
Beneficial Actions of Orostachys japonica and Its Compounds against Tumors via MAPK Signaling Pathways


Abstract
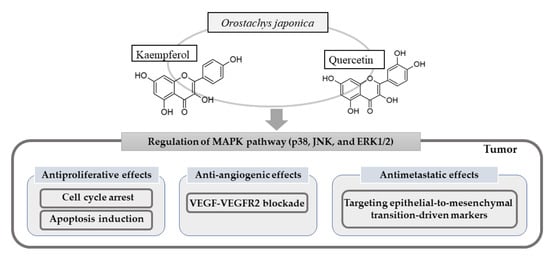
1. Introduction
2. Phytochemistry of O. japonica
3. Anticancer Properties of O. japonica
3.1. Antiproliferative Effect
3.1.1. Apoptosis Induction
3.1.2. Cell Cycle Arrest
3.2. Anti-Angiogenic Effect
3.3. Antimetastatic Effect
4. Possibility of the Combined Use of O. japonica with Other Chemotherapeutic Drugs
5. Anticancer Effects of Two Representative Flavonoids from O. japonica
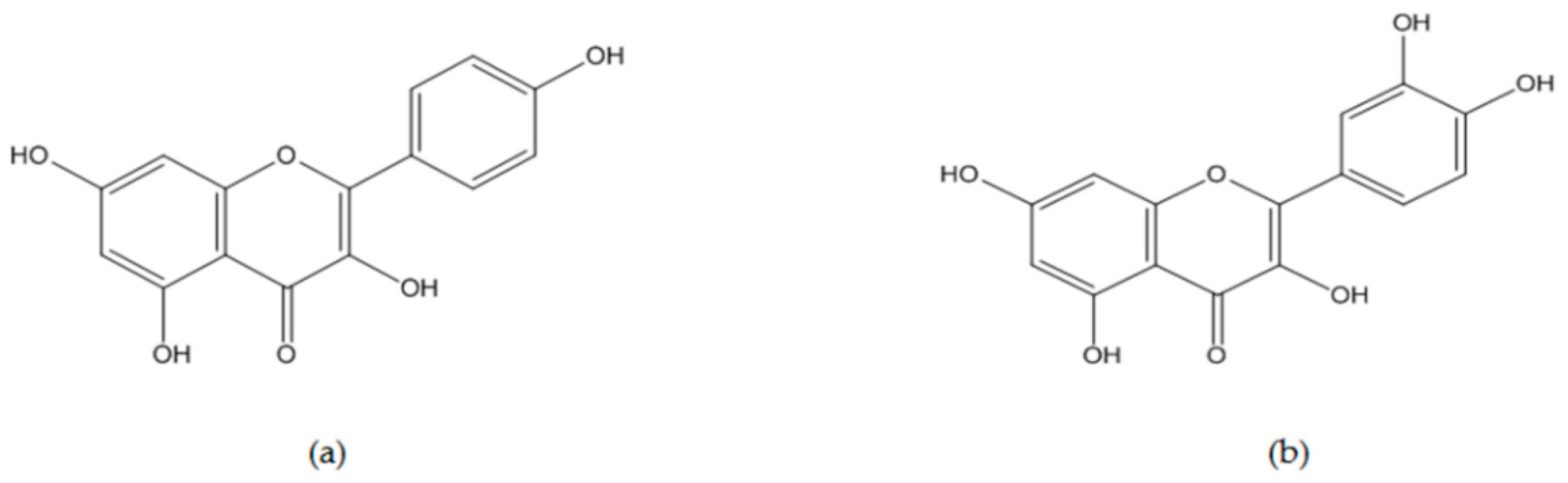
5.1. Kaempferol
5.2. Quercetin
6. Safety
7. Discussion
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- WHO. Global Health Estimates 2016: Disease Burden by Cause, Age, Sex, by Country and by Region, 2000–2016. Available online: https://www.who.int/healthinfo/global_burden_disease/estimates/en/ (accessed on 4 January 2021).
- Mattiuzzi, C.; Lippi, G. Current cancer epidemiology. J. Epidemiol. Glob. Health 2019, 9, 217–222. [Google Scholar] [CrossRef]
- Sun, J.; Wei, Q.; Zhou, Y.; Wang, J.; Liu, Q.; Xu, H. A systematic analysis of FDA-approved anticancer drugs. BMC Syst. Biol. 2017, 11, 27–43. [Google Scholar] [CrossRef]
- Akhdar, H.; Legendre, C.; Aninat, C.; Morel, F. Anticancer drug metabolism: Chemotherapy resistance and new therapeutic approaches. In Topics on Drug Metabolism; InTech: Rijeka, Croatia, 2012; pp. 138–170. [Google Scholar]
- De la Torre, B.G.; Albericio, F. The pharmaceutical industry in 2017. An analysis of FDA drug approvals from the perspective of molecules. Molecules 2018, 23, 533. [Google Scholar] [CrossRef] [PubMed]
- Zugazagoitia, J.; Guedes, C.; Ponce, S.; Ferrer, I.; Molina-Pinelo, S.; Paz-Ares, L. Current challenges in cancer treatment. Clin. Ther. 2016, 38, 1551–1566. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Zhou, L.; Xie, N.; Nice, E.C.; Zhang, T.; Cui, Y.; Huang, C. Overcoming cancer therapeutic bottleneck by drug repurposing. Signal Transduct. Target 2020, 5, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Ruan, W.J.; Lai, M.D.; Zhou, J.G. Anticancer effects of Chinese herbal medicine, science or myth? J. Zhejiang Univ. Sci. B 2006, 7, 1006–1014. [Google Scholar] [CrossRef]
- Sajadimajd, S.; Bahramsoltani, R.; Iranpanah, A.; Patra, J.K.; Das, G.; Gouda, S.; Rahimi, R.; Rezaeiamiri, E.; Cao, H.; Giampieri, F. Advances on natural polyphenols as anticancer agents for skin cancer. Pharm. Res. 2020, 151, 1–14. [Google Scholar] [CrossRef]
- Jin, J. Korean Plant Ecology Trace; Nature and Ecology: Seoul, Korea, 2013; pp. 1063–1064. [Google Scholar]
- Cho, H.D.; Lee, K.W.; Won, Y.S.; Kim, J.H.; Seo, K.I. Cultivated Orostachys japonicus extract inhibits VEGF-induced angiogenesis via regulation of VEGFR2 signaling pathway in vitro and in vivo. J. Ethnopharmacol. 2020, 256, 1–11. [Google Scholar] [CrossRef]
- Park, H.J.; Young, H.S.; Park, K.Y.; Rhee, S.H.; Chung, H.Y.; Choi, J.S. Flavonoids from the whole plants of Orostachys japonicus. Arch. Pharmacal Res. 1991, 14, 167–171. [Google Scholar] [CrossRef]
- Park, J.G.; Park, J.C.; Hur, J.M.; Park, S.J.; Choi, D.R.; Shin, D.Y.; Park, K.Y.; Cho, H.W.; Kim, M.S. Phenolic compounds from Orostachys japonicus having anti-HIV-1 protease activity. Nat. Prod. Sci. 2000, 6, 117–121. [Google Scholar]
- Park, H.J.; Lim, S.C.; Lee, M.S.; Young, H.S. Triterpene and steroids from Orostachys japonicus. Korean J. Pharm. 1994, 25, 20–23. [Google Scholar]
- National Pharmacopoeia Committee of China Ministry of Health. Pharmacopoeia of the People’s Republic of China; People’s Medical Publishing House: Beijing, China, 2020; pp. 71–72. [Google Scholar]
- The Korean Herbal Pharmacopoeia. 2020. Available online: https://www.mfds.go.kr/brd/m_211/view.do?seq=14443&srchFr=&srchTo=&srchWord=&srchTp=&itm_seq_1=0&itm_seq_2=0&multi_itm_seq=0&company_cd=&company_nm=&page=1 (accessed on 4 January 2021).
