewski, K.; Blaner, W.S. Delivery of retinoid-based therapies to target tissues. Biochemistry 2007, 46, 4449–4458. [Google Scholar] [CrossRef] [PubMed]
- Heinonen, M. Food groups as the source of retinoids, carotenoids, and vitamin A in Finland. Int. J. Vitam. Nutr. Res. 1991, 61, 3–9. [Google Scholar]
- Snodgrass, S.R. Vitamin neurotoxicity. Mol. Neurobiol. 1992, 6, 41–73. [Google Scholar] [CrossRef] [PubMed]
- Penniston, K.L.; Tanumihardjo, S.A. The acute and chronic toxic effects of vitamin A. Am. J. Clin. Nutr. 2006, 83, 191–201. [Google Scholar] [CrossRef] [PubMed]
- Lowe, N.J.; David, M. New retinoids for dermatologic diseases. Uses and toxicity. Dermatol. Clin. 1988, 6, 539–552. [Google Scholar] [CrossRef]
- Biesalski, H.K. Comparative assessment of the toxicology of vitamin A and retinoids in man. Toxicology 1989, 57, 117–161. [Google Scholar] [CrossRef]
- Patatanian, E.; Thompson, D.F. Retinoic acid syndrome: A review. J. Clin. Pharm. Ther. 2008, 33, 331–338. [Google Scholar] [CrossRef]
- Setty, O.H.; Misra, U.K. Effect of heparin on vitamin A induced hyperlipidemia in rats. Int. J. Vitam. Nutr. Res. 1981, 51, 325–330. [Google Scholar]
