
Identification and Purification of Novel Low-Molecular-Weight Lupine Allergens as Components for Personalized Diagnostics

 , , , ,
, , , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Group
2.2. Lupine Extract Production and Protein Identification According to the Molecular Weight
2.2.1. Alkaline Extraction
2.2.2. Acidic Extraction
2.2.3. Gel Electrophoresis of Different Lupine Extracts
2.2.4. In Silico Analysis
2.2.5. 2D Fluorescence Difference Gel Electrophoresis (DIGE)
2.3. Identification and Purification of New Single Allergens
2.3.1. Recombinant Production of the Lupinus albus Profilin
2.3.2. Expression
2.3.3. Purification under Native Conditions
2.3.4. Preparative SDS-PAGE
2.3.5. Separation, Isolation and Purification of Proteins via Different Chromatography Methods
2.4. Identification, Isolation and Purification of Natural Lupine Lipid Transfer Proteins
2.5. Immunoblotting
2.6. Immunological Antigen Detection on Blotting Membranes
2.7. Sequence Alignment
2.8. N-Terminal Sequence Analysis
2.9. Mass Spectrometry
3. Results
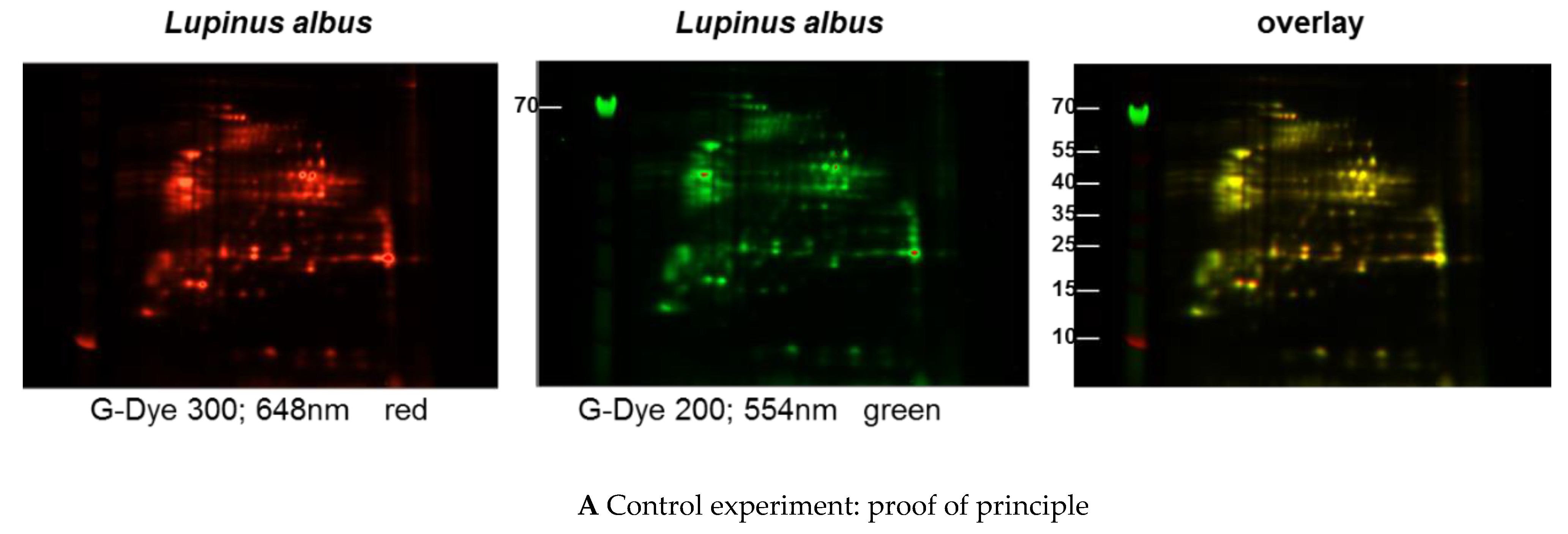
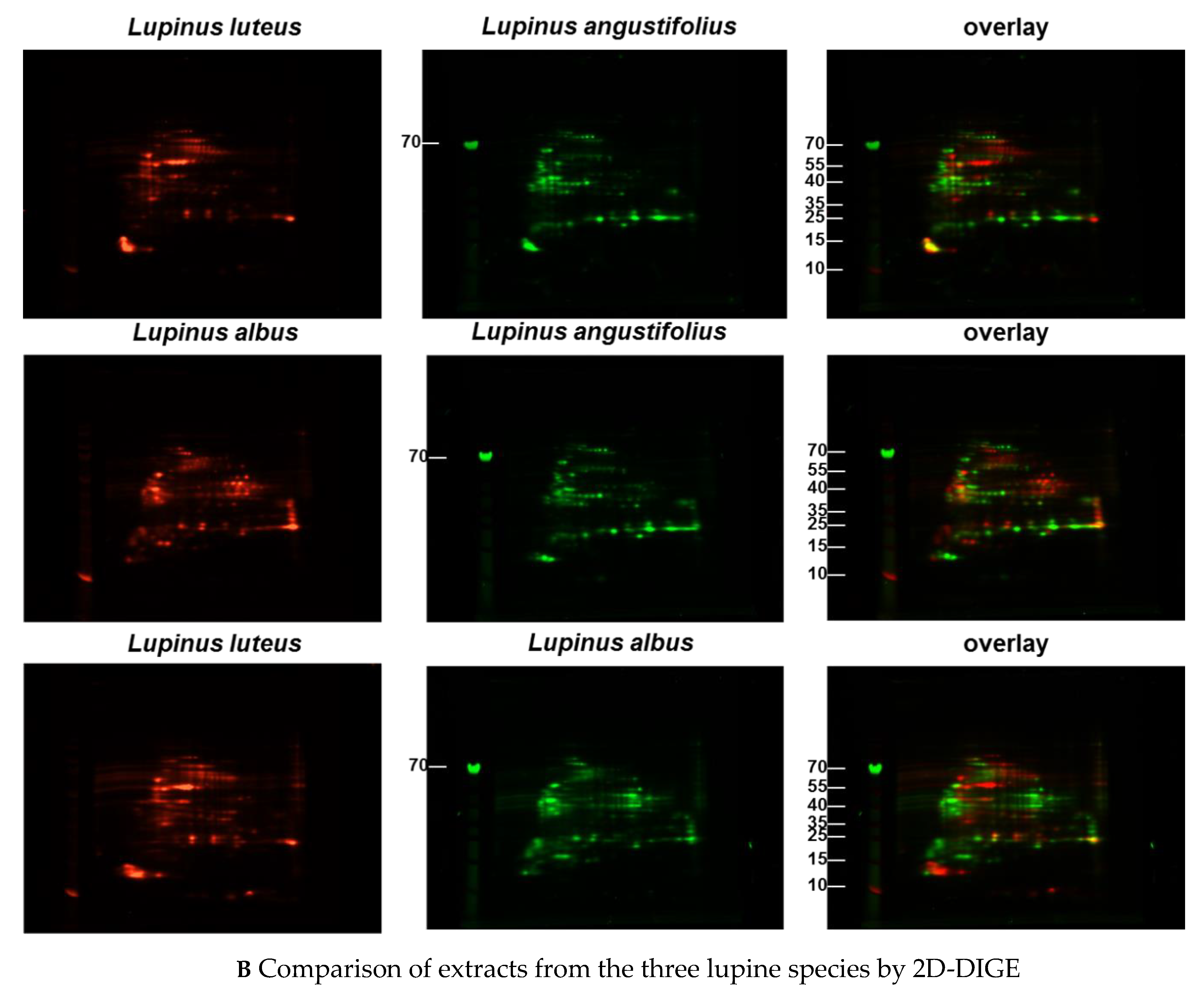
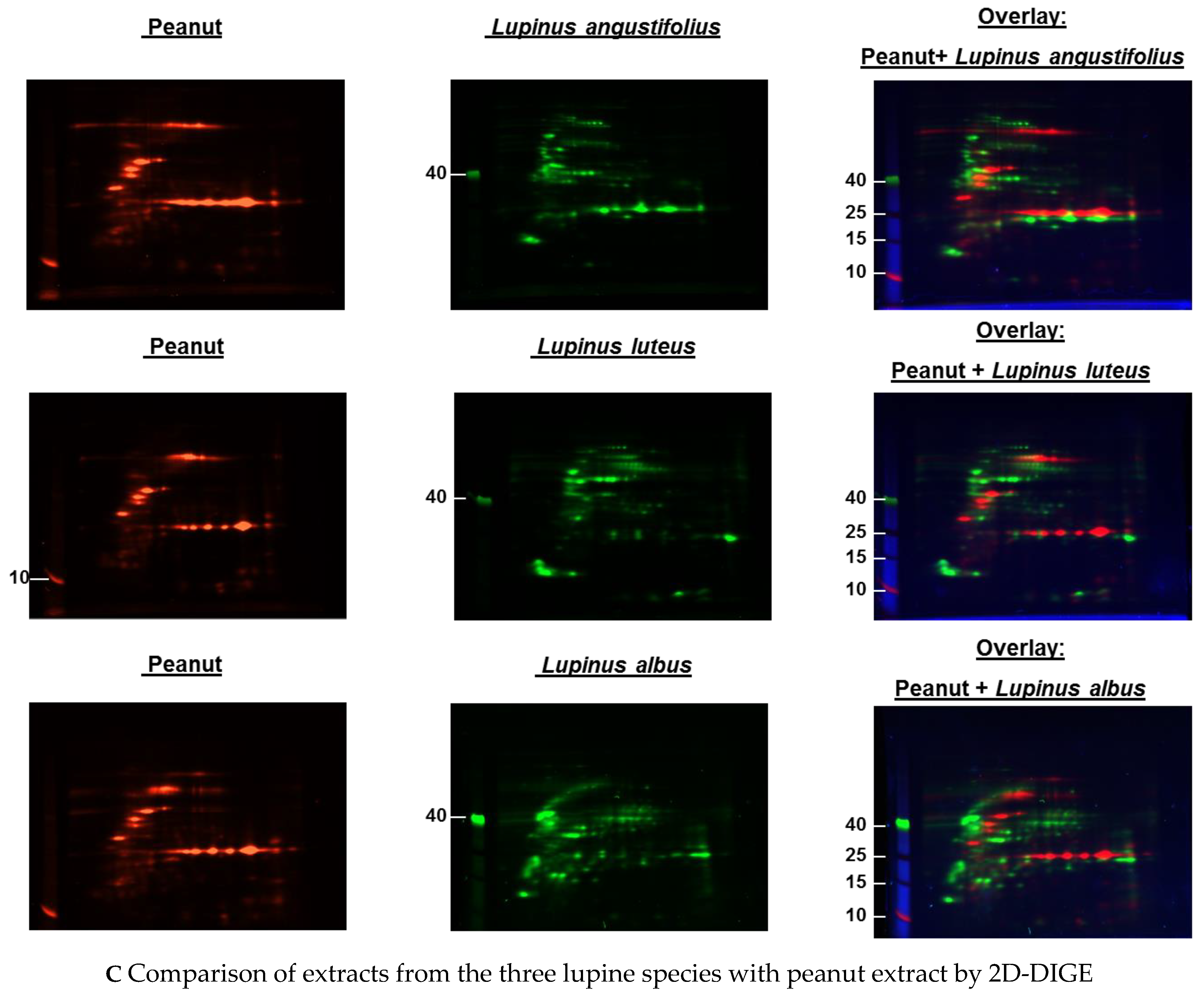
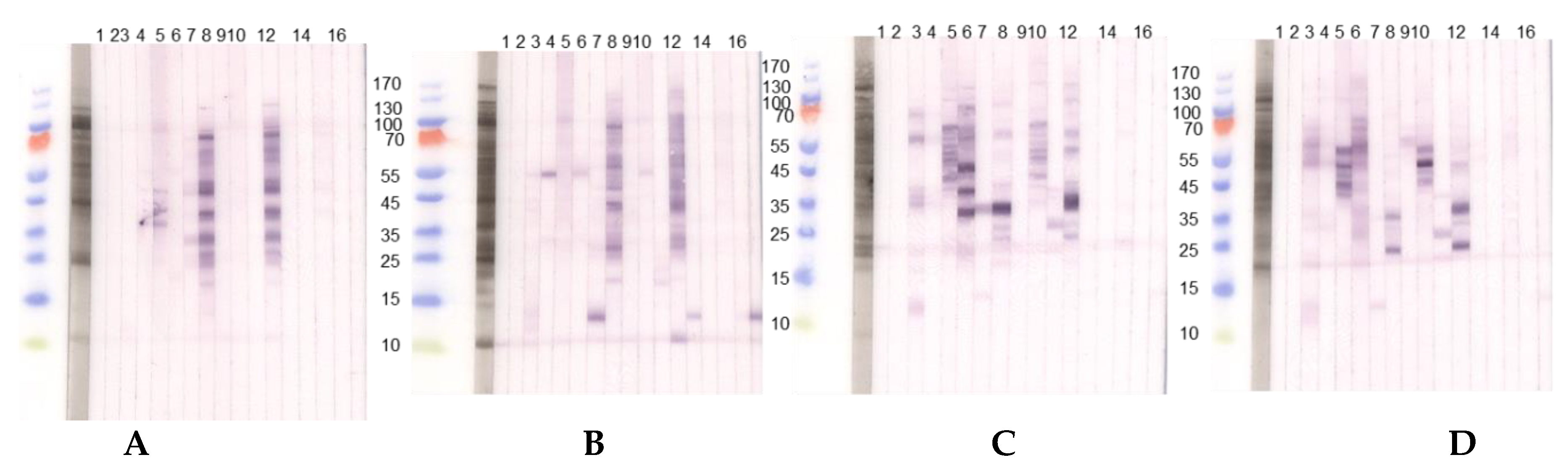
