
Effect of Trilobatin from Lithocarpus polystachyus Rehd on Gut Microbiota of Obese Rats Induced by a High-Fat Diet

Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Animals and Diet
2.3. Short-Chain Fatty Acid Analysis
2.4. Gut Microbiota Analysis
2.5. Predictive Metagenome Analysis
2.6. Statistical Analysis
3. Results
3.1. Food Intake, Morphometric Parameters, and SCFAs Are Greatly Modulated by Trilobatin from LPR Leaves
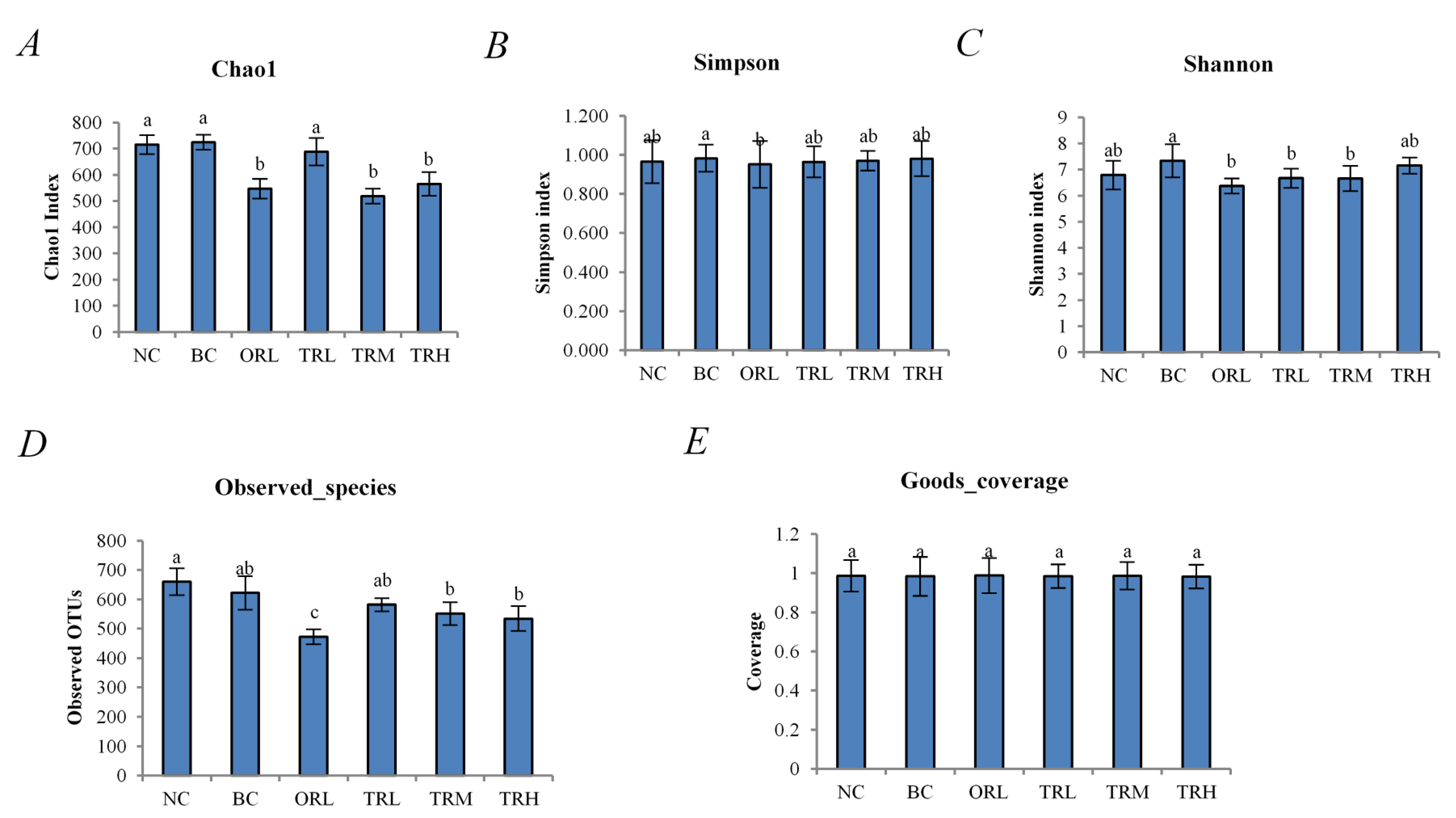
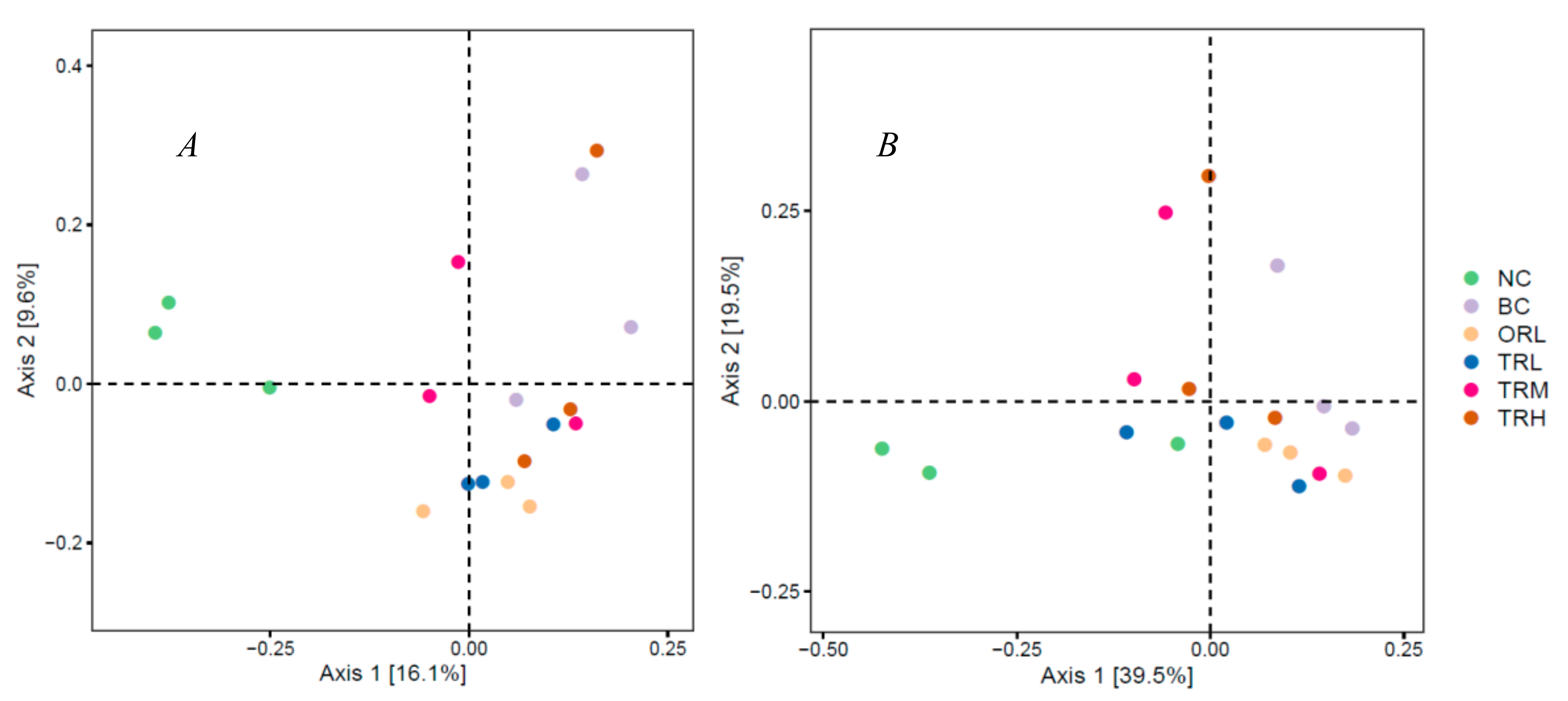
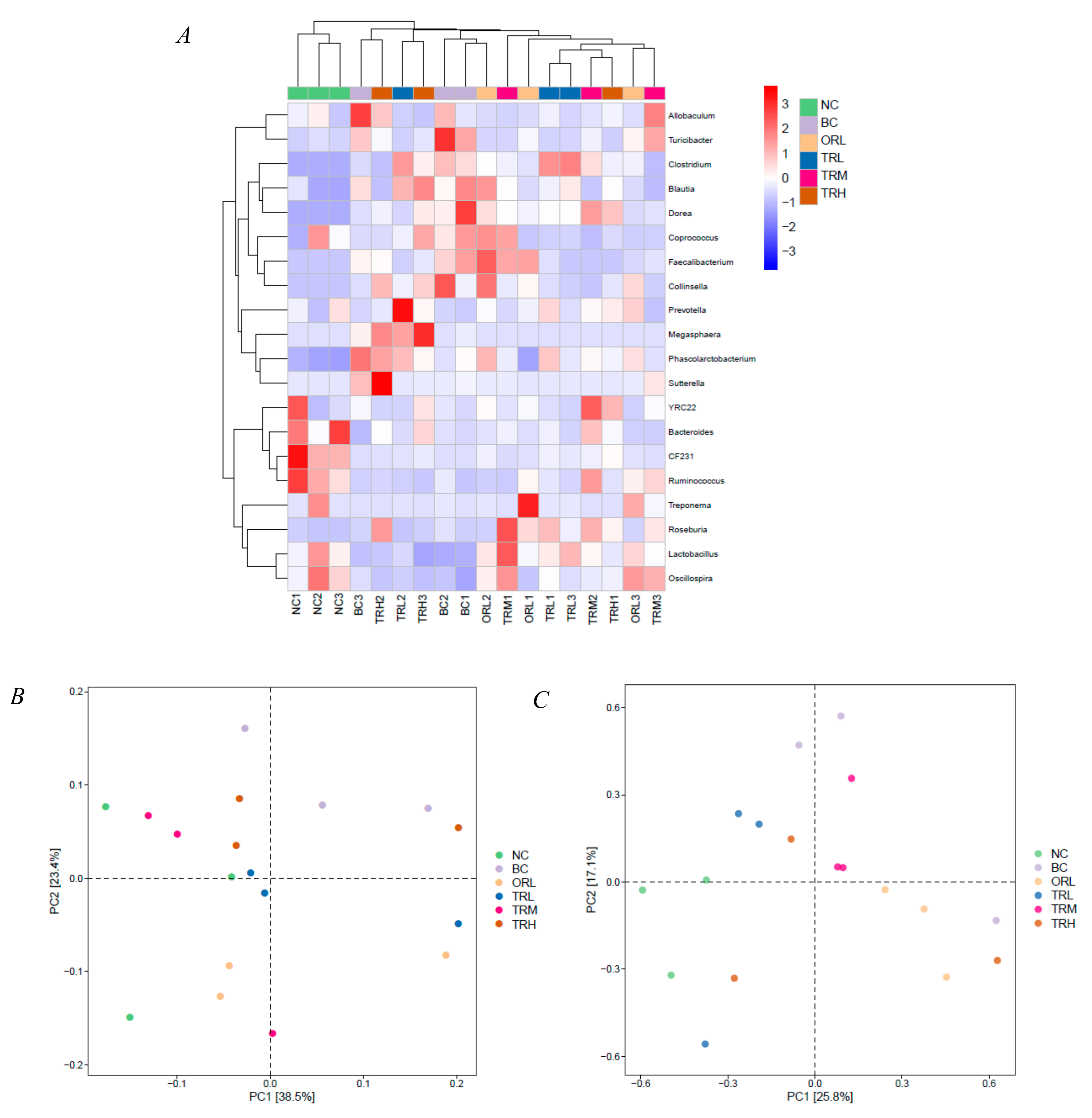
3.2. Effects of Trilobatin on the Diversity and Structure of Gut Microbiota
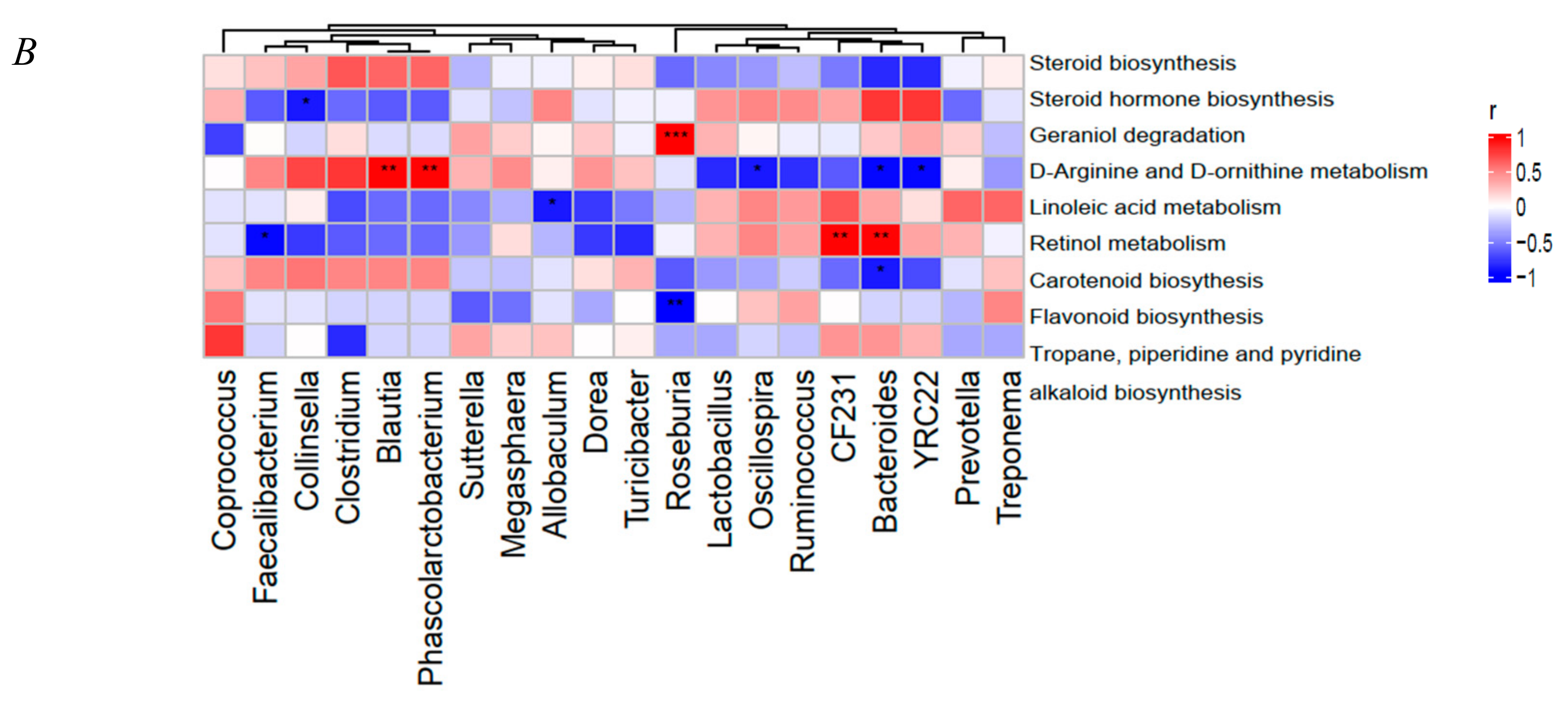
3.3. Functional Prediction Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bahmani, M.; Eftekhari, Z.; Saki, K.; Fazelimoghadam, E.; Jelodari, M. Obesity phytotherapy: Review of native herbs used in traditional medicine for obesity. Evid. Based Complement. Altern. Med. 2016, 21, 228–234. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.; Huang, S.; Yen, G. Inhibitory effect of phenolic acids on the proliferation of 3T3-L1 preadipocytes in relation to their antioxidant activity. J. Agric. Food Chem. 2006, 54, 4191–4197. [Google Scholar] [CrossRef] [PubMed]
- Choe, S.S.; Huh, J.Y.; Hwang, I.J.; Kim, J.I.; Kim, J.B. Adipose tissue remodeling: Its role in energy metabolism and metabolic disorders. Front. Endocrinol. 2016, 7, 30. [Google Scholar] [CrossRef]
- Luo, L.; Liu, M. Adipose tissue in control of metabolism. J. Endocrinol. 2016, 231, R77–R99. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.W.; Jones, P.J. Conjugated linoleic acid and obesity control: Efficacy and mechanisms. Int. J. Obes. 2004, 28, 941–955. [Google Scholar] [CrossRef] [PubMed]
