
Diversity of Plant Sterols Metabolism: The Impact on Human Health, Sport, and Accumulation of Contaminating Sterols


Abstract
1. Introduction
2. The Prevalence and Variety of Plant Sterols
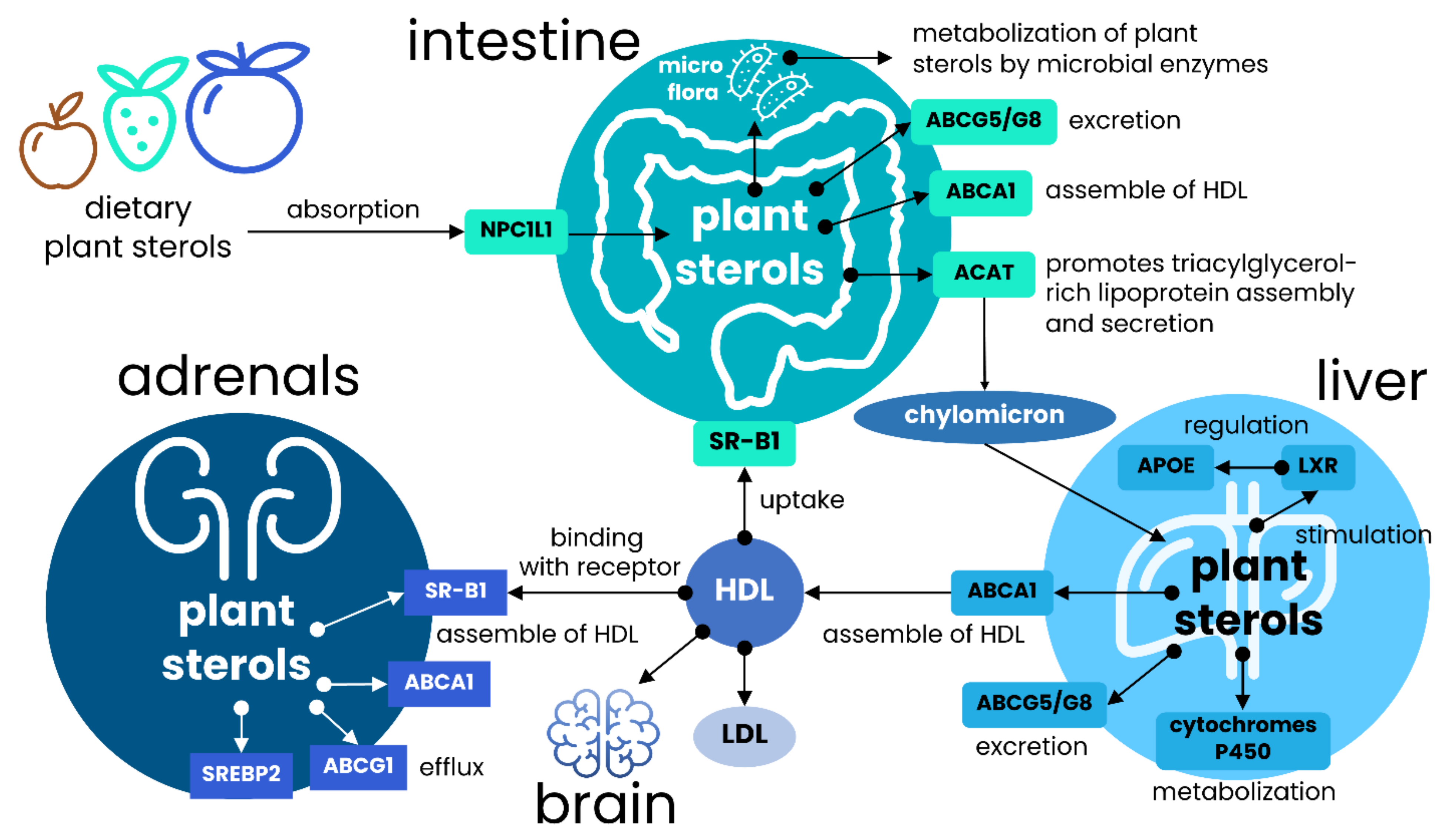
3. Factor Associated with the Efficacy of Plant Sterols Absorption
4. Benefits of Plant Sterols in Clinical Application
4.1. Cardioprotective Property of Plants Sterols
4.2. Neuroprotection and Neuroimmunomodulation
4.3. Anti-Aging and Skin Regeneration Effects
5. Dietary Sterols in Sport
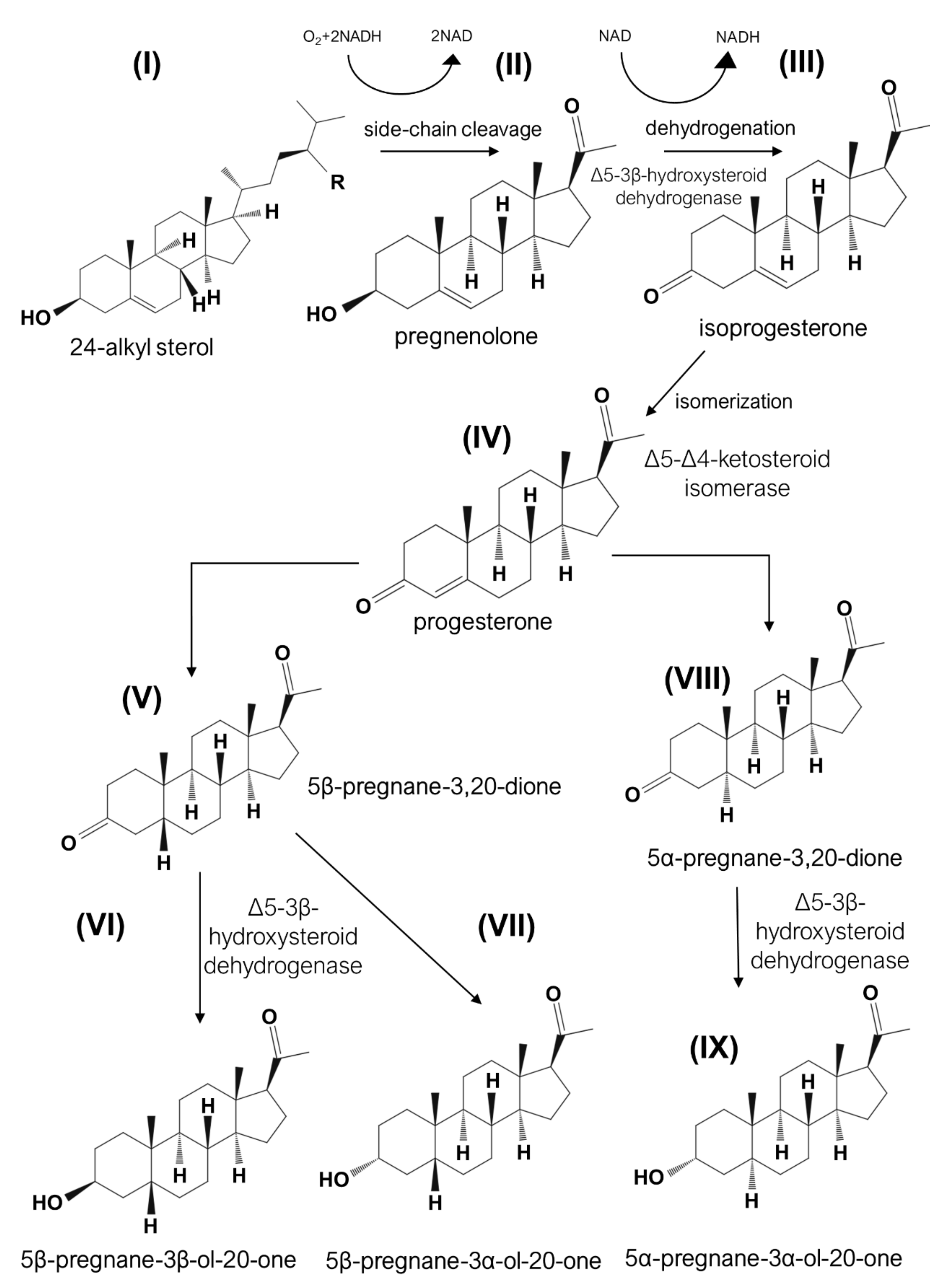
6. Fake Sterols and Contaminations
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Trautwein, E.A.; Vermeer, M.A.; Hiemstra, H.; Ras, R.T. LDL-Cholesterol Lowering of Plant Sterols and Stanols—Which Factors Influence Their Efficacy? Nutrients 2018, 10, 1262. [Google Scholar] [CrossRef]
- Marrone, G.; Guerriero, C.; Palazzetti, D.; Lido, P.; Marolla, A.; Di Daniele, F. Vegan Diet Health Benefits in Metabolic Syn-drome. Nutrients 2021, 13, 817. [Google Scholar] [CrossRef]
- Jones, P.J.H. Inter-individual Variability in Response to Plant Sterol and Stanol Consumption. J. AOAC Int. 2015, 98, 724–728. [Google Scholar] [CrossRef]
- Morand, C.; Tomás Barberán, F.A. Contribution of plant food bioactives in promoting health effects of plant foods: Why look at interindividual variability? Eur. J. Nutr. 2019, 58, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Milenkovic, D.; Berghe WVanden Morand, C.; Claude, S.; van de Sandt, A.; Gorressen, S. A systems biology network analy-sis of nutri(epi)genomic changes in endothelial cells exposed to epicatechin metabolites. Sci. Rep. 2018, 8, 15487. [Google Scholar] [CrossRef] [PubMed]
- Espín, J.C.; González-Sarrías, A.; Tomás-Barberán, F.A. The gut microbiota: A key factor in the therapeutic effects of (poly)phenols. Biochem. Pharmacol. 2017, 139, 82–93. [Google Scholar] [CrossRef] [PubMed]
- Barber-Chamoux, N.; Milenkovic, D.; Verny, M.-A.; Habauzit, V.; Pereira, B.; Lambert, C. Substantial Variability Across Individuals in the Vascular and Nutrigenomic Response to an Acute Intake of Curcumin: A Randomized Controlled Trial. Mol. Nutr. Food Res. 2018, 62, 1700418. [Google Scholar] [CrossRef]
- Heiss, C.; Sansone, R.; Karimi, H.; Krabbe, M.; Schuler, D.; Rodriguez-Mateos, A.; Kraemer, T.; Cortese-Krott, M.M.; Kuhnle, G.G.C.; Spencer, J.P.E.; et al. Impact of cocoa flavanol intake on age-dependent vascular stiffness in healthy men: A randomized, controlled, double-masked trial. AGE 2015, 37, 1–12. [Google Scholar] [CrossRef]
- Carlson, J.J.; Eisenmann, J.C.; Norman, G.J.; Ortiz, K.A.; Young, P.C. Dietary Fiber and Nutrient Density Are Inversely Associated with the Metabolic Syndrome in US Adolescents. J. Am. Diet. Assoc. 2011, 111, 1688–1695. [Google Scholar] [CrossRef]
- Ziaee, R.S.; Keshani, P.; Salehi, M.; Ghaem, H. Diet Quality Indices and Their Correlation with Glycemic Status and Lipid Profile in Patients with Type 2 Diabetes. Adv. Prev. Med. 2021, 2021, 1–8. [Google Scholar] [CrossRef]
- Turner-McGrievy, G.M.; Barnard, N.D.; Cohen, J.; Jenkins, D.J.; Gloede, L.; Green, A.A. Changes in Nutrient Intake and Dietary Quality among Participants with Type 2 Diabetes Following a Low-Fat Vegan Diet or a Conventional Diabetes Diet for 22 Weeks. J. Am. Diet. Assoc. 2008, 108, 1636–1645. [Google Scholar] [CrossRef]
- Dinu, M.; Abbate, R.; Gensini, G.F.; Casini, A.; Sofi, F. Vegetarian, vegan diets and multiple health outcomes: A systematic review with meta-analysis of observational studies. Crit. Rev. Food Sci. Nutr. 2017, 57, 3640–3649. [Google Scholar] [CrossRef]
- Baumgartner, S.; Mensink, R.P.; Plat, J. Plant Sterols and Stanols in the Treatment of Dyslipidemia: New Insights into Targets and Mechanisms Related to Cardiovascular Risk. Curr. Pharm. Des. 2011, 17, 922–932. [Google Scholar] [CrossRef]
- Bruckert, E.; Rosenbaum, D. Lowering LDL-cholesterol through diet: Potential role in the statin era. Curr. Opin. Lipidol. 2011, 22, 43–48. [Google Scholar] [CrossRef]
- Weingärtner, O.; Teupser, D.; Patel, S.B. The Atherogenicity of Plant Sterols: The Evidence from Genetics to Clinical Trials. J. AOAC Int. 2015, 98, 742–749. [Google Scholar] [CrossRef]
- Tundis, R.; Loizzo, M.; Bonesi, M.; Menichini, F. Potential Role of Natural Compounds Against Skin Aging. Curr. Med. Chem. 2015, 22, 1515–1538. [Google Scholar] [CrossRef]
- Trautwein, E.A.; McKay, S. The Role of Specific Components of a Plant-Based Diet in Management of Dyslipidemia and the Impact on Cardiovascular Risk. Nutrients 2020, 12, 2671. [Google Scholar] [CrossRef]
- Moreau, R.A. Composition of Plant Sterols and Stanols in Supplemented Food Products. J. AOAC Int. 2015, 98, 685–690. [Google Scholar] [CrossRef]
- Yang, R.; Xue, L.; Zhang, L.; Wang, X.; Qi, X.; Jiang, J. Phytosterol Contents of Edible Oils and Their Contributions to Estimat-ed Phytosterol Intake in the Chinese Diet. Foods 2019, 8, 334. [Google Scholar] [CrossRef]
- Patel, S.B. Plant sterols and stanols: Their role in health and disease. J. Clin. Lipidol. 2008, 2, S11–S19. [Google Scholar] [CrossRef]
- Cabral, C.E.; Klein, M.R.S.T. Phytosterols in the Treatment of Hypercholesterolemia and Prevention of Cardiovascular Diseases. Arq. Bras. Cardiol. 2017, 109, 475–482. [Google Scholar] [CrossRef]
- Cusack, L.K.; Fernandez, M.L.; Volek, J.S. The Food Matrix and Sterol Characteristics Affect the Plasma Cholesterol Lowering of Phytosterol/Phytostanol. Adv. Nutr. 2013, 4, 633–643. [Google Scholar] [CrossRef]
- Valitova, J.N.; Sulkarnayeva, A.G.; Minibayeva, F.V. Plant sterols: Diversity, biosynthesis, and physiological functions. Biochemistry 2016, 81, 819–834. [Google Scholar] [CrossRef]
- Ferrer, A.; Altabella, T.; Arró, M.; Boronat, A. Emerging roles for conjugated sterols in plants. Prog. Lipid Res. 2017, 67, 27–37. [Google Scholar] [CrossRef]
- Benveniste, P. Biosynthesis and Accumulation of Sterols. Annu. Rev. Plant Biol. 2004, 55, 429–457. [Google Scholar] [CrossRef]
- Patel, S.B.; Honda, A.; Salen, G. Sitosterolemia: Exclusion of genes involved in reduced cholesterol biosynthesis. J. Lipid Res. 1998, 39, 1055–1061. [Google Scholar] [CrossRef]
- Malina, D.M.T.; Fonseca, F.A.; Barbosa, S.A.; Kasmas, S.H.; Machado, V.A.; França, C.N. Additive effects of plant sterols supple-mentation in addition to different lipid-lowering regimens. J. Clin. Lipidol. 2015, 9, 542–552. [Google Scholar] [CrossRef]
- Rysz, J.; Franczyk, B.; Olszewski, R.; Banach, M.; Gluba-Brzozka, A. The Use of Plant Sterols and Stanols as Lipid-Lowering Agents in Cardiovascular Disease. Curr. Pharm. Des. 2017, 23, 2488–2495. [Google Scholar] [CrossRef]
- Sahebkar, A.; Serban, M.-C.; Gluba-Brzózka, A.; Mikhailidis, D.P.; Cicero, A.F.; Rysz, J. Lipid-modifying effects of nutraceuticals: An evidence-based approach. Nutrition 2016, 32, 1179–1192. [Google Scholar] [CrossRef]
