
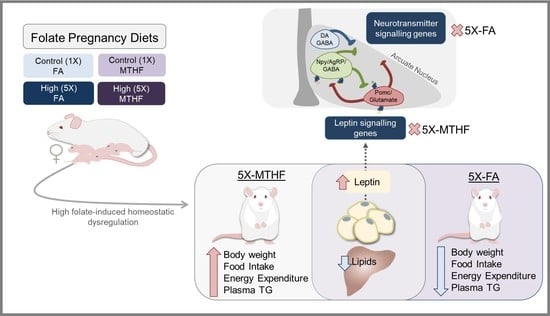
High Intakes of [6S]-5-Methyltetrahydrofolic Acid Compared with Folic Acid during Pregnancy Programs Central and Peripheral Mechanisms Favouring Increased Food Intake and Body Weight of Mature Female Offspring


 ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Diets
2.2. Phenotypic and Biochemical Analyses
2.3. Folate and Related 1-Carbon Metabolites
2.4. Energy Expenditure and Locomotor Activity
2.5. Brain Dissections and qRT-PCR
2.6. Statistical Analyses
3. Results
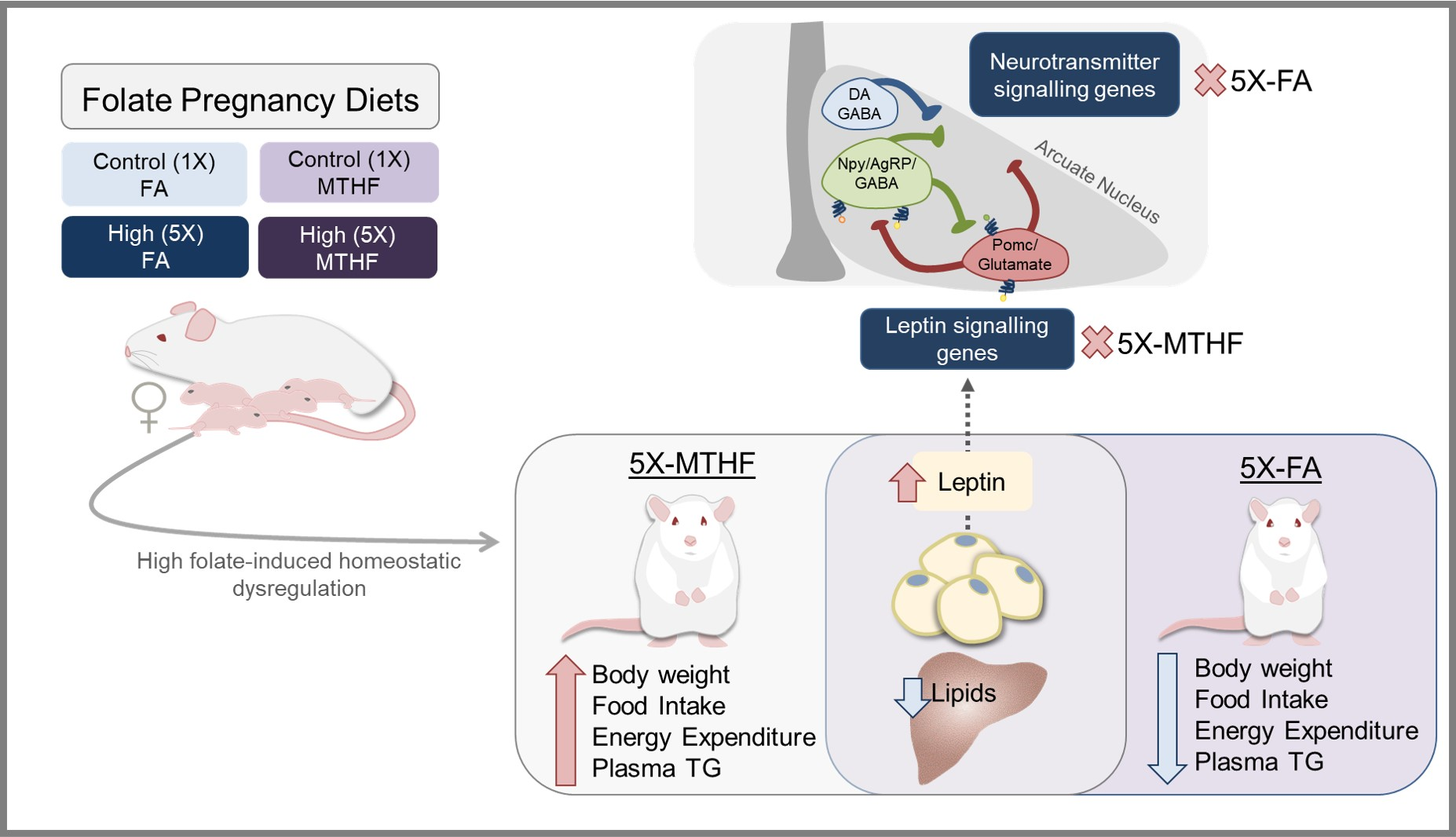
3.1. X-MTHF Offspring Gained more Weight and Ate more Food Than 5X-FA Offspring
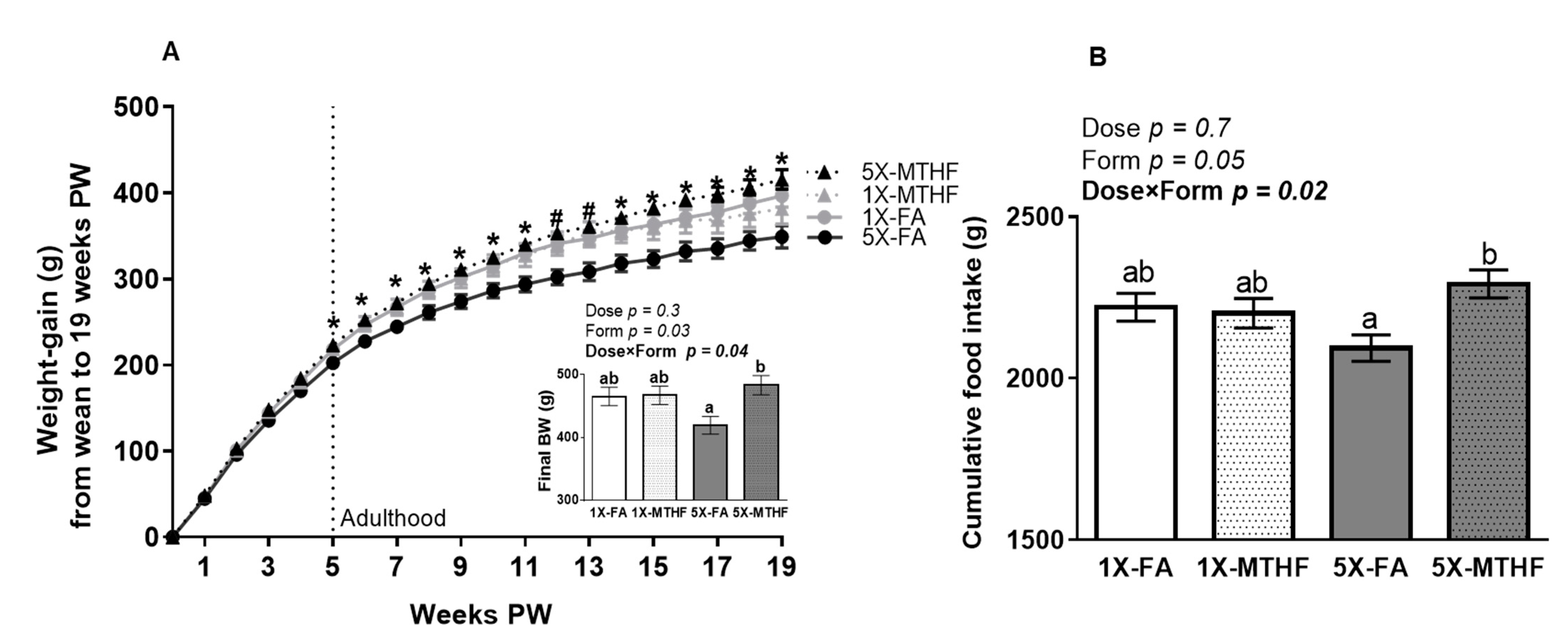
3.2. X offspring have Higher Plasma Leptin at Birth and at 19 Weeks Post-Weaning Than 1X Offspring
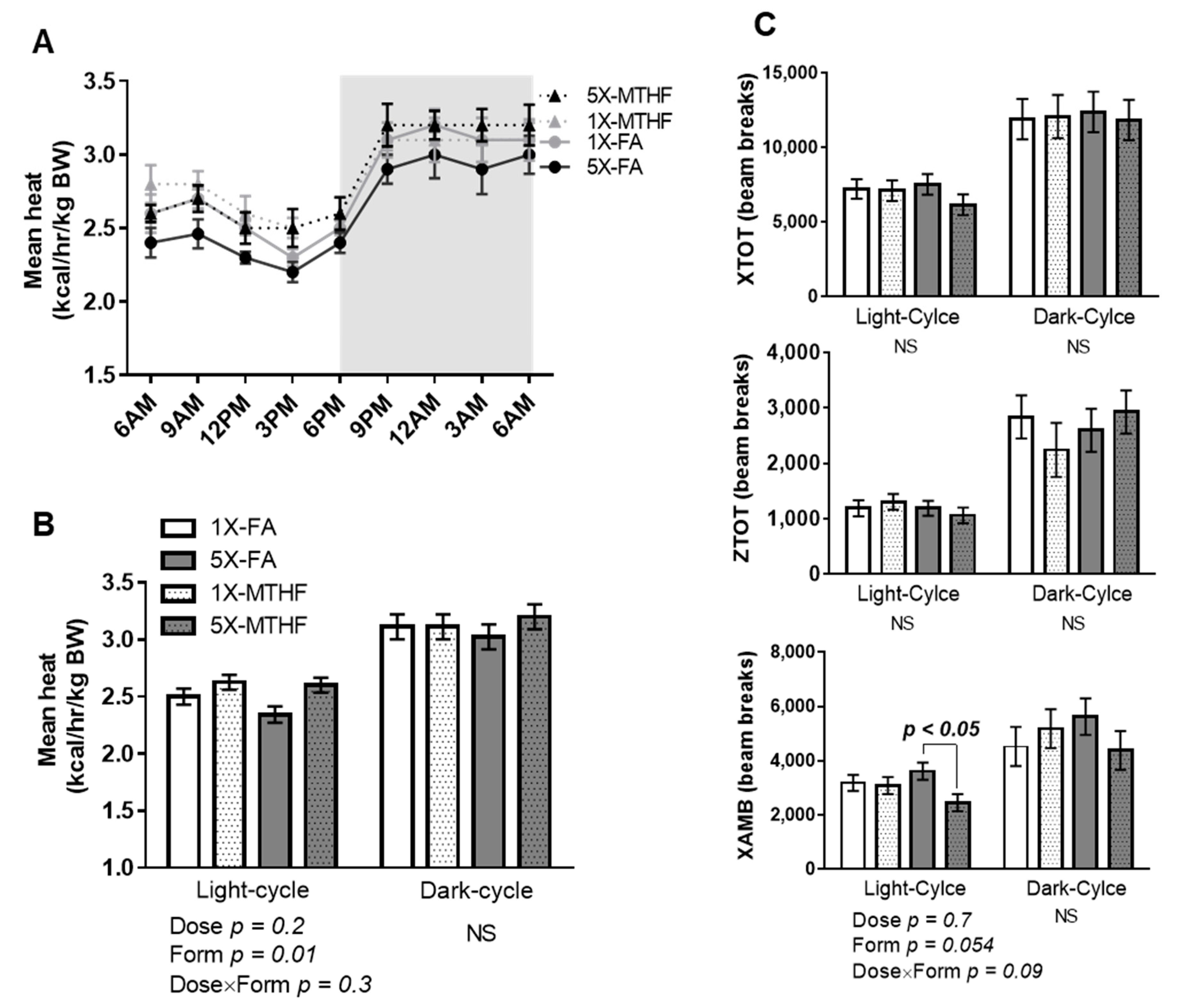
3.3. Post-Weaning Energy Expenditure and Locomotor Activity Are Affected by Gestational Folate Diets
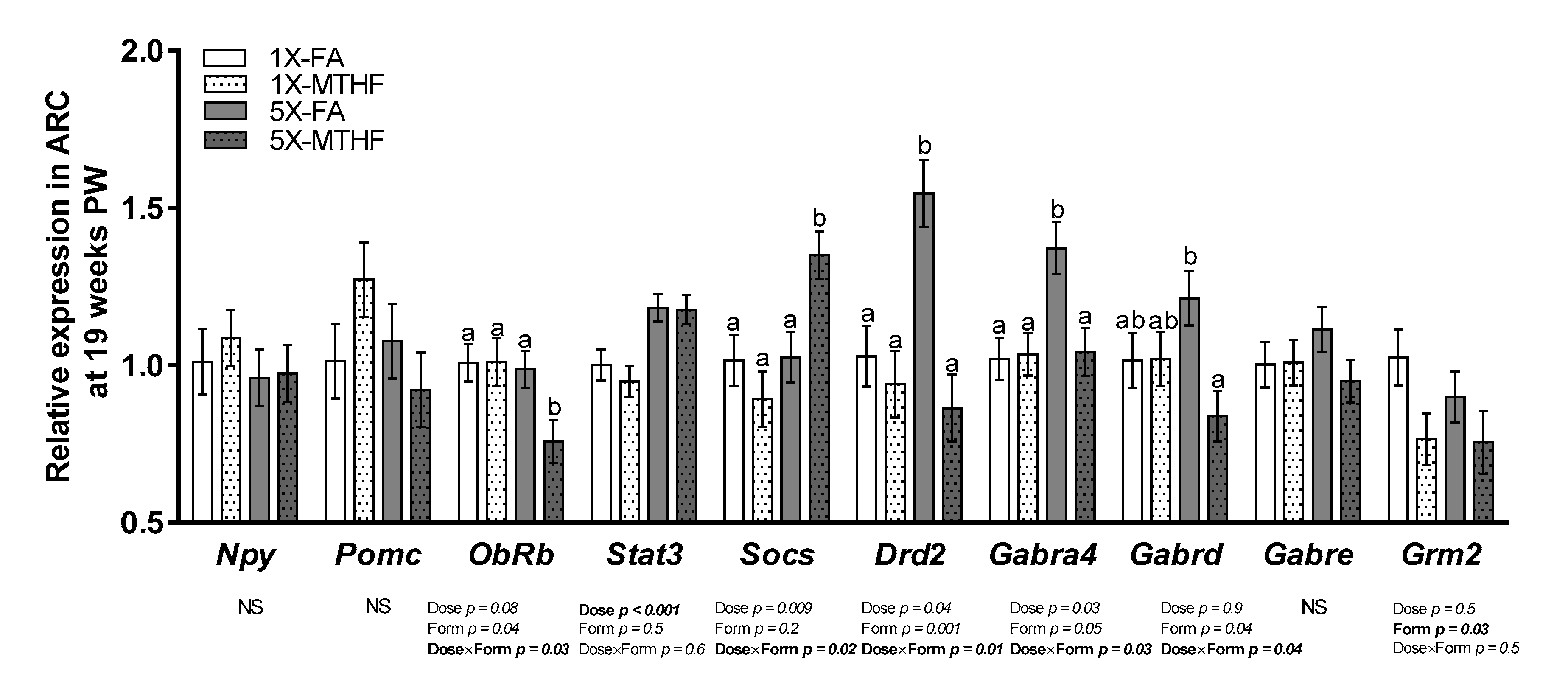
3.4. Post-Weaning Hypothalamic Energy Regulatory Genes Are Affected by Gestational Folate Diets
3.5. Plasma 5-MTHF and Related 1-Carbon Metabolites Are Affected by Folate Gestational Diets
4. Discussion
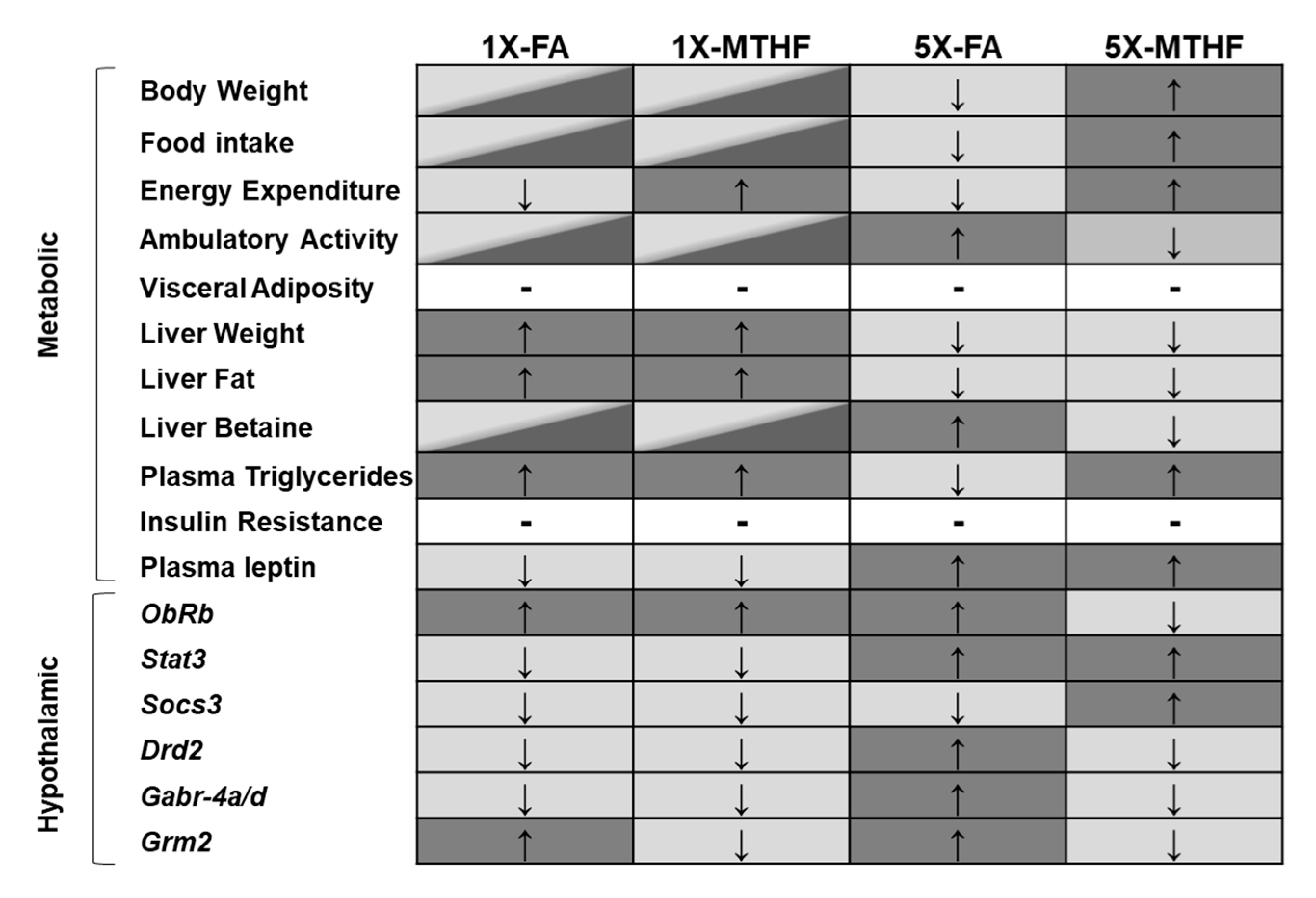
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Padmanabhan, V.; Cardoso, R.C.; Puttabyatappa, M. Developmental Programming, a Pathway to Disease. Endocrinology 2016, 157, 1328–1340. [Google Scholar] [CrossRef]
- Pannia, E.; Cho, C.E.; Kubant, R.; Sánchez-Hernández, D.; Huot, P.S.; Anderson, G.H. Role of maternal vitamins in programming health and chronic disease. Nutr. Rev. 2016, 74, 166–180. [Google Scholar] [CrossRef] [PubMed]
- Stover, P.J.; Field, M.S. Trafficking of intracellular folates. Adv. Nutr. 2011, 2, 325–331. [Google Scholar] [CrossRef]
- Christensen, K.E.; Mikael, L.G.; Leung, K.-Y.; Lévesque, N.; Deng, L.; Wu, Q.; Malysheva, O.V.; Best, A.; Caudill, M.A.; Greene, E.N.D.; et al. High folic acid consumption leads to pseudo-MTHFR deficiency, altered lipid metabolism, and liver injury in mice. Am. J. Clin. Nutr. 2015, 101, 646–658. [Google Scholar] [CrossRef] [PubMed]
- Mudryj, A.N.; de Groh, M.; Aukema, H.M.; Yu, N. Folate intakes from diet and supplements may place certain Canadians at risk for folic acid toxicity. Corrigendum. Br. J. Nutr. 2016, 116, 1995. [Google Scholar] [CrossRef] [PubMed]
- Bailey, R.L.; Pac, S.G.; Fulgoni, V.L.; Reidy, K.C.; Catalano, P.M. Estimation of Total Usual Dietary Intakes of Pregnant Women in the United States. JAMA Netw. Open 2019, 2, e195967. [Google Scholar] [CrossRef]
- Plumptre, L.; Masih, S.P.; Ly, A.; Aufreiter, S.; Sohn, K.-J.; Croxford, R.; Lausman, A.Y.; Berger, H.; O’Connor, D.L.; Kim, Y.-I. High concentrations of folate and unmetabolized folic acid in a cohort of pregnant Canadian women and umbilical cord blood. Am. J. Clin. Nutr. 2015, 102, 848–857. [Google Scholar] [CrossRef]
