
Vitamin D Supplementation and Impact on Skeletal Muscle Function in Cell and Animal Models and an Aging Population: What Do We Know So Far?



Abstract
:1. Introduction
2. Methods
2.1. Search Strategy
2.2. Inclusion Criteria
2.3. Exclusion Criteria
2.4. Data Extraction
3. Results and Discussion
3.1. Cell Lines
3.2. Animal Studies
3.2.1. In Vitro Cell and Animal Studies
Myotube Formation, Muscle Mass, Strength and Force
Muscle function and protein synthesis
Mitochondria and Lipid Metabolism
Glucose and Insulin Metabolism
Oxidative Stress, AGES
3.3. Human Studies
Muscle Mass, Strength and Function
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Shea, M.K.; Fielding, R.A.; Dawson-Hughes, B. The effect of vitamin D supplementation on lower-extremity power and function in older adults: A randomized controlled trial. Am. J. Clin. Nutr. 2019, 109, 369–379. [Google Scholar] [CrossRef] [PubMed]
- Meehan, M.; Penckofer, S. The Role of Vitamin D in the Aging Adult. J. Aging Gerontol. 2014, 2, 60–71. [Google Scholar] [CrossRef]
- Cuellar, W.A.; Blizzard, L.; Hides, J.A.; Callisaya, M.L.; Jones, G.; Cicuttini, F.; Wluka, A.E.; Ding, C.; Winzenberg, T.M. Vitamin D supplements for trunk muscle morphology in older adults: Secondary analysis of a randomized controlled trial. J. Cachexia Sarcopenia Muscle 2019, 10, 177–187. [Google Scholar] [CrossRef]
- Santilli, V.; Bernetti, A.; Mangone, M.; Paoloni, M. Clinical definition of sarcopenia. Clin. Cases Miner. Bone Metab. 2014, 11, 177–180. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Jentoft, A.J.; Baeyens, J.P.; Bauer, J.M.; Boirie, Y.; Cederholm, T.; Landi, F.; Martin, F.C.; Michel, J.P.; Rolland, Y.; Schneider, S.M.; et al. Sarcopenia: European consensus on definition and diagnosis: Report of the European Working Group on Sarcopenia in Older People. Age Ageing 2010, 39, 412–423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rizzoli, R.; Reginster, J.Y.; Arnal, J.F.; Bautmans, I.; Beaudart, C.; Bischoff-Ferrari, H.; Biver, E.; Boonen, S.; Brandi, M.L.; Chines, A.; et al. Quality of life in sarcopenia and frailty. Calcif. Tissue Int. 2013, 93, 101–120. [Google Scholar] [CrossRef] [PubMed]
- Wimalawansa, S.J. Associations of vitamin D with insulin resistance, obesity, type 2 diabetes, and metabolic syndrome. J. Steroid Biochem. Mol. Biol. 2018, 175, 177–189. [Google Scholar] [CrossRef] [PubMed]
- Robinson, S.M.; Reginster, J.Y.; Rizzoli, R.; Shaw, S.C.; Kanis, J.A.; Bautmans, I.; Bischoff-Ferrari, H.; Bruyere, O.; Cesari, M.; Dawson-Hughes, B.; et al. Does nutrition play a role in the prevention and management of sarcopenia? Clin. Nutr. 2018, 37, 1121–1132. [Google Scholar] [CrossRef] [Green Version]
- Bislev, L.S.; Langagergaard Rodbro, L.; Rolighed, L.; Sikjaer, T.; Rejnmark, L. Effects of Vitamin D3 Supplementation on Muscle Strength, Mass, and Physical Performance in Women with Vitamin D Insufficiency: A Randomized Placebo-Controlled Trial. Calcif. Tissue Int. 2018, 103, 483–493. [Google Scholar] [CrossRef]
- Girgis, C.M.; Clifton-Bligh, R.J.; Hamrick, M.W.; Holick, M.F.; Gunton, J.E. The roles of vitamin D in skeletal muscle: Form, function, and metabolism. Endocr. Rev. 2013, 34, 33–83. [Google Scholar] [CrossRef] [Green Version]
- Chang, E. 1,25-Dihydroxyvitamin D Decreases Tertiary Butyl-Hydrogen Peroxide-Induced Oxidative Stress and Increases AMPK/SIRT1 Activation in C2C12 Muscle Cells. Molecules 2019, 24, 3903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, E.; Kim, Y. Vitamin D Ameliorates Fat Accumulation with AMPK/SIRT1 Activity in C2C12 Skeletal Muscle Cells. Nutrients 2019, 11, 2806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benetti, E.; Mastrocola, R.; Chiazza, F.; Nigro, D.; D’Antona, G.; Bordano, V.; Fantozzi, R.; Aragno, M.; Collino, M.; Minetto, M.A. Effects of vitamin D on insulin resistance and myosteatosis in diet-induced obese mice. PLoS ONE 2018, 13, e0189707. [Google Scholar] [CrossRef]
- Fan, Y.; Futawaka, K.; Koyama, R.; Fukuda, Y.; Hayashi, M.; Imamoto, M.; Miyawaki, T.; Kasahara, M.; Tagami, T.; Moriyama, K. Vitamin D3/VDR resists diet-induced obesity by modulating UCP3 expression in muscles. J. Biomed. Sci. 2016, 23, 56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jefferson, G.E.; Schnell, D.M.; Thomas, D.T.; Bollinger, L.M. Calcitriol concomitantly enhances insulin sensitivity and alters myocellular lipid partitioning in high fat-treated skeletal muscle cells. J. Physiol. Biochem. 2017, 73, 613–621. [Google Scholar] [CrossRef] [Green Version]
- Taneja, N. Mass Spectrometric Analysis of Proteins of L6 Skeletal Muscle Cells Under Different Glucose Conditions and Vitamin D Supplementation. Protei. Pept. Lett. 2018, 25, 356–361. [Google Scholar] [CrossRef]
- Romeu Montenegro, K.; Carlessi, R.; Cruzat, V.; Newsholme, P. Effects of vitamin D on primary human skeletal muscle cell proliferation, differentiation, protein synthesis and bioenergetics. J. Steroid Biochem. Mol. Biol. 2019, 193, 105423. [Google Scholar] [CrossRef]
- Antinozzi, C.; Marampon, F.; Sgro, P.; Tombolini, V.; Lenzi, A.; Crescioli, C.; Di Luigi, L. Comparative study of testosterone and vitamin D analogue, elocalcitol, on insulin-controlled signal transduction pathway regulation in human skeletal muscle cells. J. Endocrinol. Investig. 2019, 42, 897–907. [Google Scholar] [CrossRef]
