
Immunomodulating Effects of Fungal Beta-Glucans: From Traditional Use to Medicine


Abstract
1. Introduction
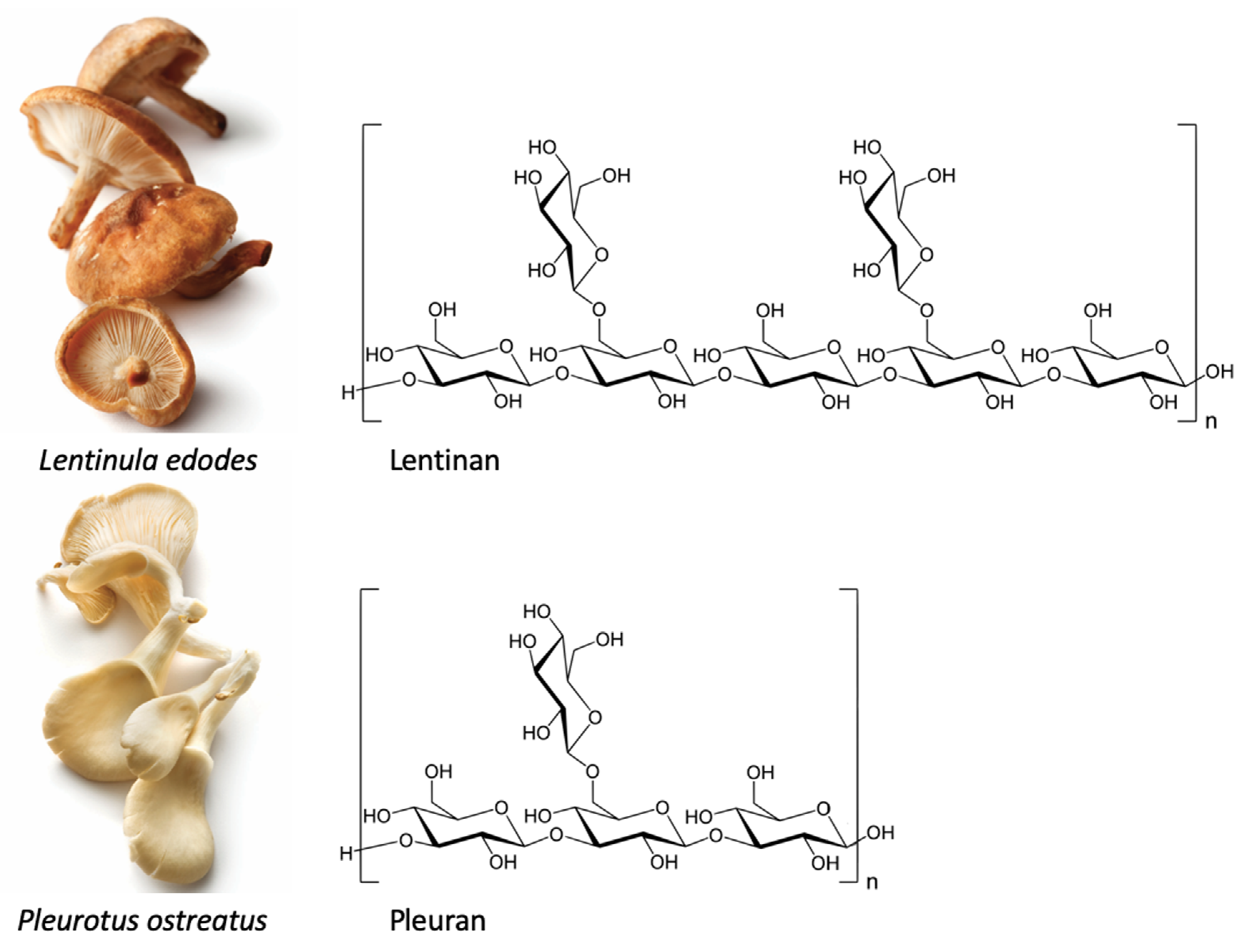
Structure, Chemical Properties, and Natural Sources of Beta-Glucans
2. Beta-Glucans in Immunomodulating Dietary Supplements
2.1. Health Claims in Europe
2.2. Immune Functioning Health Claims
2.3. Yestimun®
2.4. Pleuran
3. Trained Immunity
3.1. BCG Vaccine
3.2. Mifamurtide
3.3. Fungal Glucans and Trained Immunity
3.3.1. Lentinan
3.3.2. Imprime PGG
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Javelle, E.; Raoult, D. COVID-19 pandemic more than a century after the Spanish flu. Lancet Infect. Dis. 2021, 21, e78. [Google Scholar] [CrossRef]
- Rao, K.S.; Suryaprakash, V.; Senthilkumar, R.; Preethy, S.; Katoh, S.; Ikewaki, N.; Abraham, S.J.K. Role of Immune Dysregu-lation in Increased Mortality Among a Specific Subset of COVID-19 Patients and Immune-Enhancement Strategies for Com-batting Through Nutritional Supplements. Front. Immunol. 2020, 11, 154. [Google Scholar] [CrossRef] [PubMed]
- Fara, A.; Mitrev, Z.; Rosalia, R.A.; Assas, B.M. Cytokine storm and COVID-19: A chronicle of pro-inflammatory cytokines. Open Biol. 2020, 10, 200160. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.; Cao, Y.; Lu, X.; Zhang, J.; Du, H.; Yan, Y.; Akdis, C.A.; Gao, Y. Eleven faces of coronavirus disease 2019. Allergy 2020, 75, 1699–1709. [Google Scholar] [CrossRef] [PubMed]
- Wichers, H. Immunomodulation by food: Promising concept for mitigating allergic disease? Anal. Bioanal. Chem. 2009, 395, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Mogensen, T.H. Pathogen Recognition and Inflammatory Signaling in Innate Immune Defenses. Clin. Microbiol. Rev. 2009, 22, 240–273. [Google Scholar] [CrossRef]
- Ma, H.-D.; Deng, Y.-R.; Tian, Z.; Lian, Z.-X. Traditional Chinese Medicine and Immune Regulation. Clin. Rev. Allergy Immunol. 2013, 44, 229–241. [Google Scholar] [CrossRef] [PubMed]
- Vetvicka, V.; Vannucci, L.; Sima, P.; Richter, J. Beta Glucan: Supplement or Drug? From Laboratory to Clinical Trials. Molecules 2019, 24, 1251. [Google Scholar] [CrossRef]
- Wasser, S. Medicinal mushrooms as a source of antitumor and immunomodulating polysaccharides. Appl. Microbiol. Biotechnol. 2002, 60, 258–274. [Google Scholar]
- Stuart, I.; Loi, L.; Fincher, G. Immunological comparison of (1→3,1→4)-β-glucan endohydrolases in germinating cereals. J. Cereal Sci. 1987, 6, 45–52. [Google Scholar] [CrossRef]
- Nakashima, A.; Yamada, K.; Iwata, O.; Sugimoto, R.; Atsuji, K.; Ogawa, T.; Ishibashi-Ohgo, N.; Suzuki, K. β-Glucan in Foods and Its Physiological Functions. J. Nutr. Sci. Vitaminol. 2018, 64, 8–17. [Google Scholar] [CrossRef]
- Ripsin, C.M.; Keenan, J.M.; Jacobs, D.R.; Elmer, P.J.; Welch, R.R.; Van Horn, L.; Liu, K.; Turnbull, W.H.; Thye, F.W.; Kestin, M.; et al. Oat Products and Lipid Lowering. JAMA 1992, 267, 3317–3325. [Google Scholar] [CrossRef]
- Kim, S.Y.; Song, H.J.; Lee, Y.Y.; Cho, K.H.; Roh, Y.K. Biomedical issues of dietary fiber beta-glucan. J. Korean Med. Sci. 2006, 21, 781. [Google Scholar] [CrossRef]
- Novak, M.; Vetvicka, V. β-Glucans, History, and the Present: Immunomodulatory Aspects and Mechanisms of Action. J. Immunotoxicol. 2008, 5, 47–57. [Google Scholar] [CrossRef] [PubMed]
- Goodridge, H.S.; Wolf, A.J.; Underhill, D.M. β-glucan recognition by the innate immune system. Immunol. Rev. 2009, 230, 38–50. [Google Scholar] [CrossRef]
- Bashir, K.M.I.; Choi, J.-S. Clinical and Physiological Perspectives of β-Glucans: The Past, Present, and Future. Int. J. Mol. Sci. 2017, 18, 1906. [Google Scholar] [CrossRef] [PubMed]
- Lv, G.; Fan, L.; Zhang, Z.; Pan, H. Development of research on lentinan. Acta Agric. Zhejiangensis 2009, 21, 183–188. [Google Scholar]
- Majtan, J. Pleuran (β-Glucan fromPleurotus ostreatus): An Effective Nutritional Supplement against Upper Respiratory Tract Infections? Pediatric Fitness 2012, 59, 57–61. [Google Scholar] [CrossRef]
- Rop, O.; Mlcek, J.; Jurikova, T. Beta-glucans in higher fungi and their health effects. Nutr. Rev. 2009, 67, 624–631. [Google Scholar] [CrossRef] [PubMed]
- Lenssen, K.G.; Bast, A.; de Boer, A. Clarifying the health claim assessment procedure of EFSA will benefit functional food innovation. J. Funct. Foods 2018, 47, 386–396. [Google Scholar] [CrossRef]
- Avermaete, T.; Viaene, J.; Morgan, E.J.; Crawford, N. Determinants of innovation in small food firms. Eur. J. Innov. Manag. 2003, 6, 8–17. [Google Scholar] [CrossRef]
- Costa, A.; Jongen, W. New insights into consumer-led food product development. Trends Food Sci. Technol. 2006, 17, 457–465. [Google Scholar] [CrossRef]
- The European Commission Commission Regulation (EU) No 432/2012. Off. J. Eur. Union. 2012, 13, 1–40.
- De Boer, A.; Vos, E.; Bast, A. Implementation of the nutrition and health claim regulation—The case of antioxidants. Regul. Toxicol. Pharmacol. 2014, 68, 475–487. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.; Terry, S.; Detken, D. 10 years of the European Food Safety Authority (EFSA) and the EU Food Safety System. Eur. Food Feed Law Rev. EFFL 2012, 7, 111–116. [Google Scholar]
- Verhagen, H.; van Loveren, H. Status of nutrition and health claims in Europe by mid 2015. Trends Food Sci. Technol. 2016, 56, 39–45. [Google Scholar] [CrossRef]
- Mainous, M.R.; Deitch, E.A. Nutrition and Infection. Surg. Clin. N. Am. 1994, 74, 659–676. [Google Scholar] [CrossRef]
- Van Buren, C.T. Nutrition and Immunology. J. Parenter. Enter. Nutr. 1991, 15. [Google Scholar] [CrossRef]
- Khedkar, S.; Ciliberti, S.; Bröring, S. The EU health claims regulation: Implications for innovation in the EU food sector. Br. Food J. 2016, 118, 2647–2665. [Google Scholar] [CrossRef]
