
High β-Glucan Barley Supplementation Improves Glucose Tolerance by Increasing GLP-1 Secretion in Diet-Induced Obesity Mice


Abstract
1. Introduction
2. Materials and Methods
2.1. Chemical Composition of High β-Glucan Barley
2.2. Experimental Design
2.3. Biochemical Analyses of the Serum
2.4. Analysis of Short-Chain Fatty Acids in Cecal Digesta
2.5. Measurement of Total GLP-1 Level in the Cecum
2.6. mRNA Expression Analysis in the Ileum
2.7. Measurement of the Number of L Cells in the Ileum
2.8. Statistical Analyses
3. Results
3.1. Gross Changes to Mice during the Study Period
3.2. Biochemical Markers in the Serum
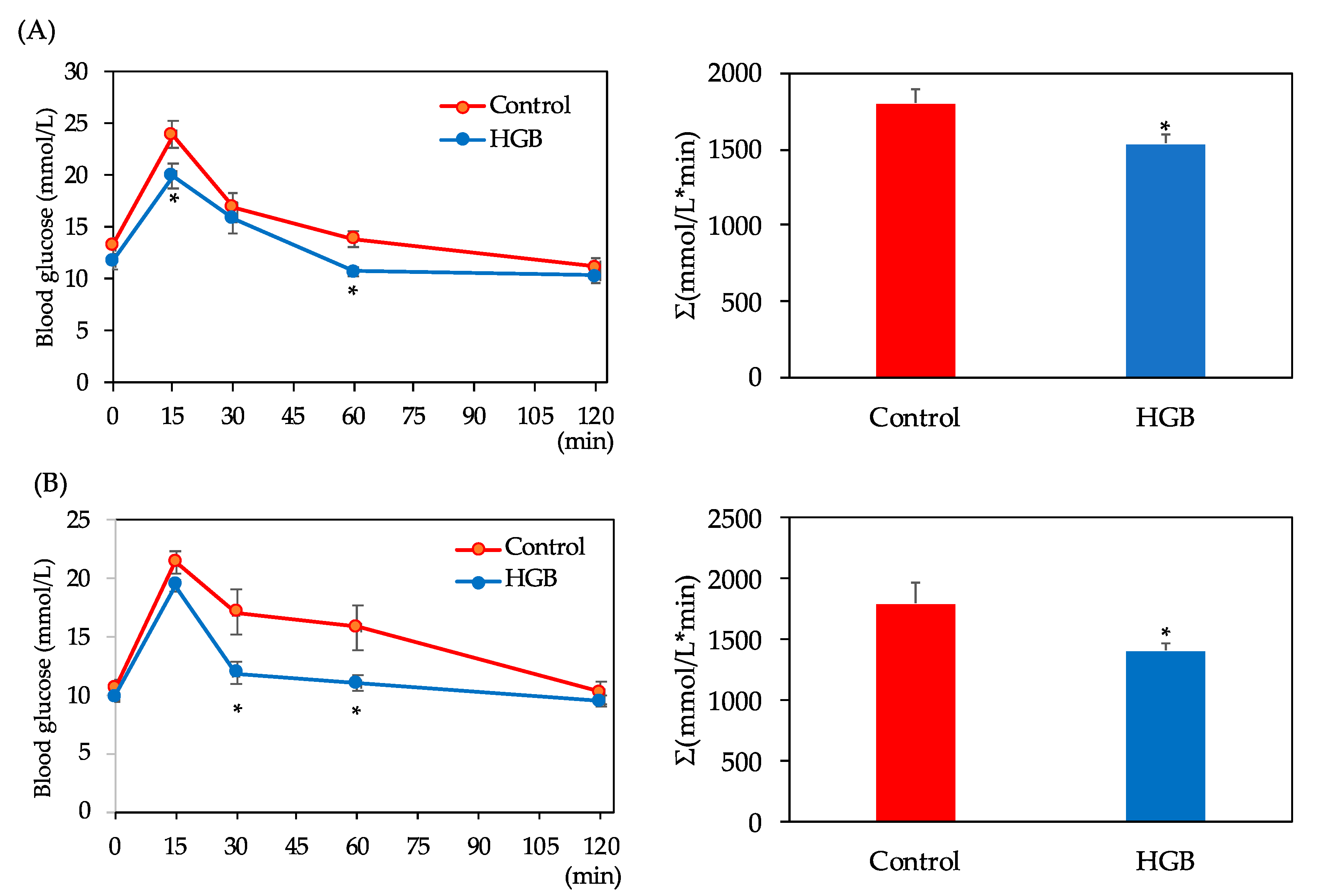
3.3. Assessment of Glucose Tolerance
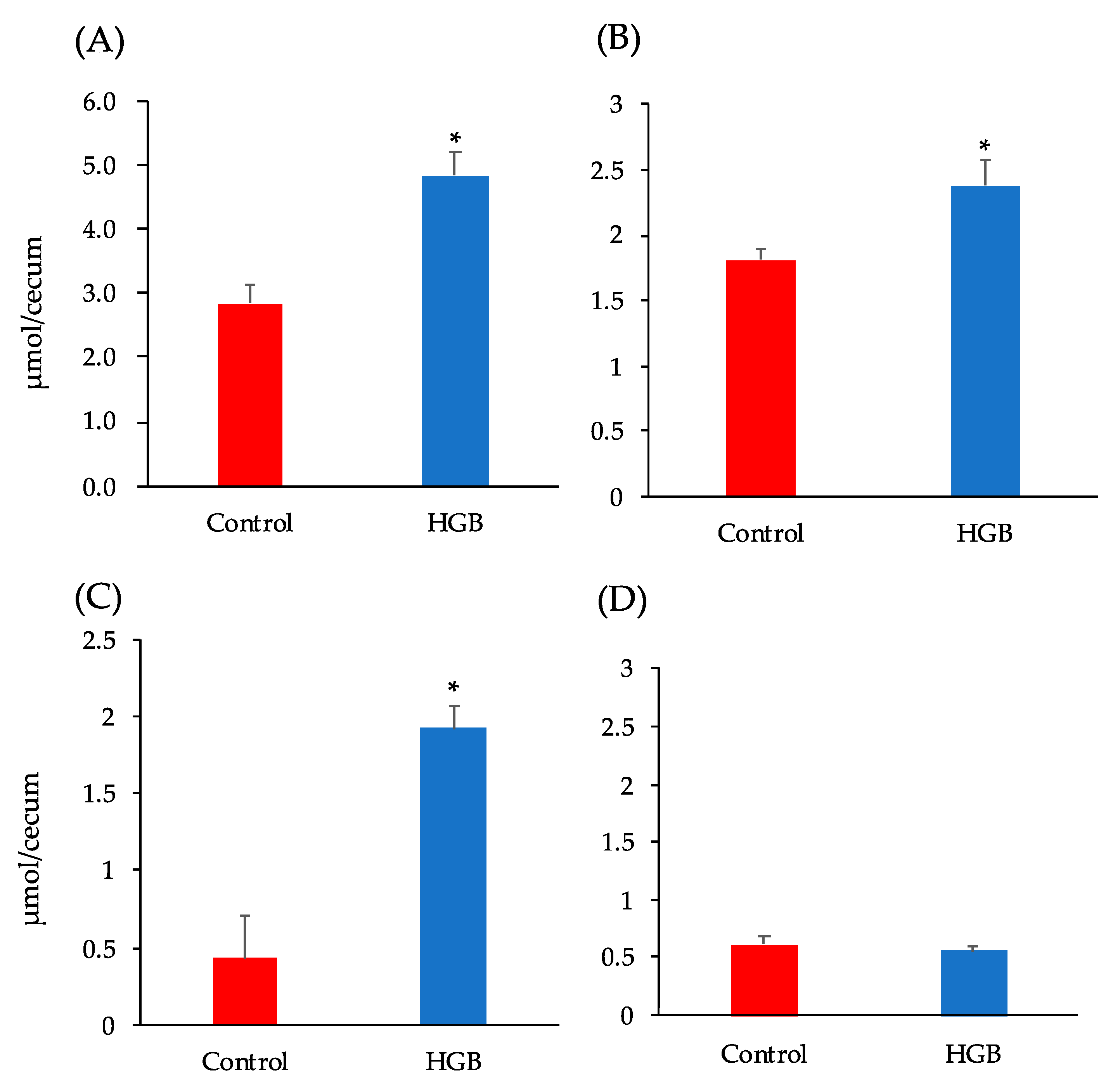
3.4. Analysis of Short-Chain Fatty Acids in Cecal Digesta
3.5. Concentrations of GLP-1 in the Portal Vein and Cecum
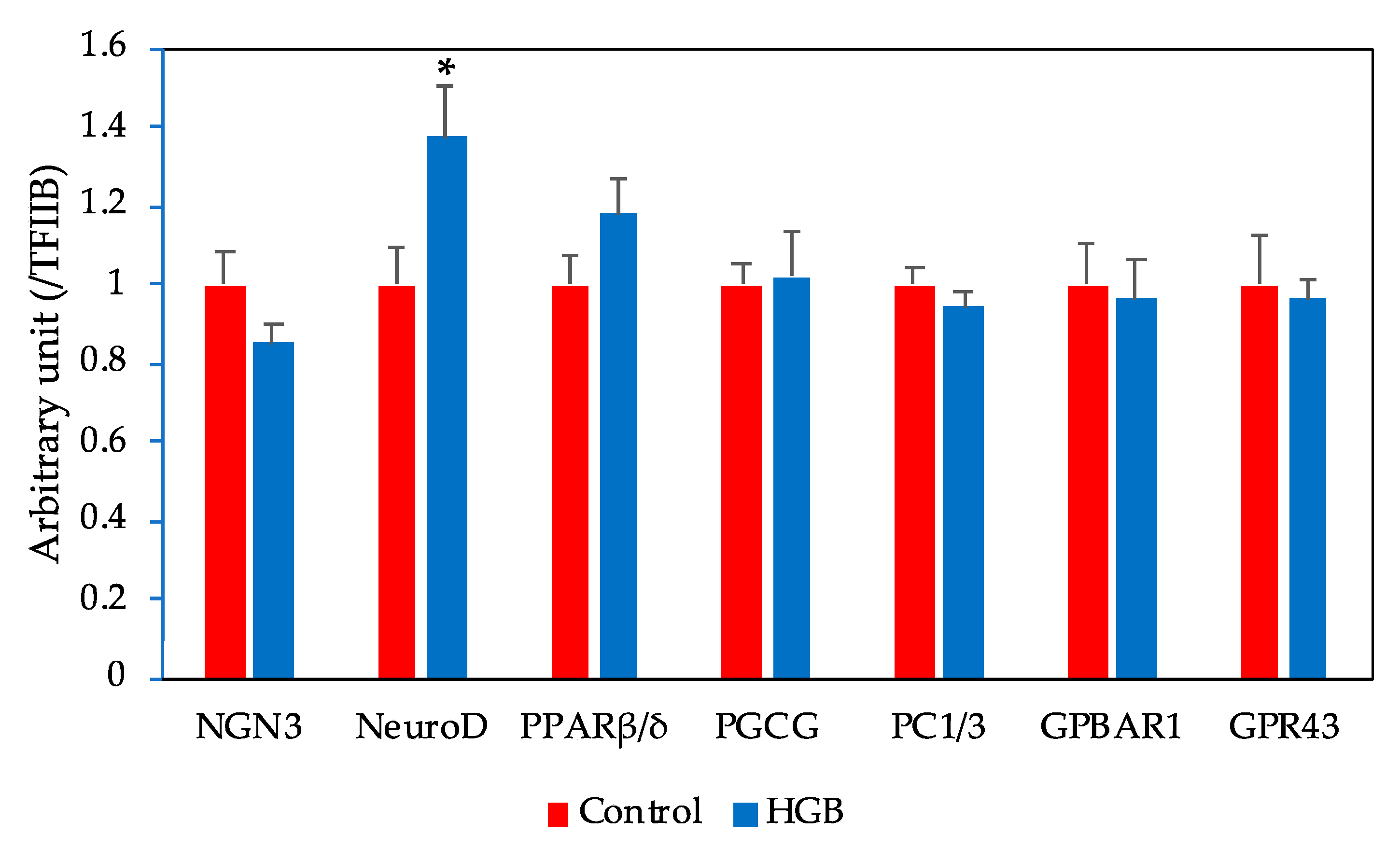
3.6. mRNA Expression Levels in the Ileum Determined by q-PCR
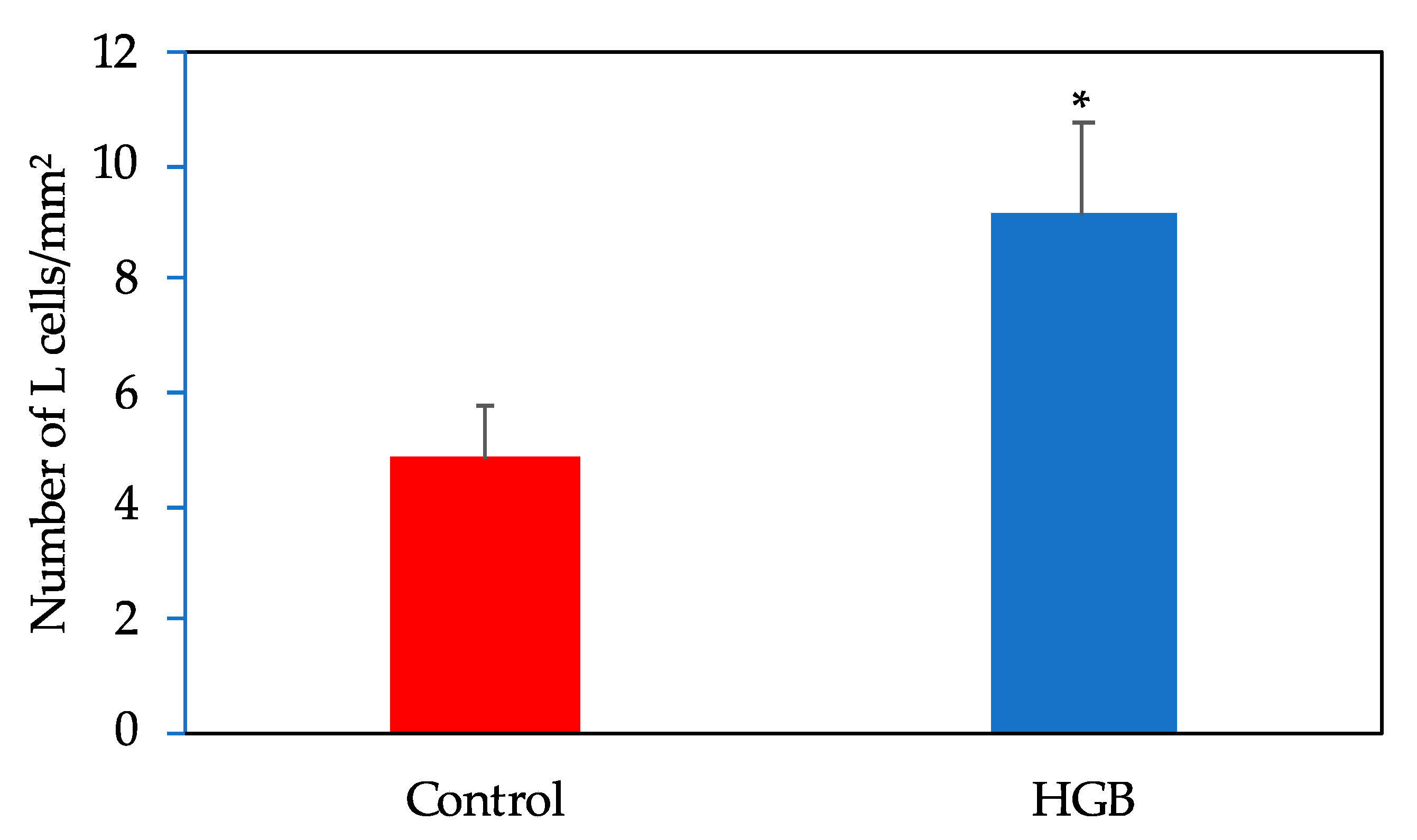
3.7. Number of L Cells in the Ileum
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Behall, K.M.; Scholfield, D.J.; Hallfrisch, J. Comparison of hormone and glucose responses of overweight women to barley and oats. J. Am. Coll. Nutr. 2005, 24, 182–188. [Google Scholar] [CrossRef]
- Casiraghi, M.C.; Garsetti, M.; Testolin, G.; Brighenti, F. Post-prandial responses to cereal products enriched with barley β-glucan. J. Am. Coll. Nutr. 2006, 25, 313–320. [Google Scholar] [CrossRef]
- Aldughpassi, A.; Abdel-Aal, E.-S.M.; Wolever, T.M.S. Barley cultivar, kernel composition, and processing affect the glycemic index. J. Nutr. 2012, 142, 1666–1671. [Google Scholar] [CrossRef]
- AbuMweis, S.S.; Jew, S.; Ames, N.P. Beta-glucan from barley and its lipid-lowering capacity, a meta-analysis of randomized, controlled trials. Eur. J. Clin. Nutr. 2010, 64, 1472–1480. [Google Scholar] [CrossRef]
- Behall, K.M.; Scholfield, D.J.; Hallfrisch, J. Diets containing barley significantly reduce lipids in mildly hypercholesterolemic men and women. Am. J. Clin. Nutr. 2004, 80, 1185–1193. [Google Scholar] [CrossRef]