- Lammer, E.J.; Chen, D.T.; Hoar, R.M.; Agnish, N.D.; Benke, P.J.; Braun, J.T.; Curry, C.J.; Fernhoff, P.M.; Grix, A.W., Jr.; Lott, I.T.; et al. Retinoic acid embryopathy. N. Engl. J. Med. 1985, 313, 837–841. [Google Scholar] [CrossRef]
- Lascari, A.D. Carotenemia. A review. Clin. Pediatr. 1981, 20, 25–29. [Google Scholar] [CrossRef] [PubMed]







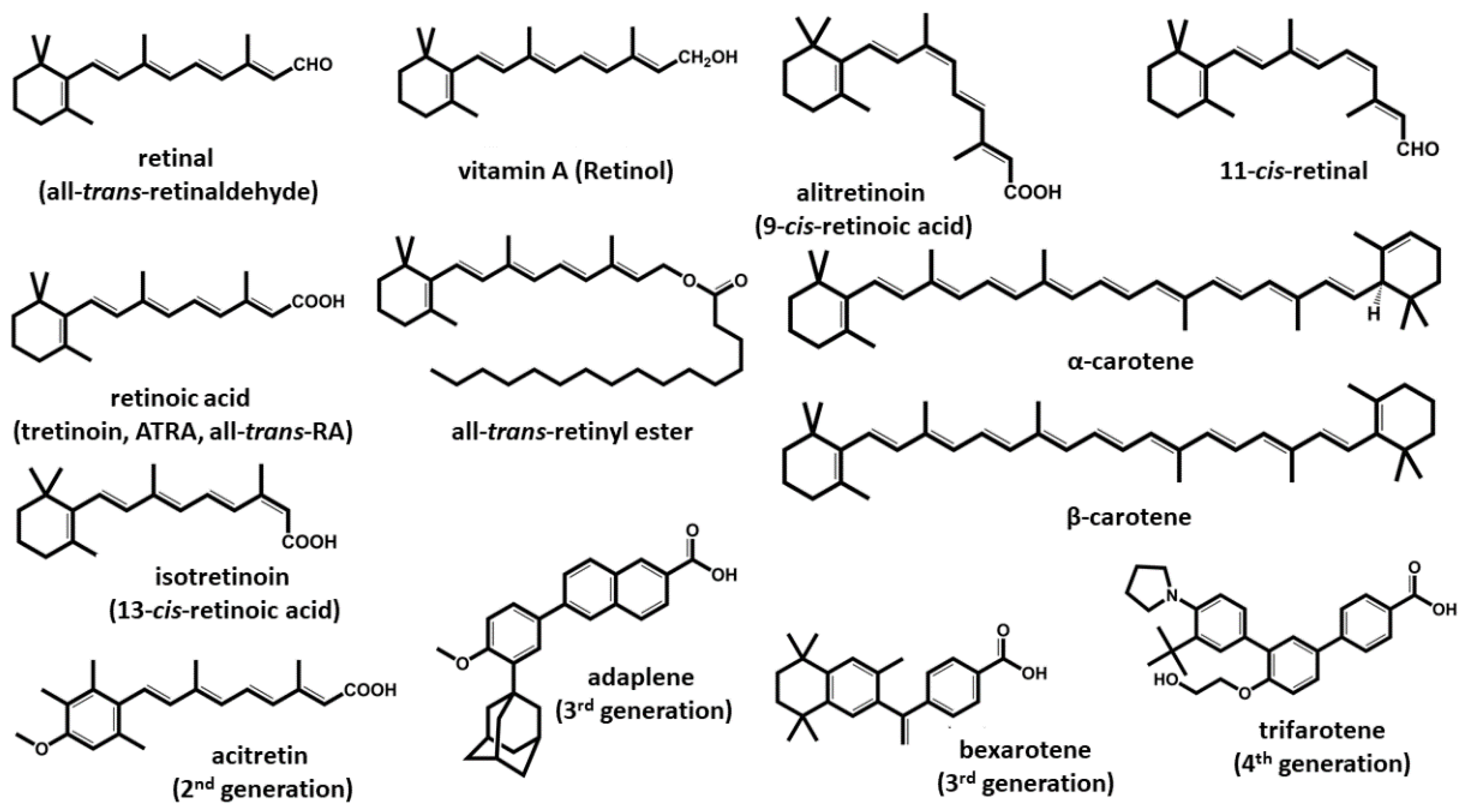{kind=link}
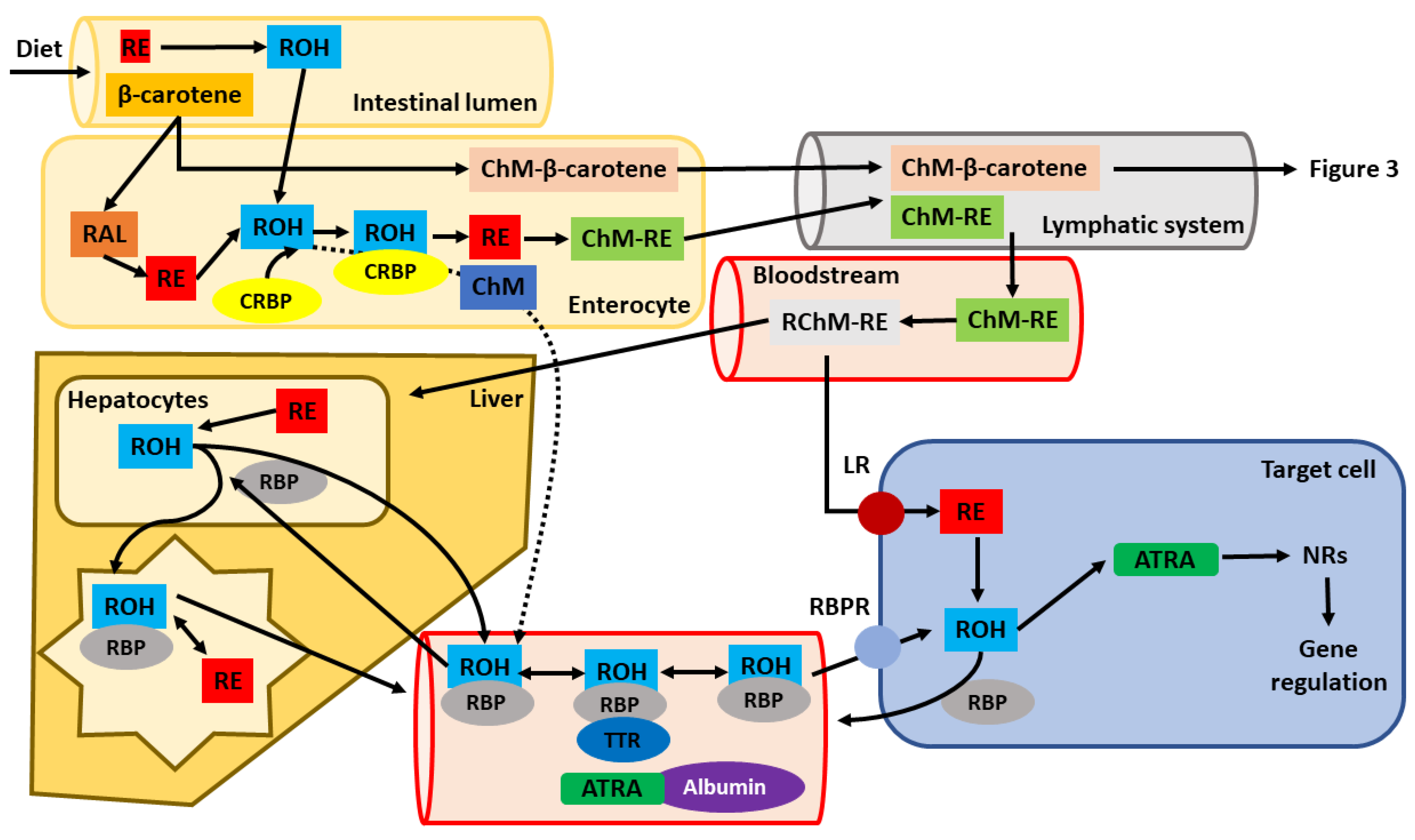{kind=link}
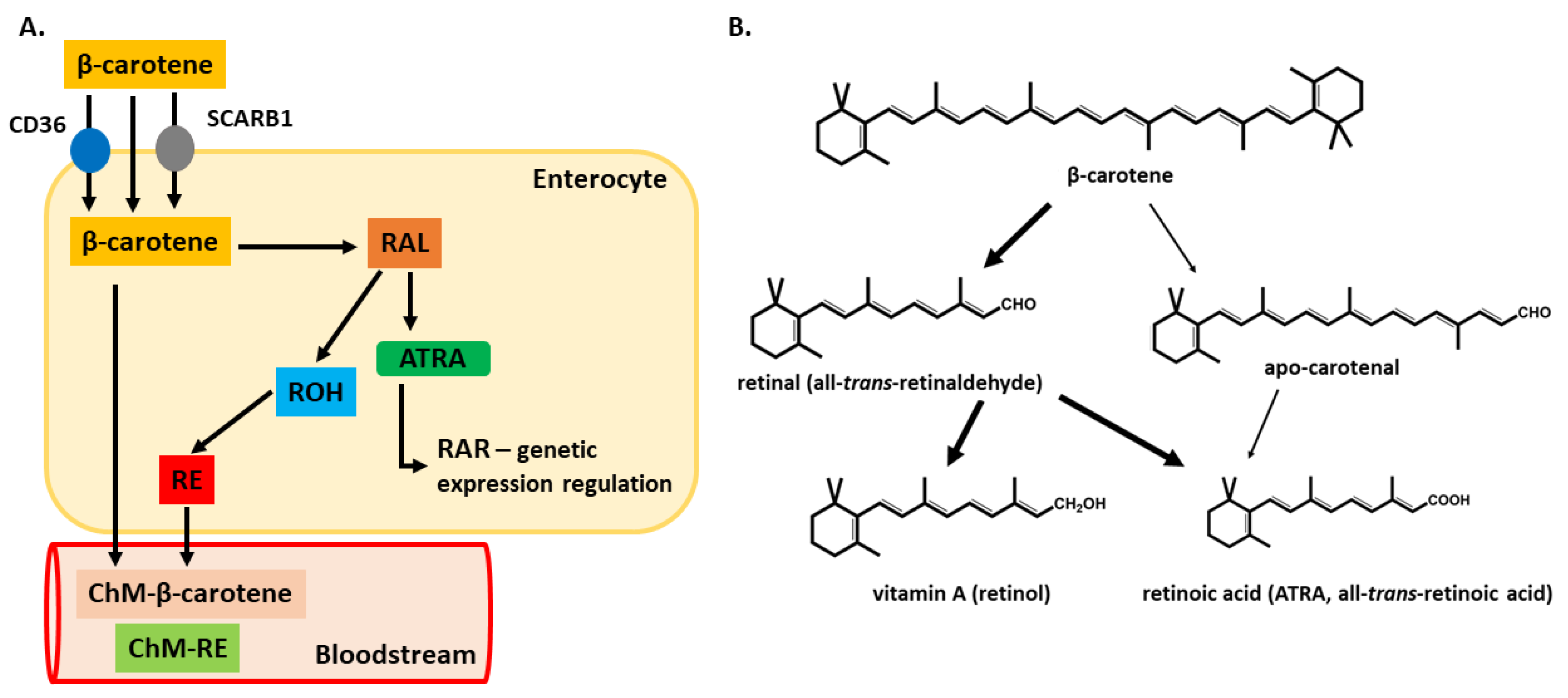{kind=link}
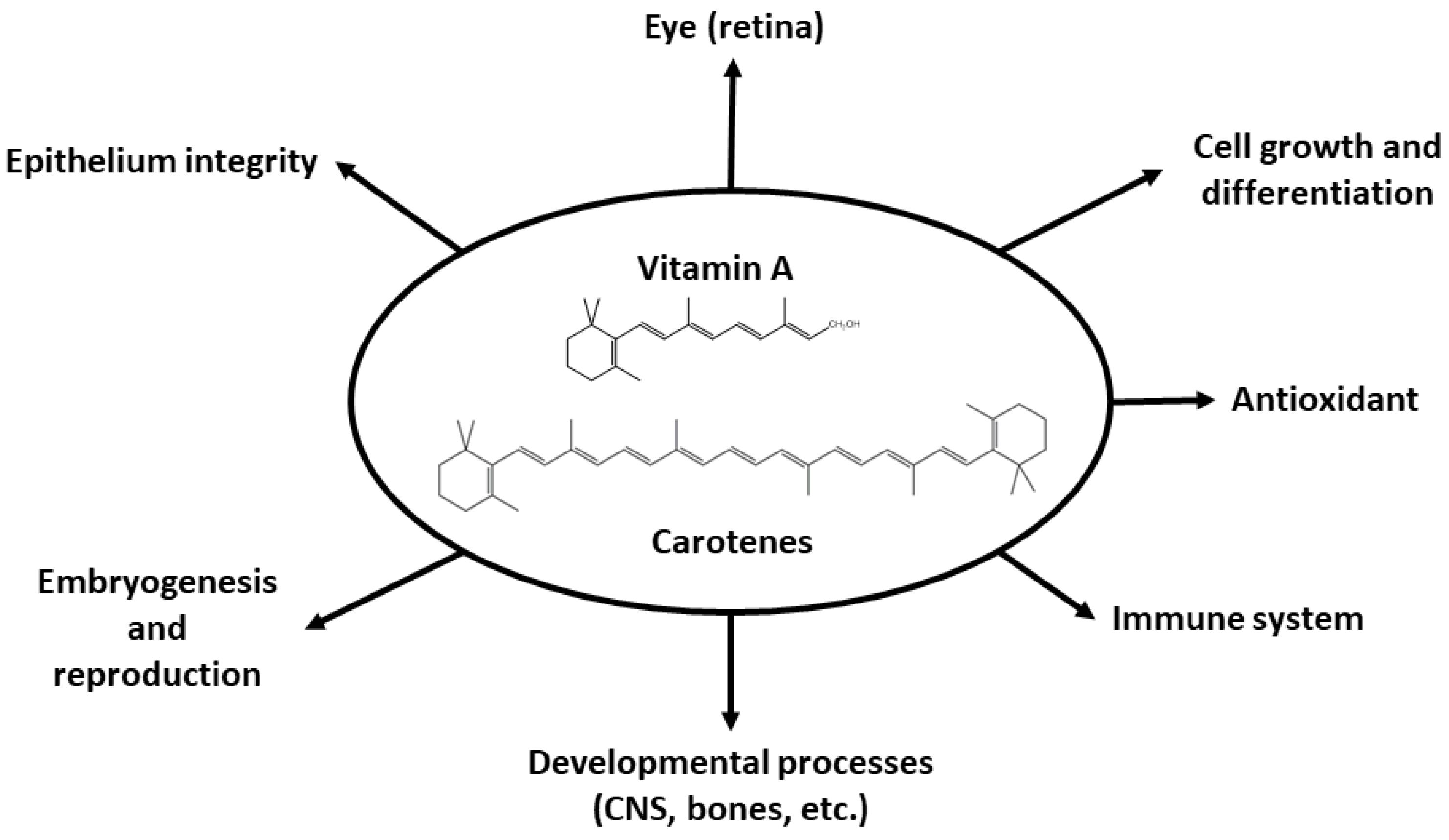{kind=link}
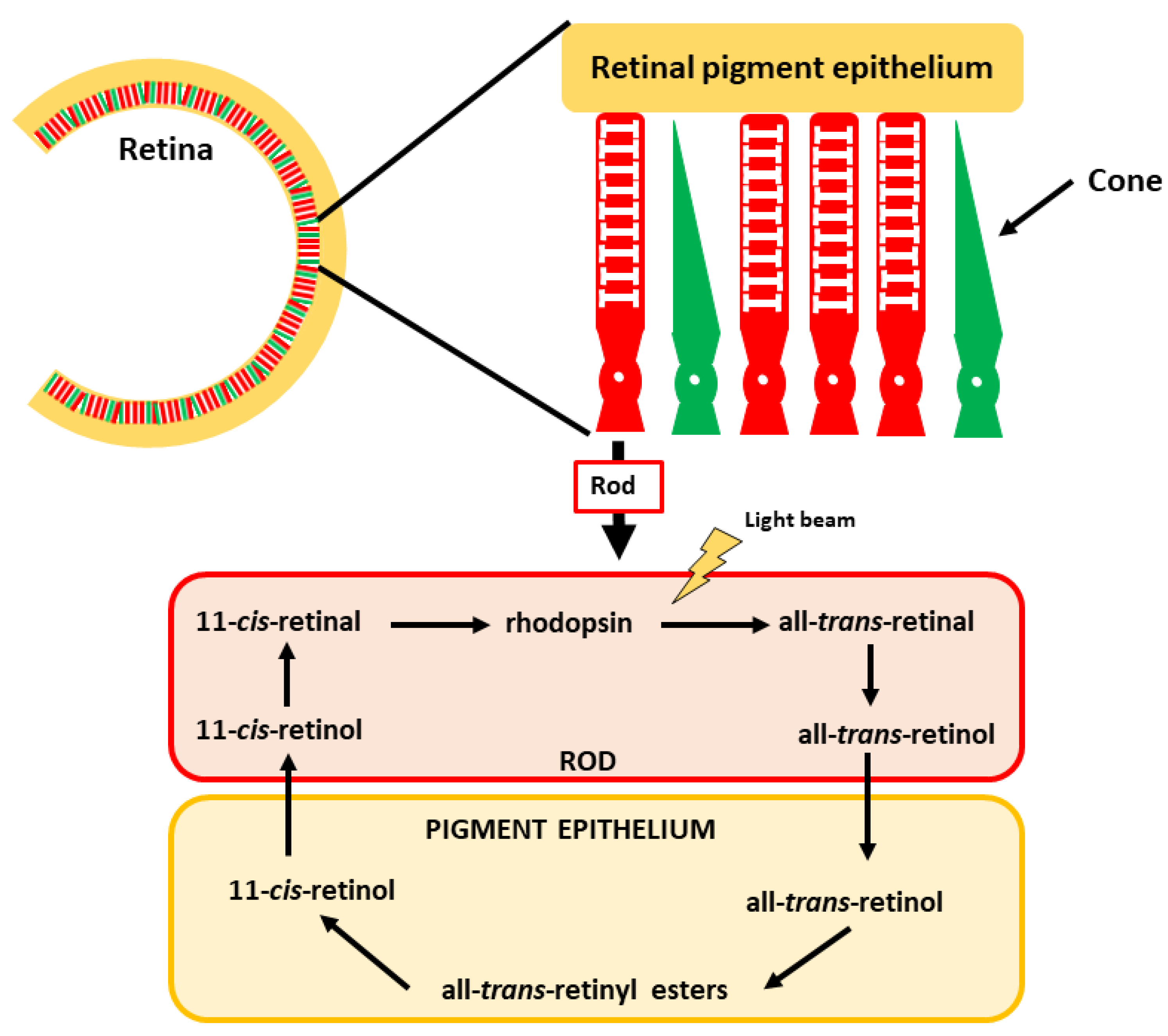{kind=link}
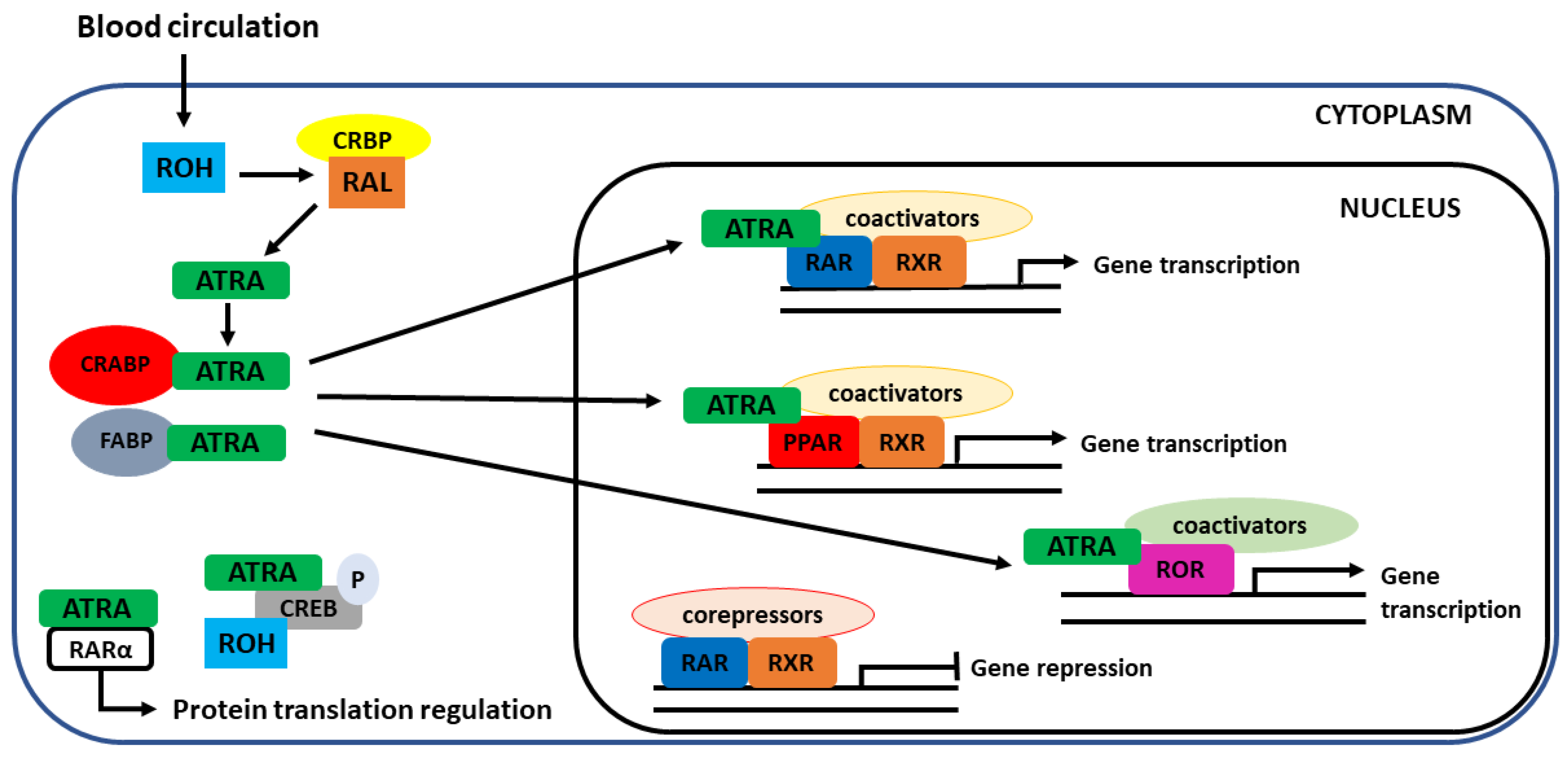{kind=link}
| Source | Latin Name | Family | β-Carotene Content | Refs. |
|---|---|---|---|---|
| Orange-red vegetables | ||||
| Sweet potato | Ipomoea batatas (L.) Lam. | Convolvulaceae | 20–22,600 a | [12,18,22,35] |
| Bitter gourd | Momordica charantia L. | Cucurbitaceae | 17,040 b | [36] |
| Winter squash (butternut) | Cucurbita moschata Duchesne | Cucurbitaceae | 12,340–15,770 a | [21] |