- Khedkar, S.; Bröring, S.; Ciliberti, S. Exploring the Nutrition and Health Claims Regulation (EC) No. 1924/2006: What is the impact on innovation in the EU food sector? Int. J. Food Sci. Nutr. 2016, 68, 10–17. [Google Scholar] [CrossRef]
- EFSA Panel on Dietetic Products, Nutrition and Allergies (NDA). Guidance on the scientific requirements for health claims related to gut and immune function. EFSA J. 2011, 9, 2474. [Google Scholar]
- EFSA Panel on Dietetic Products, Nutrition and Allergies (NDA). Scientific Opinion on the substantiation of health claims related to vitamin D and normal function of the immune system and inflammatory response (ID 154, 159), maintenance of normal muscle function (ID 155) and maintenance of normal cardio-vascular function (ID 159) pursuant to Article 13(1) of Regulation (EC) No 1924/2006. EFSA J. 2010, 8, 1468. [Google Scholar]
- EFSA Panel on Dietetic Products, Nutrition and Allergies (NDA). Scientific Opinion on the substantiation of a health claim related to vitamin D and contribution to the normal function of the immune system pursuant to Article 14 of Regulation (EC) No 1924/2006. EFSA J. 2015, 13, 4096. [Google Scholar] [CrossRef][Green Version]
- EFSA Panel on Dietetic Products, Nutrition and Allergies (NDA). Scientific Opinion on the substantiation of health claims related to vitamin B6 and protein and glycogen metabolism (ID 65, 70, 71), function of the nervous system (ID 66), red blood cell formation (ID 67, 72, 186), function of the immune system (ID 68). EFSA J. 2009, 7, 1225. [Google Scholar] [CrossRef]
- EFSA Panel on Dietetic Products, Nutrition and Allergies (NDA). Scientific Opinion on the substantiation of a health claim related to zinc and normal function of the immune system pursuant to Article 14 of Regulation (EC) No 1924/2006. EFSA J. 2014, 12, 3653. [Google Scholar] [CrossRef][Green Version]
- EFSA Panel on Dietetic Products, Nutrition and Allergies (NDA). Scientific Opinion on the substantiation of health claims related to copper and protection of DNA, proteins and lipids from oxidative damage (ID 263, 1726), function of the immune system (ID 264), maintenance of connective tissues (ID 265, 271, 1722), ene. EFSA J. 2009, 7, 1211. [Google Scholar] [CrossRef]
- EFSA Panel on Dietetic Products, Nutrition and Allergies (NDA). Scientific Opinion on the substantiation of health claims related to copper and reduction of tiredness and fatigue (ID 272), maintenance of the normal function of the nervous system (ID 1723), maintenance of the normal function of the immune system (ID 17). EFSA J. 2011, 9, 2079. [Google Scholar] [CrossRef]
- EFSA Panel on Dietetic Products, Nutrition and Allergies (NDA). Scientific Opinion on the substantiation of health claims related to selenium and protection of DNA, proteins and lipids from oxidative damage (ID 277, 283, 286, 1289, 1290, 1291, 1293, 1751), function of the immune system (ID 278), thyroid function (ID 2. EFSA J. 2009, 7, 1220. [Google Scholar] [CrossRef]
- EFSA Panel on Dietetic Products, Nutrition and Allergies (NDA). Scientific Opinion on the substantiation of health claims related to iron and formation of red blood cells and haemoglobin (ID 249, ID 1589), oxygen transport (ID 250, ID 254, ID 256), energy-yielding metabolism (ID 251, ID 1589), function of the immune s. EFSA J. 2009, 7, 1215. [Google Scholar] [CrossRef]
- EFSA Panel on Dietetic Products, Nutrition and Allergies (NDA). Guidance on the scientific requirements for health claims related to the immune system, the gastrointestinal tract and defence against pathogenic microorganisms. EFSA J. 2016, 14, 4369. [Google Scholar] [CrossRef]
- Douglas, R.; Hemilä, H.; Chalker, E.; Treacy, B. Cochrane review: Vitamin C for preventing and treating the common cold. Evid. Child Health A Cochrane Rev. J. 2008, 3, 672–720. [Google Scholar] [CrossRef]
- Douglas, R.M.; Hemilä, H. Vitamin C for preventing and treating the common cold. PLoS Med. 2005, 2, e168. [Google Scholar] [CrossRef]
- Hemilä, H.; Chalker, E. Vitamin C for preventing and treating the common cold. Cochrane Database Syst. Rev. 2013. [Google Scholar] [CrossRef] [PubMed]
- Galmés, S.; Serra, F.; Palou, A. Current State of Evidence: Influence of Nutritional and Nutrigenetic Factors on Immunity in the COVID-19 Pandemic Framework. Nutrients 2020, 12, 2738. [Google Scholar] [CrossRef]
- Donabedian, H. Nutritional therapy and infectious diseases: A two-edged sword. Nutr. J. 2006, 5, 21. [Google Scholar] [CrossRef]
- Glanville, J.; King, S.; Guarner, F.; Hill, C.; Sanders, M.E. A review of the systematic review process and its applicability for use in evaluating evidence for health claims on probiotic foods in the European Union. Nutr. J. 2015, 14, 16. [Google Scholar] [CrossRef]
- Rijkers, G.T.; De Vos, W.M.; Brummer, R.-J.; Morelli, L.; Corthier, G.; Marteau, P. Health benefits and health claims of probiotics: Bridging science and marketing. Br. J. Nutr. 2011, 106, 1291–1296. [Google Scholar] [CrossRef]
- Miquel, S.; Beaumont, M.; Martín, R.; Langella, P.; Braesco, V.; Thomas, M. A proposed framework for an appropriate evalu-ation scheme for microorganisms as novel foods with a health claim in Europe. Microb. Cell Fact. 2015, 14, 48. [Google Scholar] [CrossRef]
- EFSA Panel on Dietetic Products, Nutrition and Allergies (NDA). Scientific Opinion on the substantiation of health claims related to non characterised bacteria and yeasts pursuant to Article 13(1) of Regulation (EC) No 1924/2006. EFSA J. 2010, 8, 1470. [Google Scholar] [CrossRef]
- Stier, H.; Ebbeskotte, V.; Gruenwald, J. Immune-modulatory effects of dietary Yeast Beta-1,3/1,6-D-glucan. Nutr. J. 2014, 13, 38. [Google Scholar] [CrossRef]
- Wójcik, R. Effect of Biolex Beta-HP on phagocytic activity and oxidative metabolism of peripheral blood granulocytes and monocytes in rats intoxicated by cyclophosphamide. Pol. J. Vet. Sci. 2010, 13, 181–188. [Google Scholar] [PubMed]