- Li, D.H.; Wu, H.; Dou, H. Weight loss effect of sweet orange essential oil microcapsules on obese SD rats induced by high-fat diet. Biosci. Biotechnol. Biochem. 2019, 29, 1699–1704. [Google Scholar] [CrossRef]
- Zhang, Z.C.; Li, S.; Cao, H.Y.; Shen, P.; Liu, J.X.; Fu, Y.H.; Cao, Y.G.; Zhang, N.S. The protective role of phloretin against dextran sulfate sodium-induced ulcerative colitis in mice. Food Funct. 2019, 10, 422–431. [Google Scholar] [CrossRef] [PubMed]
- Nie, R.; Tanaka, T.; Zhou, J.; Tanaka, O. Phlorizin and trilobatin, sweet dihydrochalcone-glucosides from leaves of Lithocarpus litseifolius (Hance) Rehd. (Fagaceae). Agric. Biol. Chem. 1982, 46, 1933–1934. [Google Scholar] [CrossRef]
- Qin, X.; Liu, J. A new sweet dihydrochalcone-glucoside from leaves of Lithocarpus pachyphyllus (Kutz) rehd. (Fagaceae). Z. Naturforschung C 2003, 58, 759–762. [Google Scholar] [CrossRef]
- Shang, A.; Liu, H.Y.; Luo, M.; Xia, Y.; Yang, X.; Li, H.Y.; Wu, D.T.; Sun, Q.; Geng, F.; Li, H.B.; et al. Sweet tea (Lithocarpus polystachyus rehd.) as a new natural source of bioactive dihydrochalcones with multiple health benefits. Crit. Rev. Food Sci. Nutr. 2020. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Huang, Y.Y.; Yang, Z.; Zhou, C.; Hu, X.J. Identification and quantitative evaluation of major sweet ingredients in sweet tea (Lithocarpus polystachyus Rehd.) based upon location, harvesting time, leaf age. J. Chem. Soc. Pak. 2018, 40, 158–164. [Google Scholar]
- Zhao, Y.; Li, X.; Zeng, S.; Huang, S.; Hou, S.Z.; Lai, X.P. Characterization of phenolic constituents in Lithocarpus polystachyus. Anal. Method 2014, 6, 1359–1363. [Google Scholar] [CrossRef]
- Shin, S.K.; Cho, S.J.; Jung, U.J.; Ryu, R.; Choi, M.S. Phlorizin supplementation attenuates obesity, inflammation, and hyperglycemia in diet-induced obese mice fed a high-fat diet. Nutrients 2016, 8, 92. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.J.; Huang, S.; Liu, J.Q.; Qiu, S.Q.; Xie, F.Y.; Song, H.P.; Li, Y.S.; Hou, S.Z.; Lai, X.P. Sweet tea leaves extract improve leptin resistance in diet-induced obese rats. J. Ethnopharmacol. 2013, 145, 386–392. [Google Scholar] [CrossRef] [PubMed]