- Ryu, D.S.; Lee, H.S.; Lee, G.S.; Lee, D.S. Effects of the ethylacetate extract of Orostachys japonicus on induction of apoptosis through the p53-mediated signaling pathway in human gastric cancer cells. Biol. Pharm. Bull. 2012, 35, 660–665. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.Y.; Choi, J.Y.; Lim, H.W.; Kim, J.; Kim, S.Y.; Han, C.W. Research on immune enhancing effect and safety of Wasong (Orostachys japonicus) extract: Study protocol for a single center, randomized, double-blind, placebo-controlled, clinical trial. Herb. Formula Sci. 2017, 25, 135–143. [Google Scholar] [CrossRef]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef]
- Galmarini, C.M.; Popowycz, F.; Joseph, B. Cytotoxic nucleoside analogues: Different strategies to improve their clinical efficacy. Curr. Med. Chem. 2008, 15, 1072–1082. [Google Scholar] [CrossRef] [PubMed]
- Ediriweera, M.K.; Tennekoon, K.H.; Samarakoon, S.R. In vitro assays and techniques utilized in anticancer drug discovery. J. Appl. Toxicol. 2019, 39, 38–71. [Google Scholar] [CrossRef]
- Kim, J.H.; Nam, G.S.; Kim, S.H.; Ryu, D.S.; Lee, D.S. Orostachys japonicus exerts antipancreatic cancer activity through induction of apoptosis and cell cycle arrest in PANC-1 cells. Food Sci. Nutr. 2019, 7, 3549–3559. [Google Scholar] [CrossRef]
- Ryu, D.H.; Ryu, D.S. Anticancer and signaling mechanisms of biologically active substances from Orostachys japonicus through arrest of cell cycle in human melanoma cells. J. Korean Med. Ophthalmol. Otolaryngol. Dermatol. 2019, 32, 1–12. [Google Scholar]
- Elmore, S. Apoptosis: A review of programmed cell death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef]
- Brown, J.M.; Attardi, L.D. The role of apoptosis in cancer development and treatment response. Nat. Rev. Cancer 2005, 5, 231–237. [Google Scholar] [CrossRef]
- Logue, S.E.; Martin, S.J. Caspase activation cascades in apoptosis. Biochem. Soc. Trans. 2008, 36, 1–9. [Google Scholar] [CrossRef]
- Yue, J.; López, J.M. Understanding MAPK signaling pathways in apoptosis. Int. J. Mol. Sci. 2020, 21, 2346. [Google Scholar] [CrossRef] [PubMed]
- Lunghi, P.; Tabilio, A.; Dall’Agilo, P.; Ridolo, E.; Carlo-Stela, C.; Pelicci, P.; Bonati, A. Downmodulation of ERK activity inhibits the proliferation and induces the apoptosis of primary acute myelogenous leukemia blasts. Leukemia 2003, 17, 1783–1793. [Google Scholar] [CrossRef]
- Tomita, Y.; Marchenko, N.; Erster, S.; Nemajerova, A.; Dehner, A.; Klein, C.; Pan, H.; Kessler, H.; Pancoska, P.; Moll, U.M. WT p53, but not tumor-derived mutants, bind to Bcl2 via the DNA binding domain and induce mitochondrial permeabilization. J. Biol. Chem. 2006, 281, 8600–8606. [Google Scholar] [CrossRef]
- Sax, J.K.; Fei, P.; Murphy, M.E.; Bernhard, E.; Korsmeyer, S.J.; El-Deiry, W.S. BID regulation by p53 contributes to chemosensitivity. Nat. Cell Biol. 2002, 4, 842–849. [Google Scholar] [CrossRef] [PubMed]
- Khazaei, S.; Esa, N.M.; Ramachandran, V.; Hamid, R.A.; Pandurangan, A.K.; Etemad, A.; Ismail, P. In vitro antiproliferative and apoptosis inducing effect of Allium atroviolaceum bulb extract on breast, cervical, and liver cancer cells. Front. Pharm. 2017, 8, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Lee, G.S.; Lee, H.S.; Kim, S.H.; Suk, D.H.; Ryu, D.S.; Lee, D.S. Anti-cancer activity of the ethylacetate fraction from Orostachys japonicus for modulation of the signaling pathway in HepG2 human hepatoma cells. Food Sci. Biotechnol. 2014, 23, 269–275. [Google Scholar] [CrossRef]
- Kim, J.Y.; Jung, E.J.; Won, Y.S.; Lee, J.H.; Shin, D.Y.; Seo, K.I. Cultivated Orostachys japonicus induces apoptosis in human colon cancer cells. Korean J. Food Sci. Technol. 2012, 44, 317–323. [Google Scholar] [CrossRef]
- Won, Y.S.; Lee, J.H.; Kwon, S.J.; Ahn, D.U.; Shin, D.Y.; Seo, K.I. Anticancer effects of cultivated Orostachys japonicus on human prostate cancer cells. J. Korean Soc. Food Sci. Nutr. 2014, 43, 67–73. [Google Scholar] [CrossRef]
- Ryu, D.S.; Kim, S.H.; Kwon, J.H.; Lee, D.S. Orostachys japonicus induces apoptosis and cell cycle arrest through the mitochondria-dependent apoptotic pathway in AGS human gastric cancer cells. Int. J. Oncol. 2014, 45, 459–469. [Google Scholar] [CrossRef]
- Choi, E.S.; Lee, J.H. Orostachys japonicus DW and EtOH extracts induce apoptosis in cholangiocarcinoma cell line SNU-1079. J. Korean Med. 2015, 36, 19–34. [Google Scholar] [CrossRef]
- Kwon, J.; Lee, D.; Jung, E.; Kim, H.; Kim, S.; Ryu, D. Anti-cancer activity of the ethylactate fraction from Orostachys japonicus in A549 human lung cancer cells by induction of apoptosis and cell cycle arrest. AJMAHS 2017, 7, 395–405. [Google Scholar] [CrossRef]
- Ryu, D.S.; Lee, H.J.; Kwon, J.H.; Lee, D.S. Anti-cancer effect of ethylacetate fraction from Orostachys japonicus on HT-29 human colon cancer cells by induction of apoptosis through caspase-dependent signaling pathway. Asian Pac. J. Trop. Med. 2018, 11, 330–335. [Google Scholar]
- Kim, S.H. Anti-Cancer Mechanisms in Human Cervical Cancer Cells and Pre-Clinical Safety of the Biologically Active Substances from Orostachydis Herba; Inje University: Gimhae, Korea, 2014; pp. 1–134. [Google Scholar]
- Lee, K.S.; Kim, S.W.; Lee, H.S. Orostachys japonicus induce p53-dependent cell cycle arrest through the MAPK signaling pathway in OVCAR-3 human ovarian cancer cells. Food Sci. Nutr. 2018, 6, 2395–2401. [Google Scholar] [CrossRef]
- Kwon, J.H.; Kim, J.H.; Ryu, D.S.; Lee, H.J.; Lee, D.S. Anticancer effect of the ethyl acetate fraction from Orostachys japonicus on MDA-MB-231 human breast cancer cells through extensive induction of apoptosis, cell cycle arrest, and antimetastasis. Evid. Based Complement. Altern. Med. 2019, 2019, 1–10. [Google Scholar] [CrossRef]