- Maruvada, P.; Stover, P.J.; Mason, J.B.; Bailey, R.L.; Davis, C.D.; Field, M.S.; Finnell, R.H.; Garza, C.; Green, R.; Gueant, J.-L.; et al. Knowledge gaps in understanding the metabolic and clinical effects of excess folates/folic acid: A summary, and perspectives, from an NIH workshop. Am. J. Clin. Nutr. 2020, 112, 1390–1403. [Google Scholar] [CrossRef]
- Naninck, E.F.G.; Stijger, P.C.; Brouwer-Brolsma, E.M. The Importance of Maternal Folate Status for Brain Development and Function of Offspring. Adv. Nutr. 2019, 10, 502–519. [Google Scholar] [CrossRef]
- Keating, E.; Correia-Branco, A.; Araújo, J.R.; Meireles, M.; Fernandes, R.; Guardão, L.; Guimarães, J.T.; Martel, F.; Calhau, C. Excess perigestational folic acid exposure induces metabolic dysfunction in post-natal life. J. Endocrinol. 2015, 224, 245–259. [Google Scholar] [CrossRef]
- Giudicelli, F.; Brabant, A.-L.; Grit, I.; Parnet, P.; Amarger, V. Excess of Methyl Donor in the Perinatal Period Reduces Postnatal Leptin Secretion in Rat and Interacts with the Effect of Protein Content in Diet. PLoS ONE 2013, 8, e68268. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Goh, C.E.; Demmer, R.T.; Whitcomb, B.W.; Du, P.; Liu, Z. Association between Serum Folate and Insulin Resistance among U.S. Nondiabetic Adults. Sci. Rep. 2017, 7, 9187. [Google Scholar] [CrossRef]
- Narin, F.; Atabek, M.E.; Karakukcu, M.; Narin, N.; Kurtoglu, S.; Gumus, H.; Çoksevim, B.; Erez, R. The association of plasma homocysteine levels with serum leptin and apolipoprotein B levels in childhood obesity. Ann. Saudi Med. 2005, 25, 209–214. [Google Scholar] [CrossRef] [PubMed]
- Cho, C.E.; Pannia, E.; Huot, P.S.P.; Sánchez-Hernández, D.; Kubant, R.; Dodington, D.W.; Ward, W.E.; Bazinet, R.P.; Anderson, G.H. Methyl vitamins contribute to obesogenic effects of a high multivitamin gestational diet and epigenetic alterations in hypothalamic feeding pathways in Wistar rat offspring. Mol. Nutr. Food Res. 2014, 59, 476–489. [Google Scholar] [CrossRef] [PubMed]
- Huot, P.S.; Ly, A.; Szeto, I.M.; Reza-López, S.A.; Cho, D.; Kim, Y.-I.; Anderson, G.H. Maternal and postweaning folic acid supplementation interact to influence body weight, insulin resistance, and food intake regulatory gene expression in rat offspring in a sex-specific manner. Appl. Physiol. Nutr. Metab. 2016, 41, 411–420. [Google Scholar] [CrossRef]
- Yang, N.V.; Pannia, E.; Chatterjee, D.; Kubant, R.; Ho, M.; Hammoud, R.; Pausova, Z.; Anderson, G.H. Gestational folic acid content alters the development and function of hypothalamic food intake regulating neurons in Wistar rat offspring post-weaning. Nutr. Neurosci. 2018, 23, 149–160. [Google Scholar] [CrossRef] [PubMed]
- Xie, K.; Fu, Z.; Li, H.; Gu, X.; Cai, Z.; Xu, P.; Cui, X.; You, L.; Wang, X.; Zhu, L.; et al. High folate intake contributes to the risk of large for gestational age birth and obesity in male offspring. J. Cell. Physiol. 2018, 233, 9383–9389. [Google Scholar] [CrossRef]