- Casanova-Marti, A.; Serrano, J.; Portune, K.J.; Sanz, Y.; Blay, M.T.; Terra, X.; Ardevol, A.; Pinent, M. Grape seed proanthocyanidins influence gut microbiota and enteroendocrine secretions in female rats. Food Funct. 2018, 9, 1672. [Google Scholar] [CrossRef]
- Shen, R.; Dang, X.Y.; Dong, J.L.; Hu, X.Z. Effects of Oat β-glucan and barley β-glucan on fecal characteristics, intestinal microflora, and intestinal bacterial metabolites in rats. J. Agric. Food Chem. 2012, 60, 11301–11308. [Google Scholar] [CrossRef] [PubMed]
- Anhê, F.F.; Roy, D.; Pilon, G.; Dudonné, S.; Matamoros, S.; Varin, T.V.; Garofalo, C.; Moine, Q.; Desjardins, Y.; Levy, E.; et al. A polyphenol-rich cranberry extract protects from diet-induced obesity, insulin resistance and intestinal inflammation in association with increased Akkermansia spp. Population in the gut microbiota of mice. Gut 2015, 64, 872–883. [Google Scholar] [CrossRef] [PubMed]
- Jing, N.; Liu, X.; Jin, M.; Yang, X.; Hu, X.; Li, C.; Zhao, K. Fubrick tea attenuates high-fat diet induced fat deposition and metabolic disorder by regulating gut microbiota and caffeine metabolism. Food Funct. 2020, 11, 6971. [Google Scholar] [CrossRef]
- Wu, W.; Hu, J.; Gao, H.; Chen, H.; Fang, X.; Mu, H.; Han, Y.; Liu, R. The potential cholesterol-lowering and prebiotic effects of bamboo shoot dietary fibers and their structure characteristics. Food Chem. 2020, 332, 127372. [Google Scholar] [CrossRef] [PubMed]