- Kuwabara, A.; Tsugawa, N.; Kondo, H.; Ao, M.; Fujiwara, H.; Hosokawa, N.; Matsumoto, S.; Tanaka, K.; Nakano, T. Associations between serum 25-hydroxyvitamin D3 level and skeletal muscle mass and lower limb muscle strength in Japanese middle-aged subjects. Osteoporos. Sarcopenia 2017, 3, 53–58. [Google Scholar] [CrossRef]
- Pirotta, S.; Kidgell, D.J.; Daly, R.M. Effects of vitamin D supplementation on neuroplasticity in older adults: A double-blinded, placebo-controlled randomised trial. Osteoporos. Int. 2015, 26, 131–140. [Google Scholar] [CrossRef] [Green Version]
- Grimnes, G.; Kubiak, J.; Jorde, R. Four months vitamin D supplementation to vitamin D insufficient individuals does not improve muscular strength: A randomized controlled trial. PLoS ONE 2019, 14, e0225600. [Google Scholar] [CrossRef] [PubMed]
- Girgis, C.M.; Baldock, P.A.; Downes, M. Vitamin D, muscle and bone: Integrating effects in development, aging and injury. Mol. Cell Endocrinol. 2015, 410, 3–10. [Google Scholar] [CrossRef]
- Garcia, M.; Seelaender, M.; Sotiropoulos, A.; Coletti, D.; Lancha, A.H., Jr. Vitamin D, muscle recovery, sarcopenia, cachexia, and muscle atrophy. Nutrition 2019, 60, 66–69. [Google Scholar] [CrossRef]
- Chiang, C.M.; Ismaeel, A.; Griffis, R.B.; Weems, S. Effects of Vitamin D Supplementation on Muscle Strength in Athletes: A Systematic Review. J. Strength Cond. Res. 2017, 31, 566–574. [Google Scholar] [CrossRef] [PubMed]
- Girgis, C.M.; Clifton-Bligh, R.J.; Turner, N.; Lau, S.L.; Gunton, J.E. Effects of vitamin D in skeletal muscle: Falls, strength, athletic performance and insulin sensitivity. Clin. Endocrinol. 2014, 80, 169–181. [Google Scholar] [CrossRef]
- Braga, M.; Simmons, Z.; Norris, K.C.; Ferrini, M.G.; Artaza, J.N. Vitamin D induces myogenic differentiation in skeletal muscle derived stem cells. Endocr. Connect. 2017, 6, 139–150. [Google Scholar] [CrossRef] [PubMed]
- Hosoyama, T.; Iida, H.; Kawai-Takaishi, M.; Watanabe, K. Vitamin D Inhibits Myogenic Cell Fusion and Expression of Fusogenic Genes. Nutrients 2020, 12, 2192. [Google Scholar] [CrossRef]
- Hayakawa, N.; Fukumura, J.; Yasuno, H.; Fujimoto-Ouchi, K.; Kitamura, H. 1alpha,25(OH)2D3 downregulates gene expression levels of muscle ubiquitin ligases MAFbx and MuRF1 in human myotubes. Biomed. Res. 2015, 36, 71–80. [Google Scholar] [CrossRef] [Green Version]
- van der Meijden, K.; Bravenboer, N.; Dirks, N.F.; Heijboer, A.C.; den Heijer, M.; de Wit, G.M.; Offringa, C.; Lips, P.; Jaspers, R.T. Effects of 1,25(OH)2 D3 and 25(OH)D3 on C2C12 Myoblast Proliferation, Differentiation, and Myotube Hypertrophy. J. Cell. Physiol. 2016, 231, 2517–2528. [Google Scholar] [CrossRef] [Green Version]
- Antinozzi, C.; Corinaldesi, C.; Giordano, C.; Pisano, A.; Cerbelli, B.; Migliaccio, S.; Di Luigi, L.; Stefanantoni, K.; Vannelli, G.B.; Minisola, S.; et al. Potential role for the VDR agonist elocalcitol in metabolic control: Evidences in human skeletal muscle cells. J. Steroid Biochem. Mol. Biol. 2017, 167, 169–181. [Google Scholar] [CrossRef]
- Hirose, Y.; Onishi, T.; Miura, S.; Hatazawa, Y.; Kamei, Y. Vitamin D Attenuates FOXO1-Target Atrophy Gene Expression in C2C12 Muscle Cells. J. Nutr. Sci. Vitaminol. 2018, 64, 229–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arakawa, M.; Wagatsuma, A. 1alpha, 25(OH)2D3 regulates agrin-induced acetylcholine receptor clustering through upregulation of rapsyn expression in C2C12 myotubes. Biochem. Biophys. Res. Commun 2020. [Google Scholar] [CrossRef]
- Ryan, K.J.; Daniel, Z.C.; Craggs, L.J.; Parr, T.; Brameld, J.M. Dose-dependent effects of vitamin D on transdifferentiation of skeletal muscle cells to adipose cells. J. Endocrinol. 2013, 217, 45–58. [Google Scholar] [CrossRef] [Green Version]
- Schnell, D.M.; Walton, R.G.; Vekaria, H.J.; Sullivan, P.G.; Bollinger, L.M.; Peterson, C.A.; Thomas, D.T. Vitamin D produces a perilipin 2-dependent increase in mitochondrial function in C2C12 myotubes. J. Nutr. Biochem. 2019, 65, 83–92. [Google Scholar] [CrossRef]
- Manna, P.; Achari, A.E.; Jain, S.K. 1,25(OH)2-vitamin D3 upregulates glucose uptake mediated by SIRT1/IRS1/GLUT4 signaling cascade in C2C12 myotubes. Mol. Cell. Biochem. 2018, 444, 103–108. [Google Scholar] [CrossRef]
- Tamilselvan, B.; Seshadri, K.G.; Venkatraman, G. Role of vitamin D on the expression of glucose transporters in L6 myotubes. Indian J. Endocrinol. Metab. 2013, 17, S326–S328. [Google Scholar] [CrossRef]
- Tanaka, K.; Kanazawa, I.; Yamaguchi, T.; Yano, S.; Kaji, H.; Sugimoto, T. Active vitamin D possesses beneficial effects on the interaction between muscle and bone. Biochem. Biophys. Res. Commun. 2014, 450, 482–487. [Google Scholar] [CrossRef] [PubMed]
- Nonaka, K.; Akiyama, J.; Yoshikawa, Y.; Une, S.; Ito, K. 1,25-Dihydroxyvitamin D3 Inhibits Lipopolysaccharide-Induced Interleukin-6 Production by C2C12 Myotubes. Medicina 2020, 56, 450. [Google Scholar] [CrossRef]
- Ray, A.D.; Personius, K.E.; Williamson, D.L.; Dungan, C.M.; Dhillon, S.S.; Hershberger, P.A. Vitamin D3 intake modulates diaphragm but not peripheral muscle force in young mice. J. Appl. Physiol. 2016, 120, 1124–1131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trovato, F.M.; Castrogiovanni, P.; Szychlinska, M.A.; Purrello, F.; Musumeci, G. Impact of Western and Mediterranean Diets and Vitamin D on Muscle Fibers of Sedentary Rats. Nutrients 2018, 10, 231. [Google Scholar] [CrossRef] [Green Version]