3.1. Comparison of Different Protein Extractions of Three Lupine Species
3.1.1. Immunoblot Analyses
Comparison of Different Protein Extractions of Three Lupine Species
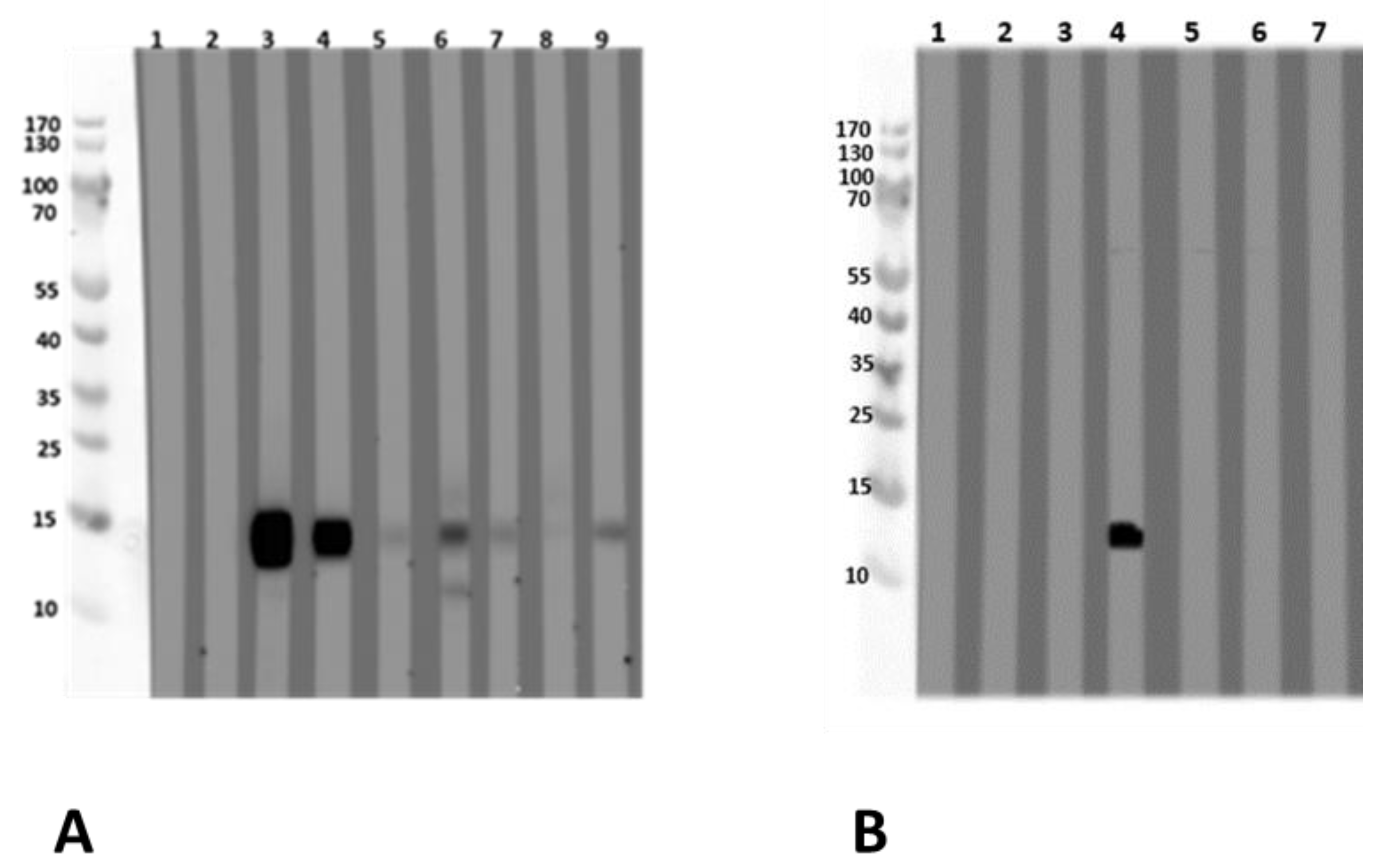
3.2. Identification of an L. albus Profilin
3.2.1. Allergen Identification via IgE-Reactivity with Sera from Lupine-Sensitized and Lupine-Allergic Patients
3.2.2. Sequence Alignment
3.2.3. Lupinus albus Profilin Allergenicity
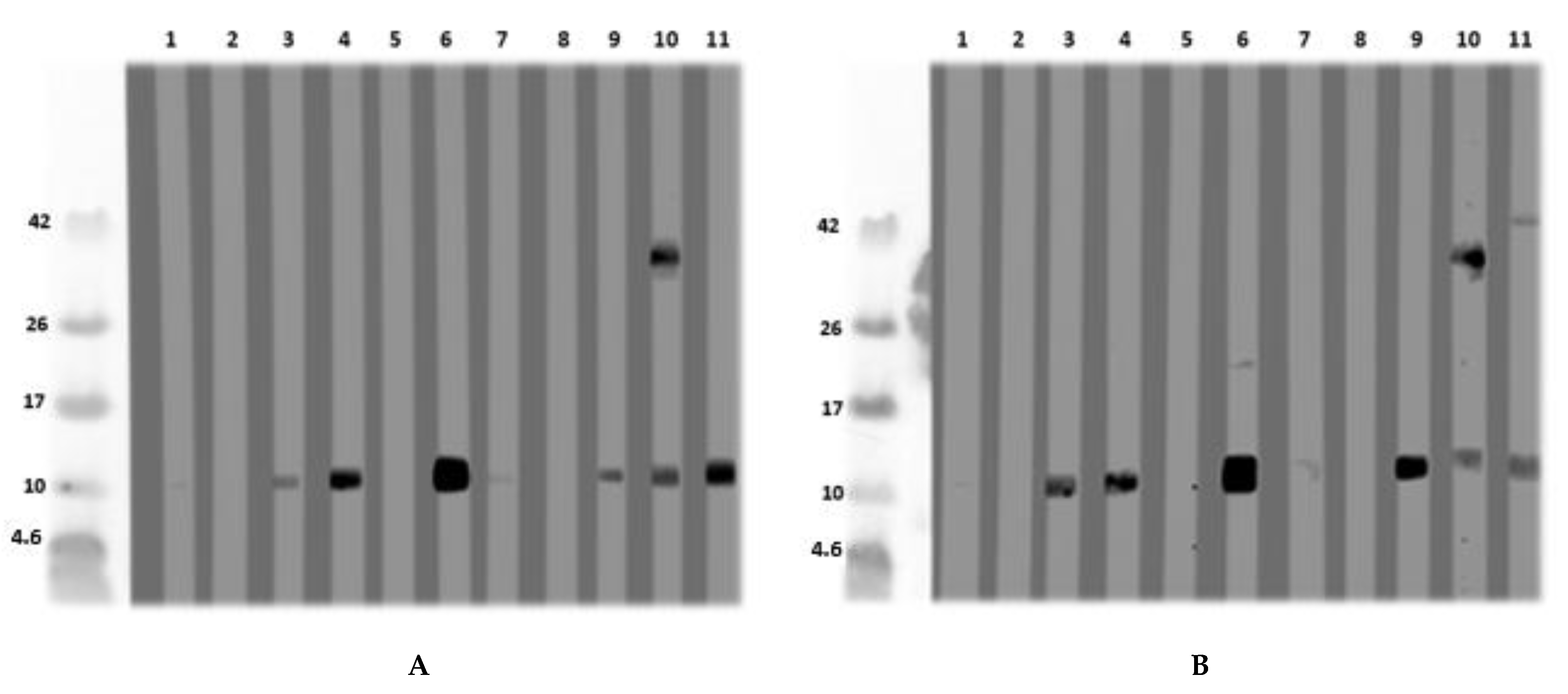
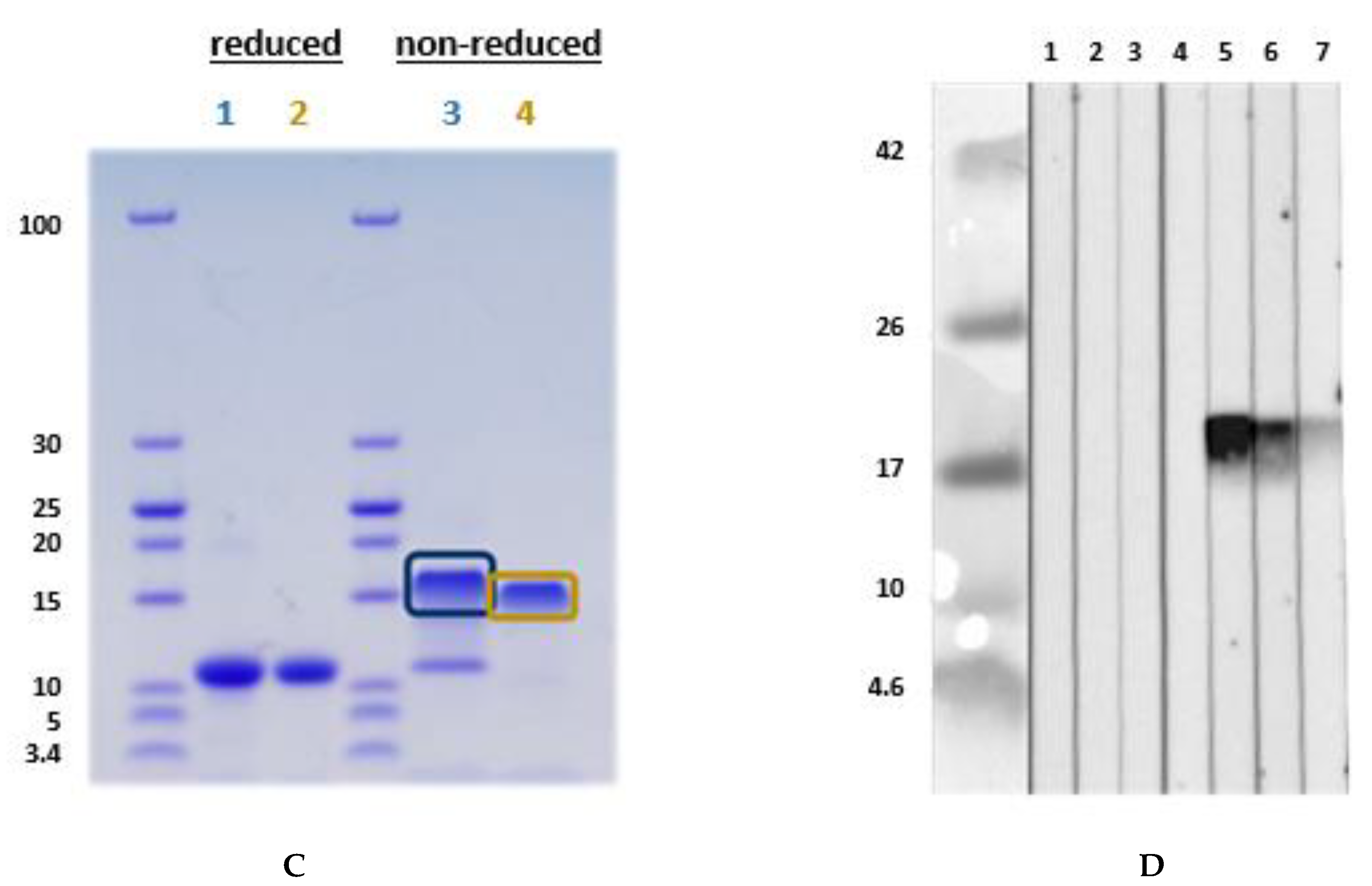
3.3. Isolation and Purification of a Non-Specific Lipid Transfer Protein from L. angustifolius Seeds