- Chung, A.; Coimbra, C.; Farias, P.; Francisco, R.; Branco, R.; Simao, F.V.; Gomes, E.; Pereira, A.; Vila, M.C.; Fiuza, A.; et al. Tailings microbial community profile and prediction of its functionality in basins of tungsten mine. Sci. Rep. 2019, 9, 19596. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.H.; Youn, J.H.; Park, J.Y.; Hong, S.K.; Park, K.S.; Park, S.W.; Suh, K.I.; Lee, K.U. Effects of high-fat diet and exercise training on intracellular glucose metabolism in rats. American Journal of Physiology. Endocrinol. Metab. 2000, 278, E977–E984. [Google Scholar] [CrossRef]
- China Pharmacopoeia Committee. Pharmacopoeia of the People’s Republic of China; China Chemical Industry Press: Beijing, China, 1999; pp. 327–329. [Google Scholar]
- Hou, S.Z.; Chen, S.X.; Huang, S.; Jiang, D.X.; Zhou, C.J.; Chen, C.Q.; Liang, Y.M.; Lai, X.P. The hypoglycemic activity of Lithocarpus polystachyus Rehd. Leaves in the experimental hyperglycemic rats. J. Ethnopharmacol. 2011, 138, 142–149. [Google Scholar] [CrossRef]
- Hou, S.Z.; Xu, S.J.; Jiang, D.X.; Chen, S.X.; Wang, L.L.; Huang, S.; Lai, X. Effect of the flavonoid fraction of Lithocarpus polystachyus Rehd. on spontaneously hypertensive and normotensive rats. J. Ethnopharmacol. 2012, 143, 441–447. [Google Scholar] [CrossRef]
- Zhao, H.; Yakar, S.; Gavrilova, O.; Sun, H.; Zhang, Y.; Kim, H.; Setser, J.; Jou, W.; Leroith, D. Phloridzin improves hyperglycemia but no hepatic insulin resistance in a transgenic mouse model of type 2 diabetes. Diabetes 2004, 53, 2901–2909. [Google Scholar] [CrossRef]
- Serrano, J.; Casanova-Marti, A.; Gual, A.; Perez-Vendrell, A.M.; Blay, M.T.; Terra, X.; Ardevol, A.; Pinent, M. A specific dose of grape seed-derived proanthocyanidins to inhibit body weight gain limits food intake and increases energy expenditure in rats. Eur. J. Nutr. 2016, 56, 1629–1636. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Y.D.; Marungruang, N.; Fak, F.; Nyman, M. Effects of two whole-grain barley varieties on caecal SCFA, gut microbiota and plasma inflammatory markers in rats consuming low- and high-fat diets. Br. J. Nutr. 2015, 113, 1558–1570. [Google Scholar] [CrossRef] [PubMed]