- Huang, Y.; He, Y.; Sun, X.; He, Y.; Li, Y.; Sun, C. Maternal High Folic Acid Supplement Promotes Glucose Intolerance and Insulin Resistance in Male Mouse Offspring Fed a High-Fat Diet. Int. J. Mol. Sci. 2014, 15, 6298–6313. [Google Scholar] [CrossRef]
- Huot, P.S.P.; Dodington, D.W.; Mollard, R.C.; Reza-López, S.A.; Sanchez-Hernandez, D.; Cho, C.E.; Kuk, J.; Ward, W.E.; Anderson, G.H. High Folic Acid Intake during Pregnancy Lowers Body Weight and Reduces Femoral Area and Strength in Female Rat Offspring. J. Osteoporos. 2013, 2013, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Sie, K.K.Y.; Li, J.; Ly, A.; Sohn, K.-J.; Croxford, R.; Kim, Y.-I. Effect of maternal and postweaning folic acid supplementation on global and gene-specific DNA methylation in the liver of the rat offspring. Mol. Nutr. Food Res. 2013, 57, 677–685. [Google Scholar] [CrossRef]
- Lamers, Y.; Macfarlane, A.J.; O’Connor, D.L.; Fontaine-Bisson, B. Periconceptional intake of folic acid among low-risk women in Canada: Summary of a workshop aiming to align prenatal folic acid supplement composition with current expert guidelines. Am. J. Clin. Nutr. 2018, 108, 1357–1368. [Google Scholar] [CrossRef] [PubMed]
- Saldanha, L.G.; Dwyer, J.T.; Haggans, C.J.; Mills, J.L.; Potischman, N. Perspective: Time to Resolve Confusion on Folate Amounts, Units, and Forms in Prenatal Supplements. Adv. Nutr. 2020, 11, 753–759. [Google Scholar] [CrossRef] [PubMed]
- US Department of Health and Human Services; Food and Drug Administration; Center for Food Safety and Applied Nutrition. Guidance for Industry: Converting Units of Measure for Folate, Niacin, And Vitamins A, D, and E on the Nutrition and Supplement Facts Labels. Available online: https://www.fda.gov/media/129863/download (accessed on 19 March 2021).
- Pannia, E.; Hammoud, R.; Simonian, R.; Arning, E.; Ashcraft, P.; Wasek, B.; Bottiglieri, T.; Pausova, Z.; Kubant, R.; Anderson, G.H. [6S]-5-Methyltetrahydrofolic Acid and Folic Acid Pregnancy Diets Differentially Program Metabolic Phenotype and Hypothalamic Gene Expression of Wistar Rat Dams Post-Birth. Nutrition 2020, 13, 48. [Google Scholar] [CrossRef]
- Delgado, T.C. Glutamate and GABA in Appetite Regulation. Front. Endocrinol. 2013, 4, 103. [Google Scholar] [CrossRef] [PubMed]
- Freeman, M.P.; Savella, G.M.; Church, T.R.; Góez-Mogollón, L.; Sosinsky, A.Z.; Noe, O.B.; Kaimal, A.; Cohen, L.S. A prenatal supplement with methylfolate for the treatment and prevention of depression in women trying to conceive and during pregnancy. Ann. Clin. Psychiatry 2019, 31, 4–16. [Google Scholar]
- Jain, R.; Manning, S.; Cutler, A.J. Good, better, best: Clinical scenarios for the use of L-methylfolate in patients with MDD. CNS Spectr. 2019, 25, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Battista, M.-C.; Hivert, M.-F.; Duval, K.; Baillargeon, J.-P. Intergenerational Cycle of Obesity and Diabetes: How Can We Reduce the Burdens of These Conditions on the Health of Future Generations? Exp. Diabetes Res. 2011, 2011, 596060. [Google Scholar] [CrossRef]