Lupinus angustifolius and L. luteus LTP Allergenicity
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Peeters, K.A.; Koppelman, S.J.; Penninks, A.H.; Lebens, A.; Bruijnzeel-Koomen, C.A.; Hefle, S.L.; Taylor, S.L.; van Hoffen, E.; Knulst, A.C. Clinical relevance of sensitization to lupine in peanut-sensitized adults. Allergy 2009, 64, 549–555. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.P.; Mori, T.A.; Sipsas, S.; Barden, A.; Puddey, I.B.; Burke, V.; Hall, R.S.; Hodgson, J.M. Lupin-enriched bread increases satiety and reduces energy intake acutely. Am. J. Clin. Nutr. 2006, 84, 975–980. [Google Scholar] [CrossRef] [PubMed]
- Jappe, U.; Vieths, S. Lupine, a source of new as well as hidden food allergens. Mol. Nutr. Food Res. 2010, 54, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Hefle, S.L.; Lemanske, R.F., Jr.; Bush, R.K. Adverse reaction to lupine-fortified pasta. J. Allergy Clin. Immunol. 1994, 94, 167–172. [Google Scholar] [CrossRef] [PubMed]
- Thermo Fisher Scientific. Lupinus albus, ImmunoCAP f335. Available online: https://www.thermofisher.com/diagnostic-education/hcp/de/de/resource-center/allergen-encyclopedia/whole-allergens.html?key=f335 (accessed on 1 December 2020).