- Wang, Y.; Harding, S.V.; Eck, P.; Thandapilly, S.J.; Gamel, T.H.; Abdel-Aal, E.-S.M.; Crow, G.H.; Tosh, S.M.; Jones, P.J.; Ames, N.P. High-Molecular-Weight beta-Glucan Decreases Serum Cholesterol Differentially Based on the CYP7A1 rs3808607 Polymorphism in Mildly Hypercholesterolemic Adults. J. Nutr. 2016, 146, 720–727. [Google Scholar] [CrossRef] [PubMed]
- Hughes, S.A.; Shewry, P.R.; Gibson, G.R.; McCleary, B.V.; Rastall, R.A. In vitro fermentation of oat and barley derived beta-glucans by human faecal microbiota. FEMS Microbiol. Ecol. 2008, 64, 482–493. [Google Scholar] [CrossRef] [PubMed]
- Bindelle, J.; Pieper, R.; Montoya, C.A.; Van Kessel, A.G.; Leterme, P. Nonstarch polysaccharide-degrading enzymes alter the microbial community and the fermentation patterns of barley cultivars and wheat products in an in vitro model of the porcine gastrointestinal tract. FEMS Microbiol. Ecol. 2011, 76, 553–563. [Google Scholar] [CrossRef] [PubMed]
- Aoe, S.; Yamanaka, C.; Fuwa, M.; Tamiya, T.; Nakayama, Y.; Miyoshi, T.; Kitazono, E. Effects of BARLEYmax and high-beta-glucan barley line on short-chain fatty acids production and microbiota from the cecum to the distal colon in rats. PLoS ONE 2019, 14, e0218118. [Google Scholar] [CrossRef] [PubMed]
- Kanamoto, I.; Koyama, S.; Murata, I.; Inoue, Y.; Kohyama, N.; Ichinose, Y.; Komae, K.; Yauagisaya, T.; Aoe, S. Effect of ingestion of high β-glucan barley breads with different particle size on postprandial blood glucose levels. Luminacoids Res. 2017, 21, 19–23. [Google Scholar]
- Aoe, S.; Komae, K.; Inoue, Y.; Murata, I.; Minegishi, Y.; Kanmotom, I.; Kohyama, N.; Ichinose, Y.; Yoshioka, T.; Yanagisawa, T. Effects of Various Blending Ratios of Rice and Waxy Barley on Postprandial Blood Glucose Levels. J. Jpn. Soc. Nutr. Food Sci. 2018, 71, 283–288. [Google Scholar] [CrossRef]