- Hayes, A.; Rybalka, E.; Debruin, D.A.; Hanson, E.D.; Scott, D.; Sanders, K. The Effect of Yearly-Dose Vitamin D Supplementation on Muscle Function in Mice. Nutrients 2019, 11, 1097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alkharfy, K.M.; Al-Daghri, N.M.; Ahmed, M.; Yakout, S.M. Effects of vitamin D treatment on skeletal muscle histology and ultrastructural changes in a rodent model. Molecules 2012, 17, 9081–9089. [Google Scholar] [CrossRef]
- Gifondorwa, D.J.; Thompson, T.D.; Wiley, J.; Culver, A.E.; Shetler, P.K.; Rocha, G.V.; Ma, Y.L.; Krishnan, V.; Bryant, H.U. Vitamin D and/or calcium deficient diets may differentially affect muscle fiber neuromuscular junction innervation. Muscle Nerve 2016, 54, 1120–1132. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, S.; Sato, Y.; Kobayashi, T.; Kaneko, Y.; Ito, E.; Soma, T.; Okada, H.; Miyamoto, K.; Oya, A.; Matsumoto, M.; et al. Vitamin D protects against immobilization-induced muscle atrophy via neural crest-derived cells in mice. Sci. Rep. 2020, 10, 12242. [Google Scholar] [CrossRef] [PubMed]
- Chanet, A.; Salles, J.; Guillet, C.; Giraudet, C.; Berry, A.; Patrac, V.; Domingues-Faria, C.; Tagliaferri, C.; Bouton, K.; Bertrand-Michel, J.; et al. Vitamin D supplementation restores the blunted muscle protein synthesis response in deficient old rats through an impact on ectopic fat deposition. J. Nutr. Biochem. 2017, 46, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Nadimi, H.; Djazayery, A.; Javanbakht, M.H.; Dehpour, A.; Ghaedi, E.; Derakhshanian, H.; Mohammadi, H.; Zarei, M.; Djalali, M. The Effect of Vitamin D Supplementation on Serum and Muscle Irisin Levels, and FNDC5 Expression in Diabetic Rats. Rep. Biochem. Mol. Biol. 2019, 8, 236–243. [Google Scholar] [PubMed]
- Xavier, S.; Sadanandan, J.; George, N.; Paulose, C.S. beta(2)-adrenoceptor and insulin receptor expression in the skeletal muscle of streptozotocin induced diabetic rats: Antagonism by vitamin D(3) and curcumin. Eur. J. Pharmacol. 2012, 687, 14–20. [Google Scholar] [CrossRef]
- Kim, D.H.; Klemp, A.; Salazar, G.; Hwang, H.S.; Yeh, M.; Panton, L.B.; Kim, J.S. High-dose vitamin D administration and resistance exercise training attenuate the progression of obesity and improve skeletal muscle function in obese p62-deficient mice. Nutr. Res. 2020. [Google Scholar] [CrossRef]
- Akagawa, M.; Miyakoshi, N.; Kasukawa, Y.; Ono, Y.; Yuasa, Y.; Nagahata, I.; Sato, C.; Tsuchie, H.; Nagasawa, H.; Hongo, M.; et al. Effects of activated vitamin D, alfacalcidol, and low-intensity aerobic exercise on osteopenia and muscle atrophy in type 2 diabetes mellitus model rats. PLoS ONE 2018, 13, e0204857. [Google Scholar] [CrossRef] [Green Version]
- Burd, N.A.; Gorissen, S.H.; van Loon, L.J. Anabolic resistance of muscle protein synthesis with aging. Exerc. Sport Sci. Rev. 2013, 41, 169–173. [Google Scholar] [CrossRef]
- Janssen, I.; Heymsfield, S.B.; Wang, Z.M.; Ross, R. Skeletal muscle mass and distribution in 468 men and women aged 18-88 yr. J. Appl. Physiol. 2000, 89, 81–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rizzoli, R.; Stevenson, J.C.; Bauer, J.M.; van Loon, L.J.; Walrand, S.; Kanis, J.A.; Cooper, C.; Brandi, M.L.; Diez-Perez, A.; Reginster, J.Y.; et al. The role of dietary protein and vitamin D in maintaining musculoskeletal health in postmenopausal women: A consensus statement from the European Society for Clinical and Economic Aspects of Osteoporosis and Osteoarthritis (ESCEO). Maturitas 2014, 79, 122–132. [Google Scholar] [CrossRef] [PubMed]
- Yoon, M.S. mTOR as a Key Regulator in Maintaining Skeletal Muscle Mass. Front. Physiol. 2017, 8, 788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lluis, F.; Perdiguero, E.; Nebreda, A.R.; Munoz-Canoves, P. Regulation of skeletal muscle gene expression by p38 MAP kinases. Trends Cell Biol. 2006, 16, 36–44. [Google Scholar] [CrossRef] [PubMed]
- Galluzzo, P.; Rastelli, C.; Bulzomi, P.; Acconcia, F.; Pallottini, V.; Marino, M. 17beta-Estradiol regulates the first steps of skeletal muscle cell differentiation via ER-alpha-mediated signals. Am. J. Physiol. Cell Physiol. 2009, 297, C1249–C1262. [Google Scholar] [CrossRef] [PubMed]
- Clapham, J.C.; Arch, J.R.; Chapman, H.; Haynes, A.; Lister, C.; Moore, G.B.; Piercy, V.; Carter, S.A.; Lehner, I.; Smith, S.A.; et al. Mice overexpressing human uncoupling protein-3 in skeletal muscle are hyperphagic and lean. Nature 2000, 406, 415–418. [Google Scholar] [CrossRef]
- Ryder, J.W.; Gilbert, M.; Zierath, J.R. Skeletal muscle and insulin sensitivity: Pathophysiological alterations. Front. Biosci. 2001, 6, D154–D163. [Google Scholar] [CrossRef] [Green Version]
- Manna, P.; Jain, S.K. Vitamin D up-regulates glucose transporter 4 (GLUT4) translocation and glucose utilization mediated by cystathionine-gamma-lyase (CSE) activation and H2S formation in 3T3L1 adipocytes. J. Biol. Chem. 2012, 287, 42324–42332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhatti, J.S.; Bhatti, G.K.; Reddy, P.H. Mitochondrial dysfunction and oxidative stress in metabolic disorders—A step towards mitochondria based therapeutic strategies. Biochim. Biophys. Acta Mol. Basis Dis. 2017, 1863, 1066–1077. [Google Scholar] [CrossRef] [PubMed]