- Wójcik, R.; Janowska, E.; Małaczewska, J.; Siwicki, A.K. Effect of β-1,3/1,6-D-glucan on the phagocytic activity and oxidative metabolism of peripheral blood granulocytes and monocytes in rats. Bull. Vet. Inst. Pulawy 2009, 53, 241–246. [Google Scholar]
- Małaczewska, J.; Wójcik, R.; Jung, L.; Siwicki, A.K. Effect of biolex β-HP on selected parameters of specific and non-specific humoral and cellular immunity in rats. Bull. Vet. Inst. Pulawy 2010, 54, 75–80. [Google Scholar]
- Rychlik, A.; Nieradka, R.; Kander, M.; Nowicki, M.; Wdowiak, M.; Kołodziejska-Sawerska, A. The effectiveness of natural and synthetic immunomodulators in the treatment of inflammatory bowel disease in dogs. Acta Vet. Hung. 2013, 61, 297–308. [Google Scholar] [CrossRef] [PubMed]
- Auinger, A.; Riede, L.; Bothe, G.; Busch, R.; Gruenwald, J. Yeast (1,3)-(1,6)-beta-glucan helps to maintain the body’s defence against pathogens: A double-blind, randomized, placebo-controlled, multicentric study in healthy subjects. Eur. J. Nutr. 2013, 52, 1913–1918. [Google Scholar] [CrossRef] [PubMed]
- Graubaum, H.-J.; Busch, R.; Stier, H.; Gruenwald, J. A Double-Blind, Randomized, Placebo-Controlled Nutritional Study Using an Insoluble Yeast Beta-Glucan to Improve the Immune Defense System. Food Nutr. Sci. 2012, 3, 738–746. [Google Scholar] [CrossRef]
- EFSA Panel on Dietetic Products, Nutrition and Allergies (NDA). Scientific Opinion on the substantiation of a health claim related to Yestimun® and defence against pathogens in the upper respiratory tract pursuant to Article 13(5) of Regulation (EC) No 1924/2006. EFSA J. 2013, 11, 3159. [Google Scholar] [CrossRef]
- Response to comments on the Scientific Opinion on the substantiation of a health claim related to Yestimun® and defence against pathogens in the upper respiratory tract pursuant to Article 13(5) of Regulation (EC) No 1924/2006. EFSA Support. Publ. 2013, 10, 500E. [CrossRef][Green Version]
- Dharsono, T.; Rudnicka, K.; Wilhelm, M.; Schoen, C. Effects of Yeast (1,3)-(1,6)-Beta-Glucan on Severity of Upper Respiratory Tract Infections: A Double-Blind, Randomized, Placebo-Controlled Study in Healthy Subjects. J. Am. Coll. Nutr. 2019, 38, 40–50. [Google Scholar] [CrossRef] [PubMed]
- Barrett, B.; Locken, K.; Maberry, R.; Schwamman, J.; Brown, R.; Bobula, J.; Stauffacher, E.A. The Wisconsin Upper Respiratory Symptom Survey (WURSS): A new research instrument for assessing the common cold. J. Fam. Pr. 2002, 51, 265. [Google Scholar]
- Turner, R.B. The Common Cold. Mandell Douglas Bennett’s Princ. Pract. Infect. Dis. 2015, 748–752.e2. [Google Scholar] [CrossRef]
- Jeseňák, M.; Sanisló, E.; Kuniaková, R.; Rennerová, Z.; Buchanec, J.; Bánovčin, P. Imunoglukan P4H® in the prevention of recurrent respiratory infections in childhood. Cesk Pediatr. 2010, 73, 639–647. [Google Scholar]
- Sapena Grau, J.; Picó Sirvent, L.; Morera Inglés, M.; Rivero Urgell, M. Beta-glucans from Pleurotus ostreatus for prevention of recurrent respiratory tract infections. Acta Pediátrica Española 2015, 73, 186–193. [Google Scholar]
- Pasnik, J.; Ślemp, A.; Cywinska-Bernas, A.; Zeman, K.; Jesenak, M. Preventive effect of pleuran (β-glucan from Pleurotus ostreatus) in children with recurrent respiratory tract infections—Open-label prospective study. Curr. Pediatr. Res. 2017, 21, 99–104. [Google Scholar]
- Moreira, A.; Kekkonen, R.A.; Delgado, L.; Fonseca, J.; Korpela, R.; Haahtela, T. Nutritional modulation of exercise-induced immunodepression in athletes: A systematic review and meta-analysis. Eur. J. Clin. Nutr. 2007, 61, 443–460. [Google Scholar] [CrossRef] [PubMed]
- Jesenak, M.; Majtan, J.; Rennerova, Z.; Kyselovic, J.; Banovcin, P.; Hrubisko, M. Immunomodulatory effect of pleuran (β-glucan from Pleurotus ostreatus) in children with recurrent respiratory tract infections. Int. Immunopharmacol. 2013, 15, 395–399. [Google Scholar] [CrossRef]
- Bergendiova, K.; Tibenska, E.; Majtan, J. Pleuran (β-glucan from Pleurotus ostreatus) supplementation, cellular immune response and respiratory tract infections in athletes. Eur. J. Appl. Physiol. 2011, 111, 2033–2040. [Google Scholar] [CrossRef]
- Bobovčák, M.; Kuniaková, R.; Gabriž, J.; Majtán, J. Effect of Pleuran (β-glucan from Pleurotus ostreatus) supplementation on cellular immune response after intensive exercise in elite athletes. Appl. Physiol. Nutr. Metab. 2010, 35, 755–762. [Google Scholar] [CrossRef] [PubMed]
- Gałązka-Franta, A.; Jura-Szołtys, E.; Smółka, W.; Gawlik, R. Upper Respiratory Tract Diseases in Athletes in Different Sports Disciplines. J. Hum. Kinet. 2016, 53, 99–106. [Google Scholar] [CrossRef][Green Version]
- Mayr, A. Taking Advantage of the Positive Side-Effects of Smallpox Vaccination. J. Vet. Med. Ser. B 2004, 51, 199–201. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Ramón, S.; Conejero, L.; Netea, M.G.; Sancho, D.; Palomares, Ó.; Subiza, J.L. Trained Immunity-Based Vaccines: A New Paradigm for the Development of Broad-Spectrum Anti-infectious Formulations. Front. Immunol. 2018, 9, 2936. [Google Scholar] [CrossRef]
- Netea, M.G.; Quintin, J.; Van Der Meer, J.W. Trained Immunity: A Memory for Innate Host Defense. Cell Host Microbe 2011, 9, 355–361. [Google Scholar] [CrossRef]
- Quintin, J.; Saeed, S.; Martens, J.H.; Giamarellos-Bourboulis, E.J.; Ifrim, D.C.; Logie, C.; Jacobs, L.; Jansen, T.; Kullberg, B.-J.; Wijmenga, C.; et al. Candida albicans Infection Affords Protection against Reinfection via Functional Reprogramming of Monocytes. Cell Host Microbe 2012, 12, 223–232. [Google Scholar] [CrossRef]
- Mourits, V.P.; Wijkmans, J.C.; Ab Joosten, L.; Netea, M.G. Trained immunity as a novel therapeutic strategy. Curr. Opin. Pharmacol. 2018, 41, 52–58. [Google Scholar] [CrossRef]