- Demonty, I.; Ras, R.T.; van der Knaap, H.C.M.; Duchateau, G.S.M.J.E.; Meijer, L.; Zock, P.L. Continuous dose-response relationship of the LDL-cholesterol-lowering effect of phytosterol intake. J. Nutr. 2009, 139, 271–284. [Google Scholar] [CrossRef]
- von Bergmann, K.; Sudhop, T.; Lütjohann, D. Cholesterol and plant sterol absorption: Recent insights. Am. J. Cardiol. 2005, 96, 10D–14D. [Google Scholar] [CrossRef] [PubMed]
- Kreuzer, J. Phytosterols and phytostanols: Is it time to rethink that supplemented margarine? Cardiovasc. Res. 2011, 90, 397–398. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Silbernagel, G.; Baumgartner, I.; März, W. Cardiovascular Safety of Plant Sterol and Stanol Consumption. J. AOAC Int. 2015, 98, 739–741. [Google Scholar] [CrossRef] [PubMed]
- Ras, R.T.; Geleijnse, J.M.; Trautwein, E.A. LDL-cholesterol-lowering effect of plant sterols and stanols across different dose rang-es: A meta-analysis of randomised controlled studies. Br. J. Nutr. 2014, 112, 214–219. [Google Scholar] [CrossRef] [PubMed]
- Thomsen, A.B.; Hansen, H.B.; Christiansen, C.; Green, H.; Berger, A. Effect of free plant sterols in low-fat milk on serum lipid profile in hypercholesterolemic subjects. Eur. J. Clin. Nutr. 2004, 58, 860–870. [Google Scholar] [CrossRef]
- Doornbos, A.M.E.; Meynen, E.M.; Duchateau, G.S.M.J.E.; Van Der Knaap, H.C.M.; A Trautwein, E. Intake occasion affects the serum cholesterol lowering of a plant sterol-enriched single-dose yoghurt drink in mildly hypercholesterolaemic subjects. Eur. J. Clin. Nutr. 2006, 60, 325–333. [Google Scholar] [CrossRef]
- Nestel, P.; Cehun, M.; Pomeroy, S.; Abbey, M.; Weldon, G. Cholesterol-lowering effects of plant sterol esters and non-esterified stanols in margarine, butter and low-fat foods. Eur. J. Clin. Nutr. 2001, 55, 1084–1090. [Google Scholar] [CrossRef]
- Amir Shaghaghi, M.; Abumweis, S.S.; Jones, P.J.H. Cholesterol-lowering efficacy of plant sterols/stanols provided in capsule and tablet formats: Results of a systematic review and meta-analysis. J Acad. Nutr. Diet. 2013, 113, 1494–1503. [Google Scholar] [CrossRef]
- Blanco Mejia, S.; Messina, M.; Li, S.S.; Viguiliouk, E.; Chiavaroli, L.; Khan, T.A. A Meta-Analysis of 46 Studies Identified by the FDA Demonstrates that Soy Protein Decreases Circulating LDL and Total Cholesterol Concentrations in Adults. J. Nutr. 2019, 149, 968–981. [Google Scholar] [CrossRef]
- Maki, K.C.; Lawless, A.L.; Reeves, M.S.; Dicklin, M.R.; Jenks, B.H.; Shneyvas, E.; Brooks, J.R. Lipid-altering effects of a dietary supplement tablet containing free plant sterols and stanols in men and women with primary hypercholesterolaemia: A randomized, placebo-controlled crossover trial. Int. J. Food Sci. Nutr. 2012, 63, 476–482. [Google Scholar] [CrossRef]
- McPherson, T.B.; Ostlund, R.E.; Goldberg, A.C.; Bateman, J.H.; Schimmoeller, L.; Spilburg, C.A. Phytostanol tablets reduce human LDL-cholesterol. J. Pharm. Pharmacol. 2010, 57, 889–896. [Google Scholar] [CrossRef]
- Ottestad, I.; Ose, L.; Wennersberg, M.H.; Granlund, L.; Kirkhus, B.; Retterstøl, K. Phytosterol capsules and serum cholesterol in hy-percholesterolemia: A randomized controlled trial. Atherosclerosis 2013, 228, 421–425. [Google Scholar] [CrossRef]
- Plösch, T.; Bloks, V.W.; Terasawa, Y.; Berdy, S.; Siegler, K.; van der Sluijs, F.; Kema, I.P.; Groen, A.K.; Shan, B.; Kuipers, F.; et al. Sitosterolemia in ABC-Transporter G5-deficient mice is aggravated on activation of the liver-X receptor. Gastroenterology 2004, 126, 290–300. [Google Scholar] [CrossRef]
- Yu, X.-H.; Qian, K.; Jiang, N.; Zheng, X.-L.; Cayabyab, F.S.; Tang, C.-K. ABCG5/ABCG8 in cholesterol excretion and atherosclerosis. Clin. Chim. Acta 2014, 428, 82–88. [Google Scholar] [CrossRef]
- Calpe-Berdiel, L.; Escolà-Gil, J.C.; Blanco-Vaca, F. New insights into the molecular actions of plant sterols and stanols in choles-terol metabolism. Atherosclerosis 2009, 203, 18–31. [Google Scholar] [CrossRef]
- Wouters, E.; De Wit, N.M.; Vanmol, J.; Van Der Pol, S.M.A.; Hof, B.V.H.; Sommer, D.; Loix, M.; Geerts, D.; Gustafsson, J.A.; Steffensen, K.R.; et al. Liver X Receptor Alpha Is Important in Maintaining Blood-Brain Barrier Function. Front. Immunol. 2019, 10, 1811. [Google Scholar] [CrossRef]
- Ostlund, R.E.J.; McGill, J.B.; Zeng, C.-M.; Covey, D.F.; Stearns, J.; Stenson, W.F. Gastrointestinal absorption and plasma kinetics of soy Delta(5)-phytosterols and phytostanols in humans. Am. J. Physiol. Endocrinol. Metab. 2002, 282, E911–E916. [Google Scholar] [CrossRef]
- Yu, L.; Hammer, R.E.; Li-Hawkins, J.; von Bergmann, K.; Lutjohann, D.; Cohen, J.C.; Hobbs, H.H. Disruption of Abcg5 and Abcg8 in mice reveals their crucial role in biliary cholesterol secretion. Proc. Natl. Acad. Sci. USA 2002, 99, 16237–16242. [Google Scholar] [CrossRef]
- Yu, L.; von Bergmann, K.; Lutjohann, D.; Hobbs, H.H.; Cohen, J.C. Selective sterol accumulation in ABCG5/ABCG8-deficient mice. J. Lipid Res. 2004, 45, 301–307. [Google Scholar] [CrossRef]
- Helgadottir, A.; Thorleifsson, G.; Alexandersson, K.F.; Tragante, V.; Thorsteinsdottir, M.; Eiriksson, F.F.; Gretarsdottir, S.; Björnsson, E.; Magnusson, O.; Sveinbjornsson, G.; et al. Genetic variability in the absorption of dietary sterols affects the risk of coronary artery disease. Eur. Heart J. 2020, 41, 2618–2628. [Google Scholar] [CrossRef]
- Ikeda, I. Factors Affecting Intestinal Absorption of Cholesterol and Plant Sterols and Stanols. J. Oleo Sci. 2015, 64, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Best, M.M.; Duncan, C.H.; Van Loon, E.J.; Wathen, J.D. Lowering of serum cholesterol by the administration of a plant sterol. Circulation 1954, 10, 201–206. [Google Scholar] [CrossRef] [PubMed]
- De Smet, E.; Mensink, R.P.; Lütjohann, D.; Plat, J. Acute effects of plant stanol esters on postprandial metabolism and its relation with changes in serum lipids after chronic intake. Eur. J. Clin. Nutr. 2015, 69, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Gylling, H.; A Miettinen, T. The effect of plant stanol- and sterol-enriched foods on lipid metabolism, serum lipids and coronary heart disease. Ann. Clin. Biochem. 2005, 42, 254–263. [Google Scholar] [CrossRef]
- Hallikainen, M.; Lyyra-Laitinen, T.; Laitinen, T.; Agren, J.J.; Pihlajamäki, J.; Rauramaa, R. Endothelial function in hypercholes-terolemic subjects: Effects of plant stanol and sterol esters. Atherosclerosis 2006, 188, 425–432. [Google Scholar] [CrossRef]
- Miettinen, T.A.; Gylling, H.; Strandberg, T.; Sarna, S. Baseline serum cholestanol as predictor of recurrent coronary events in sub-group of Scandinavian simvastatin survival study. Finnish 4S Investigators. BMJ 1998, 316, 1127–1130. [Google Scholar] [CrossRef]