- Mirhosseini, N.; Vatanparast, H.; Mazidi, M.; Kimball, S.M. Vitamin D Supplementation, Glycemic Control, and Insulin Resistance in Prediabetics: A Meta-Analysis. J. Endocr. Soc. 2018, 2, 687–709. [Google Scholar] [CrossRef] [Green Version]
- Remelli, F.; Vitali, A.; Zurlo, A.; Volpato, S. Vitamin D Deficiency and Sarcopenia in Older Persons. Nutrients 2019, 11, 2861. [Google Scholar] [CrossRef] [Green Version]
- Singh, R.; Barden, A.; Mori, T.; Beilin, L. Advanced glycation end-products: A review. Diabetologia 2001, 44, 129–146. [Google Scholar] [CrossRef] [Green Version]
- El Hajj, C.; Fares, S.; Chardigny, J.M.; Boirie, Y.; Walrand, S. Vitamin D supplementation and muscle strength in pre-sarcopenic elderly Lebanese people: A randomized controlled trial. Arch. Osteoporos. 2018, 14, 4. [Google Scholar] [CrossRef] [PubMed]
- Vaes, A.M.M.; Tieland, M.; Toussaint, N.; Nilwik, R.; Verdijk, L.B.; van Loon, L.J.C.; de Groot, L. Cholecalciferol or 25-Hydroxycholecalciferol Supplementation Does Not Affect Muscle Strength and Physical Performance in Prefrail and Frail Older Adults. J. Nutr. 2018, 148, 712–720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Vliet, S.; Fappi, A.; Reeds, D.N.; Mittendorfer, B. No independent or combined effects of vitamin D and conjugated linoleic acids on muscle protein synthesis in older adults: A randomized, double-blind, placebo-controlled clinical trial. Am. J. Clin. Nutr. 2020, 112, 1382–1389. [Google Scholar] [CrossRef] [PubMed]
- Ceglia, L.; Niramitmahapanya, S.; da Silva Morais, M.; Rivas, D.A.; Harris, S.S.; Bischoff-Ferrari, H.; Fielding, R.A.; Dawson-Hughes, B. A randomized study on the effect of vitamin D(3) supplementation on skeletal muscle morphology and vitamin D receptor concentration in older women. J. Clin. Endocrinol. Metab. 2013, 98, E1927–E1935. [Google Scholar] [CrossRef] [Green Version]
- Latham, N.K.; Anderson, C.S.; Lee, A.; Bennett, D.A.; Moseley, A.; Cameron, I.D.; Fitness Collaborative, G. A randomized, controlled trial of quadriceps resistance exercise and vitamin D in frail older people: The Frailty Interventions Trial in Elderly Subjects (FITNESS). J. Am. Geriatr. Soc. 2003, 51, 291–299. [Google Scholar] [CrossRef]
- Dhesi, J.K.; Jackson, S.H.; Bearne, L.M.; Moniz, C.; Hurley, M.V.; Swift, C.G.; Allain, T.J. Vitamin D supplementation improves neuromuscular function in older people who fall. Age Ageing 2004, 33, 589–595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sorensen, O.H.; Lund, B.; Saltin, B.; Lund, B.; Andersen, R.B.; Hjorth, L.; Melsen, F.; Mosekilde, L. Myopathy in bone loss of ageing: Improvement by treatment with 1 alpha-hydroxycholecalciferol and calcium. Clin. Sci. 1979, 56, 157–161. [Google Scholar] [CrossRef]
- Sato, Y.; Iwamoto, J.; Kanoko, T.; Satoh, K. Low-dose vitamin D prevents muscular atrophy and reduces falls and hip fractures in women after stroke: A randomized controlled trial. Cerebrovasc. Dis. 2005, 20, 187–192, Retracted in Cerebrovasc. Dis. 2017, 44, 240. [Google Scholar] [CrossRef]
- Beaudart, C.; Buckinx, F.; Rabenda, V.; Gillain, S.; Cavalier, E.; Slomian, J.; Petermans, J.; Reginster, J.Y.; Bruyere, O. The effects of vitamin D on skeletal muscle strength, muscle mass, and muscle power: A systematic review and meta-analysis of randomized controlled trials. J. Clin. Endocrinol. Metab. 2014, 99, 4336–4345. [Google Scholar] [CrossRef] [Green Version]
- Wood, A.D.; Secombes, K.R.; Thies, F.; Aucott, L.S.; Black, A.J.; Reid, D.M.; Mavroeidi, A.; Simpson, W.G.; Fraser, W.D.; Macdonald, H.M. A parallel group double-blind RCT of vitamin D3 assessing physical function: Is the biochemical response to treatment affected by overweight and obesity? Osteoporos. Int. 2014, 25, 305–315. [Google Scholar] [CrossRef] [PubMed]
- Houston, D.K.; Tooze, J.A.; Neiberg, R.H.; Hausman, D.B.; Johnson, M.A.; Cauley, J.A.; Bauer, D.C.; Cawthon, P.M.; Shea, M.K.; Schwartz, G.G.; et al. 25-hydroxyvitamin D status and change in physical performance and strength in older adults: The Health, Aging, and Body Composition Study. Am. J. Epidemiol. 2012, 176, 1025–1034. [Google Scholar] [CrossRef] [PubMed]
- Wicherts, I.S.; van Schoor, N.M.; Boeke, A.J.; Visser, M.; Deeg, D.J.; Smit, J.; Knol, D.L.; Lips, P. Vitamin D status predicts physical performance and its decline in older persons. J. Clin. Endocrinol. Metab. 2007, 92, 2058–2065. [Google Scholar] [CrossRef] [PubMed]
- Houston, D.K.; Tooze, J.A.; Davis, C.C.; Chaves, P.H.; Hirsch, C.H.; Robbins, J.A.; Arnold, A.M.; Newman, A.B.; Kritchevsky, S.B. Serum 25-hydroxyvitamin D and physical function in older adults: The Cardiovascular Health Study All Stars. J. Am. Geriatr. Soc. 2011, 59, 1793–1801. [Google Scholar] [CrossRef]
- Kuchuk, N.O.; Pluijm, S.M.; van Schoor, N.M.; Looman, C.W.; Smit, J.H.; Lips, P. Relationships of serum 25-hydroxyvitamin D to bone mineral density and serum parathyroid hormone and markers of bone turnover in older persons. J. Clin. Endocrinol. Metab. 2009, 94, 1244–1250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfeifer, M.; Begerow, B.; Minne, H.W.; Suppan, K.; Fahrleitner-Pammer, A.; Dobnig, H. Effects of a long-term vitamin D and calcium supplementation on falls and parameters of muscle function in community-dwelling older individuals. Osteoporos. Int. 2009, 20, 315–322. [Google Scholar] [CrossRef] [PubMed]