- Shimizu, C.; Kihara, M.; Aoe, S.; Araki, S.; Ito, K.; Hayashi, K.; Watari, J.; Sakata, Y.; Ikegami, S. Effect of high beta-glucan barley on serum cholesterol concentrations and visceral fat area in Japanese men—A randomized, double-blinded, placebo-controlled trial. Plant Foods Hum. Nutr. 2008, 63, 21–25. [Google Scholar] [CrossRef] [PubMed]
- Aoe, S.; Ichinose, Y.; Kohyama, N.; Komae, K.; Takahashi, A.; Abe, D.; Yoshioka, T.; Yanagisawa, T. Effects of high beta-glucan barley on visceral fat obesity in Japanese individuals: A randomized, double-blind study. Nutrition 2017, 42, 1–6. [Google Scholar] [CrossRef]
- Aoe, S.; Ikenaga, T.; Noguchi, H.; Kohashi, C.; Kakumoto, K.; Kohda, N. Effect of cooked white rice with high β-glucan barley on appetite and energy intake in healthy Japanese subjects: A randomized controlled trial. Plant Foods Hum. Nutr. 2014, 69, 325–330. [Google Scholar] [CrossRef]
- Johansson, E.V.; Nilsson, A.C.; Östman, E.M.; Björck, I.M.E. Effects of indigestible carbohydrates in barley on glucose metabolism, appetite and voluntary food intake over 16 h in healthy adults. Nutr. J. 2013, 12, 46. [Google Scholar] [CrossRef]
- Brighenti, F.; Benini, L.; Del Rio, D.; Casiraghi, C.; Pellegrini, N.; Scazzina, F.; Jenkins, D.J.; Vantini, I. Colonic fermentation of indigestible carbohydrates contributes to the second-meal effect. Am. J. Clin. Nutr. 2006, 83, 817–822. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, D.J.; Wolever, T.M.; Taylor, R.H.; Griffiths, C.; Krzeminska, K.; Lawrie, J.A.; Bennett, C.M.; Goff, D.V.; Sarson, D.L.; Bloom, S.R. Slow release dietary carbohydrate improves second meal tolerance. Am. J. Clin. Nutr. 1982, 35, 1339–1346. [Google Scholar] [CrossRef]
- Wolever, T.M.; Jenkins, D.J.; Ocana, A.M.; Rao, V.A.; Collier, G.R. Secondmeal effect: Low-glycemic-index foods eaten at dinner improve subsequent breakfast glycemic response. Am. J. Clin. Nutr. 1988, 48, 1041–1047. [Google Scholar] [CrossRef] [PubMed]
- Priyadarshini, M.; Villa, S.R.; Fuller, M.; Wicksteed, B.; Mackay, C.R.; Alquier, T.; Poitout, V.; Mancebo, H.; Mirmira, R.G.; Gilchrist, A.; et al. An Acetate-Specific GPCR, FFAR2, Regulates Insulin Secretion. Mol. Endocrinol. 2015, 29, 1055–1066. [Google Scholar] [CrossRef]