- Joo, S.; Jang, E.; Kim, Y. Effect of Orostachys japonicus on apoptosis and autophagy in human monocytic leukemia cell line THP-1 via inhibition of NF-κB and phosphorylation of p38 MAPK. J. Korean Med. 2019, 40, 35–50. [Google Scholar] [CrossRef]
- Lee, W.S.; Yun, J.W.; Nagappan, A.; Jung, J.H.; Yi, S.M.; Kim, D.H.; Kim, H.J.; Kim, G.; Ryu, C.H.; Shin, S.C. Flavonoids from Orostachys japonicus A. Berger induces caspase-dependent apoptosis at least partly through activation of p38 MAPK pathway in U937 human leukemic cells. Asian Pac. J. Cancer Prev. 2015, 16, 465–469. [Google Scholar] [CrossRef]
- Yun, K.S.; Kim, Y.C.; Lee, J.H.; Woo, H.J. Effect of Orostschys japonicus A. Berger on apoptosis in K562 cell lines. J. Int. Korean Med. 2006, 27, 166–177. [Google Scholar]
- Pellegata, N.S.; Antoniono, R.J.; Redpath, J.L.; Stanbridge, E.J. DNA damage and p53-mediated cell cycle arrest: A reevaluation. Proc. Natl. Acad. Sci. USA 1996, 93, 15209–15214. [Google Scholar] [CrossRef] [PubMed]
- Evan, G.I.; Vousden, K.H. Proliferation, cell cycle and apoptosis in cancer. Nature 2001, 411, 342–348. [Google Scholar] [CrossRef] [PubMed]
- Shah, M.A.; Schwartz, G.K. Cell cycle-mediated drug resistance: An emerging concept in cancer therapy. Clin. Cancer Res. 2001, 7, 2168–2181. [Google Scholar] [PubMed]
- Thill, M.; Schmidt, M. Management of adverse events during cyclin-dependent kinase 4/6 (CDK4/6) inhibitor-based treatment in breast cancer. Adv. Med. Oncol. 2018, 10, 1–12. [Google Scholar] [CrossRef]
- Bai, J.; Li, Y.; Zhang, G. Cell cycle regulation and anticancer drug discovery. Cancer Biol. Med. 2017, 14, 348–362. [Google Scholar] [PubMed]
- Di Sante, G.; Pagé, J.; Jiao, X.; Nawab, O.; Cristofanilli, M.; Skordalakes, E.; Pestell, R.G. Recent advances with cyclin-dependent kinase inhibitors: Therapeutic agents for breast cancer and their role in immuno-oncology. Expert Rev. Anticancer 2019, 19, 569–587. [Google Scholar] [CrossRef] [PubMed]
- Chen, J. The cell-cycle arrest and apoptotic functions of p53 in tumor initiation and progression. Cold Spring Harb. Perspect. Med. 2016, 6, 1–15. [Google Scholar] [CrossRef]
- Min, H.J. Metastatic Inhibition of Cultivated Orostachys Japonicus in Hypoxia Induced-Prostate Cancer Cells; Dong-A University: Busan, Korea, 2019; pp. 1–42. [Google Scholar]
- Kwon, J.; Han, K.S. Effects of Orostachys japonicus A. Berger on the immune system. Korean J. Med. Crop Sci. 2004, 12, 315–320. [Google Scholar]
- Oh, C.H.; Bae, J.B.; Kim, N.S.; Jeon, H.; Han, K.S.; Lee, M.J.; Kwon, J. Effect of Orostachys japonicus A. Berger on apoptosis induction of human leukemia HL60 Cells. Korean J. Pharm. 2009, 40, 118–122. [Google Scholar]
- Martineau, C.; Abed, E.; Médina, G.; Jomphe, L.A.; Mantha, M.; Jumarie, C.; Moreau, R. Involvement of transient receptor potential melastatin-related 7 (TRPM7) channels in cadmium uptake and cytotoxicity in MC3T3-E1 osteoblasts. Toxicol. Lett. 2010, 199, 357–363. [Google Scholar] [CrossRef]
- Kim, J.S.; Yoon, S.H.; Ryu, B.H.; Ryu, K.W. Anti-cancer effects of Orostachyos Herba on some kinds of cancer cells. J. Int. Korean Med. 2005, 26, 333–340. [Google Scholar]
- Sohn, S.H.; Park, H.S. Effects of Orostachys japhonicus herbal-acupuncture on transferred hepatic cancer of mouse induced by Colon26-L5 human colon cancer cells. J. Acupunct. Res. 2006, 23, 61–76. [Google Scholar]
- Park, S.; Won, J.; Park, K.; Hong, Y. Anti-cancer effects of cultivated Orostachys japonicus on human colon cancer cell line SW480. J. Life Sci. 2018, 28, 819–826. [Google Scholar]
- Kim, K.S.; Yea, S.C.; Yoo, B.C.; Cho, C.K.; Lee, Y.W.; Yoo, H.S. Altered protein expression in ovarian and cervical cancer cells by the treatment of extracts from Euonymus alatus Sieb, Oldenlandia diffusa (Willd.) Roxburgh, and Orostachys japonicus A. Berger. J. Int. Korean Med. 2011, 32, 33–42. [Google Scholar]
- Ryu, D.S.; Baek, G.O.; Kim, E.Y.; Kim, K.H.; Lee, D.S. Effects of polysaccharides derived from Orostachys japonicus on induction of cell cycle arrest and apoptotic cell death in human colon cancer cells. BMB Rep. 2010, 43, 750–755. [Google Scholar]
- Cho, H.D.; Lee, K.W.; Won, Y.S.; Shin, D.Y.; Seo, K.I. Studies on the anti-angiogenic activities of wild and cultivated Orostachys japonicus extracts in human umbilical vein endothelial cells. J. Food Sci. 2019, 84, 1764–1775. [Google Scholar] [CrossRef]
- Favier, J.; Corvol, P. Physiological angiogenesis. Therapie 2001, 56, 455–463. [Google Scholar]
- Carmeliet, P.; Jain, R.K. Angiogenesis in cancer and other diseases. Nature 2000, 407, 249–257. [Google Scholar] [CrossRef] [PubMed]
- Folkman, J. Role of angiogenesis in tumor growth and metastasis. Semin. Oncol. 2002, 29, 15–18. [Google Scholar] [CrossRef]
- Chen, H.X.; Cleck, J.N. Adverse effects of anticancer agents that target the VEGF pathway. Nat. Rev. Clin. Oncol. 2009, 6, 465–477. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.G.; Kim, J.S.; Lee, H.S.; Lim, Y.M.; So, J.H.; Hahn, D.; Ha, Y.S.; Nam, Y.O. Bioconverted Orostachys japonicas extracts suppress angiogenic activity of Ms-1 endothelial cells. Int. J. Mol. Sci. 2017, 18, 2615. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.K.; Kim, Y.C.; Lee, J.H.; Woo, H.K. The Effects of Artermisiae Capillaris Herba, Curcumae Rhizoma, Loranthi Ramulus, and Orostachys Herba on expression of angiogenic factors in HepG2 Cells. J. Int. Korean Med. 2007, 28, 149–165. [Google Scholar] [CrossRef]