- Cabanillas, B.; Jappe, U.; Novak, N. Allergy to peanut, soy bean and other legumes: Recent advances on allergen characterization, stability to processing, and IgE cross-reactivity. Mol. Nutr. Food Res. 2018, 62, 1–9. [Google Scholar] [CrossRef]
- Moneret-Vautrin, D.A.; Guerin, L.; Kanny, G.; Flabbee, J.; Frémont, S.; Morisset, M. Cross-allergenicity of peanut and lupine: The risk of lupine allergy in patients allergic to peanuts. J. Allergy Clin. Immunol. 1999, 104, 883–888. [Google Scholar] [CrossRef]
- Jappe, U.; Kull, S.; Opitz, A.; Zabel, P. Anaphylaxis to vanilla ice cream: A near fatal cross reactivity phenomenon. Letter to the Editor. J. Eur. Acad. Dermatol. Venereol. 2018, 32, e22–e23. [Google Scholar] [CrossRef]
- Campbell, C.P.; Jackson, A.S.; Johnson, A.R.; Thomas, P.S.; Yates, D.H. Occupational sensitzation to lupin in the workplace: Occupational asthma, rhinitis, and work-aggravated asthma. J. Allergy Clin. Immunol. 2007, 119, 1133–1139. [Google Scholar] [CrossRef]
- Goggin, D.E.; Mir, G.; Smith, W.; Stuckey, M.; Smith, P.M.C. Proteomic analysis of lupin seed proteins to identify conglutin ß as an allergen, Lup an 1. J. Agric. Food Chem. 2008, 56, 6370–6377. [Google Scholar] [CrossRef]
- Niggemann, B.; Beyer, K.; Erdmann, S.; Fuchs, T.; Kleine-Tebbe, J.; Lepp, U.; Raithel, M.; Reese, I.; Saloga, J.; Schäfer, C.; et al. Standardisierung von oralen Provokationstests bei Verdacht auf Nahrungsmittelallergie. Allergo J. 2011, 20, 149–160. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Thermo Fisher Scientific. User Guide: GeneJET Gel Extraktion Kit. Available online: https://tools.thermofisher.com/content/sfs/manuals/MAN0012661_GeneJET_Gel_Extraction_UG.pdf (accessed on 15 December 2016).
- Kyhse-Andersen, J. Electroblotting of multiple gels: A simple apparatus without buffer tank for rapid transfer of proteins from polyacrylamide to nitrocellulose. J. Biochem. Biophys. Methods 1984, 10, 203–209. [Google Scholar] [CrossRef]
- Petersen, A.; Kull, S.; Rennert, S.; Becker, W.M.; Krause, S.; Ernst, M.; Gutsmann, T.; Bauer, J.; Lindner, B.; Jappe, U. Peanut defensins: Novel allergens isolated from lipophilic peanut extract. J. Allergy Clin. Immunol. 2015, 136, 1295–1301. [Google Scholar] [CrossRef] [PubMed]
- Petersen, A.; Suck, R.; Lindner, B.; Georgieva, D.; Ernst, M.; Notbohm, H.; Becker, W.M. Phl p 3: Structural and immunological characterization of a major allergen of timothy grass pollen. Clin. Exp. Allergy 2006, 36, 840–849. [Google Scholar] [CrossRef] [PubMed]
- Shevchenko, A.; Wilm, M.; Vorm, O.; Mann, M. Mass spectrometric sequencing of proteins silver-stained polyacrylamide gels. Anal. Chem. 1996, 68, 850–858. [Google Scholar] [CrossRef] [PubMed]
- Available online: www.allergome.org (accessed on 20 January 2021).
- Available online: www.allergen.org (accessed on 20 January 2021).
- Jappe, U.; Nikolic, J.; Opitz, A.; Homann, A.; Zabel, P.; Gavrovic-Jankulovic, M. Apparent IgE-negative anaphylactic reaction to banana combined with kiwi-allergy-Complementary diagnostic value of purified single banana allergens. J. Eur. Acad. Dermatol. Venereol. 2016, 30, 1220–1222. [Google Scholar] [CrossRef]
- Jappe, U.; Schwager, C. Relevance of lipophilic allergens in food allergy diagnosis. Curr. Allergy Asthma Rep. 2017, 17, 61. [Google Scholar] [CrossRef]
- Alvarado, M.I.; Jimeno, L.; De La Torre, F.; Boissy, P.; Rivas, B.; Lázaro, M.J.; Barber, D. Profilin as a severe food allergen in allergic patients overexposed to grass pollen. Allergy 2014, 69, 1610–1616. [Google Scholar] [CrossRef]
- Guelsen, A.; Jappe, U. Lipid transfer protein sensitization in an apple-allergic patient: A case report from Northern Europe. Eur. Ann. Allergy Clin. Immunol. 2019, 51, 80–83. [Google Scholar] [CrossRef]
- Schocker, F.; Lüttkopf, D.; Scheurer, S.; Petersen, A.; Cisteró-Bahima, A.; Enrique, E.; San Miguel-Moncín, M.; Akkerdaas, J.; van Ree, R.; Vieths, S.; et al. Recombinant lipid transfer protein Cor a 8 from hazelnut: A new tool for in vitro diagnosis of potentially severe hazelnut allergy. J. Allergy Clin. Immunol. 2004, 113, 141–147. [Google Scholar] [CrossRef]