- Arts, R.J.W.; Joosten, L.A.B.; Netea, M.G. The potential role of trained immunity in autoimmune and autoinflammatory dis-orders. Front. Immunol. 2018, 9, 298. [Google Scholar] [CrossRef] [PubMed]
- Rusek, P.; Wala, M.; Druszczyńska, M.; Fol, M. Infectious agents as stimuli of trained innate immunity. Int. J. Mol. Sci. 2018, 19, 456. [Google Scholar] [CrossRef] [PubMed]
- Lérias, J.R.; de Sousa, E.; Paraschoudi, G.; Martins, J.; Condeço, C.; Figueiredo, N.; Carvalho, C.; Dodoo, E.; Maia, A.; Cas-tillo-Martin, M.; et al. Trained Immunity for Personalized Cancer Immunotherapy: Current Knowledge and Future Opportu-nities. Front. Microbiol. 2020, 10, 2924. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization Global Tuberculosis Report 2018; World Health Organization: Geneva, Switzerland, 2018.
- Butkeviciute, E.; Jones, C.E.; Smith, S.G. Heterologous effects of infant BCG vaccination: Potential mechanisms of immunity. Futur. Microbiol. 2018, 13, 1193–1208. [Google Scholar] [CrossRef]
- Asín, M.A.P.-J.; Fernández-Ruiz, M.; López-Medrano, F.; Lumbreras, C.; Tejido, Á.; Juan, R.S.; Arrebola-Pajares, A.; Lizasoain, M.; Prieto, S.; Aguado, J.M. Bacillus Calmette-Guérin (BCG) infection following intravesical BCG administration as adjunctive therapy for bladder cancer: Incidence, risk factors, and outcome in a single-institution series and review of the literature. Medicine 2014, 93, 236–254. [Google Scholar] [CrossRef] [PubMed]
- Bistoni, F.; Vecchiarelli, A.; Cenci, E.; Puccetti, P.; Marconi, P.; Cassone, A. Evidence for macrophage-mediated protection against lethal Candida albicans infection. Infect. Immun. 1986, 51, 668–674. [Google Scholar] [CrossRef]
- Bistoni, F.; Verducci, G.; Perito, S.; Vecchiarelli, A.; Puccetti, P.; Marconi, P.; Cassone, A. Immunomodulation by a low-virulence, agerminative variant ofCandida albicans. Further evidence for macrophage activation as one of the effector mechanisms of nonspecific anti-infectious protection. Med. Mycol. 1988, 26, 285–299. [Google Scholar] [CrossRef]
- Wout, J.W.V.; Poell, R.; Van Furth, R. The Role of BCG/PPD-Activated Macrophages in Resistance against Systemic Candidiasis in Mice. Scand. J. Immunol. 1992, 36, 713–720. [Google Scholar] [CrossRef]
- Kleinnijenhuis, J.; Quintin, J.; Preijers, F.; Joosten, L.A.B.; Ifrim, D.C.; Saeed, S.; Jacobs, C.; Van Loenhout, J.; De Jong, D.; Stunnenberg, H.G.; et al. Bacille Calmette-Guerin induces NOD2-dependent nonspecific protection from reinfection via epigenetic reprogramming of monocytes. Proc. Natl. Acad. Sci. USA 2012, 109, 17537–17542. [Google Scholar] [CrossRef]
- Uthayakumar, D.; Paris, S.; Chapat, L.; Freyburger, L.; Poulet, H.; De Luca, K. Non-specific Effects of Vaccines Illustrated Through the BCG Example: From Observations to Demonstrations. Front. Immunol. 2018, 9, 2869. [Google Scholar] [CrossRef] [PubMed]
- Kleinnijenhuis, J.; Quintin, J.; Preijers, F.; Benn, C.S.; Joosten, L.A.; Jacobs, C.; Van Loenhout, J.; Xavier, R.J.; Aaby, P.; Van Der Meer, J.W.; et al. Long-Lasting Effects of BCG Vaccination on Both Heterologous Th1/Th17 Responses and Innate Trained Immunity. J. Innate Immun. 2014, 6, 152–158. [Google Scholar] [CrossRef] [PubMed]
- Arts, R.J.; Moorlag, S.J.; Novakovic, B.; Li, Y.; Wang, S.-Y.; Oosting, M.; Kumar, V.; Xavier, R.J.; Wijmenga, C.; Joosten, L.A.; et al. BCG Vaccination Protects against Experimental Viral Infection in Humans through the Induction of Cytokines Associated with Trained Immunity. Cell Host Microbe 2018, 23, 89–100. [Google Scholar] [CrossRef] [PubMed]
- Walk, J.; De Bree, L.C.J.; Graumans, W.; Stoter, R.; Van Gemert, G.-J.; Van De Vegte-Bolmer, M.; Teelen, K.; Hermsen, C.C.; Arts, R.J.W.; Behet, M.C.; et al. Outcomes of controlled human malaria infection after BCG vaccination. Nat. Commun. 2019, 10, 874. [Google Scholar] [CrossRef] [PubMed]
- Freyne, B.; Donath, S.; Germano, S.; Gardiner, K.; Casalaz, D.; Robins-Browne, R.M.; Amenyogbe, N.; Messina, N.L.; Netea, M.G.; Flanagan, K.L.; et al. Neonatal BCG Vaccination Influences Cytokine Responses to Toll-like Receptor Ligands and Heterologous Antigens. J. Infect. Dis. 2018, 217, 1798–1808. [Google Scholar] [CrossRef]
- Leentjens, J.; Kox, M.; Stokman, R.; Gerretsen, J.; Diavatopoulos, D.A.; Van Crevel, R.; Rimmelzwaan, G.F.; Pickkers, P.; Netea, M.G. BCG Vaccination Enhances the Immunogenicity of Subsequent Influenza Vaccination in Healthy Volunteers: A Randomized, Placebo-Controlled Pilot Study. J. Infect. Dis. 2015, 212, 1930–1938. [Google Scholar] [CrossRef]
- Arts, R.J.W.; Blok, B.A.; Aaby, P.; Joosten, L.A.B.; De Jong, D.; Van Der Meer, J.W.M.; Benn, C.S.; Van Crevel, R.; Netea, M.G. Long-term in vitro and in vivo effects of γ-irradiated BCG on innate and adaptive immunity. J. Leukoc. Biol. 2015, 98, 995–1001. [Google Scholar] [CrossRef]
- Giamarellos-Bourboulis, E.J.; Tsilika, M.; Moorlag, S.; Antonakos, N.; Kotsaki, A.; Domínguez-Andrés, J.; Kyriazopoulou, E.; Gkavogianni, T.; Adami, M.-E.; Damoraki, G.; et al. Activate: Randomized Clinical Trial of BCG Vaccination against Infection in the Elderly. Cell 2020, 183, 315–323. [Google Scholar] [CrossRef]
- Nissen, T.N.; Birk, N.M.; Blok, B.A.; Arts, R.J.W.; Andersen, A.; Kjærgaard, J.; Thøstesen, L.M.; Hoffmann, T.; Jeppesen, D.L.; Nielsen, S.D.; et al. Bacillus Calmette-Guérin vaccination at birth and in vitro cytokine responses to non-specific stimulation. A randomized clinical trial. Eur. J. Clin. Microbiol. Infect. Dis. 2018, 37, 29–41. [Google Scholar] [CrossRef]
- Venkatakrishnan, K.; Kramer, W.G.; Synold, T.W.; Goodman, D.B.; Sides, E.; Oliva, C. A pharmacokinetic, pharmacodynamic, and electrocardiographic study of liposomal mifamurtide (L-MTP-PE) in healthy adult volunteers. Eur. J. Clin. Pharmacol. 2012, 68, 1347–1355. [Google Scholar] [CrossRef]