- Bravo-Cordero, J.J.; Hodgson, L.; Condeelis, J. Directed cell invasion and migration during metastasis. Curr. Opin. Cell Biol. 2012, 24, 277–283. [Google Scholar] [CrossRef]
- Mehlen, P.; Puisieux, A. Metastasis: A question of life or death. Nat. Rev. Cancer 2006, 6, 449–458. [Google Scholar] [CrossRef]
- Qian, C.N.; Mei, Y.; Zhang, J. Cancer metastasis: Issues and challenges. Chin. J. Cancer 2017, 36, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Yeung, K.T.; Yang, J. Epithelial–mesenchymal transition in tumor metastasis. Mol. Oncol. 2017, 11, 28–39. [Google Scholar] [CrossRef]
- Orlichenko, L.S.; Radisky, D.C. Matrix metalloproteinases stimulate epithelial-mesenchymal transition during tumor development. Clin. Exp. Metastasis 2008, 25, 593–600. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.I.; Park, S.W.; Yoon, Y.K.; Lee, K.W.; Lee, J.H.; Woo, H.J.; Kim, Y. Orostachys japonicus inhibits the expression of MMP-2 and MMP-9 mRNA and modulates the expression of iNOS and COX-2 genes in human PMA-differentiated THP-1 cells via inhibition of NF-κB and MAPK activation. Mol. Med. Rep. 2015, 12, 657–662. [Google Scholar] [CrossRef] [PubMed]
- Li, L.H. Orostachys Japonicus Extract Inhibits Matrix Metalloproteinase-2 and -9 Expression in HT1080 Cells; Chonnam National University: Gwangju, Korea, 2006; pp. 1–23. [Google Scholar]
- Shin, D.Y.; Lee, W.S.; Jung, J.H.; Hong, S.H.; Park, C.; Kim, H.J.; Kim, G.Y.; Hwang, H.J.; Kim, G.S.; Jung, J.M. Flavonoids from Orostachys japonicus A. Berger inhibit the invasion of LnCaP prostate carcinoma cells by inactivating Akt and modulating tight junctions. Int. J. Mol. Sci. 2013, 14, 18407–18420. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Ryu, S.Y. Antimetastatic effect of several crude drugs with antitumor activity on B16-Fo and A549 cells (I). J. Korean Med. 1996, 17, 111–131. [Google Scholar]
- Gay, L.J.; Felding-Habermann, B. Contribution of platelets to tumour metastasis. Nat. Rev. Cancer 2011, 11, 123–134. [Google Scholar] [CrossRef]
- Fu, B.; Wang, N.; Tan, H.-Y.; Li, S.; Cheung, F.; Feng, Y. Multi-component herbal products in the prevention and treatment of chemotherapy-associated toxicity and side effects: A review on experimental and clinical evidences. Front. Pharm. 2018, 9, 1–15. [Google Scholar] [CrossRef]
- Wu, J.; Liu, Y.; Fang, C.; Zhao, L.; Lin, L.; Lu, L. Traditional Chinese medicine preparation combined therapy may improve chemotherapy efficacy: A systematic review and meta-analysis. Evid. Based Complement. Altern. Med. 2019, 1, 1–8. [Google Scholar] [CrossRef]
- Zhang, X.P.; Li, Y.D.; Luo, L.L.; Liu, Y.Q.; Li, Y.; Guo, C.; Li, Z.D.; Xie, X.R.; Song, H.X.; Yang, L.P.; et al. Astragalus Saponins and liposome constitute an efficacious adjuvant formulation for cancer vaccines. Cancer Biother. Radiopharm. 2018, 33, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Chang, J.M.; Hung, L.M.; Chyan, Y.J.; Cheng, C.M.; Wu, R.Y. Carthamus tinctorius enhances the antitumor activity of dendritic cell vaccines via polarization toward Th1 cytokines and increase of cytotoxic T lymphocytes. Evid. Based Complement. Altern. Med. 2011, 2011, 1–10. [Google Scholar]
- Lee, H.Y.; Park, Y.M.; Kim, J.; Oh, H.G.; Kim, K.S.; Kang, H.J.; Kim, R.R.; Kim, M.J.; Kim, S.H.; Yang, H.J.; et al. Orostachys japonicus A. Berger extracts induce immunity-enhancing effects on cyclophosphamide-treated immunosuppressed rats. Biomed. Res. Int. 2019, 2019, 1–9. [Google Scholar] [CrossRef]
- Vyas, D.; Laput, G.; Vyas, A.K. Chemotherapy-enhanced inflammation may lead to the failure of therapy and metastasis. Oncotargets 2014, 7, 1015–1023. [Google Scholar] [CrossRef]
- Yoon, Y.K.; Woo, H.J.; Kim, Y. Orostachys japonicus inhibits expression of the TLR4, NOD2, iNOS, and COX-2 genes in LPS-stimulated human PMA-differentiated THP-1 cells by inhibiting NF-κB and MAPK activation. Evid. Based Complement. Altern. Med. 2015, 2015, 1–9. [Google Scholar] [CrossRef]
- Xin, M.G. Anti-Inflammatory Effects of Orostachys Japonicus Fractions in TNF-α-Stimulated HT-29 Human Colon Epithelial Cells; Won Kwang University: Iksan, Korea, 2011; pp. 1–40. [Google Scholar]
- Arif, H.; Sohail, A.; Farhan, M.; Rehman, A.A.; Ahmad, A.; Hadi, S. Flavonoids-induced redox cycling of copper ions leads to generation of reactive oxygen species: A potential role in cancer chemoprevention. Int. J. Biol. Macromol. 2018, 106, 569–578. [Google Scholar] [CrossRef]
- Uusitalo, L.; Salmenhaara, M.; Isoniemi, M.; Garcia-Alvarez, A.; Serra-Majem, L.; Ribas-Barba, L.; Finglas, P.; Plumb, J.; Tuominen, P.; Savela, K. Intake of selected bioactive compounds from plant food supplements containing fennel (Foeniculum vulgare) among Finnish consumers. Food Chem. 2016, 194, 619–625. [Google Scholar] [CrossRef]
- Zhang, L.; Gao, H.Y.; Baba, M.; Okada, Y.; Okuyama, T.; Wu, L.J.; Zhan, L.B. Extracts and compounds with anti-diabetic complications and anti-cancer activity from Castanea mollissina Blume (Chinese chestnut). BMC Complement. Altern. Med. 2014, 14, 1–9. [Google Scholar] [CrossRef]
- Ko, M.J.; Nam, H.H.; Chung, M.S. Subcritical water extraction of bioactive compounds from Orostachys japonicus A. Berger (Crassulaceae). Sci. Rep. 2020, 10, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Devi, K.P.; Malar, D.S.; Nabavi, S.F.; Sureda, A.; Xiao, J.; Nabavi, S.M.; Daglia, M. Kaempferol and inflammation: From chemistry to medicine. Pharm. Res. 2015, 99, 1–10. [Google Scholar] [CrossRef]
- Chen, A.Y.; Chen, Y.C. A review of the dietary flavonoid, kaempferol on human health and cancer chemoprevention. Food Chem. 2013, 138, 2099–2107. [Google Scholar] [CrossRef]