- Asero, R.; Mistrello, G.; Roncarolo, D.; Amato, S.; Caldironi, G.; Barocci, F.; van Ree, R. Immunological cross-reactivity between lipid transfer proteins from botanically unrelated plant-derived foods: A clinical study. Allergy 2002, 57, 900–906. [Google Scholar] [CrossRef] [PubMed]
- Pastorello, E.A.; Robino, A.M. Clinical role of lipid transfer proteins in food allergy. Mol. Nutr. Food Res. 2004, 48, 356–362. [Google Scholar] [CrossRef] [PubMed]
- Van Winkle, R.C.; Chang, C. The biochemical basis and clinical evidence of food allergy due to lipid transfer proteins: A comprehensive review. Clin. Rev. Allergy Immunol. 2014, 46, 211–224. [Google Scholar] [CrossRef] [PubMed]
- Scala, E.; Till, S.J.; Asero, R.; Abeni, D.; Guerra, E.C.; Pirrotta, L.; Paganelli, R.; Pomponi, D.; Giani, M.; De Pità, O.; et al. Lipid transfer protein sensitization: Reactivity profiles and clinical risk assessment in an Italian cohort. Allergy 2015, 70, 933–943. [Google Scholar] [CrossRef] [PubMed]
- Pastorello, E.A.; Farioli, L.; Pravettoni, V.; Scibilia, J.; Mascheri, A.; Borgonovo, L.; Piantanida, M.; Primavesi, L.; Stafylaraki, C.; Pasqualetti, S.; et al. Pru p 3-sensitised Italian peach-allergic patients are less likely to develop severe symptoms when also presenting IgE antibodies to Pru p 1 and Pru p 4. Int. Arch. Allergy Immunol. 2011, 156, 362–372. [Google Scholar] [CrossRef]
- Gomez, F.; Aranda, A.; Campo, P.; Diaz-Perales, A.; Blanca-Lopez, N.; Perkins, J.; Garrido, M.; Blanca, M.; Mayorga, C.; Torres, M.J. High prevalence of lipid transfer protein sensitization in apple allergic patients with systemic symptoms. PLoS ONE 2014, 9, e107304. [Google Scholar] [CrossRef]
- Pastorello, E.A.; Farioli, L.; Pravettoni, V.; Ortolani, C.; Ispano, M.; Monza, M.; Baroglio, C.; Scibola, E.; Ansaloni, R.; Incorvaia, C.; et al. The major allergen of peach (Prunus persica) is a lipid transfer protein. J. Allergy Clin. Immunol. 1999, 103 Pt 1, 520–526. [Google Scholar] [CrossRef]
- Commission Directive 2006/142/EC of 22 December 2006 amending Directive 2000/13/EC listing the ingredients which must under all circumstances appear on the labelling of foodstuffs. Off. J. Union 2006, L368, 110.








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| P-Code | Gender | Age | Peanut Allergy | Peanut Sensitization | Lupine Allergy | Lupine Sensitization | BLOT | CAP | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| L. luteus (Whole Extract) | L. angustifolius (Whole Extract) | L. luteus LTP | L. angustifolius LTP | L. albus Profilin | Total-IgE | Lupine Seed | Peanut Extract | Ara h 2 | Ara h 8 | Ara h 9 | |||||||
| P 1 | M | 35 | x | x | + | + | + | + | n. d. | >5000 | n. d. | 84.7 | n. d. | n. d. | n. d. | ||
| P 2 | F | 25 | x | x | (x) | x | + | + | − | − | n. d. | 110 | 4.75 | n. d. | n. d. | n. d. | n. d. |
| P 3 | F | 24 | x | x | (x) | x | + | + | (+) | (+) | n. d. | n. d. | 5.54 | n. d. | >100 | 0 | n. d. |
| P 4 | M | 76 | x | x | ? | x | + | + | + | + | n. d. | 1053 | n. d. | n. d. | n. d. | n. d. | n. d. |
| P 5 | F | 27 | x | x | ? | x | + | + | − | − | n. d. | 1828 | n. d. | 47.5 | 28.9 | 11.4 | 0.06 |
| P 6 | F | 63 | x | x | x | x | + | + | + | + | n. d. | 199 | 26.6 | 1.27 | n. d. | n. d. | 5.83 |
| P 7 | F | 24 | x | x | x | x | + | + | + | + | n. d. | n. d. | 6.5 | >100 | n. d. | n. d. | n. d. |
| P 8 | F | 42 | x | x | x | + | + | + | + | n. d. | >5000 | 4.9 | 18 | 0 | 50 | 0 | |
| P 9 | M | 15 | x | x | ? | x | + | + | + | + | n. d. | 705.5 | n. d. | 21.3 | 3.21 | 1.34 | 3.7 |
| P 10 | M | 63 | x | x | + | 0.75 | n. d. | n. d. | n. d. | n. d. | |||||||
| P 11 | F | 30 | x | x | ? | x | + | + | − | − | + | 1696 | n. d. | 80.7 | n. d. | >100 | n. d. |
| P 12 | F | 57 | x | x | x | + | − | − | − | + | 357 | 3.69 | 4.2 | <0.1 | 6.2 | 0 | |
| P 13 | F | 27 | x | x | x | + | − | − | − | + | 1324 | 0.71 | 13.7 | 8.56 | 4.7 | <0.01 | |
| P 14 | F | 50 | (x) | x | + | − | − | − | + | n. d. | 0.64 | n. d. | n. d. | n. d. | n. d. | ||