- Holst, J.J. Glucagon like peptide 1: A newly discovered gastrointestinal hormone. Gastroenterology 1994, 107, 1848–1855. [Google Scholar] [CrossRef]
- Orskov, C.; Rabenhoj, L.; Wettergren, A.; Kofod, H.; Holst, J.J. Tissue and plasma concentrations of amidated and glycine-extended glucagonlike peptide I in humans. Diabetes 1994, 43, 535–539. [Google Scholar] [CrossRef] [PubMed]
- Reimer, R.A.; McBurney, M.I. Dietary fiber modulates intestinal proglucagon messenger ribonucleic acid and postprandial secretion of glucagon-like peptide-1 and insulin in rats. Endocrinology 1996, 137, 3948–3956. [Google Scholar] [CrossRef]
- Reimer, R.A.; Thomson, A.B.; Rajotte, R.V.; Basu, T.K.; Ooraikul, B.; McBurney, M.I. A physiological level of rhubarb fiber increases proglucagon gene expression and modulates intestinal glucose uptake in rats. J. Nutr. 1997, 127, 1923–1928. [Google Scholar] [CrossRef][Green Version]
- Reimer, R.A.; Thomson, A.B.R.; Rajotte, R.V.; Basu, T.K.; Ooraikul, B.; McBurney, M.I. Proglucagon messenger ribonucleic acid and intestinal glucose uptake are modulated by fermentable fiber and food intake in diabetic rats. Nutr. Res. 2000, 20, 851–864. [Google Scholar] [CrossRef]
- Mio, K.; Yamanaka, C.; Matsuoka, T.; Kobayashi, T.; Aoe, S. Effects of beta-glucan rich barley flour on glucose and lipid metabolism in the ileum, liver, and adipose tissues of high-fat diet induced-obesity model male mice analyzed by DNA microarray. Nutrients 2020, 12, 3546. [Google Scholar] [CrossRef] [PubMed]
- Miyamoto, J.; Watanabe, K.; Taira, S.; Kasubuchi, M.; Li, X.; Irie, J.; Itoh, H.; Kimura, I. Barley beta-glucan improves metabolic condition via short-chain fatty acids produced by gut microbial fermentation in high fat diet fed mice. PLoS ONE 2018, 13, e0196579. [Google Scholar] [CrossRef]