- Powe, C.E.; Evans, M.K.; Wenger, J.; Zonderman, A.B.; Berg, A.H.; Nalls, M.; Tamez, H.; Zhang, D.; Bhan, I.; Karumanchi, S.A.; et al. Vitamin D-binding protein and vitamin D status of black Americans and white Americans. N. Engl. J. Med. 2013, 369, 1991–2000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herbert, R.; Moseley, A.; Sherrington, C. PEDro: A database of randomised controlled trials in physiotherapy. Health Inf. Manag. 1998, 28, 186–188. [Google Scholar] [CrossRef]
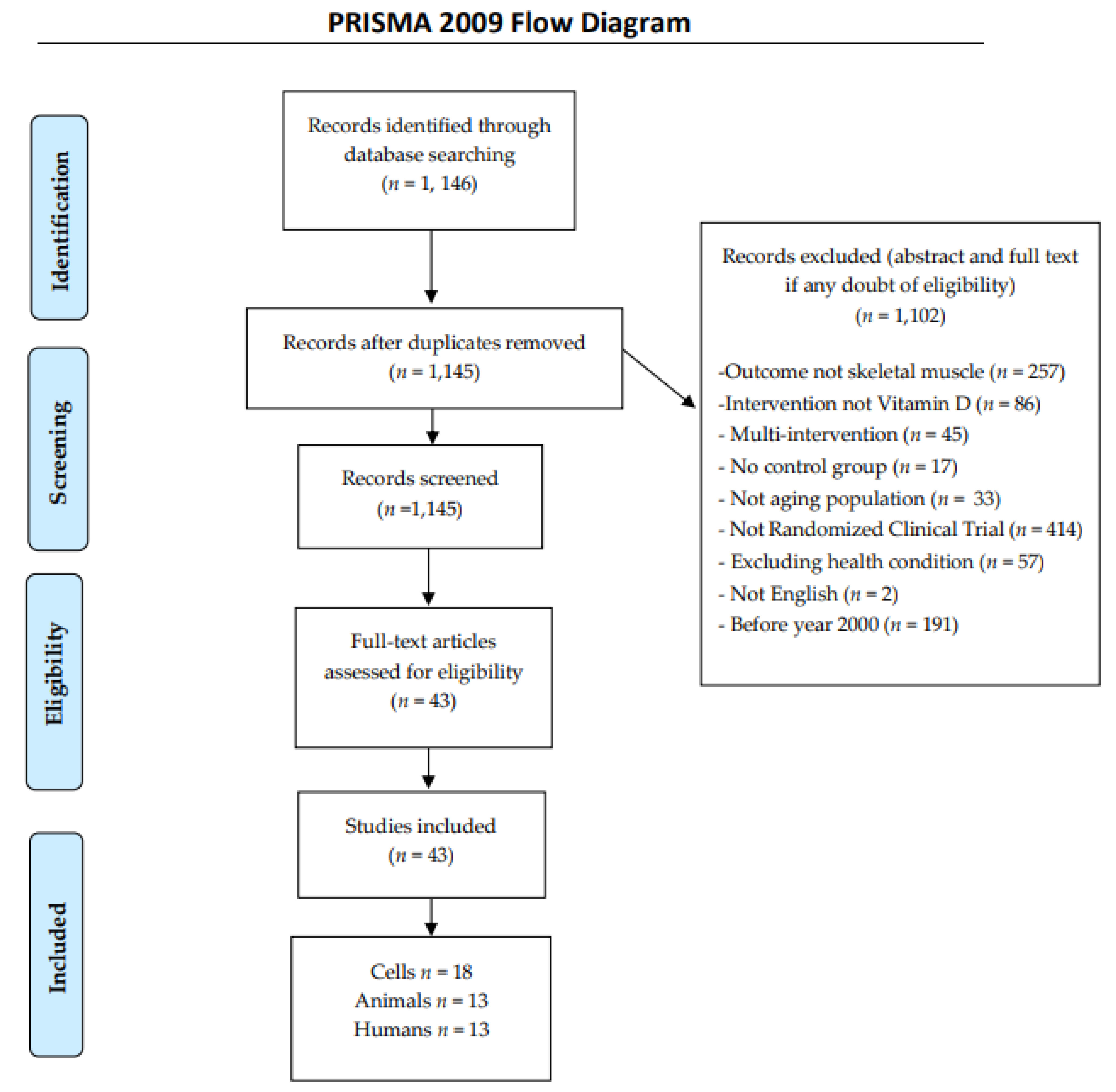

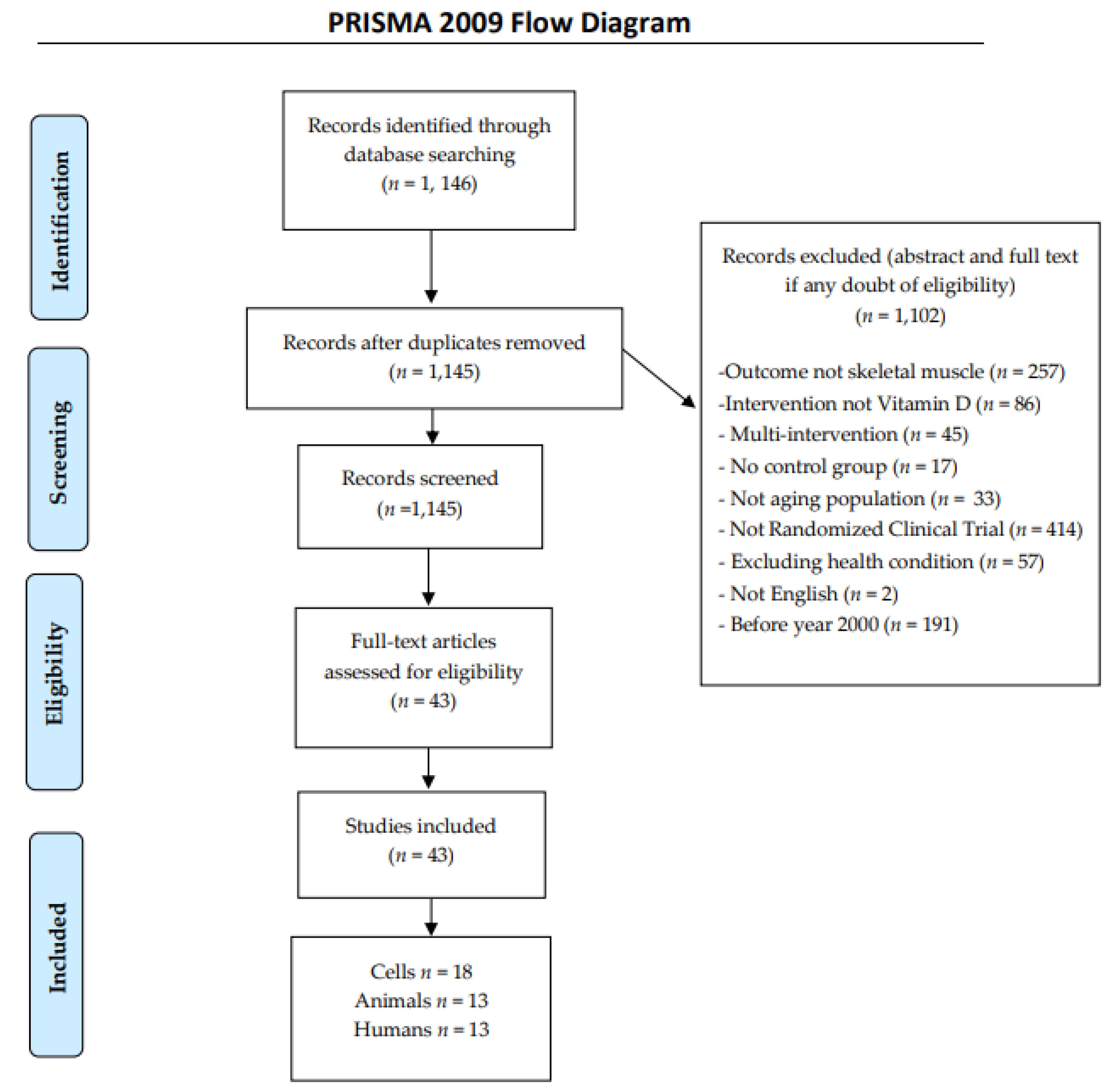{kind=link}
| The authors, Year | Cell Line/Type | Maturation | VitD Dose, Form, Time | Significant Findings and Effects | Comments |
|---|---|---|---|---|---|
| Myotube formation, muscle mass, strength and force | |||||
| Braga et al., 2017 [26] | Mice satellite cells | Myoblasts | 100 nM, 1,25(OH)2D, 1–12d | ↑ MYOD, MYOG, MYC2, skeletal muscle fast troponin I and T, MYH1, IGF1 IGF2, FGF1 and 2, BMP4, MMP9 and FST. | VitD3 promoted a robust myogenic effect on satellite cells responsible for the regeneration of muscle after injury or muscle waste. |
| Romeu et al., 2019 [17] | HSMM | Myoblasts and myotubes | 100 nM, 1,25(OH)2D, 48 h/or 5 d | ↑ differentiation by altering the expression of myogenic regulatory factors. ↑ protein synthesis signalling and synthesis (AKT, mTOR, GSK3B); ↑ OCR in myoblasts and myotubes. | At day 6, there were notably higher number and diameter of myotubes per field in VitD3 group when compared with vehicle group. |
| Hosoyama et al., 2020 [27] | Mouse Ric10 and human myogenic cell clone Hu5KD3 | Myoblasts and myotubes | 1000 nM, 1,25(OH)2D, 24 h | Induced hypertrophy of multinucleated myotubes by stimulating protein anabolism | ↓ expression of MRFs, Myf5 and myogenin in proliferating myoblasts. High concentration of VitD reduced myoblast-to-myoblast and myoblast-to-myotube fusion through the inhibition of Tmem8c (myomaker) and Gm7325 (myomerger). |
| Muscle function and protein synthesis | |||||
| Hayakawa et al., 2015 [28] | HSMM | Myotubes | 10 nM, 1,25(OH)2D, 24/48/or 72 h | ↑ interleukin-6 expression and inhibited expression of TNF-α, MAFbx, MuRF1 and ubiquitin ligases involved in muscle atrophy after VitD3 treatment. | VitD3 suppresses muscle degradation and its likely to be involved in the regulation of apoptosis, insulin responsiveness and myogenesis in skeletal muscle. |
| Van der Meijden et al., 2016 [29] | C2C12 | Myoblasts and myotubes | 0/400/1000/or 2000 nmol/L, 1,25(OH)2D, 24 h | Myoblasts and myotubes were able to convert 25(OH)D3 to 24,25(OH)2D3 locally (skeletal muscle); ↑ VDR and MHC mRNA expression. | Skeletal muscle is not only a direct target for VitD3 metabolites, but is also to its precursor. |
| Antinozzi et al., 2017 [30] | Human fetal skeletal muscle cells | Myoblasts | 10/or 100 nM, Elocalcitol, 1–24 h | Elocalcitol exerted an I-like effect, promoting GLUT4 re-localization in Flotillin-1, Caveolin-3 and Caveolin-1 positive sites and mTOR, AKT, ERK, 4E-BP1 activation; it enhanced Interleukin-6 myokine release. | VDR agonists as elocalcitol may be therapeutic tools for skeletal muscle integrity/function maintenance, an indispensable condition for health homeostasis. |
| Hirose et al., 2018 [31] | C2C12 | Myoblasts | 1000 and 10,000 nM, 1,25(OH)2D, 24 h | ↓ FOXO1-mediated; glucocorticoid-induced gene expression of atrogin 1 and cathepsin L | VitD3 may prevent muscle atrophy via the FOXO1-mediated pathway in muscle cells. |
| Arakawa & Wagatsuma, 2020 [32] | C2C12 | Myotubes | 100 nM, 1,25(OH)2D, 24 h | Enhanced agrin- induced AChR clustering in myotubes compared to treatment with agrin alone. | VitD3 -VDR signalling may regulate rapsin expression, resulting in the up-regulation of agrin- induced AChR clustering. |
| Mitochondria and lipid metabolism | |||||
| Ryan et al., 2013 [33] | C2C12 | Myotubes | 0.1–10000 nM, 1,25(OH)2D, ≤ 6 d | Low physiological concentrations (10K13 and 10K11 M) of VitD3 increased fat droplet accumulation; high physiological (10K9 M) and supraphysiological concentrations (R10K7 M) inhibited fat accumulation. | Low VitD concentration was associated with a sequential up-regulation of Pparg2 and Fabp4 mRNA, indicating formation of adipocytes, whereas higher concentrations reduced all these effects. |
| Jefferson et al., 2017 [15] | C2C12 | Myotubes | 100 nM, 1,25(OH)2D, 96 h | ↑ insulin-stimulated pAkt; ↑ total ceramides and DAG in a subspecies specific manner; ↓ the proportion of lipid within myotubes. | VitD3 altered myocellular lipid partitioning and lipid droplet packaging, lipid turnover and partially explained improvements in insulin sensitivity. |
| Chang & Kim, 2019 [11] | C2C12 | Myotubes | 100 nM, 1,25(OH)2D, 24 h | ↑ ATP levels and mitochondrial function gene expression: CPT1, PPARα, VLCAD, LCAD, MCAD, UCP2 and UCP3. ↑ SIRT1 mRNA expression, ↑ activation of AMPK and SIRT1 | Protective effect of VitD3 on muscle fat accumulation and mitochondrial dysfunction ↑ mtDNA, NRF1, PGC-1α and mitochondrial transcription factor A (Tfam) in C2C12 myotubes. |