- Hosseini, E.; Grootaert, C.; Verstraete, W.; van de Wiele, T. Propionate as a health-promoting microbial metabolite in the human gut. Nutr. Rev. 2011, 69, 245–258. [Google Scholar] [CrossRef] [PubMed]
- Schwiertz, A.; Taras, D.; Schafer, K.; Beijer, S.; Bos, N.A.; Donus, C.; Hardt, P.D. Microbiota and SCFA in lean and overweight healthy subjects. Obesity 2010, 18, 190–195. [Google Scholar] [CrossRef] [PubMed]
- Xia, X.; Li, G.; Ding, Y.; Ren, T.; Zheng, J.; Kan, J. Effect of whole grain Qingke (Tibetan Hordeum Vulgare L. Zangqing 320) on the serum lipid levels and intestinal microbiota of rats under high-fat diet. J. Agric. Food Chem. 2017, 65, 2686–2693. [Google Scholar] [CrossRef]
- Hu, H.W.; Zhang, L.M.; Dai, Y.; Di, H.J.; He, J.Z. pH-dependent distribution of soil ammonia oxidizers across a large geographical scale as revealed by high-throughput pyrosequencing. J. Soils Sediments 2013, 13, 1439–1449. [Google Scholar] [CrossRef]
- Zhang, J.; Guo, Z.; Xue, Z.; Sun, Z.; Zhang, M.; Wang, L.; Wang, G.; Wang, F.; Xu, J.; Cao, H.; et al. A phylo-functional core of gut microbiota in healthy young Chinese cohorts across lifestyles, geography and ethnicities. ISME J. 2015, 9, 1979–1990. [Google Scholar] [CrossRef] [PubMed]
- Gil-Gardoso, K.; Gines, I.; Pinent, M.; Ardevol, A.; Blay, M.; Terra, X. Effects of flavonoids on intestinal inflammation, barrier integrity and changes in gut microbiota during diet-induced obesity. Nutr. Res. Rev. 2016, 29, 234–248. [Google Scholar] [CrossRef] [PubMed]
- Ducan, S.H.; Lobley, G.E.; Holtrop, G.; Ince, J.; Johnstone, A.M.; Louis, P.; Flint, H.J. Human colonic microbiota associate with diet, obesity and weight loss. Int. J. Obes. 2008, 32, 1720–1724. [Google Scholar] [CrossRef] [PubMed]