- Rajaratnam, R.A.; Gylling, H.; Miettinen, T.A. Independent association of serum squalene and noncholesterol sterols with coro-nary artery disease in postmenopausal women. J. Am. Coll. Cardiol. 2000, 35, 1185–1191. [Google Scholar] [CrossRef]
- Silbernagel, G.; Fauler, G.; Renner, W.; Landl, E.M.; Hoffmann, M.M.; Winkelmann, B.R. The relationships of cholesterol metab-olism and plasma plant sterols with the severity of coronary artery disease. J. Lipid Res. 2009, 50, 334–341. [Google Scholar] [CrossRef]
- Escurriol, V.; Cofán, M.; Moreno-Iribas, C.; Larrañaga, N.; Martínez, C.; Navarro, C.; Rodríguez, L.; González, C.A.; Corella, D.; Ros, E. Phytosterol plasma concentrations and coronary heart disease in the prospective Spanish EPIC cohort. J. Lipid Res. 2010, 51, 618–624. [Google Scholar] [CrossRef]
- Pinedo, S.; Vissers, M.N.; von Bergmann, K.; Elharchaoui, K.; Lütjohann, D.; Luben, R.; Wareham, N.J.; Kastelein, J.J.P.; Khaw, K.-T.; Boekholdt, S.M. Plasma levels of plant sterols and the risk of coronary artery disease: The prospective EPIC-Norfolk Population Study. J. Lipid Res. 2007, 48, 139–144. [Google Scholar] [CrossRef]
- Calpe-Berdiel, L.; Escolà-Gil, J.C.; Benítez, S.; Bancells, C.; González-Sastre, F.; Palomer, X.; Blanco-Vaca, F. Dietary phytosterols modulate T-helper immune response but do not induce apparent anti-inflammatory effects in a mouse model of acute, aseptic inflammation. Life Sci. 2007, 80, 1951–1956. [Google Scholar] [CrossRef]
- Brüll, F.; Mensink, R.P.; van den Hurk, K.; Duijvestijn, A.; Plat, J. TLR2 activation is essential to induce a Th1 shift in human pe-ripheral blood mononuclear cells by plant stanols and plant sterols. J. Biol. Chem. 2010, 285, 2951–2958. [Google Scholar] [CrossRef]
- Adetutu, A.; Morgan, W.A.; Corcoran, O. Ethnopharmacological survey and in vitro evaluation of wound-healing plants used in South-western Nigeria. J. Ethnopharmacol. 2011, 137, 50–56. [Google Scholar] [CrossRef]
- Esposito, D.; Rathinasabapathy, T.; Schmidt, B.; Shakarjian, M.P.; Komarnytsky, S.; Raskin, I. Acceleration of cutaneous wound healing by brassinosteroids. Wound Repair Regen. 2013, 21, 688–696. [Google Scholar] [CrossRef]
- Smiljanic, K.; Vanmierlo, T.; Djordjevic, A.M.; Perovic, M.; Loncarevic-Vasiljkovic, N.; Tešić, V.; Rakić, L.; Ruzdijic, S.; Lütjohann, D.; Kanazir, S. Aging Induces Tissue-Specific Changes in Cholesterol Metabolism in Rat Brain and Liver. Lipids 2013, 48, 1069–1077. [Google Scholar] [CrossRef]
- Teunissen, C.E.; De Vente, J.; Von Bergmann, K.; Bosma, H.; Van Boxtel, M.P.J.; De Bruijn, C.; Jolles, J.; Steinbusch, H.W.M.; Lütjohann, D. Serum cholesterol, precursors and metabolites and cognitive performance in an aging population. Neurobiol. Aging 2002, 24, 147–155. [Google Scholar] [CrossRef]
- Gylling, H.; Radhakrishnan, R.; Miettinen, T.A. Reduction of serum cholesterol in postmenopausal women with previous myo-cardial infarction and cholesterol malabsorption induced by dietary sitostanol ester margarine: Women and dietary sitostanol. Circulation 1997, 96, 4226–4231. [Google Scholar] [CrossRef]
- Verheyden, K.; Noppe, H.; Vanhaecke, L.; Wille, K.; Bussche JVanden Bekaert, K. Excretion of endogenous boldione in hu-man urine: Influence of phytosterol consumption. J. Steroid. Biochem. Mol. Biol. 2009, 117, 8–14. [Google Scholar] [CrossRef]
- Theuwissen, E.; Plat, J.; Van Der Kallen, C.J.; Van Greevenbroek, M.M.; Mensink, R.P. Plant Stanol Supplementation Decreases Serum Triacylglycerols in Subjects with Overt Hypertriglyceridemia. Lipids 2009, 44, 1131–1140. [Google Scholar] [CrossRef]
- Esposito, D.; Damsud, T.; Wilson, M.; Grace, M.H.; Strauch, R.; Li, X.; Lila, M.A.; Komarnytsky, S. Black Currant Anthocyanins Attenuate Weight Gain and Improve Glucose Metabolism in Diet-Induced Obese Mice with Intact, but Not Disrupted, Gut Microbiome. J. Agric. Food Chem. 2015, 63, 6172–6180. [Google Scholar] [CrossRef]
- Saeed, A.A.; Genové, G.; Li, T.; Lütjohann, D.; Olin, M.; Mast, N. Effects of a disrupted blood-brain barrier on cholesterol ho-meostasis in the brain. J. Biol. Chem. 2014, 289, 23712–23722. [Google Scholar] [CrossRef]
- Saeed, A.A.; Genové, G.; Li, T.; Hülshorst, F.; Betsholtz, C.; Björkhem, I.; Lütjohann, D. Increased flux of the plant sterols campesterol and sitosterol across a disrupted blood brain barrier. Steroids 2015, 99, 183–188. [Google Scholar] [CrossRef]
- Jansen, P.J.; Lütjohann, D.; Abildayeva, K.; Vanmierlo, T.; Plösch, T.; Plat, J.; von Bergmann, K.; Groen, A.K.; Ramaekers, F.C.; Kuipers, F. Dietary plant sterols accumulate in the brain. Biochim. Biophys. Acta 2006, 1761, 445–453. [Google Scholar] [CrossRef]
- Kim, H.-J.; Fan, X.; Gabbi, C.; Yakimchuk, K.; Parini, P.; Warner, M. Liver X receptor beta (LXRbeta): A link between be-ta-sitosterol and amyotrophic lateral sclerosis-Parkinson’s dementia. Proc. Natl. Acad. Sci. USA 2008, 105, 2094–2099. [Google Scholar] [CrossRef]
- Schött, H.-F.; Husche, C.; Friedrichs, S.; Miller, C.M.; McCarthy, F.O.; Laufs, U. 7β-Hydroxysitosterol crosses the blood-brain barrier more favored than its substrate sitosterol in ApoE−/− mice. Steroids 2015, 99 (Pt B), 178–182. [Google Scholar] [CrossRef]
- Nieminen, P.; Mustonen, A.-M.; Lindström-Seppä, P.; Asikainen, J.; Mussalo-Rauhamaa, H.; Kukkonen, J.V.K. Phytosterols act as endocrine and metabolic disruptors in the European polecat (Mustela putorius). Toxicol. Appl. Pharmacol. 2002, 178, 22–28. [Google Scholar] [CrossRef]
- Ryökkynen, A.; Käyhkö, U.-R.; Mustonen, A.-M.; Kukkonen, J.V.K.; Nieminen, P. Multigenerational exposure to phytosterols in the mouse. Reprod. Toxicol. 2005, 19, 535–540. [Google Scholar] [CrossRef]
- Aldini, R.; Micucci, M.; Cevenini, M.; Fato, R.; Bergamini, C.; Nanni, C. Antiinflammatory effect of phytosterols in experimental murine colitis model: Prevention, induction, remission study. PLoS ONE 2014, 9, e108112. [Google Scholar] [CrossRef] [PubMed]
- Balekar, N.; Katkam, N.G.; Nakpheng, T.; Jehtae, K.; Srichana, T. Evaluation of the wound healing potential of Wedelia trilobata (L.) leaves. J. Ethnopharmacol. 2012, 141, 817–824. [Google Scholar] [CrossRef] [PubMed]
- Van Kampen, J.M.; Robertson, H.A. The BSSG rat model of Parkinson’s disease: Progressing towards a valid, predictive model of disease. EPMA J. 2017, 8, 261–271. [Google Scholar] [CrossRef] [PubMed]
- Bogie, J.; Hoeks, C.; Schepers, M.; Tiane, A.; Cuypers, A.; Leijten, F. Dietary Sargassum fusiforme improves memory and re-duces amyloid plaque load in an Alzheimer’s disease mouse model. Sci. Rep. 2019, 9, 4908. [Google Scholar] [CrossRef]
- Berghoff, S.A.; Gerndt, N.; Winchenbach, J.; Stumpf, S.K.; Hosang, L.; Odoardi, F. Dietary cholesterol promotes repair of demy-elinated lesions in the adult brain. Nat. Commun. 2017, 8, 14241. [Google Scholar] [CrossRef]