| Chili pepper | Capsicum annuum L., C. chinense Jacq., C. frutescens L. | Solanaceae | 100–15,400 a | [12,37] |
| Carrot | Daucus carota L. | Apiaceae | 4350–8840 a | [35,38] |
| Pumpkins | Cucurbita maxima Duchesne | Cucurbitaceae | 70–6070 a | [12,17,35,38] |
| Cantaloupe | Cucumis melo L. | Cucurbitaceae | 2448–3861 a | [39] |
| Red pepper | Capsicum annuum L. | Solanaceae | 1441–2390 a | [12] |
| Tomato | Solanum lycopersicum L. | Solanaceae | 59–1500 a | [12,35,38] |
| Green vegetables | ||||
| Drumstick leaves | Moringa oleifera Lam. | Moringaceae | 19,700 a | [35] |
| Amaranth | Amaranthus gangeticus L. | Amaranthaceae | 8600 a | [35] |
| Kale | Brassica oleracea var. acephala DC. | Brassicaceae | 1020–10,000 a | [12,40] |
| Garden rocket | Eruca vesicaria (L.) Cav. | Brassicaceae | 7960 a | [41] |
| Chicory | Cichorium intybus L. | Asteraceae | 3940–7310 a | [41] |
| Wild rocket | Diplotaxis tenuifolia (L.) DC. | Brassicaceae | 7010 a | [41] |
| Dandelion | Taraxacum officinale (L.) Weber ex F.H. Wigg. | Asteraceae | 6340 a | [41] |