- Murphy, E.F.; Cotter, P.D.; Healy, S.; Marques, T.M.; Sullivan, O.; Fouhy, F.; Clarke, S.F.; Toole, P.W.; Quigley, E.M.; Stanton, C.; et al. Composition and energy harvesting capacity of the gut microbiota: Relationship to diet, obesity and time in mouse models. Gut 2010, 59, 1635–1642. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.C.; Lin, S.H.; Chang, J.S.; Chien, Y.W. High-fat diet with a high monounsaturated fatty acid and polyunsaturated/saturated fatty acid ratio suppresses body fat accumulation and weight gain in obese hamsters. Nutrients 2017, 9, 1148. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Yang, Z.; Chen, X.; Huang, Y.; Yin, B.; Guo, F.; Zhao, H.; Huang, J.; Wu, Y.; Gu, R. Effects of Lactobacillus rhamnosus hsryfm 1301 on the gut microbiota and lipid metabolism in rats fed a high-fat diet. J. Microbiol. Biotechnol. 2015, 25, 687–695. [Google Scholar] [CrossRef]
- Konikoff, T.; Gophna, U. Oscillospira: A central, enigmatic component of the human gut microbiota. Trends Microbiol. 2016, 24, 523–524. [Google Scholar] [CrossRef] [PubMed]
- Hatti, K.S.; Diwakar, L.; Rao, G.V.; Kush, A.; Reddy, G.C. Abyssinones and related flavonoids as potential steroidogenesis modulators. Bioinformation 2009, 3, 399–402. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Panche, A.N.; Diwan, A.D.; Chandra, S.R. Flavonoids: An overview. J. Nutr. Sci. 2016, 5, 1–15. [Google Scholar] [CrossRef]
- Pan, J.J.; Solbiati, J.O.; Ramamoorthy, G.; Hillerich, B.S.; Seidel, R.D.; Gronan, J.E.; Almo, S.C.; Poulter, C.D. Biosynthesis of squalene from farnesyl diphosphate in bacteria: Three steps catalyzed by three enzymes. ACS Cent. Sci. 2015, 1, 77–82. [Google Scholar] [CrossRef] [PubMed]