| Schnell et al., 2019 [34] | C2C12 | Myotubes | 100 nM, 1,25(OH)2D, 24 h | ↑ mitochondrial function in myotubes (↑ lipolytic genes ATGL and CGI-58, OCR | ↑ mRNA expression of triglyceride synthesizing genes DGAT1 and DGAT2; in part mediated by Perilipin-2 |
| Glucose and insulin metabolism | |||||
| Manna et al., 2017 [35] | C2C12 | Myotubes | 25 or 50 nM, 1,25(OH)2D, 2 h | VitD3 + insulin ↑ GLUT4 translocation and glucose uptake | VitD3 ↑ glucose consumption by inducing SIRT1 activation, which in turn increases IRS1 phosphorylation and GLUT4 translocation in myotubes. |
| Tamilselvan et al., 2013 [36] | L6 | Myotubes | 100 nM, 1,25(OH)2D, 24 h | ↑ GLUT1, GLUT4, VDR and insulin receptor expression. | Potential antidiabetic role of VitD in regulation of expression of the glucose transporters in muscle cells. |
| Antinozzi et al., 2019 [18] | Human fetal skeletal muscle cells | Myoblasts | 10 and 100 nM, Elocalcitol, 15 min | Elocalcitol induced GLUT4 protein translocation likely in lipid raft microdomains; rapid IRS1 phosphorylation; inflammatory myopathy subjects, had VitD deficiency and a high lipidemic and resistin profile, possibly increasing the risk to develop metabolic diseases. | Elocalcitol might be a therapeutic tool for skeletal muscle integrity/function maintenance and important for health homeostasis. |
| Oxidative stress and AGES | |||||
| Tanaka et al., 2014 [37] | C2C12 | Myoblasts | 0.1 nM, 1,25(OH)2D, 48 h | ↑ expression of type 1 collagen; AGE2 and AGE3 suppressed the expression of MyoD, myogenin and OGN. 1,25D blunted the AGEs’ effects. | VitD3 may rescue the AGEs-induced sarcopenia as well as–suppressed osteoblastic differentiation via OGN expression in myoblasts. |
| Chang et al., 2019 [11] | C2C12 | Myotubes | 1, 10 and 100 nM, 1,25(OH)2D, 24 h | ↑ mtDNA, PGC1α, NRF1, Tfam, NRF2, NAD levels, activities of AMPK, SIRT1, expression of HMOX1 and TXNRD1. | ↓ muscle oxidative stress, lipid peroxidation, intracellular damage and cell death |
| Nonaka et al., 2020 [38] | C2C12 | Myotubes | 0/0.1/1/or 10 nM, 1,25(OH)2D, 4 d | VitD3 inhibited increases in Interleukin-6 protein, suggesting that VitD3 inhibits inflammation in muscle cells. | VitD3 can prevent or improve sarcopenia, which is associated with interleukin-6. |
| The authors, Year | Specie, N, Age | Groups (VitD Form and Dose), Time | Significant Findings and Effects | Comments |
|---|---|---|---|---|
| Myotube formation, muscle mass, strength and force | ||||
| Ray et al., 2016 [39] | A/J mice, n = 20 (5 per group), 4- or 12-weeks old female | G1 (Low, 100 IU); G2 (Reference, 1000 IU); G3 (10,000 IU 1,25(OH)2D/kg), 6 weeks | Low VitD3 group had ↓ maximal diaphragm (DIA) force, twitch force and fiber CSA (26%, 28% and 10% respectively). | Potential role of VitD3 in regulating DIA development and insulin sensitivity. |
| Trovato et al., 2018 [40] | Sprague/Dawley rats, n = 28 (4 per group), 7–9 weeks old male | RD (regular diet); R-DS (R + 4000 IU/KgVitD); RDR = regular diet without VitD; HFB-DS (high-fat diet +VitD); HFB-DR (high-fat w/o VitD); HFEVO-DS (high-fat + VitD); HFEVO-DR (high-fat + w/o VitD),10 weeks | Muscle fibres of high fat diet + VitD3 rats were hypertrophic comparing to those of regular diet + VitD3. | VitD associated with a Mediterranean diet showed trophic action on the muscle fibres. |
| Hayes et al., 2019 [41] | C57BL/6J mice n = 32, 8 weeks old | Control (standard chow + 1000 IU/kgVitD3); High (same diet with 20,000 IU/kg of 25-hydroxyvitamin D); YEAR (injected bolus of 1500 IU25-hydroxyvitamin D) 4 weeks | YEAR ↓ forces in both muscles compared to High, as well as lower force during fatigue and early recovery. | Mice ingesting the same amount of food + VitD3 over four weeks did not demonstrate the same detrimental effects. |
| Muscle function and protein synthesis | ||||
| Alkharfy et al., 2012 [42] | C57BL/6J mice, n = 44 (11 pergroup), 4–5 weeks old | Low fat diet (LFD); High fat diet (HFD) with and without 150 IU/kg/day 1,25(OH)2D, 16 weeks | HFD with VitD3 showed less weight gain as compared to controls (6.8% vs. 28.7%, respectively). | Muscle structural abnormalities caused by HFD were attenuated by VitD3; tissues have regained their normal structural appearance. |
| Gifondorwa et al., 2016 [43] | C57BL/6J mice, n = 24 (6 per group), 3 weeks old male | G1 (VitD+/Ca+: 1000 IU/kg/0.50%); G2 (VitD+/Ca-1000 IU/kg/0.01%); G3 (VitD-/Ca+: 0 IU/kg/0.47%); G4: (Vit. D2-/Ca-: 0.02%; 0 IU/kg), 9 weeks | VitD3 lead to metabolic changes, NMJ-related and protein chaperoning and refolding genes. | VitD deficient or a VitD and Ca+2 deficient diet resulted in detrimental changes in the structure and function of the NMJ. |
| Nakamura et al., 2020 [44] | C57BL/6J mice, n = 5 per group, 9 weeks old | Standard (S) diet, High 1,25(OH)2D diet = (VitD3 and Ca: 0.47%, P: 0.3%,) and Low 1,25(OH)2D diet = (Ca: 2%, P: 1.25%), 4–8 weeks | VitD low status worsens immobilization-induced muscle atrophy in mice. Mice globally lacking VDR exhibited more severe muscle atrophy following limb immobilization than controls. | Maintaining VitD status at an appropriate level before injury or decline in physical activity is likely crucial to prevent deterioration and muscle atrophy. |