| Onion leaf | Allium cepa L. | Amaryllidaceae | 4900 a | [35] |
| Coriander | Coriandrum sativum L. | Apiaceae | 4800 a | [35] |
| Parsley | Petroselinum crispum (Mill.) Fuss | Apiaceae | 4440–4680 a | [12] |
| Spinach | Spinacia oleracea L. | Amaranthaceae | 3100–4810 a | [12] |
| Endive | Cichorium endivia L. | Asteraceae | 1340–4350 a | [12] |
| Cress | Lepidium sativum L. | Brassicaceae | 2720–3690 a | [12] |
| Leek | Allium ampeloprasum L. | Amaryllidaceae | 3190 a | [12] |
| Lettuce | Lactuca sativa L. | Asteraceae | 870–2960 a | [12] |
| Broccoli | Brassica oleraceae var. italica Plenck. | Brassicaceae | 291–1750 a | [12] |
| Fruits | ||||
| Apricot | Prunus sect. Armeniaca (Scop.) Koch | Rosaceae | 585–3800 a | [12,19] |
| Mango | Mangifera indica L. | Anacardiaceae | 109–3210 a | [12,35,42] |
| Persimmon | Diospyros kaki L.f. | Ebenaceae | 3000 b | [36] |
| Dates | Phoenix sp. L. | Arecaceae | 2950 a | [35] |
| Guava | Psidium guajava L. | Myrtaceae | 1–2669 a | [12,35] |
| Red grapefruit | Citrus paradisi Macfad. | Rutaceae | 2580 a | [27] |
| Papaya | Carica papaya L. | Caricaceae | 190–1050 a | [35,42,43] |