- Belobrajdic, D.P.; Jobling, S.A.; Morell, M.K.; Taketa, S.; Bird, A.R. Wholegrain barley beta-glucan fermentation does not improve glucose tolerance in rats fed a high-fat diet. Nutr. Res. 2015, 35, 162–168. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.C.; Rodriguez, F.; Storey, M.; Farmakalidis, E.; Prosky, L. Determination of soluble and insoluble dietary fiber in psyllium containing cereal products. J. AOAC Int. 1995, 78, 724–729. [Google Scholar] [CrossRef]
- McCleary, B.V.; Codd, R. Measurement of (1–3), (1–4)-β-D-glucan in barley and oats: A streamlined enzymic procedure. J. Sci. Food Agric. 1991, 55, 303–312. [Google Scholar] [CrossRef]
- Atarashi, K.; Tanoue, T.; Oshima, K.; Suda, W.; Nagano, Y.; Nishikawa, H.; Fukuda, S.; Saito, T.; Narushima, T.; Hase, K.; et al. Treg induction by a rationally selected mixture of Clostridia strains from the human microbiota. Nature 2013, 500, 232–236. [Google Scholar] [CrossRef]
- Knapper, J.M.; Morgan, L.M.; Fletcher, J.M.; Marks, V. Plasma and intestinal concentrations of GIP and GLP-1 (7-36) amide during suckling and after weaning in pigs. Horm. Metab. Res. 1995, 27, 485–490. [Google Scholar] [CrossRef] [PubMed]
- Thondre, P.S.; Shafat, A.; Clegg, M.E. Molecular weight of barley beta-glucan influences energy expenditure, gastric emptying and glycaemic response in human subjects. Br. J. Nutr. 2013, 110, 2173–2179. [Google Scholar] [CrossRef]
- El Khoury, D.; Cuda, C.; Luhovy, B.L.; Anderson, G.H. Beta glucan: Health benefits in obesity and metabolic syndrome. J. Nutr. Metab. 2012, 2012, 851362. [Google Scholar] [CrossRef]
- Mio, K.; Yamanaka, C.; Ichinose, Y.; Kohyama, N.; Yanagisawa, T.; Aoe, S. Effects of barley β-glucan with various molecular weights partially hydrolyzed by endogenous β-glucanase on glucose tolerance and lipid metabolism in mice. Cereal Chem. 2020, 97, 1056–1065. [Google Scholar] [CrossRef]
- Nilsson, A.C.; Ostman, E.M.; Granfeldt, Y.; Björck, I.M. Effect of cereal test breakfasts differing in glycemic index and content of indigestible carbohydrates on daylong glucose tolerance in healthy subjects. Am. J. Clin. Nutr. 2008, 87, 645–654. [Google Scholar] [CrossRef]