- Leentjens, J.; Quintin, J.; Gerretsen, J.; Kox, M.; Pickkers, P.; Netea, M.G. The Effects of Orally Administered Beta-Glucan on Innate Immune Responses in Humans, a Randomized Open-Label Intervention Pilot-Study. PLoS ONE 2014, 9, e108794. [Google Scholar] [CrossRef] [PubMed]
- Mourits, V.P.; Koeken, V.A.C.M.; De Bree, L.C.J.; Moorlag, S.J.C.F.M.; Chu, W.C.; Xu, X.; Dijkstra, H.; Lemmers, H.; Joosten, L.A.B.; Wang, Y.; et al. BCG-Induced Trained Immunity in Healthy Individuals: The Effect of Plasma Muramyl Dipeptide Concentrations. J. Immunol. Res. 2020, 2020, 5812743. [Google Scholar] [CrossRef]
- Ellouz, F.; Adam, A.; Ciorbaru, R.; Lederer, E. Minimal structural requirements for adjuvant activity of bacterial peptidoglycan derivatives. Biochem. Biophys. Res. Commun. 1974, 59, 1317–1325. [Google Scholar] [CrossRef]
- Huang, Z.; Wang, J.; Xu, X.; Wang, H.; Qiao, Y.; Chu, W.C.; Xu, S.; Chai, L.; Cottier, F.; Pavelka, N.; et al. Antibody neutral-ization of microbiota-derived circulating peptidoglycan dampens inflammation and ameliorates autoimmunity. Nat. Microbiol. 2019, 4, 766–773. [Google Scholar] [CrossRef] [PubMed]
- Jimmy, R.; Stern, C.; Lisy, K.; White, S. Effectiveness of mifamurtide in addition to standard chemotherapy for high-grade osteosarcoma: A systematic review. JBI Evid. Syst. 2017, 15, 2113–2152. [Google Scholar] [CrossRef]
- Liu, Y.-J.; Kobayashi, K.S. Muramyl Dipeptide and its Derivatives: Peptide Adjuvant in Immunological Disorders and Cancer Therapy. Curr. Bioact. Compd. 2011, 7, 180–197. [Google Scholar] [CrossRef]
- Nelson, R.P.; Ballow, M. 26. Immunomodulation and immunotherapy: Drugs, cytokines, cytokine receptors, and antibodies. J. Allergy Clin. Immunol. 2003, 111, S720–S732. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, M.; Jiang, Y.; Li, X.; He, Y.; Zeng, P.; Guo, Z.; Chang, Y.; Luo, H.; Liu, Y.; et al. Lentinan as an immuno-therapeutic for treating lung cancer: A review of 12 years clinical studies in China. J. Cancer Res. Clin. Oncol. 2018, 144, 2177–2186. [Google Scholar] [CrossRef]
- Chihara, G.; Hamuro, J.; Maeda, Y.Y.; Shiio, T.; Suga, T.; Takasuka, N.; Sasaki, T. Antitumor and metastasis-inhibitory ac-tivities of lentinan as an immunomodulator: An overview. Cancer Detect. Prev. Suppl. 1987, 1, 423–443. [Google Scholar] [PubMed]
- Maeda, Y.Y.; Takahama, S.; Kohara, Y.; Yonekawa, H. Two genes controlling acute phase responses by the antitumor polysaccharide, lentinan. Immunogenetics 1996, 43, 215–219. [Google Scholar] [CrossRef]
- Jung, B.-G.; Lee, J.-A.; Lee, B.-J. Immunoprophylactic effects of shiitake mushroom (Lentinula edodes) against Bordetella bronchiseptica in mice. J. Microbiol. 2012, 50, 1003–1008. [Google Scholar] [CrossRef] [PubMed]
- Masihi, K.N.; Madaj, K.; Hintelmann, H.; Gast, G.; Kaneko, Y. Down-regulation of tumor necrosis factor-α, moderate reduction of interleukin-1β, but not interleukin-6 or interleukin-10, by glucan immunomodulators curdlan sulfate and lentinan. Int. J. Immunopharmacol. 1997, 19, 463–468. [Google Scholar] [CrossRef]
- Wang, J. Clinical observation of lentinan in treatment of 60 cases of hepatitis B. J. Chin. Med. 1994, 16, 23–24. [Google Scholar]
- Wu, B.; Zhang, Y.; Zhang, L. III clinical trial summary of lentinan in the treatment of 108 cases with chronic viral hepatitis. Fujian J. Tradit. Chin. Med. 1993, 24, 10–13. [Google Scholar]
- Gordon, M.; Bihari, B.; Goosby, E.; Gorter, R.; Greco, M.; Guralnik, M.; Mimura, T.; Rudinicki, V.; Wong, R.; Kaneko, Y. A placebo-controlled trial of the immune modulator, lentinan, in HIV-positive patients: A phase I/II trial. J. Med. 1998, 29, 305–330. [Google Scholar] [PubMed]
- Sullivan, R.; Smith, J.E.; Rowan, N.J. Medicinal Mushrooms and Cancer Therapy: Translating a traditional practice into Western medicine. Perspect. Biol. Med. 2006, 49, 159–170. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.E.; Rowan, N. Medicinal Mushrooms: Their Therapeutic Properties and Current Medical Usage with Special Emphasis on Cancer Treatments; Cancer Research UK: London, UK, 2002. [Google Scholar]
- De Boer, A.; Bast, A. Demanding safe foods—Safety testing under the novel food regulation (2015/2283). Trends Food Sci. Technol. 2018, 72, 125–133. [Google Scholar] [CrossRef]
- Andrade, E.; Bento, A.; Cavalli, J.; Oliveira, S.; Schwanke, R.; Siqueira, J.; Freitas, C.; Marcon, R.; Calixto, J. Non-clinical studies in the process of new drug development—Part II: Good laboratory practice, metabolism, pharmacokinetics, safety and dose translation to clinical studies. Braz. J. Med. Biol. Res. 2016, 49, e5646. [Google Scholar] [CrossRef]
- Zhou, C.; Liu, M. 30 cases of adverse reaction caused by lentinan. Zhejiang J. Integr. Tradit. Chin. West. Med. 2010, 20, 635. [Google Scholar]
- De Mendonça, C.N.; Silva, P.M.C.E.; Avelleira, J.C.R.; Nishimori, F.S.; Cassia, F.D.F. Shiitake dermatitis. An. Bras. de Dermatol. 2015, 90, 276–278. [Google Scholar] [CrossRef] [PubMed]
- Normile, D. ASIAN MEDICINE: The New Face of Traditional Chinese Medicine. Science 2003, 299, 188–190. [Google Scholar] [CrossRef] [PubMed]
- Wasser, S.P.; Sokolov, D.; Reshetnikov, S.V.; Timor-Tismenetsky, M. Dietary supplements from medicinal mushrooms: Di-versity of types and variety of regulations. Int. J. Med. Mushrooms 2000, 2, 1–19. [Google Scholar] [CrossRef]
- Andrade, E.; Bento, A.; Cavalli, J.; Oliveira, S.; Freitas, C.; Marcon, R.; Schwanke, R.; Siqueira, J.; Calixto, J.B. Non-clinical studies required for new drug development—Part I: Early in silico and in vitro studies, new target discovery and validation, proof of principles and robustness of animal studies. Braz. J. Med. Biol. Res. 2016, 49. [Google Scholar] [CrossRef]