| Cereals | ||||
| Golden rice | Oryza sativa L. | Poaceae | 160–3700 a | [44] |
| Maize | Zea mays L. | Poaceae | 171–1500 a | [38,44] |
| Medicinal plants and herbs | ||||
| Rose hips | Rosa rubiginosa L. | Rosaceae | 3600 a | [45] |
| Marigold flowers | Calendula officinalis L. | Asteraceae | 940–20,600 a | [46] |
| Dill | Anethum graveolens L. | Apiaceae | 5450 a | [12] |
| Basil | Ocimum basilicum L. | Lamiaceae | 4820 a | [12] |
| Others | ||||
| Spirulina | Spirulina sp. Turpin ex Gomont | Spirulinaceae | 184,100–272,500 a | [31] |
| Sea buckthorn oil | Elaeagnus rhamnoides (L.) A. Nelson | Elaeagnaceae | 16,740 c | [47] |
| Red palm oil | Elaeis guineensis Jacq. | Arecaceae | 5000–5602 c | [48] |
| Technique | Sensitivity (nmol/L) | Matrix | Analytes | Advantages | Disadvantages | References |
|---|---|---|---|---|---|---|
| HPLC-UV-vis/DAD | 1 0.1 × 10−3–209.46 | Serum, plasma, seminal plasma, mouse embryos and kidney, dried whole blood spots, breast milk, red blood cells, adipose tissue | Retinoids (retinol, retinal, RA, retinyl esters) and