- Lamers, Y.; Prinz-Langenohl, R.; Moser, R.; Pietrzik, K. Supplementation with [6S]-5-methyltetrahydrofolate or folic acid equally reduces plasma total homocysteine concentrations in healthy women. Am. J. Clin. Nutr. 2004, 79, 473–478. [Google Scholar] [CrossRef]
- Lamers, Y.; Prinz-Langenohl, R.; Brämswig, S.; Pietrzik, K. Red blood cell folate concentrations increase more after supplementation with [6 S]-5-methyltetrahydrofolate than with folic acid in women of childbearing age. Am. J. Clin. Nutr. 2006, 84, 156–161. [Google Scholar] [CrossRef]
- Council, N.R. Nutrient requirements of the laboratory rat. In Nutrient Requirements of Laboratory Animals, 4th ed.; National Academy Press: Washington, DC, USA, 1995; pp. 11–79. [Google Scholar]
- Folch, J.; Lees, M.; Sloane-Stanley, G.H. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Arning, E.; Bottiglieri, T. Quantitation of S-Adenosylmethionine and S-Adenosylhomocysteine in Plasma Using Liquid Chromatography-Electrospray Tandem Mass Spectrometry. Adv. Struct. Saf. Stud. 2016, 1378, 255–262. [Google Scholar] [CrossRef]
- Nandania, J.; Kokkonen, M.; Euro, L.; Velagapudi, V. Simultaneous measurement of folate cycle intermediates in different biological matrices using liquid chromatography–tandem mass spectrometry. J. Chromatogr. B 2018, 1092, 168–178. [Google Scholar] [CrossRef]
- Rooney, M.; Bottiglieri, T.; Wasek-Patterson, B.; McMahon, A.; Hughes, C.F.; McCann, A.; Horigan, G.; Strain, J.; McNulty, H.; Ward, M. Impact of the MTHFR C677T polymorphism on one-carbon metabolites: Evidence from a randomised trial of riboflavin supplementation. Biochimie 2020, 173, 91–99. [Google Scholar] [CrossRef]
- Ducros, V.; Belva-Besnet, H.; Casetta, B.; Favier, A. A robust liquid chromatography tandem mass spectrometry method for total plasma homocysteine determination in clinical practice. Clin. Chem. Lab. Med. 2006, 44, 987–990. [Google Scholar] [CrossRef] [PubMed]
- Paxinos, G.; Watson, C.R.; Emson, P.C. AChE-stained horizontal sections of the rat brain in stereotaxic coordinates. J. Neurosci. Methods 1980, 3, 129–149. [Google Scholar] [CrossRef]
- Herrera, E.; Lasunción, M.; Huerta, L.; Martín-Hidalgo, A. Plasma leptin levels in rat mother and offspring during pregnancy and lactation. Biol. Neonate 2000, 78, 315–320. [Google Scholar] [CrossRef] [PubMed]
- Simonds, S.E.; Cowley, M.A.; Enriori, P.J. Leptin increasing sympathetic nerve outflow in obesity. Adipocyte 2012, 1, 177–181. [Google Scholar] [CrossRef] [PubMed]
- Licholai, J.A.; Nguyen, K.P.; Fobbs, W.C.; Schuster, C.J.; Ali, M.A.; Kravitz, A.V. Why Do Mice Overeat High-Fat Diets? How High-Fat Diet Alters the Regulation of Daily Caloric Intake in Mice. Obesity 2018, 26, 1026–1033. [Google Scholar] [CrossRef]