- Matsuoka, T.; Tsuchida, A.; Yamaji, A.; Kurosawa, C.; Shinohara, M.; Takayama, I.; Nakagomi, H.; Izumi, K.; Ichikawa, Y.; Hariya, N.; et al. Consumption of a meal containing refined barley flour bread is associated with a lower postprandial blood glucose concentration after a second meal compared with one containing refined wheat flour bread in healthy Japanese, A randomized control trial. Nutrition 2020, 72, 110637. [Google Scholar] [CrossRef] [PubMed]
- Burcelin, R.; Cani, P.D.; Knauf, C. Glucagon-like peptide-1 and energy homeostasis. J. Nutr. 2007, 137, 2534S–2538S. [Google Scholar] [CrossRef]
- Cani, P.D.; Holst, J.J.; Drucker, D.J.; Delzenne, N.M.; Thorens, B.; Burcelin, R.; Knauf, C. GLUT2 and the incretin receptors are involved in glucoseinduced incretin secretion. Mol. Cell. Endocrinol. 2007, 276, 18–23. [Google Scholar] [CrossRef] [PubMed]
- Knauf, C.; Cani, P.D.; Ait-Belgnaoui, A.; Benani, A.; Dray, C.; Cabou, C.; Colom, A.; Uldry, M.; Rastrelli, S.; Sabatier, E.; et al. Brain glucagon-like peptide 1 signaling controls the onset of high-fat diet-induced insulin resistance and reduces energy expenditure. Endocrinology 2008, 149, 4768–4777. [Google Scholar] [CrossRef] [PubMed]
- Knauf, C.; Cani, P.D.; Kim, D.H.; Iglesias, M.A.; Chabo, C.; Waget, A.; Colom, A.; Rastrelli, S.; Delzenne, N.M.; Drucker, D.J.; et al. Role of central nervous system glucagon-like Peptide-1 receptors in enteric glucose sensing. Diabetes 2008, 57, 2603–2612. [Google Scholar] [CrossRef]
- Kok, N.N.; Morgan, L.M.; Williams, C.M.; Roberfroid, M.B.; Thissen, J.P.; Delzenne, N.M. Insulin, glucagon-like peptide 1, glucose-dependent insulinotropic polypeptide and insulin-like growth factor I as putative mediators of the hypolipidemic effect of oligofructose in rats. J. Nutr. 1998, 128, 1099–1103. [Google Scholar] [CrossRef]