- Imran, M.; Salehi, B.; Sharifi-Rad, J.; Aslam Gondal, T.; Saeed, F.; Imran, A.; Shahbaz, M.; Tsouh Fokou, P.V.; Umair Arshad, M.; Khan, H. Kaempferol: A key emphasis to its anticancer potential. Molecules 2019, 24, 2277. [Google Scholar] [CrossRef]
- Wang, X.; Yang, Y.; An, Y.; Fang, G. The mechanism of anticancer action and potential clinical use of kaempferol in the treatment of breast cancer. Biomed. Pharm. 2019, 117, 1–6. [Google Scholar] [CrossRef]
- Sharma, V.; Joseph, C.; Ghosh, S.; Agarwal, A.; Mishra, M.K.; Sen, E. Kaempferol induces apoptosis in glioblastoma cells through oxidative stress. Mol. Cancer Ther. 2007, 6, 2544–2553. [Google Scholar] [CrossRef]
- Li, Q.; Wei, L.; Lin, S.; Chen, Y.; Lin, J.; Peng, J. Synergistic effect of kaempferol and 5-fluorouracil on the growth of colorectal cancer cells by regulating the PI3K/Akt signaling pathway. Mol. Med. Rep. 2019, 20, 728–734. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Chen, A.Y.; Li, M.; Chen, C.; Yao, Q. Ginkgo biloba extract kaempferol inhibits cell proliferation and induces apoptosis in pancreatic cancer cells. J. Surg. Res. 2008, 148, 17–23. [Google Scholar] [CrossRef] [PubMed]
- El-Kott, A.; Shati, A.; Al-Kahtani, M.; Alharbi, S. Kaempferol induces cell death in A2780 ovarian cancer cells and increases their sensitivity to cisplatin by activation of cytotoxic endoplasmic reticulum-mediated autophagy and inhibition of protein kinase B. Folia Biol. 2020, 66, 36–46. [Google Scholar]
- Luo, H.; Daddysman, M.K.; Rankin, G.O.; Jiang, B.-H.; Chen, Y.C. Kaempferol enhances cisplatin’s effect on ovarian cancer cells through promoting apoptosis caused by down regulation of cMyc. Cancer Cell Int. 2010, 10, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Riahi-Chebbi, I.; Souid, S.; Othman, H.; Haoues, M.; Karoui, H.; Morel, A.; Srairi-Abid, N.; Essafi, M.; Essafi-Benkhadir, K. The Phenolic compound Kaempferol overcomes 5-fluorouracil resistance in human resistant LS174 colon cancer cells. Sci. Rep. 2019, 9, 1–20. [Google Scholar] [CrossRef]
- Nair, B.; Anto, R.J.; Sabitha, M.; Nath, L.R. Kaempferol-mediated sensitization enhances chemotherapeutic efficacy of sorafenib against hepatocellular carcinoma: An in silico and in vitro approach. Adv. Pharm. Bull. 2020, 10, 472–476. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J.; Sun, G.B.; Sun, B.; Wu, Y.; He, L.; Wang, X.; Chen, R.C.; Cao, L.; Ren, X.Y.; Sun, X.B. Kaempferol protects against doxorubicin-induced cardiotoxicity in vivo and in vitro. Toxicology 2012, 292, 53–62. [Google Scholar] [CrossRef]
- Wu, W.; Yang, B.; Qiao, Y.; Zhou, Q.; He, H.; He, M. Kaempferol protects mitochondria and alleviates damages against endotheliotoxicity induced by doxorubicin. Biomed. Pharm. 2020, 126, 1–8. [Google Scholar] [CrossRef]
- Wang, Z.; Sun, W.; Sun, X.; Wang, Y.; Zhou, M. Kaempferol ameliorates Cisplatin induced nephrotoxicity by modulating oxidative stress, inflammation and apoptosis via ERK and NF-κB pathways. AMB Express 2020, 10, 1–11. [Google Scholar] [CrossRef]
- Xu, L.; Zhang, Y.; Zhang, P.; Dai, X.; Gao, Y.; Lv, Y.; Qin, S.; Xu, F. Integrated metabolomics and network pharmacology strategy-driven active traditional Chinese medicine ingredients discovery for the alleviation of cisplatin nephrotoxicity. Chem. Res. Toxicol. 2019, 32, 2411–2421. [Google Scholar] [CrossRef]
- Gao, S.S.; Choi, B.M.; Chen, X.Y.; Zhu, R.Z.; Kim, Y.; So, H.S.; Park, R.; Sung, M.; Kim, B.R. Kaempferol suppresses cisplatin-induced apoptosis via inductions of heme oxygenase-1 and glutamate-cysteine ligase catalytic subunit in HEI-OC1 cells. Pharm. Res. 2010, 27, 235–245. [Google Scholar] [CrossRef]
- Dabeek, W.M.; Marra, M.V. Dietary quercetin and kaempferol: Bioavailability and potential cardiovascular-related bioactivity in humans. Nutrients 2019, 11, 2288. [Google Scholar] [CrossRef]
- Eid, H.M.; Haddad, P.S. The antidiabetic potential of quercetin: Underlying mechanisms. Curr. Med. Chem. 2017, 24, 355–364. [Google Scholar] [PubMed]
- Ebrahimpour, S.; Zakeri, M.; Esmaeili, A. Crosstalk between obesity, diabetes, and Alzheimer’s disease: Introducing quercetin as an effective triple herbal medicine. Ageing Res. Rev. 2020, 62, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Sato, S.; Mukai, Y. Modulation of chronic inflammation by quercetin: The beneficial effects on obesity. J. Inflamm. Res. 2020, 13, 421–431. [Google Scholar] [CrossRef]
- Li, Y.; Yao, J.; Han, C.; Yang, J.; Chaudhry, M.T.; Wang, S.; Liu, H.; Yin, Y. Quercetin, inflammation and immunity. Nutrients 2016, 8, 167. [Google Scholar] [CrossRef]
- McCann, S.E.; Ambrosone, C.B.; Moysich, K.B.; Brasure, J.; Marshall, J.R.; Freudenheim, J.L.; Wilkinson, G.S.; Graham, S. Intakes of selected nutrients, foods, and phytochemicals and prostate cancer risk in western New York. Nutr. Cancer 2005, 53, 33–41. [Google Scholar] [CrossRef]
- Sargiacomo, C.; Sotgia, F.; Lisanti, M.P. COVID-19 and chronological aging: Senolytics and other anti-aging drugs for the treatment or prevention of corona virus infection? Aging 2020, 12, 6511–6517. [Google Scholar] [CrossRef]
- Jana, N.; Břetislav, G.; Pavel, S.; Pavla, U. Potential of the flavonoid quercetin to prevent and treat cancer-current status of research. Klin. Onkol. 2018, 31, 184–190. [Google Scholar]
- Parvaresh, A.; Razavi, R.; Rafie, N.; Ghiasvand, R.; Pourmasoumi, M.; Miraghajani, M. Quercetin and ovarian cancer: An evaluation based on a systematic review. J. Res. Med. Sci. 2016, 21, 1–7. [Google Scholar]
- Fernández-Palanca, P.; Fondevila, F.; Méndez-Blanco, C.; Tuñón, M.J.; González-Gallego, J.; Mauriz, J.L. Antitumor effects of quercetin in hepatocarcinoma in vitro and in vivo models: A systematic review. Nutrients 2019, 11, 2875. [Google Scholar] [CrossRef] [PubMed]