- Bjursell, M.; Gerdin, A.-K.; Lelliott, C.J.; Egecioglu, E.; Elmgren, A.; Törnell, J.; Oscarsson, J.; Bohlooly-Y, M. Acutely reduced locomotor activity is a major contributor to Western diet-induced obesity in mice. Am. J. Physiol. Metab. 2008, 294, E251–E260. [Google Scholar] [CrossRef]
- Cosín-Tomás, M.; Luan, Y.; Leclerc, D.; Malysheva, O.V.; Lauzon, N.; Bahous, R.H.; Christensen, K.E.; Caudill, M.A.; Rozen, R. Moderate Folic Acid Supplementation in Pregnant Mice Results in Behavioral Alterations in Offspring with Sex-Specific Changes in Methyl Metabolism. Nutrients 2020, 12, 1716. [Google Scholar] [CrossRef]
- Chu, D.; Li, L.; Jiang, Y.; Tan, J.; Ji, J.; Zhang, Y.; Jin, N.; Liu, F. Excess Folic Acid Supplementation Before and During Pregnancy and Lactation Activates Fos Gene Expression and Alters Behaviors in Male Mouse Offspring. Front. Neurosci. 2019, 13, 313. [Google Scholar] [CrossRef]
- Shen, W.; Wang, C.; Xia, L.; Fan, C.; Dong, H.; Deckelbaum, R.J.; Qi, K. Epigenetic Modification of the Leptin Promoter in Diet-Induced Obese Mice and the Effects of N-3 Polyunsaturated Fatty Acids. Sci. Rep. 2014, 4, 5282. [Google Scholar] [CrossRef] [PubMed]
- Wilhelm, J.; Birkenstock, A.; Buchholz, V.; Müller, A.; Aly, S.A.; Gruner-Labitzke, K.; Koehler, H.; Lichtinghagen, R.; Jahn, K.; Groh, A.; et al. Promoter Methylation of LEP and LEPR before and after Bariatric Surgery: A Cross-Sectional Study. Obes. Facts 2021, 14, 93–99. [Google Scholar] [CrossRef]
- Lecoutre, S.; Oger, F.; Pourpe, C.; Butruille, L.; Marousez, L.; Dickes-Coopman, A.; Laborie, C.; Guinez, C.; Lesage, J.; Vieau, D.; et al. Maternal obesity programs increased leptin gene expression in rat male offspring via epigenetic modifications in a depot-specific manner. Mol. Metab. 2017, 6, 922–930. [Google Scholar] [CrossRef] [PubMed]
- Forhead, A.J.; Fowden, A.L. The hungry fetus? Role of leptin as a nutritional signal before birth. J. Physiol. 2009, 587, 1145–1152. [Google Scholar] [CrossRef]
- Glavas, M.M.; Kirigiti, M.A.; Xiao, X.Q.; Enriori, P.J.; Fisher, S.K.; Evans, A.E.; Grayson, B.E.; Cowley, M.A.; Smith, M.S.; Grove, K.L. Early Overnutrition Results in Early-Onset Arcuate Leptin Resistance and Increased Sensitivity to High-Fat Diet. Endocrinology 2010, 151, 1598–1610. [Google Scholar] [CrossRef]
- Alexe, D.-M.; Syridou, G.; Petridou, E.T. Determinants of Early Life Leptin Levels and Later Life Degenerative Outcomes. Clin. Med. Res. 2006, 4, 326–335. [Google Scholar] [CrossRef]
- Zhou, Y.; Rui, L. Leptin signaling and leptin resistance. Front. Med. 2013, 7, 207–222. [Google Scholar] [CrossRef]
- Reed, A.S.; Unger, E.K.; Olofsson, L.E.; Piper, M.L.; Myers, M.G.; Xu, A.W. Functional Role of Suppressor of Cytokine Signaling 3 Upregulation in Hypothalamic Leptin Resistance and Long-Term Energy Homeostasis. Diabetes 2010, 59, 894–906. [Google Scholar] [CrossRef]
- Wauman, J. Leptin receptor signaling: Pathways to leptin resistance. Front. Biosci. 2011, 16, 2771–2793. [Google Scholar] [CrossRef]