- Nomaguchi, K.; Tanaka, M.; Misawa, E.; Yamada, M.; Toida, T.; Iwatsuki, K.; Goto, T.; Kawada, T. Aloe vera phytosterols act as ligands for PPAR and improve the expression levels of PPAR target genes in the livers of mice with diet-induced obesity. Obes. Res. Clin. Pract. 2011, 5, e190–e201. [Google Scholar] [CrossRef]
- Klaikeaw, N.; Wongphoom, J.; Werawatganon, D.; Chayanupatkul, M.; Siriviriyakul, P. Anti-inflammatory and anti-oxidant effects of Aloe vera in rats with non-alcoholic steatohepatitis. World J. Hepatol. 2020, 12, 363–377. [Google Scholar] [CrossRef]
- Esposito, D.; Rathinasabapathy, T.; Poulev, A.; Komarnytsky, S.; Raskin, I. Akt-dependent anabolic activity of natural and syn-thetic brassinosteroids in rat skeletal muscle cells. J. Med. Chem. 2011, 54, 4057–4066. [Google Scholar] [CrossRef]
- Cheng, D.M.; Kutzler, L.W.; Boler, D.D.; Drnevich, J.; Killefer, J.; Lila, M.A. Continuous Infusion of 20-Hydroxyecdysone Increased Mass of Triceps Brachii in C57BL/6 Mice. Phytother. Res. 2013, 27, 107–111. [Google Scholar] [CrossRef]
- Hirunsai, M.; Yimlamai, T.; Suksamrarn, A. Effect of 20-Hydroxyecdysone on Proteolytic Regulation in Skeletal Muscle Atrophy. In Vivo 2016, 30, 869–878. [Google Scholar] [CrossRef]
- Gorelick-Feldman, J.; Cohick, W.; Raskin, I. Ecdysteroids elicit a rapid Ca2+ flux leading to Akt activation and increased protein synthesis in skeletal muscle cells. Steroids 2010, 75, 632–637. [Google Scholar] [CrossRef]
- Josefsen, K.D.; Nordborg, A.; Sletta, H. Bioconversion of Phytosterols into Androstenedione by Mycobacterium. Methods Mol. Biol. 2017, 1645, 177–197. [Google Scholar] [CrossRef]
- Rideout, T.C.; Chan, Y.-M.; Harding, S.V.; Jones, P.J. Low and moderate-fat plant sterol fortified soymilk in modulation of plasma lipids and cholesterol kinetics in subjects with normal to high cholesterol concentrations: Report on two randomized crosso-ver studies. Lipids Health Dis. 2009, 8, 45. [Google Scholar] [CrossRef]
- Mensink, R.P.; de Jong, A.; Lütjohann, D.; Haenen, G.R.; Plat, J. Plant stanols dose-dependently decrease LDL-cholesterol concen-trations, but not cholesterol-standardized fat-soluble antioxidant concentrations, at intakes up to 9 g/d. Am. J. Clin. Nutr. 2010, 92, 24–33. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Davidson, M.H.; Maki, K.C.; Umporowicz, D.M.; Ingram, K.A.; Dicklin, M.R.; Schaefer, E.; Lane, R.W.; McNamara, J.R.; Ribaya-Mercado, J.D.; Perrone, G.; et al. Safety and tolerability of esterified phytosterols administered in reduced-fat spread and salad dressing to healthy adult men and women. J. Am. Coll. Nutr. 2001, 20, 307–319. [Google Scholar] [CrossRef] [PubMed]
- Jones, P.J.H.; Shamloo, M.; MacKay, D.S.; Rideout, T.C.; Myrie, S.B.; Plat, J.; Roullet, J.-B.; Baer, D.J.; Calkins, K.L.; Davis, H.R.; et al. Progress and perspectives in plant sterol and plant stanol research. Nutr. Rev. 2018, 76, 725–746. [Google Scholar] [CrossRef] [PubMed]
- Okada, K.; Iwahashi, N.; Endo, T.; Himeno, H.; Fukui, K.; Kobayashi, S.; Shimizu, M.; Iwasawa, Y.; Morita, Y.; Wada, A.; et al. Long-term effects of ezetimibe-plus-statin therapy on low-density lipoprotein cholesterol levels as compared with double-dose statin therapy in patients with coronary artery disease. Atherosclerosis 2012, 224, 454–456. [Google Scholar] [CrossRef] [PubMed]
- Rideout, T.C.; Harding, S.V.; Mackay, D.S. Metabolic and genetic factors modulating subject specific LDL-C responses to plant sterol therapy. Can. J. Physiol. Pharmacol. 2012, 90, 509–514. [Google Scholar] [CrossRef] [PubMed]
- Kawabata, K.; Yoshioka, Y.; Terao, J. Role of Intestinal Microbiota in the Bioavailability and Physiological Functions of Dietary Polyphenols. Molecules 2019, 24, 370. [Google Scholar] [CrossRef]
- Panzenboeck, U.; Balazs, Z.; Sovic, A.; Hrzenjak, A.; Levak-Frank, S.; Wintersperger, A.; Malle, E.; Sattler, W. ABCA1 and Scavenger Receptor Class B, Type I, Are Modulators of Reverse Sterol Transport at an in Vitro Blood-Brain Barrier Constituted of Porcine Brain Capillary Endothelial Cells. J. Biol. Chem. 2002, 277, 42781–42789. [Google Scholar] [CrossRef]
- Dietschy, J.M.; Turley, S.D. Cholesterol metabolism in the brain. Curr. Opin. Lipidol. 2001, 12, 105–112. [Google Scholar] [CrossRef]
- Wang, Y.; Muneton, S.; Sjövall, J.; Jovanovic, J.N.; Griffiths, W.J. The Effect of 24S-Hydroxycholesterol on Cholesterol Homeostasis in Neurons: Quantitative Changes to the Cortical Neuron Proteome. J. Proteome Res. 2008, 7, 1606–1614. [Google Scholar] [CrossRef]
- Hac-Wydro, K.; Wydro, P.; Dynarowicz-Łatka, P.; Paluch, M. Cholesterol and phytosterols effect on sphingomye-lin/phosphatidylcholine model membranes--thermodynamic analysis of the interactions in ternary monolayers. J. Colloid Interface Sci. 2009, 329, 265–272. [Google Scholar] [CrossRef]
- Vanmierlo, T.; Popp, J.; Kölsch, H.; Friedrichs, S.; Jessen, F.; Stoffel-Wagner, B.; Bertsch, T.; Hartmann, T.; Maier, W.; Von Bergmann, K.; et al. The plant sterol brassicasterol as additional CSF biomarker in Alzheimer’s disease. Acta Psychiatr. Scand. 2011, 124, 184–192. [Google Scholar] [CrossRef]
- Burg, V.K.; Grimm, H.S.; Rothhaar, T.L.; Grösgen, S.; Hundsdörfer, B.; Haupenthal, V.J.; Zimmer, V.C.; Mett, J.; Weingärtner, O.; Laufs, U.; et al. Plant Sterols the Better Cholesterol in Alzheimer’s Disease? A Mechanistical Study. J. Neurosci. 2013, 33, 16072–16087. [Google Scholar] [CrossRef]
- Lobo, A.; Quintanilla, M.A. The search of new biomarkers to identify Alzheimer’s disease: An editorial comment to T. Vanmierlo et al. ‘The Plant Sterol Brassicasterol and Additional CFS Biomarker in Alzheimer’s Disease ‘(1). Acta Psychiatr. Scand. 2011, 124, 163–164. [Google Scholar] [CrossRef]
- Vanmierlo, T.; Rutten, K.; Friedrichs, S.; Bloks, V.W.; Blokland, A. Cerebral accumulation of dietary derivable plant sterols does not interfere with memory and anxiety related behavior in Abcg5−/− mice. Plant Foods Hum. Nutr. 2011, 66, 149–156. [Google Scholar] [CrossRef]
- Shafaati, M.; Marutle, A.; Pettersson, H.; Lövgren-Sandblom, A.; Olin, M.; Pikuleva, I. Marked accumulation of 27-hydroxycholesterol in the brains of Alzheimer’s patients with the Swedish APP 670/671 mutation. J. Lipid Res. 2011, 52, 1004–1010. [Google Scholar] [CrossRef]
- Liu, R.; Hao, D.; Xu, W.; Li, J.; Li, X.; Shen, D.; Sheng, K.; Zhao, L.; Xu, W.; Gao, Z.; et al. β-Sitosterol modulates macrophage polarization and attenuates rheumatoid inflammation in mice. Pharm. Biol. 2019, 57, 161–168. [Google Scholar] [CrossRef]
- Valerio, M.; Liu, H.-B.; Heffner, R.; Zivadinov, R.; Ramanathan, M.; Weinstock-Guttman, B.; Awad, A.B. Phytosterols ameliorate clinical manifestations and inflammation in experimental autoimmune encephalomyelitis. Inflamm. Res. 2010, 60, 457–465. [Google Scholar] [CrossRef]