- Ebose, N.; Chan, A.S.H.; Eguerrero, F.; Maristany, C.M.; Eqiu, X.; Walsh, R.M.; Ertelt, K.E.; Jonas, A.B.; Gorden, K.B.; Dudney, C.M.; et al. Binding of Soluble Yeast β-Glucan to Human Neutrophils and Monocytes is Complement-Dependent. Front. Immunol. 2013, 4, 230. [Google Scholar] [CrossRef]
- Bose, N.; Chan, A.S.; Jonas, A.; Qiu, X.; Ottoson, N.R.; Kangas, T.; Graff, J.R. Abstract LB-228: Imprime PGG treatment elicits a coordinated antitumor immune response that triggers enhanced expression of PD-L1 on tumor cells as well as mono-cyte-derived macrophages and dendritic cells. In Proceedings of the AACR 106th Annual Meeting 2015, Philadelphia, PA, USA, 18–22 April 2015. [Google Scholar]
- Fulton, R.B.; Leonardo, S.M.; Michel, K.S.; Danielson, M.E.; Gorden, K.B.; Graff, J.R. Abstract B019: Imprime PGG, a soluble β-glucan, binds to and activates dendritic cells resulting in enhanced T cell priming, expansion, and cytokine production. Drug Development 2016, 4. [Google Scholar] [CrossRef]
- Li, B.; Allendorf, D.J.; Hansen, R.; Marroquin, J.; Ding, C.; Cramer, D.E.; Yan, J. Yeast β-Glucan Amplifies Phagocyte Killing of iC3b-Opsonized Tumor Cells via Complement Receptor 3-Syk-Phosphatidylinositol 3-Kinase Pathway. J. Immunol. 2006, 177, 1661–1669. [Google Scholar] [CrossRef] [PubMed]
- Qi, C.; Cai, Y.; Gunn, L.; Ding, C.; Li, B.; Kloecker, G.; Qian, K.; Vasilakos, J.; Saijo, S.; Iwakura, Y.; et al. Differential pathways regulating innate and adaptive antitumor immune responses by particulate and soluble yeast-derived β-glucans. Blood 2011, 117, 6825–6836. [Google Scholar] [CrossRef]
- Li, B.; Allendorf, D.J.; Hansen, R.; Marroquin, J.; Cramer, D.E.; Harris, C.L.; Yan, J. Combined Yeast β-Glucan and Antitumor Monoclonal Antibody Therapy Requires C5a-Mediated Neutrophil Chemotaxis via Regulation of Decay-Accelerating Factor CD55. Cancer Res. 2007, 67, 7421–7430. [Google Scholar] [CrossRef]
- Jonas, A.; Qiu, X.; Chan, A.; McMurray, D.; Bose, N. Imprime PGG, a yeast beta-glucan immunomodulator, can engage Fc gamma receptor (FcgR) in addition to complement receptor 3 (CR3) on human neutrophils and monocytes. In Proceedings of the Keystone Symposium on Tumor Immunology, Multidisciplinary Science Driving Combination Therapy, Banff, AB, Canada, 8–13 February 2015. [Google Scholar]
- Bose, N.; Jonas, A.; Qiu, X.; Chan, A.S.; Ottoson, N.R.; Graff, J.R. Abstract A015: Imprime PGG treatment enhances anti-body-dependent cellular phagocytosis (ADCP) of tumor cells by monocyte-derived macrophages. In Proceedings of the CRI-CIMT-EATI-AACR Inaugural International Cancer Immunotherapy Conference: Translating Science into Survival, New York, NY, USA, 16–19 September 2016. [Google Scholar]
- Leonardo, S.M.; Fulton, R.B.; Wurst, L.R.; Gorden, K.B.; Jonas, A.B.; Qui, X.; Chan, A.S.; Graff, J.R. Abstract A160: Imprime PGG binds to neutrophils through complement, Fc, and dectin-1 receptors, priming these cells for enhanced ROS production and tumor cell cytotoxicity. Other Topics 2016, 4. [Google Scholar] [CrossRef]
- Chan, A.S.; Qiu, X.; Jonas, A.B.; Patchen, M.L.; Bose, N. Imprime PGG, a yeast β-glucan immunomodulator, has the potential to repolarize human monocyte-derived M2 macrophages to M1 phenotype. J. Immunother. Cancer 2014, 2, P191. [Google Scholar] [CrossRef]
- Chan, A.S.; Qiu, X.; Jonas, A.; Kangas, T.; Ottoson, N.R.; Bose, N. Abstract LB-225: Imprime PGG modulates the function of monocyte-derived M2 macrophages and dendritic cells to drive T-cell expansion. Immunology 2015, 75, LB-225. [Google Scholar]
- Fraser, K.; Ottoson, N.; Qiu, X.; Chan, A.S.; Jonas, A.; Kangas, T.; Graff, J.; Bose, N. Abstract A02: Imprime PGG modulates the myeloid component of the tumor microenvironment to coordinate an antitumor immune response. Immunol. Immunother. 2016, 76, A02. [Google Scholar] [CrossRef]
- Leonardo, S.M.; Gorden, K.; Fulton, R.; Wurst, L. Abstract 5034: Imprime PGG decreases regulatory T cell suppression and enhances T cell proliferation and differentiation revealing additional mechanisms for its anti-tumor activity. Immunology 2015, 75, 5034. [Google Scholar]
- Jonas, A.B.; Chan, A.S.; Qiu, X.; Fraser, K.; Ottoson, N.; Kangas, T.; Walsh, R.; Leonardo, S.M.; Fulton, R.; Gorden, K.; et al. Abstract LB-199: Imprime PGG modulates immunosuppressive myeloid components of the tumor microenvironment and drives enhanced antitumor efficacy in combination with checkpoint inhibitor therapies. Immunology 2017, 77, LB-199. [Google Scholar]
- Chan, A.S.H.; Jonas, A.B.; Qiu, X.; Ottoson, N.R.; Walsh, R.M.; Gorden, K.B.; Harrison, B.; Maimonis, P.J.; Leonardo, S.M.; Ertelt, K.E.; et al. Imprime PGG-Mediated Anti-Cancer Immune Activation Requires Immune Complex Formation. PLoS ONE 2016, 11, e0165909. [Google Scholar] [CrossRef]
- Halstenson, C.E.; Shamp, T.; Gargano, M.A.; Walsh, R.M.; Patchen, M.L. Two randomized, double-blind, placebo-controlled, dose-escalation phase 1 studies evaluating BTH1677, a 1, 3-1,6 beta glucan pathogen associated molecular pattern, in healthy volunteer subjects. Investig. New Drugs 2016, 34, 202–215. [Google Scholar] [CrossRef]
- Bose, N.; Ottoson, N.R.; Qiu, X.; Harrison, B.; Lowe, J.R.; Uhlik, M.T.; Rathmann, B.T.; Kangas, T.O.; Jordan, L.R.; Ertelt, K.E.; et al. Immune Pharmacodynamic Responses of the Novel Cancer Immunotherapeutic Imprime PGG in Healthy Volunteers. J. Immunol. 2019, 202, 2945–2956. [Google Scholar] [CrossRef]
- Segal, N.H.; Gada, P.; Senzer, N.; Gargano, M.A.; Patchen, M.L.; Saltz, L.B. A Phase II Efficacy and Safety, Open-Label, Multicenter Study of Imprime PGG Injection in Combination with Cetuximab in Patients With Stage IV KRAS-Mutant Colorectal Cancer. Clin. Color. Cancer 2016, 15, 222–227. [Google Scholar] [CrossRef] [PubMed]