- Fan, X.; Zhang, Y.; Dong, H.; Wang, B.; Ji, H.; Liu, X. Trilobatin attenuates the LPS-mediated inflammatory response by suppressing the NF-kappaB signaling pathway. Food Chem. 2015, 166, 609–615. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.S.; Zhou, Y. Excess vitamin intake: An unrecognized risk factor for obesity. World J. Diabetes 2014, 5, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Saleem, F.; Rizvi, S.W. New therapeutic approaches in obesity and metabolic syndrome with polycystic ovary syndrome. Cureus 2017, 9, e1844. [Google Scholar] [CrossRef] [PubMed]
- Dong, H.; Li, M.; Zhu, F.; Liu, F.; Huang, J. Inhibitory potential of trilobatin from Lithocarpus polystachyus Rehd against a-glucosidase and a-amylase linked to type 2 diabetes. Food Chem. 2012, 130, 261–266. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NC | BC | ORL | TRL | TRM | TRH | ||
|---|---|---|---|---|---|---|---|
| Food intake | Sum total intake (g) | 124.72 ± 4.57 a | 135.08 ± 2.09 a | 130.99 ± 6.65 a | 126.71 ± 5.86 a | 125.20 ± 10.2 a | 126.11 ± 3.02 a |
| Morphometric parameters | Final body weight (g) | 453 ± 11.4 d | 492.42 ± 10.29 a | 467.08 ± 12.9 cd | 472.50 ± 11.26 b | 457.75 ± 10.4 cd | 475.58 ± 9.57 b |
| Gain body weight (g) | 59.17 ± 3.50 cd | 85.92 ± 4.07 a | 66.58 ± 8.00 bc | 71.50 ± 6.78 b | 56.17 ± 5.09 d | 74.75 ± 5.35 b | |
| Liver weight (g) | 12.35 ± 1.23 c | 17.67 ± 1.11 a | 16.18 ± 1.21 b | 16.63 ± 1.28 bc | 16.43 ± 1.2 bc | 16.40 ± 1.41 bc | |
| PAT (g) | 3.59 ± 0.46 c | 8.01 ± 1.01 a | 5.19 ± 0.42 bc | 6.19 ± 0.41 b | 5.20 ± 0.47 bc | 5.07 ± 0.49 bc | |
| EAT (g) | 4.06 ± 0.16 b | 5.23 ± 0.36 a | 5.07 ± 0.41 ab | 5.03 ± 0.19 ab | 4.30 ± 0.32 b | 4.52 ± 0.27 b | |
| BAT (g) | 0.47 ± 0.06 c | 0.68 ± 0.11 a | 0.57 ± 0.09 b | 0.61 ± 0.11 ab | 0.59 ± 0.09 ab | 0.55 ± 0.07 ab | |