- Dierckx, T.; Bogie, J.F.; Hendriks, J.J. The Impact of Phytosterols on the Healthy and Diseased Brain. Curr. Med. Chem. 2019, 26, 6750–6765. [Google Scholar] [CrossRef]
- Saher, G.; Rudolphi, F.; Corthals, K.; Ruhwedel, T.; Schmidt, K.-F.; Löwel, S.; Dibaj, P.; Barrette, B.; Möbius, W.; Nave, K.-A. Therapy of Pelizaeus-Merzbacher disease in mice by feeding a cholesterol-enriched diet. Nat. Med. 2012, 18, 1130–1135. [Google Scholar] [CrossRef]
- Miron, V.E.; Zehntner, S.P.; Kuhlmann, T.; Ludwin, S.K.; Owens, T.; Kennedy, T.E.; Bedell, B.J.; Antel, J.P. Statin Therapy Inhibits Remyelination in the Central Nervous System. Am. J. Pathol. 2009, 174, 1880–1890. [Google Scholar] [CrossRef]
- Eroli, F.; Johnell, K.; Latorre Leal, M.; Adamo, C.; Hilmer, S.; Wastesson, J.W. Chronic polypharmacy impairs explorative be-havior and reduces synaptic functions in young adult mice. Aging (Albany NY) 2020, 12, 10147–10161. [Google Scholar] [CrossRef]
- Treuter, E. New wrestling rules of anti-inflammatory transrepression by oxysterol receptor LXR revealed. Cell Res. 2011, 21, 711–714. [Google Scholar] [CrossRef][Green Version]
- Shchelkunova, T.A.; Morozov, I.A.; Rubtsov, P.M.; Bobryshev, Y.V.; Sobenin, I.A.; Orekhov, A.N. Lipid regulators during atherogenesis: Expression of LXR, PPAR, and SREBP mRNA in the human aorta. PLoS ONE 2013, 8, e63374. [Google Scholar] [CrossRef]
- Siriwattanasatorn, M.; Itharat, A.; Thongdeeying, P.; Ooraikul, B. In Vitro Wound Healing Activities of Three Most Commonly Used Thai Medicinal Plants and Their Three Markers. Evid. Based Complement. Altern. Med. 2020, 2020, 1–11. [Google Scholar] [CrossRef]
- Blaauw, B.; Canato, M.; Agatea, L.; Toniolo, L.; Mammucari, C.; Masiero, E. Inducible activation of Akt increases skeletal muscle mass and force without satellite cell activation. FASEB J. 2009, 23, 3896–3905. [Google Scholar] [CrossRef]
- Parr, M.K.; Botrè, F.; Naß, A.; Hengevoss, J.; Diel, P.; Wolber, G. Ecdysteroids: A novel class of anabolic agents? Biol. Sport 2015, 32, 169–173. [Google Scholar] [CrossRef]
- Tóth, N.; Szabó, A.; Kacsala, P.; Héger, J.; Zádor, E. 20-Hydroxyecdysone increases fiber size in a muscle-specific fashion in rat. Phytomedicine 2008, 15, 691–698. [Google Scholar] [CrossRef]
- Bathori, M.; Toth, N.; Hunyadi, A.; Marki, A.; Zador, E. Phytoecdysteroids and Anabolic-Androgenic Steroids—Structure and Effects on Humans. Curr. Med. Chem. 2008, 15, 75–91. [Google Scholar] [CrossRef] [PubMed]
- Csábi, J.; Rafai, T.; Hunyadi, A.; Zádor, E. Poststerone increases muscle fibre size partly similar to its metabolically parent compound, 20-hydroxyecdysone. Fitoterapia 2019, 134, 459–464. [Google Scholar] [CrossRef] [PubMed]
- Dai, W.; Zhang, H.; Zhong, Z.A.; Jiang, L.; Chen, H.; Lay, Y.-A.E.; Kot, A.; Ritchie, R.O.; Lane, N.E.; Yao, W. β-Ecdysone Augments Peak Bone Mass in Mice of Both Sexes. Clin. Orthop. Relat. Res. 2015, 473, 2495–2504. [Google Scholar] [CrossRef] [PubMed]
- Kapur, P.; Wuttke, W.; Jarry, H.; Seidlova-Wuttke, D. Beneficial effects of β-Ecdysone on the joint, epiphyseal cartilage tissue and trabecular bone in ovariectomized rats. Phytomedicine 2010, 17, 350–355. [Google Scholar] [CrossRef]
- Seidlova-Wuttke, D.; Christel, D.; Kapur, P.; Nguyen, B.T.; Jarry, H.; Wuttke, W. Beta-ecdysone has bone protective but no estro-genic effects in ovariectomized rats. Phytomedicine 2010, 17, 884–889. [Google Scholar] [CrossRef]
- Zhao, X.-F. G protein-coupled receptors function as cell membrane receptors for the steroid hormone 20-hydroxyecdysone. Cell Commun. Signal. 2020, 18, 1–9. [Google Scholar] [CrossRef]
- Kicman, A.T.; Gower, D.B. Anabolic steroids in sport: Biochemical, clinical and analytical perspectives. Ann. Clin. Biochem. Int. J. Lab. Med. 2003, 40, 321–356. [Google Scholar] [CrossRef]
- Mareck, U.; Geyer, H.; Opfermann, G.; Thevis, M.; Schänzer, W. Factors influencing the steroid profile in doping control analysis. J. Mass Spectrom. 2008, 43, 877–891. [Google Scholar] [CrossRef]
- Rozner, S.; Garti, N. The activity and absorption relationship of cholesterol and phytosterols. Colloids Surfaces A Physicochem. Eng. Asp. 2006, 282–283, 435–456. [Google Scholar] [CrossRef]
- Fernández de las Heras, L.; van der Geize, R.; Drzyzga, O.; Perera, J.; María Navarro Llorens, J. Molecular characterization of three 3-ketosteroid-Δ1-dehydrogenase isoenzymes of Rhodococcus ruber strain Chol-4. J. Steroid Biochem. Mol. Biol. 2012, 132, 271–281. [Google Scholar] [CrossRef]
- Guevara, G.; Heras, L.F.D.L.; Perera, J.; Llorens, J.M.N. Functional differentiation of 3-ketosteroid Δ(1)-dehydrogenase isozymes in Rhodococcus ruber strain Chol-4. Microb. Cell Factories 2017, 16, 42. [Google Scholar] [CrossRef]
- van der Geize, R.; Hessels, G.I.; Dijkhuizen, L. Molecular and functional characterization of the kstD2 gene of Rhodococcus erythropolis SQ1 encoding a second 3-ketosteroid Delta(1)-dehydrogenase isoenzyme. Microbiology 2002, 148 (Pt 10), 3285–3292. [Google Scholar] [CrossRef]
- Rohman, A.; van Oosterwijk, N.; Thunnissen, A.-M.W.H.; Dijkstra, B.W. Crystal structure and site-directed mutagenesis of 3-ketosteroid Δ1-dehydrogenase from Rhodococcus erythropolis SQ1 explain its catalytic mechanism. J. Biol. Chem. 2013, 288, 35559–35568. [Google Scholar] [CrossRef]
- Wang, X.; Feng, J.; Zhang, D.; Wu, Q.; Zhu, D.; Ma, Y. Characterization of new recombinant 3-ketosteroid-Δ1-dehydrogenases for the biotransformation of steroids. Appl. Microbiol. Biotechnol. 2017, 101, 6049–6060. [Google Scholar] [CrossRef] [PubMed]
- Donova, M.V.; Dovbnya, D.V.; Sukhodolskaya, G.V.; Khomutov, S.M.; Nikolayeva, V.M.; Kwon, I.; Han, K. Microbial conversion of sterol-containing soybean oil production waste. J. Chem. Technol. Biotechnol. 2004, 80, 55–60. [Google Scholar] [CrossRef]
- Malaviya, A.; Gomes, J. Androstenedione production by biotransformation of phytosterols. Bioresour. Technol. 2008, 99, 6725–6737. [Google Scholar] [CrossRef]
- Turck, D.; Castenmiller, J.; De Henauw, S.; Hirsch-Ernst, K.I.; Kearney, J.; Maciuk, A. Safety of the extension of use of plant sterol esters as a novel food pursuant to Regulation (EU) 2015/2283. EFSA J. Eur. Food Saf. Auth. 2020, 18, e06135. [Google Scholar]
- Lobato, L.P.; Pereira, A.E.I.C.; Lazaretti, M.M.; Barbosa, D.S.; Carreira, C.M.; Mandarino, J.M.G.; Grossmann, M.V.E. Snack bars with high soy protein and isoflavone content for use in diets to control dyslipidaemia. Int. J. Food Sci. Nutr. 2012, 63, 49–58. [Google Scholar] [CrossRef]
- Polagruto, J.A.; Wang-Polagruto, J.F.; Braun, M.M.; Lee, L.; Kwik-Uribe, C.; Keen, C.L. Cocoa Flavanol-Enriched Snack Bars Containing Phytosterols Effectively Lower Total and Low-Density Lipoprotein Cholesterol Levels. J. Am. Diet. Assoc. 2006, 106, 1804–1813. [Google Scholar] [CrossRef]