- Thomas, M.; Sadjadian, P.; Kollmeier, J.; Lowe, J.; Mattson, P.; Trout, J.R.; Gargano, M.; Patchen, M.L.; Walsh, R.; Beliveau, M.; et al. A randomized, open-label, multicenter, phase II study evaluating the efficacy and safety of BTH1677 (1,3–1,6 beta glucan; Imprime PGG) in combination with cetuximab and chemotherapy in patients with advanced non-small cell lung cancer. Investig. New Drugs 2017, 35, 345–358. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Described Changes | Examples |
|---|---|
|
|
|
|
|
|
|
|
|
|
| Claim Type | Nutrient, Substance, Food or Food Category | Claim | Non-Authorization/Discontinuation Based on Criteria | Health Relationship | EFSA Opinion/Journal Reference | Entry ID |
|---|---|---|---|---|---|---|
| Art. 13(1) | Beta-glucan (WGP) | For immunity. Strengthens immunity. | (2) Rejected on the basis of an unclear health relationship or no clear association with health. | Immune system | 2011;9(6):2228 | 1792 |
| Art. 13(1) | Beta-glucan + olive leaf extract | Supports the body’s own defense mechanism/immunity. Maintains natural defense mechanism/immunity. Helps strengthen natural immunity. | (2) Rejected on the basis of an unclear health relationship or no clear association with health. | Immune function/immune system | 2011;9(4):2061 | 1793 |
| Art. 13(1) | Beta-glucan of Saccharomyces cerevisiae | Beta-glucan from yeast as immunomodulators. Beta-glucan from yeast support of natural defenses. | (2) Rejected on the basis of an unclear health relationship or no clear association with health. | Immune system | 2011;9(6):2228 | 847 |
| Art. 13(1) | Beta-glucan of Saccharomyces cerevisiae | Beta-glucan from yeast as immunomodulators. Beta-glucan from yeast support of natural defenses. | (2) Rejected on the basis of an unclear health relationship or no clear association with health. | Increasing nonspecific serum IgA secretion | 2011;9(6):2228 | 1944 |
| Art. 13(1) | WGP beta-glucan; (WGP® (1,3)-b-d-glucan); (from Saccharomyces cerevisiae) | WGP beta-glucan contributes to the normal function to the immune system. WGP beta-glucan naturally contributes to adequate immune responses. The daily dietary supplementation with WGP beta-glucan promotes the normal function of the immune system. WGP beta-glucan enhances the production and activity of macrophages and neutrophils. Thus, it plays an important role in the adequate function of the immune system. WGP beta-glucan contributes to maintain the normal function of the upper respiratory tract. | (3) Rejected on the basis of an unproven cause and effect relationship: no evidence (yet) for a relationship between intake and effect. | Maintenance of the upper respiratory tract defense against pathogens by maintaining immune defenses. | 2011;9(6):2248 | 1910 |
| Art. 13(5) | Yestimun® | Daily administration of Yestimun® helps to maintain the body’s defense against pathogens. | (3) Rejected on the basis of an unproven cause and effect relationship: no evidence (yet) for a relationship between intake and effect. | N/A | Q-2012-00761Commission Regulation (EU) No 1154/2014 of 29/10/2014 | N/A |
| Art. 13(5) | Yestimun®, consisting of (1,3)-(1,6)-β-d-glucans of brewer’s yeast cell wall (100% Saccharomyces cerevisiae) | Daily administration of Yestimun® strengthens the body’s defense during the cold season. | (3) Rejected on the basis of an unproven cause and effect relationship: no evidence (yet) for a relationship between intake and effect. | N/A | Q-2008-667Commission Regulation (EU) 432/2011 of 04/05/2011 | N/A |
| Art. 13(1) | Lentinula edodes (common name: Shiitake) | Contributes to natural immunological defenses. | (3) Rejected on the basis of an unproven cause and effect relationship: no evidence (yet) for a relationship between intake and effect. | Immune function/immune system | 2011;9(4):2061 | 3774 |
| Art. 13(1) | Lentinula edodes (common name: shiitake) | Contributes to natural immunological defenses. | (2) Rejected on the basis of an unclear health relationship or no clear association with health. | Stimulation of immunological responses | 2011;9(4):2061 | 2075 |
| Art. 13(1) | Pleurotus ostreatus (oyster mushroom) | Contributes to natural immunological defenses. | (2) Rejected on the basis of an unclear health relationship or no clear association with health. | Immune function/immune system | 2011;9(4):2061 | 3521 |
| Art. 13(1) | Brewer’s yeast | Strengthens immunity | (2) Rejected on the basis of an unclear health relationship or no clear association with health. | Immune function/immune system | 2010;8(10):1799 | 1384 |
| Art. 13(5) | Immune balance drink, containing vitamin C, green tea, grape skin, grape seed, and shiitake mushroom extract | The immune balance drink activates body’s defense. | (3) Rejected on the basis of an unproven cause and effect relationship: no evidence (yet) for a relationship between intake and effect. | N/A | Q-2009-517Commission Regulation (EU) No 958/2010 of 22/10/2010 | N/A |
| Art. 13(1) | Active hexose correlated compound (AHCC) | Activates immune system, exert potential effects on the immune system—stimulating immunity. | (2) Rejected on the basis of an unclear health relationship or no clear association with health. | Stimulation of immunological responses | 2011;9(4):2061 | 3139 |
| Art. 13(1) | Herbal yeast plasmolysate (Saccharomyces cerevisiae) | Strengthens the body’s defense system. Increases immunity. | (2) Rejected on the basis of an unclear health relationship or no clear association with health. | Immune function/immune system | 2011;9(4):2061 | 1817 |
| Population | Intervention | Conclusions | Ref. |
|---|---|---|---|