| Mitochondria and lipid metabolism | ||||
| Fan et al., 2016 [14] | C57BL/6J mice, n = 15, 8 weeks old male | NFD (control normal-fat diet); HFD (high-fat diet); HFVD (45 kcal % fat; 50 μg/kg body weight/d 25-hydroxyvitamin D),9 weeks | HFVD ↓ body weight and adipose tissue weight and ↑ expression of UCP3 compared to the other groups. | Changes in the expression of genes correlated with VitD3/VDR. VitD3/VDR inhibits weight gain by activating UCP3 in the muscles. |
| Chanet et al., 2017 [45] | Wistar rats, n = 50, 15 months old male | Control (1 IU VitD3/g); VitD-depleted [VDD, diet 0 IU 1,25(OH)2D)], 6 months | Weight gain was associated with ↑ in fat mass (+63%, p < 0.05), intramyocellular lipids (+75%, p < 0.05) in VDD. | VitD3 deficiency in old rats ↑ adiposity and leads to reduced muscle protein synthesis through activation of eIF2α. These disorders are restored by VitD3. |
| Glucose and insulin metabolism | ||||
| Benetti et al., 2018 [13] | C57BL/6J mice, n = 40, 4 weeks old male | Control or High Fat-High Sugar (HFHS) diet for 4 months; Then, another subset of animals: 1,25(OH)2D (7 μg/kg−1, 3 times a week) for 2 months | VitD3 ↓ body weight and ↑ systemic glucose tolerance. VitD3 restored the impaired muscle insulin signalling and reverted myosteatosis diet-induced. | VitD3 ↓ activation NFKB and ↓ TNFα, ↓ activation of the SCAP/SREBP lipogenic pathway, ↓ CML protein adducts and RAGE expression. |
| Nadimi et al., 2019 [46] | Sprague-Dawley rats, n = 36, adult male | G1 (healthy control); G2 (healthy receiving sesame oil as placebo); G3 (diabetics receiving sesame oil as placebo); G4 (diabetics treated with 4300 IU/kg/week native cholecalciferol), 4 weeks | VitD ↑ FNDC5 gene expression and muscle irisin levels. | Potential therapeutic effect of VitD3 supplementation for diabetes mellitus. |
| Xavier et al., 2012 [47] | Wistar rats, n = 6–8 per group, 6 months old male | 12 μg/kg VitD3 to (a) control; (b) diabetic; (c) insulin-treated diabetic; (d) 1,25(OH)2D -treated diabetic; (e) curcumin-treated diabetic rats, 2 weeks | ↑ β2-adrenoceptor and CREB gene expression were observed in the diabetic group and ↓ insulin receptor expression, resulting in ↑ glycogenolysis, gluconeogenesis and ↓ glycogenesis in the muscles. | These results were reversed with VitD3 and curcumin treatment. VitD3 and curcumin might help in the management of peripheral complications associated with diabetes. |
| Kim et al., 2020 [48] | p62-deficient mice, n = 10 per group, 24 weeks old male | Control (no treatment); cholecalciferol = 1000 IU VitD3/kg/d), RT = ladder climbing, 3 times per week or combined treatment, VRT = VitD3 + RT), 10 weeks VitD3 | Total body mass increased in all groups, but fat mass increased only in control group. Loss of skeletal muscle function was reported only in control group. Improved blood glucose levels and ↓ spleen mass was reported in RT and VRT compared to control. | VitD3 attenuated the progression of obesity and preserved skeletal muscle function. |
| Akagawa et al., 2018 [49] | Otsuka Long-Evans Tokushima sedentary fatty rats (8–10/group) 20 weeks old | ALF (alfacalcidol 0.1 μg/kg/day); Exe (low-intensity aerobic exercise training); Comb (alfacalcidol + low-intensity aerobic exercise training); T2DM control group, 2 or 6 weeks | ALF, Exe and Comb treatments for 2 and 6 weeks recovered the CSA compared to Control. ALF and Comb for 6 weeks increased femoral BMD compared to Control. ALF or Exe monotherapy significantly decreased Atrogin-1 or MuRF1 expression after 2 weeks. After 6 weeks, ALF and Comb decreased Atrogin-1 and REDD1. | A combination of ALF and Exe improved CSA from the early phase of treatment by stimulating skeletal muscle differentiation and suppressing muscle catabolic genes. Improvements in blood glucose, BMD and CSA were observed as long-term effects of the combination therapy. |
| The authors, Year | Male/ Female (n) | Baseline Serum [25(OH)D] | N | Dose (IU) | Type | Frequency | Duration (weeks) | INTERVENTION Outcomes | N | PLACEBO Outcomes | Comments |
|---|---|---|---|---|---|---|---|---|---|---|---|
| El Hajj et al., 2018 [63] | 59/56 | Deficient | 60 | 10000 | Cholecalciferol | 3x/week | 24 | Handgrip (kg) ↑ Muscle mass (kg) | 55 | Handgrip (kg) Muscle mass (kg) | Serum [25(OH)D] had significantly change at the end of the study between groups. |
| Bislev LS et al., 2018 [9] | 0/81 | Deficient | 40 | 2800 | Cholecalciferol | daily | 12 | ↓ Handgrip (N) ↓ Knee flexion 60° strength (N) Lean mass (kg) ↑ Timed up and go test (sec) Physical performance (MET score) | 41 | ↑ Handgrip (N) ↑ Knee flexion 60° strength (N) Lean mass (kg) ↓ Timed up and go test (sec) Physical performance (MET score) | Serum [25(OH)D] had significantly change at the end of the study between groups. |
| Shea MK et al., 2019 [1] | 64/36 | Deficient | 47 | 800–1600 | Cholecalciferol | daily | 48 | Double leg press power W 40% 1RM Double leg press power W 70% 1RM Double leg press strength 1RM Grip strength (kg) Total lean body mass (kg) | 50 | Double leg press power W 40% 1RM Double leg press power W 70% 1RM Double leg press strength 1RM Grip strength (kg) Total lean body mass (kg) | At the 4-mo visit, if a participant from VitD group had serum [25(OH)D] <28 ng/mL, it was given an additional VitD3 capsule (800 IU)/day. To everyone else was given an additional placebo pill. There was only significant change on serum [25(OH)D] at the end of the study between groups. |