- Rahmouni, K.; Morgan, D.A.; Morgan, G.M.; Mark, A.L.; Haynes, W.G. Role of Selective Leptin Resistance in Diet-Induced Obesity Hypertension. Diabetes 2005, 54, 2012–2018. [Google Scholar] [CrossRef]
- Lee, A.K.; Mojtahed-Jaberi, M.; Kyriakou, T.; Astarloa, E.A.-O.; Arno, M.; Marshall, N.J.; Brain, S.D.; O’Dell, S.D. Effect of high-fat feeding on expression of genes controlling availability of dopamine in mouse hypothalamus. Nutrition 2010, 26, 411–422. [Google Scholar] [CrossRef]
- Qiu, J.; Rivera, H.M.; Bosch, M.A.; Padilla, S.L.; Stincic, T.L.; Palmiter, R.D.; Kelly, M.J.; Rønnekleiv, O.K. Estrogenic-dependent glutamatergic neurotransmission from kisspeptin neurons governs feeding circuits in females. eLife 2018, 7. [Google Scholar] [CrossRef]
- Billes, S.K.; Simonds, S.E.; Cowley, M.A. Leptin reduces food intake via a dopamine D2 receptor-dependent mechanism. Mol. Metab. 2012, 1, 86–93. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Vicchi, F.; Ladyman, S.R.; Ornstein, A.M.; Gustafson, P.; Knowles, P.; Luque, G.M.; Grattan, D.R.; Becu-Villalobos, D. Chronic high prolactin levels impact on gene expression at discrete hypothalamic nuclei involved in food intake. FASEB J. 2020, 34, 3902–3914. [Google Scholar] [CrossRef]
- Suyama, S.; Yada, T. New insight into GABAergic neurons in the hypothalamic feeding regulation. J. Physiol. Sci. 2018, 68, 717–722. [Google Scholar] [CrossRef] [PubMed]
- Lorenz-Guertin, J.M.; Jacob, T.C. GABA type a receptor trafficking and the architecture of synaptic inhibition. Dev. Neurobiol. 2017, 78, 238–270. [Google Scholar] [CrossRef] [PubMed]
- Ge, T.T.; Yao, X.X.; Zhao, F.L.; Zou, X.H.; Yang, W.; Cui, R.J.; Li, B.J. Role of leptin in the regulation of food intake in fasted mice. J. Cell. Mol. Med. 2020, 24, 4524–4532. [Google Scholar] [CrossRef]
- Bekdash, R.A.; Harrison, N.L. Downregulation of Gabra4 expression during alcohol withdrawal is mediated by specific micro RNA s in cultured mouse cortical neurons. Brain Behav. 2015, 5, e00355. [Google Scholar] [CrossRef] [PubMed]
- Mercader, J.M.; Lozano, J.J.; Sumoy, L.; Dierssen, M.; Visa, J.; Gratacòs, M.; Estivill, X. Hypothalamus transcriptome profile suggests an anorexia-cachexia syndrome in the anx/anx mouse model. Physiol. Genom. 2008, 35, 341–350. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Sequence (5’-3’) | Reverse Sequence (3’-5’) | Acc. No |
|---|---|---|---|
| B2M | GTGCTTGCCATTCAGAAAACTCC | AACTGAGACACGTAGCAGTTGAG | NM_012512.2 |
| Pomc | GACCTCACCACGGAAAGCAA | TGTTCATCTCCGTTGCCTGG | NM_139326.3 |
| Npy | TGGCCAGATACTACTCCGCT | TTCAAGCCTTGTTCTGGGGG | NM_012614.2 |
| ObRb | CCAGTACCCAGAGCCAAAGT | GGATCGGGCTTCACAACAAGC | NM_012596.1 |
| Stat3 | TGCATTGATAAGGACTCTGGGG | CTGCCGTTGTTGGACTCCTC | NM_012747.2 |
| Socs3 | CCTCCAGCATCTTTGTCGGAAGAC | TACTGGTCCAGGAACTCCCGAATG | NM_053565.1 |
| Drd2 | GTTGTCTACCTGGAGGTGGTG | AGGTTCAGGATGCTTGCTGTG | NM_012547 |