- Keenan, M.J.; Zhou, J.; McCutcheon, K.L.; Raggio, A.M.; Bateman, H.G.; Todd, E.; Jones, C.K.; Tulley, R.T.; Melton, S.; Martin, R.J.; et al. Effects of resistant starch, a non-digestible fermentable fiber, on reducing body fat. Obesity 2006, 14, 1523–1534. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Hegsted, M.; McCutcheon, K.L.; Keenan, M.J.; Xi, X.; Raggio, A.M.; Martin, R.J. Peptide YY and proglucagon mRNA expression patterns and regulation in the gut. Obesity 2006, 14, 683–689. [Google Scholar] [CrossRef] [PubMed]
- Goodlad, R.A.; Lenton, W.; Ghatei, M.A.; Adrian, T.E.; Bloom, S.R.; Wright, N.A. Effects of an elemental diet, inert bulk and different types of dietary fibre on the response of the intestinal epithelium to refeeding in the rat and relationship to plasma gastrin, enteroglucagon, and PYY concentrations. Gut 1987, 28, 171–180. [Google Scholar] [CrossRef] [PubMed]
- Goodlad, R.A.; Lenton, W.; Ghatei, M.A.; Adrian, T.E.; Bloom, S.R.; Wright, N.A. Proliferative effects of ‘fibre’ on the intestinal epithelium epithelium: Relationship to gastrin, enteroglucagon and PYY. Gut 1987, 28, 221–226. [Google Scholar] [CrossRef]
- Roberfroid, M.B.; Delzenne, N.M. Dietary fructans. Annu. Rev. Nutr. 1998, 18, 117–143. [Google Scholar] [CrossRef] [PubMed]
- Karaki, S.; Mitsui, R.; Hayashi, H.; Kato, I.; Sugiya, H.; Iwanaga, T.; Furness, J.B.; Kuwahara, A. Short-chain fatty acid receptor, GPR43, is expressed by enteroendocrine cells and mucosal mast cells in rat intestine. Cell Tissue Res. 2006, 324, 353–360. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| (g/100 g) | |
|---|---|
| Moisture | 9.1 |
| Fat | 1.8 |
| Protein | 12.8 |
| Ash | 0.7 |
| Available carbohydrate | 65.7 |
| Total dietary fiber | 9.9 |
| β-glucan | 5.4 |
| (g/kg Diet) | ||
|---|---|---|
| Control | HGB | |
| Casein | 200 | 120.6 |
| L-cystine | 3 | 3 |
| Corn starch | 197.486 | - |
| Dextrinized corn starch | 132 | - |
| Sucrose | 100 | 62.886 |
| Soybean oil | 70 | 70 |
| Lard | 200 | 190.9 |
| Cellulose | 50 | - |
| HGB flour | - | 505.1 * |
| AIN-93G mineral mixture | 35 | 35 |
| AIN-93 vitamin mixture | 10 | 10 |
| Choline bitartrate | 2.5 | 2.5 |
| t-Butylhydroquinone | 0.014 | 0.014 |
| Control | HGB | |
|---|---|---|