- Shen, X.; Si, Y.; Wang, Z.; Wang, J.; Guo, Y.; Zhang, X. Quercetin inhibits the growth of human gastric cancer stem cells by inducing mitochondrial-dependent apoptosis through the inhibition of PI3K/Akt signaling. Int. J. Mol. Med. 2016, 38, 619–626. [Google Scholar] [CrossRef]
- Choi, E.J.; Bae, S.M.; Ahn, W.S. Antiproliferative effects of quercetin through cell cycle arrest and apoptosis in human breast cancer MDA-MB-453 cells. Arch. Pharmacal. Res. 2008, 31, 1281–1285. [Google Scholar] [CrossRef] [PubMed]
- Chou, C.C.; Yang, J.S.; Lu, H.F.; Ip, S.W.; Lo, C.; Wu, C.C.; Lin, J.P.; Tang, N.Y.; Chung, J.G.; Chou, M.J. Quercetin-mediated cell cycle arrest and apoptosis involving activation of a caspase cascade through the mitochondrial pathway in human breast cancer MCF-7 cells. Arch. Pharmacal. Res. 2010, 33, 1181–1191. [Google Scholar] [CrossRef]
- Pratheeshkumar, P.; Budhraja, A.; Son, Y.O.; Wang, X.; Zhang, Z.; Ding, S.; Wang, L.; Hitron, A.; Lee, J.C.; Xu, M. Quercetin inhibits angiogenesis mediated human prostate tumor growth by targeting VEGFR-2 regulated AKT/mTOR/P70S6K signaling pathways. PLoS ONE 2012, 7, e47516. [Google Scholar] [CrossRef]
- Balakrishnan, S.; Bhat, F.; Raja Singh, P.; Mukherjee, S.; Elumalai, P.; Das, S.; Patra, C.; Arunakaran, J. Gold nanoparticle–conjugated quercetin inhibits epithelial–mesenchymal transition, angiogenesis and invasiveness via EGFR/VEGFR-2-mediated pathway in breast cancer. Cell Prolif. 2016, 49, 678–697. [Google Scholar] [CrossRef]
- Li, S.Z.; Yuan, S.; Zhao, Q.; Wang, B.; Wang, X.; Li, K. Quercetin enhances chemotherapeutic effect of doxorubicin against human breast cancer cells while reducing toxic side effects of it. Biomed. Pharm. 2018, 100, 441–447. [Google Scholar] [CrossRef]
- Wang, G.; Zhang, J.; Liu, L.; Sharma, S.; Dong, Q. Quercetin potentiates doxorubicin mediated antitumor effects against liver cancer through p53/Bcl-xl. PLoS ONE 2012, 7, e51764. [Google Scholar] [CrossRef]
- Atashpour, S.; Fouladdel, S.; Movahhed, T.K.; Barzegar, E.; Ghahremani, M.H.; Ostad, S.N.; Azizi, E. Quercetin induces cell cycle arrest and apoptosis in CD133+ cancer stem cells of human colorectal HT29 cancer cell line and enhances anticancer effects of doxorubicin. Iran. J. Basic Med. Sci. 2015, 18, 635–643. [Google Scholar]
- Najafi, M.; Tavakol, S.; Zarrabi, A.; Ashrafizadeh, M. Dual role of quercetin in enhancing the efficacy of cisplatin in chemotherapy and protection against its side effects: A review. Arch. Physiol. Biochem. 2020, 2020, 1–15. [Google Scholar] [CrossRef]
- Dai, W.; Gao, Q.; Qiu, J.; Yuan, J.; Wu, G.; Shen, G. Quercetin induces apoptosis and enhances 5-FU therapeutic efficacy in hepatocellular carcinoma. Tumor Biol. 2016, 37, 6307–6313. [Google Scholar] [CrossRef] [PubMed]
- Brito, A.F.; Ribeiro, M.; Abrantes, A.M.; Mamede, A.C.; Laranjo, M.; Casalta-Lopes, J.E.; Gonçalves, A.C.; Sarmento-Ribeiro, A.B.; Tralhão, J.G.; Botelho, M.F. New approach for treatment of primary liver tumors: The role of quercetin. Nutr. Cancer 2016, 68, 250–266. [Google Scholar] [CrossRef]
- Lei, C.S.; Hou, Y.C.; Pai, M.H.; Lin, M.T.; Yeh, S.L. Effects of quercetin combined with anticancer drugs on metastasis-associated factors of gastric cancer cells: In vitro and in vivo studies. J. Nutr. Biochem. 2018, 51, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Zhang, J.; Wang, K.; Han, W.; Wang, X.; Gao, M.; Wang, Z.; Sun, Y.; Yan, H.; Zhang, H. Quercetin overcomes colon cancer cells resistance to chemotherapy by inhibiting solute carrier family 1, member 5 transporter. Eur. J. Pharm. 2020, 881, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R.; Gatchie, L.; Williams, I.S.; Jain, S.K.; Vishwakarma, R.A.; Chaudhuri, B.; Bharate, S.B. Glycyrrhiza glabra extract and quercetin reverses cisplatin resistance in triple-negative MDA-MB-468 breast cancer cells via inhibition of cytochrome P450 1B1 enzyme. Bioorg. Med. Chem. Lett. 2017, 27, 5400–5403. [Google Scholar] [CrossRef]
- Ahmed, Z.A.; Abtar, A.N.; Othman, H.H.; Aziz, T.A. Effects of quercetin, sitagliptin alone or in combination in testicular toxicity induced by doxorubicin in rats. Drug Des. Devel. Ther. 2019, 13, 3321–3329. [Google Scholar] [CrossRef]
- Chen, X.; Peng, X.; Luo, Y.; You, J.; Yin, D.; Xu, Q.; He, H.; He, M. Quercetin protects cardiomyocytes against doxorubicin-induced toxicity by suppressing oxidative stress and improving mitochondrial function via 14-3-3γ. Toxicol. Mech. Methods 2019, 29, 344–354. [Google Scholar] [CrossRef]
- Mahabady, M.K.; Gholami, M.R.; Varzi, H.N.; Zendedel, A.; Doostizadeh, M. Protective effect of quercetin on skeletal and neural tube teratogenicity induced by cyclophosphamide in rat fetuses. Vet. Res. Forum 2016, 7, 133–138. [Google Scholar]
- Hait, W.N. Anticancer drug development: The grand challenges. Nat. Rev. Drug Discov. 2010, 9, 253–254. [Google Scholar] [CrossRef]
- Ryu, D.S.; Lee, M.Y.; Lee, H.S.; Kim, S.H.; Lee, G.S.; Kwon, J.H.; Lee, D.S. A Repeated-dose oral toxicity study of Orostachys japonicus extract in Sprague-Dawley rats. J. Exp. Biomed. Sci. 2012, 18, 10–15. [Google Scholar]
- Kim, S.H.; Ryu, D.S.; Lee, H.S.; Shin, H.R.; Kwon, J.H.; Lee, D.S. Acute oral toxicity of the ethyl acetate fraction of Orostachys japonicus in mice. Pharm. Biol. 2014, 52, 1345–1350. [Google Scholar] [CrossRef]
- Orostachys japonica in Database for Raw Food Materials. Available online: https://www.foodsafetykorea.go.kr/portal/safefoodlife/foodMeterial/foodMeterialDB.do (accessed on 4 January 2021).