| Sum AT (g) | 8.12 ± 0.13 c | 13.92 ± 0.57 a | 10.83 ± 0.54 b | 11.83 ± 0.19 bc | 10.09 ± 0.25 b | 10.14 ± 0.23 b | |
| Short-chain fatty acids (ng/μL) | Acetic acid | 5.11 ± 0.25 a | 2.49 ± 0.11 b | 0.67 ± 0.03 c | 4.65 ± 0.20 ab | 2.37 ± 0.12 bc | 1.66 ± 0.09 bc |
| Propionic acid | 4.89 ± 0.31 b | 4.54 ± 0.21 b | 1.55 ± 0.12 c | 3.85 ± 0.18 bc | 6.17 ± 0.25 a | 2.47 ± 0.11 bc | |
| Isobutyric acid | 5.23 ± 0.54 a | 3.18 ± 0.39 a | 4.19 ± 0.46 a | 3.54 ± 0.61 a | 3.30 ± 0.87 a | 4.32 ± 0.35 a | |
| Butyric acid | 3.37 ± 0.46 b | 2.52 ± 0.13 c | 5.44 ± 0.45 a | 2.68 ± 0.18 c | 2.51 ± 0.16 c | 5.29 ± 1.05 a | |
| Isovaleric acid | 5.95 ± 0.66 a | 6.04 ± 1.01 a | 2.23 ± 0.11 a | 5.23 ± 0.56 a | 3.62 ± 0.23 a | 2.61 ± 0.31 a | |
| Valeric acid | 4.79 ± 0.35 a | 4.97 ± 0.23 a | 2.95 ± 0.12 a | 4.50 ± 0.55 a | 3.19 ± 0.42 a | 2.99 ± 0.36 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shen, H.; Huang, L.; Dou, H.; Yang, Y.; Wu, H. Effect of Trilobatin from Lithocarpus polystachyus Rehd on Gut Microbiota of Obese Rats Induced by a High-Fat Diet. Nutrients 2021, 13, 891. https://doi.org/10.3390/nu13030891
Shen H, Huang L, Dou H, Yang Y, Wu H. Effect of Trilobatin from Lithocarpus polystachyus Rehd on Gut Microbiota of Obese Rats Induced by a High-Fat Diet. Nutrients. 2021; 13(3):891. https://doi.org/10.3390/nu13030891
Chicago/Turabian StyleShen, Hailiang, Linhua Huang, Huating Dou, Yali Yang, and Houjiu Wu. 2021. "Effect of Trilobatin from Lithocarpus polystachyus Rehd on Gut Microbiota of Obese Rats Induced by a High-Fat Diet" Nutrients 13, no. 3: 891. https://doi.org/10.3390/nu13030891
APA StyleShen, H., Huang, L., Dou, H., Yang, Y., & Wu, H. (2021). Effect of Trilobatin from Lithocarpus polystachyus Rehd on Gut Microbiota of Obese Rats Induced by a High-Fat Diet. Nutrients, 13(3), 891. https://doi.org/10.3390/nu13030891