- Kristensen, M.; Jensen, M.; Kudsk, J.; Henriksen, M.; Mølgaard, C. Short-term effects on bone turnover of replacing milk with cola beverages: A 10-day interventional study in young men. Osteoporos. Int. 2005, 16, 1803–1808. [Google Scholar] [CrossRef]
- Beveridge, T.H.J.; Li, T.S.C.; Drover, J.C.G. Phytosterol content in American ginseng seed oil. J. Agric. Food Chem. 2002, 50, 744–750. [Google Scholar] [CrossRef]
- Olkkonen, V.M.; Gylling, H.; Ikonen, E. [Plant sterols, cholesterol precursors and oxysterols: Small amounts, big effects]. Duodecim; Laaketieteellinen Aikakauskirja 2015, 131, 235–241. [Google Scholar]
- Prager, N.; Bickett, K.; French, N.; Marcovici, G. A randomized, double-blind, placebo-controlled trial to determine the effective-ness of botanically derived inhibitors of 5-alpha-reductase in the treatment of androgenetic alopecia. J. Altern. Complementary Med. 2002, 8, 143–152. [Google Scholar] [CrossRef]
- Bain, B.J.; Chakravorty, S. Phytosterolemia. Am. J. Hematol. 2016, 91, 643. [Google Scholar] [CrossRef] [PubMed]
- Mignarri, A.; Magni, A.; Del Puppo, M.; Gallus, G.N.; Björkhem, I.; Federico, A. Evaluation of cholesterol metabolism in cere-brotendinous xanthomatosis. J. Inherit. Metab. Dis. 2016, 39, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Nieminen, P.; Mustonen, A.-M.; Lindström-Seppä, P.; Kärkkäinen, V.; Mussalo-Rauhamaa, H.; Kukkonen, J.V.K. Phytosterols affect endocrinology and metabolism of the field vole (Microtus agrestis). Exp. Biol. Med. 2003, 228, 188–193. [Google Scholar] [CrossRef]
- Janeczko, A.; Skoczowski, A. Mammalian sex hormones in plants. Folia Histochem. Cytobiol. 2005, 43, 71–79. [Google Scholar]
- Finsterbusch, A.; Lindemann, P.; Grimm, R.; Eckerskorn, C.; Luckner, M. Delta(5)-3beta-hydroxysteroid dehydrogenase from Dig-italis lanata Ehrh—A multifunctional enzyme in steroid metabolism? Planta 1999, 209, 478–486. [Google Scholar] [CrossRef]
- Bonner, J.; Heftmann, E.; Zeevaart, J.A.D. Suppression of Floral Induction by Inhibitors of Steroid Biosynthesis. Plant Physiol. 1963, 38, 81–88. [Google Scholar] [CrossRef]
- Janeczko, A.; Filek, W. Stimulation of generative development in partly vernalized winter wheat by animal sex hormones. Acta Physiol. Plant. 2002, 24, 291–295. [Google Scholar] [CrossRef]
- Milanesi, L.; Boland, R. Presence of estrogen receptor (ER)-like proteins and endogenous ligands for ER in solanaceae. Plant Sci. 2004, 166, 397–404. [Google Scholar] [CrossRef]
- Speranza, A. Into the world of steroids: A biochemical “keep in touch” in plants and animals. Plant Signal Behav. 2010, 5, 940–943. [Google Scholar] [CrossRef]
- Gauthaman, K.; Ganesan, A.P. The hormonal effects of Tribulus terrestris and its role in the management of male erectile dysfunction—An evaluation using primates, rabbit and rat. Phytomedicine 2008, 15, 44–54. [Google Scholar] [CrossRef]
- Ma, Y.; Guo, Z.; Wang, X. Tribulus terrestris extracts alleviate muscle damage and promote anaerobic performance of trained male boxers and its mechanisms: Roles of androgen, IGF-1, and IGF binding protein-3. J. Sport Health Sci. 2017, 6, 474–481. [Google Scholar] [CrossRef]
- Simerský, R.; Novák, O.; Morris, D.A.; Pouzar, V.; Strnad, M. Identification and Quantification of Several Mammalian Steroid Hormones in Plants by UPLC-MS/MS. J. Plant Growth Regul. 2009, 28, 125–136. [Google Scholar] [CrossRef]
- Šaden-Krehula, M.; Tajić, M.; Kolbah, D. Testosterone, epitestosterone and androstenedione in the pollen of scotch pine P. silvestris L. Cell. Mol. Life Sci. 1971, 27, 108–109. [Google Scholar] [CrossRef]
- Hartmann, S.; Lacorn, M.; Steinhart, H. Natural occurrence of steroid hormones in food. Food Chem. 1998, 62, 7–20. [Google Scholar] [CrossRef]
- Hirotani, M.; Furuya, T. Biotransformation of testosterone and other androgens by suspension cultures of Nicotiana tabacum “Bright yellow”. Phytochemistry 1974, 13, 2135–2142. [Google Scholar] [CrossRef]
- Lindemann, P. Steroidogenesis in plants—Biosynthesis and conversions of progesterone and other pregnane derivatives. Steroids 2015, 103, 145–152. [Google Scholar] [CrossRef] [PubMed]
- Janeczko, A.; Budziszewska, B.; Skoczowski, A.; Dybała, M. Specific binding sites for progesterone and 17beta-estradiol in cells of Triticum aestivum L. Acta Biochim. Pol. 2008, 55, 707–711. [Google Scholar] [CrossRef] [PubMed]
- Janeczko, A.; Oklestkova, J.; Siwek, A.; Dziurka, M.; Pociecha, E.; Kocurek, M.; Novak, O. Endogenous progesterone and its cellular binding sites in wheat exposed to drought stress. J. Steroid Biochem. Mol. Biol. 2013, 138, 384–394. [Google Scholar] [CrossRef]
- Geyer, H.; Parr, M.K.; Koehler, K.; Mareck, U.; Schänzer, W.; Thevis, M. Nutritional supplements cross-contaminated and faked with doping substances. J. Mass Spectrom. 2008, 43, 892–902. [Google Scholar] [CrossRef]
- Walpurgis, K.; Thomas, A.; Geyer, H.; Mareck, U.; Thevis, M. Dietary Supplement and Food Contaminations and Their Implica-tions for Doping Controls. Foods 2020, 9, 1012. [Google Scholar] [CrossRef]
- Hamid, H.; Eskicioglu, C. Fate of estrogenic hormones in wastewater and sludge treatment: A review of properties and ana-lytical detection techniques in sludge matrix. Water Res. 2012, 46, 5813–5833. [Google Scholar] [CrossRef] [PubMed]
- Huang, B.; Wang, B.; Ren, D.; Jin, W.; Liu, J.; Peng, J. Occurrence, removal and bioaccumulation of steroid estrogens in Dian-chi Lake catchment, China. Environ. Int. 2013, 59, 262–273. [Google Scholar] [CrossRef] [PubMed]
- Grill, G.; Li, J.; Khan, U.; Zhong, Y.; Lehner, B.; Nicell, J.; Ariwi, J. Estimating the eco-toxicological risk of estrogens in China’s rivers using a high-resolution contaminant fate model. Water Res. 2018, 145, 707–720. [Google Scholar] [CrossRef] [PubMed]
- Stavreva, D.A.; George, A.A.; Klausmeyer, P.; Varticovski, L.; Sack, D.; Voss, T.C. Prevalent glucocorticoid and androgen activ-ity in US water sources. Sci. Rep. 2012, 2, 937. [Google Scholar] [CrossRef]
- Mnif, W.; Dagnino, S.; Escande, A.; Pillon, A.; Fenet, H.; Gomez, E. Biological analysis of endocrine-disrupting compounds in Tunisian sewage treatment plants. Arch. Environ. Contam. Toxicol. 2010, 59, 1–12. [Google Scholar] [CrossRef]
- Tan, B.L.; Hawker, D.W.; Müller, J.F.; Leusch, F.D.; Tremblay, L.A.; Chapman, H.F. Comprehensive study of endocrine disrupting compounds using grab and passive sampling at selected wastewater treatment plants in South East Queensland, Australia. Environ. Int. 2007, 33, 654–669. [Google Scholar] [CrossRef]
- Pi, N.; Ng, J.; Kelly, B. Bioaccumulation of pharmaceutically active compounds and endocrine disrupting chemicals in aquatic macrophytes: Results of hydroponic experiments with Echinodorus horemanii and Eichhornia crassipes. Sci. Total. Environ. 2017, 601–602, 812–820. [Google Scholar] [CrossRef]