| Vaes AMM et al., 2018 [64] Vaes AMM et al., 2018 [64] | 43/32 43/32 | Deficient Deficient | 24 26 | 800 400 | Cholecalciferol 25-hydroxycholecalciferol | daily daily | 24 24 | Handgrip (kg) Timed up and go test (sec) SPPB total (points 0–12) Knee extension (Nm) Knee flexion (Nm) Total lean mass (kg) Handgrip (kg) Timed up and go test (sec) SPPB total (points 0–12) Knee extension (Nm) Knee flexion (Nm) Total lean mass (kg) | 25 | Handgrip Timed up and go test (sec) SPPB total (points 0–12) Knee extension (Nm) Knee flexion (Nm) Total lean mass (kg) | 3 intervention groups: 25(OH)D3, VitD3 and placebo. In both treatment with VitD, serum [25(OH)D] increased. |
| Grimnes G et al., 2019 [21] | 219/192 | Deficient | 208 | 20000 | Cholecalciferol | 1/week | 16 | Handgrip (kg) Hip flexion (N) Biceps flexion (N) Pectoralis (N) | 203 | Handgrip Hip flexion (N) Biceps flexion (N) Pectoralis | There was only a change in serum [25(OH)D] at the end of the study between groups. |
| Van Vliet S et al., 2020 [65] | 6/11 | Sufficient or Insufficient | 9 | 2000 | Cholecalciferol | daily | 8 | Handgrip (kg) Myofibrillar protein synthesis rate | 8 | Handgrip (kg) Myofibrillar protein synthesis rate | There was only increase in serum [25(OH)D] at the end on the treatment group. |
| Cuellar WA et al., 2019 [3] | 113/104 | Deficient to Sufficient | 104 | 50000 | Cholecalciferol | 1 capsule/ month | 96 | Trunk muscle size Chance in thickness of muscle with contraction ↑ Relaxed multifidus muscle thickness (cm) at L2/L3 when adjusted for age + sex + BMI + leg strength | 113 | Trunk muscle size Chance in thickness of muscle with contraction ↓ Relaxed multifidus muscle thickness (cm) at L2/L3 | Serum [25(OH) D] in the VitD group increased more at the end of the study. |
| Ceglia L et al., 2013 [66] | 0/21 | Insufficient | 9 | 4000 | Cholecalciferol | Daily | 16 | 40% and 70% of 1RM average power in knee extension (W) ↑ %∆ in total FCSA (type I + type II) Type I muscle FCSA (µm2) Type II muscle FCSA (µm2) ↑ %∆ in [VDR] ↑ %VDR-positive myonuclei in type II fibres | 12 | 40% and 70% of 1RM average power in knee extension (W) ↓ %∆ in total FCSA (type I + type II) Type I muscle FCSA (µm2) Type II muscle FCSA (µm2) %∆ in [VDR] %VDR-positive myonuclei in type II fibres | There was a significant increase in serum [25(OH)D] in the vitamin D compared with placebo group at the end of the study. |
| Latham NK et al., 2003 [67] | 114/129 | Deficient | 108 | 300000 | calciferol | single dose | 24 | Physical component score (mean) Quadriceps strength (kg) Timed up and go (sec) | 114 | Physical component score (mean) Quadriceps strength (kg) Timed up and go (sec) | The single dose of VitD was effective only in increasing mean [25(OH)D] in the intervention group at the end of the study. |
| Dhesi JK et al., 2004 [68] | 30/108 | Deficient | 62 | 600000 | ergocalciferol | single intramuscular injection | 24 | MVC (quadriceps strength, N) ↓ Aggregate functional performance time (sec) ↑ Choice reaction time (sec) ↓ Postural sway | 61 | MVC (quadriceps strength, N) ↑ Aggregate functional performance time (sec) ↓ Choice reaction time (sec) ↑ Postural sway | Serum [25(OH)D] increased significantly in the end of the study in the treatment group. |
| Pirotta S et al., 2015 [20] | 13/13 | Insufficient | 13 | 2000 | Cholecalciferol | daily | 10 | KE 120°/s (N/kg) KE 180°/s (N/kg) KE 240°/s (N/kg) Stair clim power (W) Timed up and go test (m/s) | 12 | KE 120°/s (N/kg) KE 180°/s (N/kg) KE 240°/s (N/kg) Stair clim power (W) Timed up and go test (m/s) | At the end of the study, only serum [25(OH)D] increased at the end of the study in the vitamin D group. |
| Wood AD et al., 2014 [67] | 0/265 | Insufficient | 84 90 | 400 1000 | Cholecalciferol | daily | 48 | Grip strength (kg) Grip strength (kg) | 91 | Grip strength (kg) | 3 intervention groups: low VitD3 dose, high VitD3 dose and placebo. Serum [25(OH)D] > 60 nmol/L in both VitD3 groups. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Romeu Montenegro, K.; Amarante Pufal, M.; Newsholme, P. Vitamin D Supplementation and Impact on Skeletal Muscle Function in Cell and Animal Models and an Aging Population: What Do We Know So Far? Nutrients 2021, 13, 1110. https://doi.org/10.3390/nu13041110
Romeu Montenegro K, Amarante Pufal M, Newsholme P. Vitamin D Supplementation and Impact on Skeletal Muscle Function in Cell and Animal Models and an Aging Population: What Do We Know So Far? Nutrients. 2021; 13(4):1110. https://doi.org/10.3390/nu13041110
Chicago/Turabian StyleRomeu Montenegro, Karina, Milene Amarante Pufal, and Philip Newsholme. 2021. "Vitamin D Supplementation and Impact on Skeletal Muscle Function in Cell and Animal Models and an Aging Population: What Do We Know So Far?" Nutrients 13, no. 4: 1110. https://doi.org/10.3390/nu13041110
APA StyleRomeu Montenegro, K., Amarante Pufal, M., & Newsholme, P. (2021). Vitamin D Supplementation and Impact on Skeletal Muscle Function in Cell and Animal Models and an Aging Population: What Do We Know So Far? Nutrients, 13(4), 1110. https://doi.org/10.3390/nu13041110