| P 15 | F | 38 | x | x | x | − | − | − | − | + | n. d. | <0.35 | 0.8 | n. d. | n. d. | n. d. | |
| P 16 | M | 24 | x | x | x | + | + | n. d. | n. d. | + | 208 | 1.28 | 39.8 | 36.4 | 0.67 | 0 | |
| P 17 | M | 28 | x | x | x | + | + | n. d. | n. d. | + | 94.3 | 0 | 0.74 | 0 | 0 | 0 | |
| P 18 | M | 24 | x | x | x | + | + | − | − | + | 1416 | 11 | 88.4 | 34.1 | 9.93 | 0.15 | |
| P 19 | M | 33 | x | x | x | + | + | n. d. | n. d. | n. d. | 595.6 | 5.55 | 75.9 | 51 | 7.95 | 0 | |
| P 21 | M | 10 | x | x | n. d. | n. d. | + | n. d. | n. d. | 531 | n. d. | 52.1 | 18.2 | 0.17 | 36.9 | ||
| P 22 | M | 43 | x | x | + | + | n. d. | n. d. | + | 83.5 | <0.01 | 0.22 | <0.01 | 1.65 | 0.30 | ||
| P 23 | F | 67 | x | x | x | + | + | − | − | − | 116 | 42.9 | 0 | n. d. | 0.46 | n. d. | |
| P 24 | M | 32 | x | x | x | + | + | n. d. | n. d. | − | n. d. | 7.02 | 37.1 | n. d. | n. d. | n. d. | |
| P 25 | F | 9 | x | x | x | x | + | (+) | n. d. | n. d. | − | n. d. | 11.2 | >100 | n. d. | n. d. | n. d. |
| P 26 | F | 39 | x | x | x | x | + | + | n. d. | n. d. | − | n. d. | 1.58 | n. d. | n. d. | n. d. | n. d. |
| P 27 | F | 34 | x | x | (x) | + | + | n. d. | n. d. | − | 137 | <0.12 | 2.76 | <0.1 | 0.29 | <0.01 | |
| P 28 | F | 32 | x | (x) | (+) | (+) | n. d. | n. d. | − | 319 | 16.7 | n. d. | 67.6 | 0 | n. d. | ||
| P 29 | F | 51 | x | − | − | n. d. | n. d. | n. d. | 67.3 | 0 | 0 | 0 | 0 | 0 | |||
| P 30 | F | 26 | x | x | x | x | + | + | n. d. | n. d. | − | 1023 | n.d. | >100 | >100 | n. d. | <0.01 |
| P 31 | M | 30 | x | x | − | − | n. d. | n. d. | − | 936 | n. d. | n. d. | 0.04 | n. d. | n. d. | ||
| Plant Food Allergens (Protein Families) | Peanut Allergen (Arachis hypogaea) Ara h x | MW [kDa] | Lupine Allergen (Lupinus angustifolius): Lup an x (Lupinus albus): Lup a x | MW [kDa] |
|---|---|---|---|---|
| Vicilin-type storage protein; 7S globulin | Ara h 1 (IUIS) | 64 | Lup an 1 β-conglutin (IUIS) Lup a 1 | 55–61 |
| Conglutin-like storage protein; 2S albumin | Ara h 2 (IUIS) | 17 | δ-conglutin Lup a δ-conglutin Lup an δ-conglutin | 2 4 |
| Legumin-type; 11S globulin | Ara h 3 (IUIS) | 60 | α-conglutin Lup a α-conglutin Lup an α-conglutin | 43 |
| Profilin | Ara h 5 (IUIS) | 15 | Lup a 5 (IUIS) | 15 |
| Conglutin; 2S albumin | Ara h 6 (IUIS) | 15 | n.a. | |
| Conglutin; 2S albumin | Ara h 7 (IUIS) | 15 | Lup a γ-conglutin? Lup an γ-conglutin? | n.a |
| PR-10 Bet v 1-super family | Ara h 8 (IUIS) | 17 | Lup a 4 Lup l 4 | 16.5 17 |
| Non-specific lipid transfer protein | Ara h 9 (IUIS) Ara h 16 (IUIS) Ara h 17 (IUIS) | 9.8 8.5 11 | Lup an 3 (IUIS) L. luteus | 11 11 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jappe, U.; Karstedt, A.; Warneke, D.; Hellmig, S.; Böttger, M.; Riffelmann, F.W.; Treudler, R.; Lange, L.; Abraham, S.; Dölle-Bierke, S.; et al. Identification and Purification of Novel Low-Molecular-Weight Lupine Allergens as Components for Personalized Diagnostics. Nutrients 2021, 13, 409. https://doi.org/10.3390/nu13020409
Jappe U, Karstedt A, Warneke D, Hellmig S, Böttger M, Riffelmann FW, Treudler R, Lange L, Abraham S, Dölle-Bierke S, et al. Identification and Purification of Novel Low-Molecular-Weight Lupine Allergens as Components for Personalized Diagnostics. Nutrients. 2021; 13(2):409. https://doi.org/10.3390/nu13020409
Chicago/Turabian StyleJappe, Uta, Arabella Karstedt, Daniela Warneke, Saskia Hellmig, Marisa Böttger, Friedrich W. Riffelmann, Regina Treudler, Lars Lange, Susanne Abraham, Sabine Dölle-Bierke, and et al. 2021. "Identification and Purification of Novel Low-Molecular-Weight Lupine Allergens as Components for Personalized Diagnostics" Nutrients 13, no. 2: 409. https://doi.org/10.3390/nu13020409
APA StyleJappe, U., Karstedt, A., Warneke, D., Hellmig, S., Böttger, M., Riffelmann, F. W., Treudler, R., Lange, L., Abraham, S., Dölle-Bierke, S., Worm, M., Wagner, N., Ruëff, F., Reese, G., Knulst, A. C., & Becker, W.-M. (2021). Identification and Purification of Novel Low-Molecular-Weight Lupine Allergens as Components for Personalized Diagnostics. Nutrients, 13(2), 409. https://doi.org/10.3390/nu13020409