- Kohno, M.; Pouyssegur, J. Targeting the ERK signaling pathway in cancer therapy. Ann. Med. 2006, 38, 200–211. [Google Scholar] [CrossRef]
- Brown, L.; Benchimol, S. The involvement of MAPK signaling pathways in determining the cellular response to p53 activation: Cell cycle arrest or apoptosis. J. Biol. Chem. 2006, 281, 3832–3840. [Google Scholar] [CrossRef]
- Luangdilok, S.; Box, C.; Harrington, K.; Rhŷs-Evans, P.; Eccles, S.; Luangdilok, S. MAPK and PI3K signalling differentially regulate angiogenic and lymphangiogenic cytokine secretion in squamous cell carcinoma of the head and neck. Eur. J. Cancer 2011, 47, 520–529. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Wang, S.; Wang, Y.; Wu, H.; Frank, J.A.; Zhang, Z.; Luo, J.; Xu, M. Role of p38γ MAPK in regulation of EMT and cancer stem cells. Biochim. Biophys. Acta Mol. Basis Dis. 2018, 1864, 3605–3617. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Extract | Model | Concentration | Result | Mechanism | Ref |
|---|---|---|---|---|---|
| E,W | SNU-1079 (in vitro) | 300 μg/mL | ↓Intrahepatic cholangio-carcinoma cells | ↑Early and late apoptosis rate ↓Bcl-2, Mcl-1, Survivin mRNA ↑Bax mRNA ↑Cleaved caspase -3 | [36] |
| Cell cycle arrest ↓Cyclin D1 mRNA | |||||
| E | PMA-induced THP-1 cells (in vitro) | 300 μg/mL | ↓Leukemia cells | ↑Early and late apoptosis rate ↓Bcl-2, Mcl-1, Survivin mRNA ↑Bax mRNA ↑Cleaved caspase-3 | [42] |
| Autophagy ↑LC3 II, beclin-1 ↓Atg5 | |||||
| MAPK pathway ↑p38 protein | |||||
| ↓NF-κB ↓mTOR mRNA | |||||
| EA | AGS (in vitro) | 100, 200 μg/mL | ↓Gastric cancer cells (IC50: 86 μg/mL) | ↑Early and late apoptosis rate ↑DNA fragmentation ↑Apoptotic bodies ↓Bcl-2 ↑Cleaved caspase-3 | [17] |
| Cell cycle arrest Sub-G1 peak and G2/M arrest | |||||
| ↑Tumor suppressor p53 | |||||
| EA | AGS (in vitro) | 100 μg/mL | ↓Gastric cancer cells | ↑Early and late apoptosis rate ↓Bcl-2 ↑Cytochrome c ↓Pro-caspase-3,8,9 ↑Cleaved caspase-3,8,9 | [35] |
| Cell cycle arrest Sub-G1 peak and G2/M arrest ↓Cyclin B1, CDK1 mRNA | |||||
| MAPK pathway ↑p38, JNK protein | |||||
| ↑Tumor suppressor p53 | |||||
| EA | HeLa (in vitro) | 10 μg/mL | ↓Cervical cancer cells | ↑Early and late apoptosis rate ↓Bcl-2 ↑Cytochrome c ↓Pro-caspase-3,8,9 ↑Cleaved caspase-3,8,9 | [39] |
| Cell cycle arrest Sub-G1 peak and G2/M arrest ↓Cyclin B1, CDK1 mRNA | |||||
| MAPK pathway ↑p38, JNK protein | |||||
| EA | HepG2 (in vitro) | 100, 200 μg/mL | ↓Liver cancer cellsc | ↑Early and late apoptosis rate ↑Condensed chromatin ↑Fragmented nuclei ↑Apoptotic bodies ↓Bcl-2 (not Bax) ↑Cytochrome c ↓Pro-caspase-3,8,9 | [32] |
| MAPK pathway ↑p-JNK, p-ERK1/2 protein | |||||
| EA | A549 (in vitro) | 75, 100 μg/mL | ↓Lung cancer cells | ↑Early and late apoptosis rate ↑Condensed chromatin ↑Fragmented nuclei ↑Apoptotic bodies ↑Bax ↓Bcl-2 | [37] |
| Cell cycle arrest ↓Cyclin B1, CDK1, Cyclin D, CDK4 mRNA | |||||
| ↑Tumor suppressor p53 protein | |||||
| EA | HT-29 (in vitro) | 50, 75, 100 μg/mL | ↓Colon cancer cells | ↑Early and late apoptosis rate ↑Apoptotic bodies ↑Bax ↓Bcl-2 ↑Cleaved caspase -3,8,9 | [38] |
| Cell cycle arrest Sub-G1 peak | |||||
| MAPK pathway ↑p38, JNK, ERK1/2 protein | |||||
| EA | OVCAR-3 (in vitro) | 50 μg/mL | ↓Ovarian cancer cells | ↑Early and late apoptosis rate ↑Apoptotic bodies ↑Bax/Bcl-2 ratio | [40] |
| Cell cycle arrest Sub-G1 peak ↑p21 ↓Cyclin E1/CDK2 mRNA | |||||
| MAPK pathway ↑p38, ERK1/2 protein | |||||
| ↑Tumor suppressor p53 protein | |||||
| EA | PANC-1 (in vitro) | 50, 100 μg/mL | ↓Pancreatic cancer cells (IC50: 50 μg/mL) | ↑Early and late apoptosis rate ↑Condensed chromatin ↑Fragmented nuclei ↑Cytochrome c ↑Cleaved caspase-3,9 ↓Pro-caspase-3,8,9 | [22] |
| Cell cycle arrest Sub-G1 peak and G2/M arrest ↓Cyclin D1, Cyclin B1, CDK4 | |||||
| MAPK pathway ↑p38, JNK, ERK protein | |||||
| EA | MDA-MB-231 (in vitro) | 20, 40, 60 μg/mL | ↓Breast cancer cells | ↑Early and late apoptosis rate ↑Condensed chromatin ↑Fragmented nuclei ↑Cytochrome c ↓Pro-caspase-3,8,9 ↑Cleaved caspase-3,9 | [41] |
| Cell cycle arrest Sub-G1 peak and G2/M arrest ↓Cyclin D1, Cyclin B1, CDK4 | |||||
| MAPK pathway ↑p38, JNK, ERK protein | |||||
| EA | A375 (in vitro) | 120, 140, 150 μg/mL | ↓Melanoma cells | Cell cycle arrest Sub-G1 peak ↓CDK1, cyclin B1 ↓CDK4, cyclin D | [23] |
| E | Male Balb/c mice xenografted RC-58T/h/SA#4 (in vivo) | 25, 50 mg/kg | ↓Prostate cancer size and volume | N.A. | [52] |
| M | L1210, U937 (in vitro) | 100 μg/mL | ↓Leukemia cells | ↑Tumor suppressor p53 mRNA | [53] |
| Cell cycle arrest Sub-G1 peak | |||||
| Male Balb/c mice transplanted L1210 cells (in vivo) | 500 mg/kg | ↑Apoptosis rate of leukemia cells from peritoneal | ↑Early and late apoptosis rate | ||
| M | HL-60 (in vitro) | 100 μg/mL | ↓Leukemia cells | ↑Early and late apoptosis rate | [54] |
| Cell cycle arrest Sub-G1 peak | |||||
| ↑Tumor suppressor p53 gene/protein | |||||
| ↑NF-κB p50 | |||||
| M | AGS, MCF-7 (in vitro) | 300 μg/mL | ↓Cancer cells | ↑Early and late apoptosis rate ↑Mitochondria depolarization(ΔΨm) ↑Cleaved caspase-3 | [55] |
| Cell cycle arrest Sub-G1 peak | |||||