| Gabra4 | ACAGTGATCCTTTCTCAAGTTTCC | CGTGAGGACTGTGGTTATTCCAA | NM_080587.3 |
| Gabrd | TTTATACAGCATCCGCATCACCT | GAAGAGTAGCCATAGCTCTCCAG | NM_017289.2 |
| Gabre | AGGAATTCTAAGAGGACCCAAGA | GCATCAATGGTCATCCTAACTGTG | NM_023091.1 |
| Grm2 | CTCCAGTGATTATCGGGTGCAG | TTCTGTGGCTGGAAAAGGATGAT | NM_001105711.1 |
| 1X | 5X | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1X-FA | 1X-MTHF | 5X-FA | 5X-MTHF | Two-way ANOVA p-Value | |||||||||||
| Mean | ± | S.E.M. | Mean | ± | S.E.M. | Mean | ± | S.E.M. | Mean | ± | S.E.M. | Dose | Form | Dose × Form | |
| Birth | |||||||||||||||
| Leptin ng/mL | 0.86 | ± | 0.25 | 1.32 | ± | 0.28 | 1.93 | ± | 0.28 | 1.64 | ± | 0.37 | 0.03 | 0.78 | 0.23 |
| Insulin ng/mL | 0.30 | ± | 0.05 | 0.19 | ± | 0.05 | 0.35 | ± | 0.05 | 0.29 | ± | 0.08 | 0.20 | 0.17 | 0.68 |
| Glucose mg/dL | 74.38 | ± | 6.39 | 70.30 | ± | 7.83 | 77.30 | ± | 7.24 | 72.05 | ± | 9.58 | 0.77 | 0.56 | 0.94 |
| HOMA-IR | 0.22 | ± | 0.05 | 0.15 | ± | 0.07 | 0.29 | ± | 0.06 | 0.21 | ± | 0.07 | 0.34 | 0.27 | 0.97 |
| 19 weeks PW | |||||||||||||||
| Leptin ng/mL | 14.08 | ± | 2.12 | 11.15 | ± | 2.12 | 17.18 | ± | 1.95 | 18.35 | ± | 2.12 | 0.02 | 0.67 | 0.33 |
| Leptin/VAT | 0.29 | ± | 0.05 | 0.27 | ± | 0.04 | 0.40 | ± | 0.04 | 0.37 | ± | 0.04 | 0.02 | 0.53 | 0.85 |
| Ghrelin ng/mL | 153.24 | ± | 18.23 | 110.19 | ± | 19.21 | 110.19 | ± | 17.38 | 102.79 | ± | 19.21 | 0.37 | 0.08 | 0.16 |
| Insulin ng/mL | 2.00 | ± | 0.24 | 1.76 | ± | 0.22 | 2.25 | ± | 0.26 | 1.95 | ± | 0.22 | 0.62 | 0.26 | 0.91 |
| Glucose mg/dL | 120.57 | ± | 7.11 | 115.81 | ± | 6.71 | 129.40 | ± | 7.60 | 127.67 | ± | 6.36 | 0.15 | 0.64 | 0.83 |
| HOMA-IR | 2.53 | ± | 0.38 | 2.54 | ± | 0.36 | 3.05 | ± | 0.41 | 2.62 | ± | 0.34 | 0.43 | 0.58 | 0.56 |
| TG nmol/dL | 70.01 | ± | 7.88 | 83.08 | ± | 7.37 | 45.17 | ± | 7.88 | 70.08 | ± | 7.88 | 0.02 | 0.02 | 0.45 |
| 1X | 5X | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1X-FA | 1X-MTHF | 5X-FA | 5X-MTHF | Two-Way ANOVA p-Value | |||||||||||
| Mean | ± | S.E.M. | Mean | ± | S.E.M. | Mean | ± | S.E.M. | Mean | ± | S.E.M. | Dose | Form | Dose×Form | |
| Birth Plasma | |||||||||||||||
| 5-MTHF (nmol/L) | 217.0 | ± | 25.34 | 174.80 | ± | 25.34 | 275.60 | ± | 25.34 | 331.2 | ± | 25.34 | 0.004 | 0.81 | 0.09 |
| 19 weeks PW Plasma | |||||||||||||||
| 5-MTHF (nmol/L) | 127.14 | ± | 7.83 | 144.38 | ± | 7.33 | 126.34 | ± | 7.33 | 133.38 | ± | 7.33 | 0.44 | 0.12 | 0.55 |
| Met (µmol/L) | 73.77 | ± | 2.87 | 73.41 | ± | 2.68 | 69.39 | ± | 2.87 | 74.26 | ± | 2.68 | 0.53 | 0.42 | 0.35 |
| SAM (nmol/L) | 269.02 | ± | 11.77 | 269.87 | ± | 11.77 | 267.97 | ± | 12.58 | 248.94 | ± | 11.77 | 0.37 | 0.45 | 0.41 |
| SAH (nmol/L) | 74.16 | ± | 8.84 | 76.43 | ± | 8.84 | 79.68 | ± | 8.84 | 72.90 | ± | 8.84 | 0.91 | 0.80 | 0.61 |
| Homocysteine (µmol/L) | 5.81 | ± | 0.88 | 4.20 | ± | 1.01 | 5.66 | ± | 0.88 | 5.84 | ± | 0.94 | 0.43 | 0.45 | 0.34 |