- Jung, C.; Park, J.; Lim, K.H.; Park, S.; Heo, J.; Her, N. Adsorption of selected endocrine disrupting compounds and pharma-ceuticals on activated biochars. J. Hazard. Mater. 2013, 263 Pt 2, 702–710. [Google Scholar] [CrossRef]
- Adeel, M.; Zain, M.; Fahad, S.; Rizwan, M.; Ameen, A.; Yi, H.; Baluch, M.A.; Lee, J.Y.; Rui, Y. Natural and synthetic estrogens in leafy vegetable and their risk associated to human health. Environ. Sci. Pollut. Res. 2018, 25, 36712–36723. [Google Scholar] [CrossRef]
- Adeel, M.; Song, X.; Wang, Y.; Francis, D.; Yang, Y. Environmental impact of estrogens on human, animal and plant life: A critical review. Environ. Int. 2017, 99, 107–119. [Google Scholar] [CrossRef]
- Su, C.; Cui, Y.; Liu, D.; Zhang, H.; Baninla, Y. Endocrine disrupting compounds, pharmaceuticals and personal care products in the aquatic environment of China: Which chemicals are the prioritized ones? Sci. Total Environ. 2020, 720, 137652. [Google Scholar] [CrossRef]
- Peng, F.-J.; Pan, C.-G.; Zhang, M.; Zhang, N.-S.; Windfeld, R.; Salvito, D.; Selck, H.; Brink, P.J.V.D.; Ying, G.-G. Occurrence and ecological risk assessment of emerging organic chemicals in urban rivers: Guangzhou as a case study in China. Sci. Total Environ. 2017, 589, 46–55. [Google Scholar] [CrossRef]
- González, A.; Kroll, K.J.; Silva-Sanchez, C.; Carriquiriborde, P.; Fernandino, J.I.; Denslow, N.D. Steroid hormones and estrogen-ic activity in the wastewater outfall and receiving waters of the Chascomús chained shallow lakes system (Argentina). Sci. Total Environ. 2020, 743, 140401. [Google Scholar] [CrossRef]
- Erdal, S.; Dumlupinar, R. Exogenously Treated Mammalian Sex Hormones Affect Inorganic Constituents of Plants. Biol. Trace Element Res. 2011, 143, 500–506. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type of Study | Characterization and Study Design | Purpose of Study | Observed Effect | Reference |
|---|---|---|---|---|
| Review study | Food enriched by plant sterols and stanols | Evaluation the contradictory effects of plant sterols on cholesterol lowering | Decrease of LDL-C by 10% | [33] |
| Meta-analysis | Data of clinical trials between January 1992 and April 2013 | Evaluation of plant sterols delivery way and matrix on cholesterol-lowering action | Decreasing of LDL-C by 12 mg/mL for 4–6 weeks; no difference between sterols supplements and food enriched by plant sterols | [38] |
| Randomized controlled clinical trial | Supplementation by sterols-enriched margarin | Effect of chronic intake of plant sterols on postprandial metabolism | Lowering of total cholesterol by 7.3%, LDL-C by 9.5% and LDL by 12.3% after intake of sterols-enriched margarine for 3 weeks | [53] |
| Randomized controlled trial | In vivo trial with both men (n = 6) and women (n = 6) for 5 weeks after consumption of phytosterols | Endogenous origin of boldenone and boldione under controlled phytosterol-enriched diet | Endogenous boldione produced between 0.75 and 1.73 ng/mL and strongly correlated with the consumption of plant sterols; in contrast, the endogenous origin of boldenone was not confirmed under the same condition | [68] |
| Randomized controlled clinical trial | The study on 28 subjects with hypertriglyceridemia (elevated fasting triacylglycerols) supplemented by stanols-enriched (2.5 g/day) margarine for 3 weeks | Effect of plant stanols supplementation on lipids metabolism | Plant stanol esters supplementation (margarines) lower triacylglycerol by 6.7% and LDL-C by 9.5% after 3 weeks intake | [69] |
| Clinical trial | Women with coronary artery disease consumed margarine without and with sitostanol (3 g/day) ester for 7 weeks | Effect of sitostanol-enriched diet on serum level cholesterol in postmenopausal women with myocardial infarction | Lowering of serum cholesterol by 13% and LDL-C by 20% for 12 weeks in home diet and by 11% and 16%, correspondingly, if combined with simvastatin | [54,67] |
| Comparative study; human subjects | The study on 144 subjects aged 30–80 years old | Age-related relation between some sterols and cognitive performance | The level of cholesterol, desmosterol, sitosterol and campesterol were not related to cognitive performance; however, serum levels of lathosterol and lanosterol inversely correlated with cognitive performance | [66] |
| Comparative study; animal model | Hypercholesterolemic (ApoE−/−) mouse with acute aseptic was fed with 2% plants sterols supplements | Investigation of immune modulating and stimulatory effects of plant sterols | Increased IL-6, IL-2 and IFN-γ secretion in spleen cells; potentiating T-helper response | [61] |
| Comparative study; animal model | Fibroblast and keratinocytes cell culture and mouse model of cutaneous wound healing | Effect of homobrassinolide on proliferative activity and migration of mouse fibroblasts and keratinocytes | A 2-fold increase rate of wound closure if mice received 10 µg topical brassinosteroids; suppressed ICAM-1 and TNF-α but weak effect on TGF-β; upregulation of Akt-1 phosphorylation in treated mice | [64,70] |
| Comparative study; animal model | Transgenic mice with disrupted BBB caused by lacking the PDGF-B retention motif | Metabolic circulation and sterols flux under the condition of disrupted blood-brain barrier in pericyte-deficient mice | A significant accumulation of campesterol and sitosterol in the brains but the degree of accumulation of sitosterol was lower; higher mRNA levels of HMG-CoA synthase. Significantly increased flux of cholesterol from circulation into brain | [71,72] |
| Comparative study; animal model | Mice deficient for ATP-binding cassette transporter G5 (abcg5)/G8 (abcg8) and deficient on ApoE | Association of plant sterols and cholesterol brain concentration with ApoE and transporters activities | Increased serum level of plant sterols (7–16-folds change); upregulation of ApoE mRNA level | [73] |
| Comparative study; animal model | LXRβ−/− and wild type study mice treated by β-sitosterol for 3 weeks at a dose of 42 mg/kg | Toxicity effect of β-sitosterol on neurons | Sever symptoms of paralysis and dopaminergic disfunction in transgenic mice; aggregation of ubiquitin in the cytoplasm of large motor neurons; increased level of 24-hydrocholesterol. Wild type mice were not affected | [74] |
| Comparative study; animal model | Loading of 7β-hydroxysitosterol in ApoE−/− mice for 28 days | Sterols and cholesterol absorption rate in plasma, liver and brain of transgenic ApoE-deficient mice | Concentration of 7β-hydroxysitosterol increased 65.8-folds and 21-folds in brain and plasma, correspondingly | [75] |
| Comparative study; animal model | European polecat (Mustela putorius) fed by phytosterols | Endocrine and metabolic effects of plant sterols | Increased plasma estradiol, TH and glucose-6-phosphatase activity, the plasms ghrelin level decreased; the total serum level of cholesterol was not affected | [76,77] |