| M | SW480 (in vitro) | 500 μg/mL | ↓Colon cancer cells | ↑Apoptosis ↑Apoptotic bodies ↑Cleaved caspase-3,9 ↓Bid ↑t-Bid ↑Bax ↓Bcl-2 ↑PARP | [33] |
| Cell cycle arrest Sub-G1 peak | |||||
| M | RC-58T/h/SA#4 (in vitro) | 300, 600 μg/mL | ↓Prostate cancer cells ↑Floating cells | ↑Early and late apoptosis rate ↑Condensed chromatin ↑Fragmented nuclei ↑Apoptotic bodies | [34] |
| Cell cycle arrest Sub-G1 peak | |||||
| M | U937 (in vitro) | 200, 400 μg/mL | ↓Leukemia cells | ↑Apoptosis ↓Pro-caspase-3,8,9 ↓MMP(ΔΨm), Bcl-2 ↓Bid, XIAP ↑PARP | [43] |
| Cell cycle arrest Sub-G1 peak | |||||
| MAPK pathway ↑p38 protein | |||||
| ↓p-Akt protein | |||||
| W | HepG2, Hep2B, AGS, KATOIII (in vitro) | 20 μg/mL | ↓Cancer cells ↑Floating cells | N.A. | [56] |
| W | Colon 26-L5 (in vitro) | 300 μg/mL | ↓Colon cancer cells | N.A. | [57] |
| W | SW480 (in vitro) | N.A. | ↓Colon cancer cells ↑Floating cells | N.A. | [58] |
| Male C57BL/6 mice transplanted SW480 cells (in vivo) | N.A. | ↓Tumor weight and volume ↓Tumor formation | N.A. | ||
| W | OVCAR-3, HeLa (in vitro) | 100, 1000 μg/mL | ↓Cancer cells | ↑Apoptosis | [59] |
| ↓Cancer metabolism ↓hnRNP A2/B1 ↓Alpha-enolase ↓PHGDH | |||||
| MAPK pathway ↑p38 protein | |||||
| W | K562 (in vitro) | 100, μg/mL | ↓Chronic myeloid leukemia cells | ↑Early and late apoptosis rate ↑DNA fragmentation ↓Bcl-2 mRNA ↑Caspase-3 mRNA | [44] |
| W | HT-29 (in vitro) | 2 mg/mL | ↓Colon cancer cells shrunken, disintegrated, rounded, detached cells | ↑Early and late apoptosis rate ↑Apoptosis gene (BAD, FADD, caspase-3,8,9) ↑Tumor suppression gene (TP53BP2 and STAT1) ↓Cell proliferation and growth gene (PTK6) ↓Anti-apoptosis gene (Bcl-2) ↓Cell cycle regulation gene (RFC5) ↓Cancer development gene (CTSH) | [60] |
| Cell cycle arrest Sub-G1 peak and G2/M arrest | |||||
| W | A549, HeLa, AGS (in vitro) | 0.5, 1 mg/mL | ↓Cancer cells | ↑Early and late apoptosis rate ↑DNA fragmentation | [61] |
| Extract | Inducer | Model | Concentration | Result | Mechanism | Ref |
|---|---|---|---|---|---|---|
| M | None | Ms-1 (in vitro) | 100 μg/mL | ↓Wound closure area ↓Migrated cells ↓Infiltrated capillary structure ↓Adherent cells | ↓p-FAK protein ↓p- Src protein | [66] |
| M | VEGF 200 ng/mL | Male C57BL/6 mice (in vivo) | 0.1 mg/mL | ↓Blood vessels infiltrated into Matrigel plug | N.A. | |
| E, M, W, EA | VEGF 20 μg/mL | HUVECs (in vitro) | 20 μg/mL | ↓Wound closure area ↓Migrated cells ↓Infiltrated capillary structure | N.A. | [61] |
| E | VEGF 20 μg/mL | HUVECs (in vitro) | 10~20 μg/mL | ↓Wound closure area | ↓p-VEGFR2 protein ↓p-FAK protein ↓p-Src protein ↓MMP-1, -9 protein ↓p-PI3K/p-Akt/p-mTOR protein ↓p-p38, p-ERK1/2 protein | [11] |
| E | VEGF-A 50 ng/mL | HUVECs (in vitro) | 10~20 μg/mL | ↓Migrated cells ↓Infiltrated capillary structure | ||
| E | VEGF 20 ng/mL | SD Rat aorta ring (ex vivo) | 10~20 μg/mL | ↓Sprouts from aortic rings | ||
| E | VEGF 150 ng/mL | Male C57BL/6 mice (in vivo) | 10~20 μg/mL for 6d | ↓Blood vessels infiltrated into Matrigel plug | ||
| E | RC-58T/h/SA#4 cells | Male Balb/c mice (in vivo) | 25, 50 mg/kg | ↓Blood vessels infiltrated into tumor | N.A. | [52] |
| W | None | HepG2 cells (in vitro) | 25 μg/mL | ↓VEGF, bFGF mRNA, protein ↓Extracellular secretion of VEGF, bFGF | ↓p-Akt protein ↓ p-ERK protein | [67] |
| Extract | Inducer | Model | Concentration | Result | Mechanism | Ref |
|---|---|---|---|---|---|---|
| EA | None | MDA-MB-231 (in vitro) | 20, 40, 60 μg/mL | ↓Wound closure area | ↓MMP-9 protein | [41] |
| M | PMA | THP-1 (in vitro) | 10, 25 μg/mL | ↓Adherent cells | ↓MMP-2/9 mRNA ↓p-p38, p-ERK protein ↓NF-κB p65 protein | [73] |
| E | None | HT1080 (in vitro) | 200, 300 μg/mL | ↓Migrated cells | ↓MMP-2/9 protein | [74] |
| M | None | LNCaP (in vitro) | 200 μg/mL | ↓Wound closure area ↓Migrated cells | ↓MMP-2/9 mRNA/protein ↑TIMP-1/2 protein ↓Claudin-1/3 mRNA ↓p-Akt prtoein | [75] |
| E | Hypoxia | RC-58T/h/SA#4 (in vitro) | 50, 100 μg/mL | ↓Wound closure area ↓Migrated cells | ↓HIF-1α protein ↓p-FAK protein ↓uPA protein ↓p-ERK1/2 protein ↓PI3K/Akt protein | [52] |
| M | Collagen I/IV | A549 (in vitro) | 400 μg/mL | ↓Adherent cells | N.A | [76] |
| Laminin | B16-F0 (in vitro) | 400 μg/mL | ↓Adherent cells |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hur, S.; Jang, E.; Lee, J.-H. Beneficial Actions of Orostachys japonica and Its Compounds against Tumors via MAPK Signaling Pathways. Nutrients 2021, 13, 555. https://doi.org/10.3390/nu13020555
Hur S, Jang E, Lee J-H. Beneficial Actions of Orostachys japonica and Its Compounds against Tumors via MAPK Signaling Pathways. Nutrients. 2021; 13(2):555. https://doi.org/10.3390/nu13020555
Chicago/Turabian StyleHur, Soyoung, Eungyeong Jang, and Jang-Hoon Lee. 2021. "Beneficial Actions of Orostachys japonica and Its Compounds against Tumors via MAPK Signaling Pathways" Nutrients 13, no. 2: 555. https://doi.org/10.3390/nu13020555
APA StyleHur, S., Jang, E., & Lee, J.-H. (2021). Beneficial Actions of Orostachys japonica and Its Compounds against Tumors via MAPK Signaling Pathways. Nutrients, 13(2), 555. https://doi.org/10.3390/nu13020555