| 20 Healthy individuals (age: 20–36 years) | Participants would receive a BCG vaccination from the public health agency for traveling to or working in countries where tuberculosis is prevalent. | The production of TNF-α and IL-1β to mycobacteria or unrelated pathogens was higher after 2 weeks and 3 months post-vaccination, but these effects were less pronounced 1 year after vaccination. However, monocytes recovered 1 year after vaccination had an increased expression of pattern recognition receptors such as CD14, toll-like receptor 4 (TLR4) and mannose receptor, and this correlated with an increase in proinflammatory cytokine production after stimulation with the TLR4 ligand lipopolysaccharide. | [86] |
| 30 Healthy Dutch male participants (age: 19–37 years) | Participants received either BCG (n = 15) or placebo (the diluent used to dissolve BCG) (n = 15). One month after placebo or BCG vaccination, all volunteers received a single dose of yellow fever vaccine. | BCG-vaccinated volunteers displayed a significant reduction of viremia compared to the placebo group, which highly correlated with enhanced IL-1β production. | [87] |
| 20 Healthy, BCG-naive volunteers (age: 18–35 years) | Ten subjects received standard dose (0.1 mL of the reconstituted vaccine) of intradermal BCG vaccination 5 weeks prior to challenge infection. Ten controls received no vaccination. Five weeks after BCG vaccination, both groups were exposed to bites of five Plasmodium falciparum NF54 strain infected Anopheles stephensi mosquitoes (sporozoite challenge). | BCG vaccination altered some of the clinical, immunological, and parasitological outcomes of malaria infection in a subset of volunteers. Earlier NK cell and monocyte activation in this subset of vaccinated volunteers is consistent with the possibility that induction of trained innate immunity in vivo may have functional activity against a heterologous pathogen in humans. | [88] |
| 212 Neonates; BCG vaccinated (n = 119) BCG naïve (n = 93) | Participants were randomized 1:1 to undergo vaccination with BCG (0.05 mL) intradermally within 10 days of birth or to receive no BCG vaccine. | BCG-vaccinated infants had increased production of IL-6 in unstimulated samples and decreased production of interleukin 1 receptor antagonist, IL-6, and IL-10 and the chemokines macrophage inflammatory protein 1α (MIP-1α), MIP-1β, and monocyte chemoattractant protein 1 (MCP-1) following stimulation with peptidoglycan (TLR2) and R848 (TLR7/8). BCG-vaccinated infants also had decreased MCP-1 responses following stimulation with heterologous pathogens. | [89] |
| 40 Healthy volunteers (age: 20–25 years) | Participants received either live attenuated BCG vaccine (n = 20) or placebo (n = 20), followed by intramuscular injection of trivalent influenza vaccine 14 days later. | In BCG-vaccinated subjects, HI antibody responses against the 2009 pandemic influenza A(H1N1) vaccine strain were significantly enhanced compared with the placebo group. Additionally, apart from enhanced proinflammatory leukocyte responses following BCG vaccination, nonspecific effects of influenza vaccination were also observed, with modulation of cytokine responses against unrelated pathogens. | [90] |
| 15 Healthy individuals (age: 20–34 years) | Participants received inactivated gamma-irradiated BCG (γBCG). The inactivated BCG was cultured for 6 weeks to confirm inactivation. | γBCG vaccination in volunteers had only minimal effects on innate immunity. The results indicate that γBCG induces long-term training of innate immunity in vitro. In vivo, γBCG induces effects on innate cytokine production are limited. | [91] |
| 198 Elderly patients (age >65 years) | Participants received BCG (n = 100) or placebo (n = 98) vaccine and were followed for 12 months for new infections. | At interim analysis, BCG vaccination significantly increased the time to first infection. The incidence of new infections was 42.3% after placebo vaccination and 25.0% after BCG vaccination; most of the protection was against respiratory tract infections of probable viral origin. | [92] |
| 158 Infants | Infants received BCG within 7 days of birth (n = 80). Controls (n = 78) were bled 4 days post-randomization, and at age 3 and 13 months. | BCG vaccination of Danish newborns did not induce nonspecific in vitro cytokine responses. | [93] |
| 21 Healthy participants(age: 21–59 years) | Participants received a single 4 mg i.v. infusion of L-MTP-PE over 30 min. | Serum concentrations of IL-6, TNF-α, and CRP increased following L-MTP-PE infusion. Maximum observed increases in IL-6 and TNF-α occurred at 4 and 2 h, respectively, returning toward baseline by 8 h post-dose. | [94] |
| 15 Healthy male participants(age: 19–24 years) | Beta-glucan (n = 10) or the control group (n = 5). Subjects in the glucan group ingested beta-glucan 1000 mg once daily for 7 days. Water-insoluble beta-glucan derived from baker’s yeast (S. cerevisiae) sold as a dietary supplement (Glucan #300, BG). This preparation has a purity of at least 83% guaranteed by the manufacturer. | Beta-glucan was barely detectable in serum of volunteers at all time points. Neither cytokine production nor microbicidal activity of leukocytes were affected by orally administered beta-glucan. | [95] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
van Steenwijk, H.P.; Bast, A.; de Boer, A. Immunomodulating Effects of Fungal Beta-Glucans: From Traditional Use to Medicine. Nutrients 2021, 13, 1333. https://doi.org/10.3390/nu13041333
van Steenwijk HP, Bast A, de Boer A. Immunomodulating Effects of Fungal Beta-Glucans: From Traditional Use to Medicine. Nutrients. 2021; 13(4):1333. https://doi.org/10.3390/nu13041333
Chicago/Turabian Stylevan Steenwijk, Hidde P., Aalt Bast, and Alie de Boer. 2021. "Immunomodulating Effects of Fungal Beta-Glucans: From Traditional Use to Medicine" Nutrients 13, no. 4: 1333. https://doi.org/10.3390/nu13041333
APA Stylevan Steenwijk, H. P., Bast, A., & de Boer, A. (2021). Immunomodulating Effects of Fungal Beta-Glucans: From Traditional Use to Medicine. Nutrients, 13(4), 1333. https://doi.org/10.3390/nu13041333