carotenoids | Usually small sample volume (30–200 µL) Combination with automation and online sample preparation Short analysis times in some multicomponent analyses Some methods have comparable sensitivity to MS detection | Long analysis time with a complicated gradient Use of a large volume of toxic solvents as the mobile phase and in sample preparation procedures, mainly in carotenoids analyses | [279,280,281,282,283,284,285,286,287,288,289,290,291,292,293,294,295] |
| HPLC-FLD | 1 2.3–34.91 | Plasma, breast milk | Retinol | Not often used No sufficient data | [296,297] | |
| LC–MS LC–MS/MS | 2 2 × 10−6–261.83 | Urine, plasma, amniotic fluid, tears, serum | Retinol, RA | Usually, simple sample preparation procedures | In the case of retinoic acid usage of large volumes of toxic solvents in sample preparation procedures with poor recovery | [298,299,300,301,302,303] |
| SFC-MS/MS | 1 0.09 × 10−6–70.31 | Whole blood, plasma, serum, colostrum | Carotenoids, apocarotenoids, epoxycarotenoids | Small sample volume (10–200 µL) Short analysis times in multicomponent analyses Combination with online SFE or robotic SLE | [304,305,306,307] | |
| HPLC-ECD | 1 0.4 × 10−3–314.19 | Serum, rat plasma, cervical tissue | Retinol, RA, carotenoids | Small sample volume (20–200 µL), Short analysis times in multicomponent analyses of retinol and RA | Long analysis time of carotenoids Toxic solvents usage | [308,309,310] |
| ELISA kits | 1 0.11–279.38 | Whole blood, serum, cell lysates, plasma, tissues, other human liquids, colostrum | Retinol, β-carotene | One kit for various matrices (serum, plasma, other biofluids, cell lysates) | For research only Cross-reactivity with retinol analogs Time and money consuming for small sample series | [311,312,313,314,315,316,317,318,319] |
| HPLC/UHPLC-UV kits | 2 23.62–1174.5 | Plasma, serum | Retinol, carotenoids | Small sample volume (50–250 µL) Some kits: possible to combine with a 96-well plate format 96 samples in 30 min Some kits: available in UHPLC mode–3 analytes in 3.5 min | Some kits have long analysis times and use large sample volumes (400 µL) Only for serum or plasma | [320,321,322,323,324] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carazo, A.; Macáková, K.; Matoušová, K.; Krčmová, L.K.; Protti, M.; Mladěnka, P. Vitamin A Update: Forms, Sources, Kinetics, Detection, Function, Deficiency, Therapeutic Use and Toxicity. Nutrients 2021, 13, 1703. https://doi.org/10.3390/nu13051703
Carazo A, Macáková K, Matoušová K, Krčmová LK, Protti M, Mladěnka P. Vitamin A Update: Forms, Sources, Kinetics, Detection, Function, Deficiency, Therapeutic Use and Toxicity. Nutrients. 2021; 13(5):1703. https://doi.org/10.3390/nu13051703
Chicago/Turabian StyleCarazo, Alejandro, Kateřina Macáková, Kateřina Matoušová, Lenka Kujovská Krčmová, Michele Protti, and Přemysl Mladěnka. 2021. "Vitamin A Update: Forms, Sources, Kinetics, Detection, Function, Deficiency, Therapeutic Use and Toxicity" Nutrients 13, no. 5: 1703. https://doi.org/10.3390/nu13051703
APA StyleCarazo, A., Macáková, K., Matoušová, K., Krčmová, L. K., Protti, M., & Mladěnka, P. (2021). Vitamin A Update: Forms, Sources, Kinetics, Detection, Function, Deficiency, Therapeutic Use and Toxicity. Nutrients, 13(5), 1703. https://doi.org/10.3390/nu13051703