| Prospective study; anima model | Examination of brain cortex and hippocampus in 6-, 12-, 18- and 24-month-old rats | Age-related changes of plasma and brain plants sterols concentrations | Reduction in lanosterol (by 28%), lathosterol (by 25%) and desmosterol (by 51%) concentrations at 24 months in the cortex and hippocampus that can be caused by the loss of synaptic plasticity | [65] |
| Research study; animal model | Murine colitis model fed by sterols-enriched food at a dose of 400 mg/kg per day | Effect of plant sterols mixture on gastrointestinal inflammation | Low level of leucocyte infiltration into the colon, preserved epithelial integrity; accelerated reparation of epithelial structure | [78] |
| Research study; cell culture | In vitro fibroblast L929 wound healing assays exposed to plant leaves ethanolic extract at a dose of 3 µg/mL | Investigation of wound healing effect of ethanolic extract of Wedelia trilobata (L.) leaves | Stimulatory effect; increased collagen content, activity against gram-positive bacteria, increased survivability by 85% after oxidative stress; induced by 70% migration rate | [79] |
| Research study; animal model | Rat model of Parkinson’s disease induced by β-sitosterol derivate for 16 weeks at a dose of 3 mg 5 days per a week | Effect of Beta-sitosterol beta-d-glucoside on dopaminergic neurons | ß-sitosterol may produce toxic glycosides and modulate lipids metabolism in neurons; induces the loss of dopaminergic neurons of the nigrostriatal pathway and results in akinesia and loss of locomotion | [80] |
| Research study; animal model | Male APPswePS1ΔE9 Alzheimer’s disease mice supplemented with 50% (w/w) pulverized dried Sargassum fusiforme diet | Effect of sterols on cognition ability through the activation of LXRβ receptors | Extracts of Sargassum fusiforme activates LXRβ at a dose 5 ug/mL and markedly decreased Aβ plaque load in the cortex (70%) and hippocampus (81%) | [81] |
| Research study; animal model | Animals with experimental autoimmune encephalomyelitis | Evaluation of motor skills performance and relationship between cholesterol and myelination in cuprizone model mouse fed by phytosterols diet | Declined expression of IFNγ, TNF and IL-17 and associated decrease of serum cholesterol for mice treated with 5% w/w dietary cholesterol | [82] |
| Research study; animal model | Type 2 diabetes mellitus rats | Effect of Aloe vera gel on expression level of PPAR targets | Activation of PPAR and decrease of Apoc3 in dose-dependent manner | [83] |
| Research study; animal model | Rats with non-alcoholic steatohepatitis daily administrated with Aloe vera extract (50 mg/kg) for 8 weeks | Effect of Aloe vera on liver enzymes and inflammation in non-alcoholic steatohepatitis model rats. | PPAR decreased; IL-18 and caspase-3 increased in rats fed with Aloe vera. Symptoms lobular inflammation and hepatocyte apoptosis were attenuated if animals treated by Aloe vera. | [84] |
| Research study; cell culture and animal model | Murine embryonic fibroblast cell line CCL-92 exposed to brassinosteroids and its synthetic analogues; topical application of synthetic brassinosteroids on mice after injury | Stimulating effect of homobrassinolide and its analogues on cell fibroblasts and keratinocytes proliferation and migration; and in vivo wound healing in animals | Brassinosteroids promotes cell migration into a wound zone with an efficacy of 30% at 5 μM after 12 h of incubation; treatment with 1–10 μM resulted in an increase in the proliferation of primary keratinocytes | [64] |
| Research study; cell culture | Rat L6 skeletal muscle cell line CRL-1458 treated by brassinosteroids analogues | Evaluation of anabolic effect on muscle cells through the selective activation of Akt by different brassinosteroids | Decrease in protein degradation by 20% after treatment with 10 uM of brassinosteroids in a dose-dependent manner | [85] |
| Research study; animal model | Mice (12 weeks old) strain C57BL/J6 were continually administrated by 20-hydroxyecdysone at a dose of 5 mg/kg/day | Anabolic effect of 20-hydroxyecdysone administration | Increase in the mass of the triceps brachii by 30%; however, no differentially expressed gene were observed compared to the control group (saline) after applying FDR. In summary, administration of 20-hydroxyecdysone did not demonstrate the pronounced anabolic effect | [86] |
| Research study; animal model | Adult male Wistar rats (10 weeks old) with tenotomy-induced muscle atrophy administrated 20-hydroxyecdystone at a dose of 5 mg/kg for 7 days | Evaluation of 20-hydroxyecdystone type-specific response in atrophy or injured muscles | Administration of 5 mg/kg of 20-hydroxyecdystone tended to alleviate tenotomy-induced reduction of muscle mass and attenuate by 63.1% tenotomy-induced ubiquitinated protein in soleus muscle, but had no effect on cross-sectional area in atrophied muscle | [87] |
| Research study; cell culture | Mouse skeletal muscle cell line C2C12 treated by 1 µM 20-hydroxyecdyson delivered in ethanol | Effect of 20-hydroxyecdyson on calcium influx and Akt activation | Treatment with 1 μM 20-hydroxyecdyson increased intracellular Ca2+ already in 35 sec and activated Akt more than 3-fold in 2 h; however, pretreatment with phospholipase C inhibitor abolished the effect of 20-hydroxyecdyson | [88] |
| Biotechnology and bioengineering | Mutants of Mycobacterium spp. | Investigation of transformation of plant sterols into androstenedione and androstadienedione in bacteria | Bacteria, growing on phytosterols, accumulate androstenedione and androstadienedione as the steroid intermediates during biotransformation of plant sterols after removing the side-chain and ring cleavage | [89] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kopylov, A.T.; Malsagova, K.A.; Stepanov, A.A.; Kaysheva, A.L. Diversity of Plant Sterols Metabolism: The Impact on Human Health, Sport, and Accumulation of Contaminating Sterols. Nutrients 2021, 13, 1623. https://doi.org/10.3390/nu13051623
Kopylov AT, Malsagova KA, Stepanov AA, Kaysheva AL. Diversity of Plant Sterols Metabolism: The Impact on Human Health, Sport, and Accumulation of Contaminating Sterols. Nutrients. 2021; 13(5):1623. https://doi.org/10.3390/nu13051623
Chicago/Turabian StyleKopylov, Arthur T., Kristina A. Malsagova, Alexander A. Stepanov, and Anna L. Kaysheva. 2021. "Diversity of Plant Sterols Metabolism: The Impact on Human Health, Sport, and Accumulation of Contaminating Sterols" Nutrients 13, no. 5: 1623. https://doi.org/10.3390/nu13051623
APA StyleKopylov, A. T., Malsagova, K. A., Stepanov, A. A., & Kaysheva, A. L. (2021). Diversity of Plant Sterols Metabolism: The Impact on Human Health, Sport, and Accumulation of Contaminating Sterols. Nutrients, 13(5), 1623. https://doi.org/10.3390/nu13051623