| Initial weight (g) | 18.5 ± 0.2 | 18.5 ± 0.2 |
| Final weight (g) | 41.2 ± 0.7 | 42.4 ± 0.9 |
| Body weight gain (g/d) | 0.27 ± 0.01 | 0.30 ± 0.01 |
| Food intake (g/d) | 2.8 ± 0.0 | 3.0 ± 0.1 |
| Food efficiency ratio (%) | 9.73 ± 0.41 | 9.92 ± 1.06 |
| Control | HGB | |
|---|---|---|
| Liver (g) | 1.47 ± 0.06 | 1.56 ± 0.09 |
| Cecum with digesta (g) | 0.28 ± 0.02 | 0.38 ± 0.02 * |
| Total abdominal fat | 4.53 ± 0.20 | 4.38 ± 0.16 |
| Retroperitoneal fat (g) | 0.95 ± 0.05 | 0.91 ± 0.02 |
| Epididymal fat (g) | 2.61 ± 0.11 | 2.48 ± 0.01 |
| Mesenteric fat (g) | 0.97 ± 0.10 | 0.99 ± 0.10 |
| Control | HGB | |
|---|---|---|
| Cholesterol (mmol/L) | 5.60 ± 0.34 | 5.96 ± 0.55 |
| Triglyceride (mmol/L) | 0.84 ± 0.11 | 0.67 ± 0.04 |
| NEFA (μmol/L) | 639.8 ± 45.5 | 728.0 ± 33.0 |
| Glucose (mmol/L) | 16.61 ± 0.86 | 14.88 ± 0.55 |
| Insulin (ng/mL) | 6.10 ± 1.03 | 8.36 ± 1.22 |
| Leptin (ng/mL) | 71.35 ± 7.22 | 73.48 ± 6.14 |
| Control | HGB | |
|---|---|---|
| Portal vein active GLP-1 (7-36) levels (pg/mL) | 50.0 ± 9.5 | 81.4 ± 10.0 * |
| Cecal total GLP-1 pool size (ng/cecum) | 31.1 ± 7.4 | 64.5 ± 8.4 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Suzuki, S.; Aoe, S. High β-Glucan Barley Supplementation Improves Glucose Tolerance by Increasing GLP-1 Secretion in Diet-Induced Obesity Mice. Nutrients 2021, 13, 527. https://doi.org/10.3390/nu13020527
Suzuki S, Aoe S. High β-Glucan Barley Supplementation Improves Glucose Tolerance by Increasing GLP-1 Secretion in Diet-Induced Obesity Mice. Nutrients. 2021; 13(2):527. https://doi.org/10.3390/nu13020527
Chicago/Turabian StyleSuzuki, Sachina, and Seiichiro Aoe. 2021. "High β-Glucan Barley Supplementation Improves Glucose Tolerance by Increasing GLP-1 Secretion in Diet-Induced Obesity Mice" Nutrients 13, no. 2: 527. https://doi.org/10.3390/nu13020527
APA StyleSuzuki, S., & Aoe, S. (2021). High β-Glucan Barley Supplementation Improves Glucose Tolerance by Increasing GLP-1 Secretion in Diet-Induced Obesity Mice. Nutrients, 13(2), 527. https://doi.org/10.3390/nu13020527