| Cystathionine (nmol/L) | 830.27 | ± | 83.80 | 827.55 | ± | 83.80 | 700.03 | ± | 83.80 | 611.89 | ± | 83.80 | 0.04 | 0.59 | 0.61 |
| Choline (µmol/L) | 10.89 | ± | 0.72 | 11.36 | ± | 0.72 | 10.70 | ± | 0.72 | 10.34 | ± | 0.72 | 0.41 | 0.94 | 0.57 |
| Betaine (µmol/L) | 90.71 | ± | 7.53 | 86.57 | ± | 6.25 | 85.18 | ± | 6.68 | 67.06 | ± | 6.25 | 0.06 | 0.09 | 0.29 |
| 19 weeks PW Liver | |||||||||||||||
| 5-MTHF (nmol/L) | 27.78 | 1.63 | 32.31 | 1.63 | 29.57 | 1.63 | 30.75 | 1.63 | 0.94 | 0.09 | 0.31 | ||||
| Met (µmol/L) | 447.80 | ± | 45.17 | 447.88 | ± | 45.17 | 506.25 | ± | 45.17 | 508.25 | ± | 45.17 | 0.20 | 0.98 | 0.98 |
| SAM (nmol/L) | 8.75 | ± | 3.83 | 15.85 | ± | 3.32 | 17.33 | ± | 3.32 | 17.15 | ± | 3.32 | 0.15 | 0.34 | 0.32 |
| SAH (nmol/L) | 15.55 | ± | 3.36 | 21.78 | ± | 2.91 | 18.41 | ± | 2.91 | 21.56 | ± | 2.91 | 0.67 | 0.13 | 0.61 |
| Cystathionine (nmol/L) | 25.48 | ± | 2.51 | 23.77 | ± | 2.51 | 26.61 | ± | 2.51 | 26.99 | ± | 2.51 | 0.51 | 0.95 | 0.54 |
| Choline (µmol/L) | 318.63 | ± | 64.36 | 384.00 | ± | 64.36 | 317.75 | ± | 64.37 | 346.38 | ± | 64.36 | 0.77 | 0.47 | 0.77 |
| Betaine (µmol/L) | 2458.75 ab | ± | 233.63 | 2585 ab | ± | 233.63 | 3045 a | ± | 249.76 | 2230.75 b | ± | 233.63 | 0.60 | 0.13 | 0.04 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pannia, E.; Hammoud, R.; Kubant, R.; Sa, J.Y.; Simonian, R.; Wasek, B.; Ashcraft, P.; Bottiglieri, T.; Pausova, Z.; Anderson, G.H. High Intakes of [6S]-5-Methyltetrahydrofolic Acid Compared with Folic Acid during Pregnancy Programs Central and Peripheral Mechanisms Favouring Increased Food Intake and Body Weight of Mature Female Offspring. Nutrients 2021, 13, 1477. https://doi.org/10.3390/nu13051477
Pannia E, Hammoud R, Kubant R, Sa JY, Simonian R, Wasek B, Ashcraft P, Bottiglieri T, Pausova Z, Anderson GH. High Intakes of [6S]-5-Methyltetrahydrofolic Acid Compared with Folic Acid during Pregnancy Programs Central and Peripheral Mechanisms Favouring Increased Food Intake and Body Weight of Mature Female Offspring. Nutrients. 2021; 13(5):1477. https://doi.org/10.3390/nu13051477
Chicago/Turabian StylePannia, Emanuela, Rola Hammoud, Ruslan Kubant, Jong Yup Sa, Rebecca Simonian, Brandi Wasek, Paula Ashcraft, Teodoro Bottiglieri, Zdenka Pausova, and G. Harvey Anderson. 2021. "High Intakes of [6S]-5-Methyltetrahydrofolic Acid Compared with Folic Acid during Pregnancy Programs Central and Peripheral Mechanisms Favouring Increased Food Intake and Body Weight of Mature Female Offspring" Nutrients 13, no. 5: 1477. https://doi.org/10.3390/nu13051477
APA StylePannia, E., Hammoud, R., Kubant, R., Sa, J. Y., Simonian, R., Wasek, B., Ashcraft, P., Bottiglieri, T., Pausova, Z., & Anderson, G. H. (2021). High Intakes of [6S]-5-Methyltetrahydrofolic Acid Compared with Folic Acid during Pregnancy Programs Central and Peripheral Mechanisms Favouring Increased Food Intake and Body Weight of Mature Female Offspring. Nutrients, 13(5), 1477. https://doi.org/10.3390/nu13051477