
Butyrate Protects Porcine Colon Epithelium from Hypoxia-Induced Damage on a Functional Level


Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Tissue Sampling
2.2. Ussing Chamber Experiments
2.3. Buffer Solutions and Gassing
2.4. Two-Step RT-qPCR
2.5. Statistics
3. Results
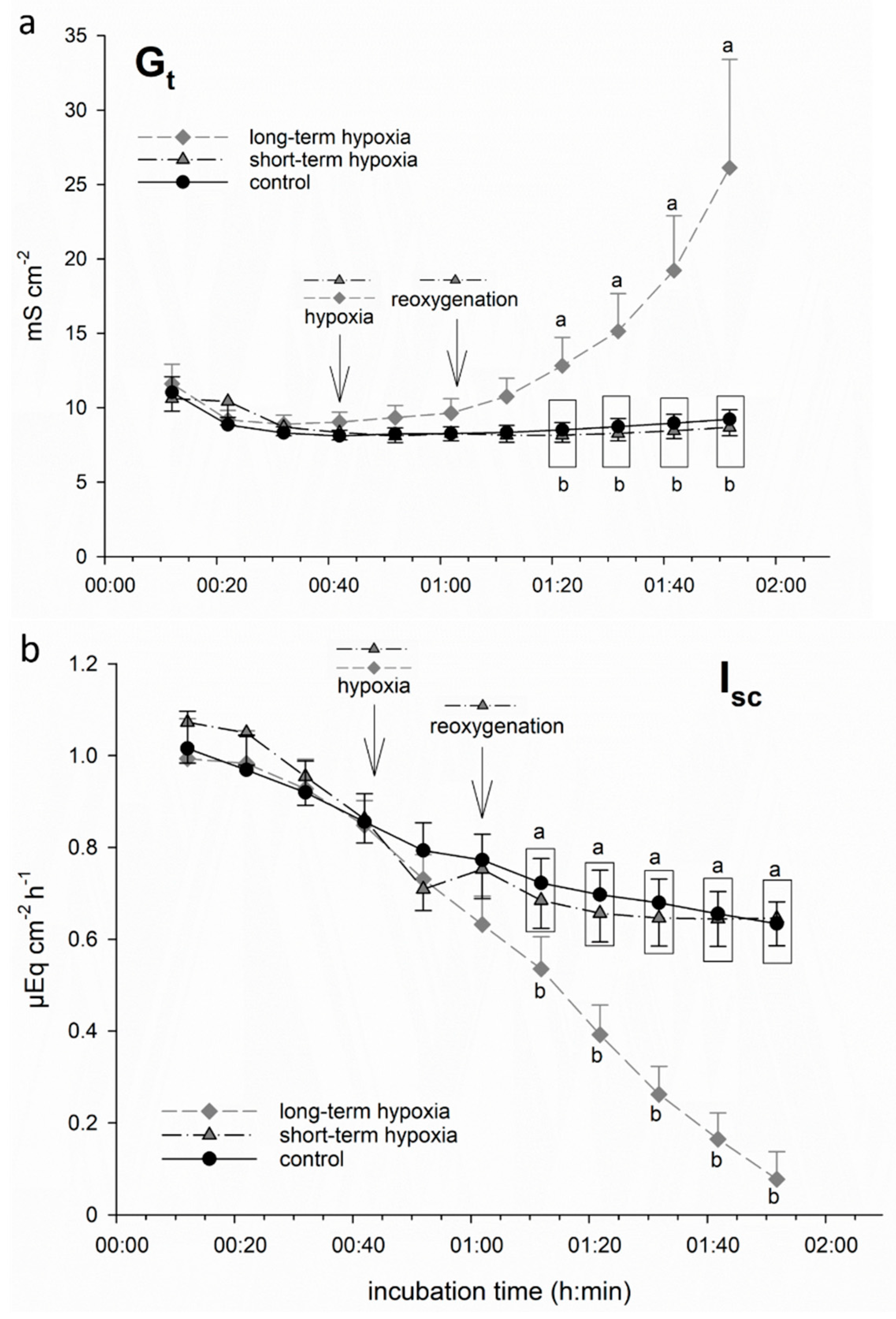
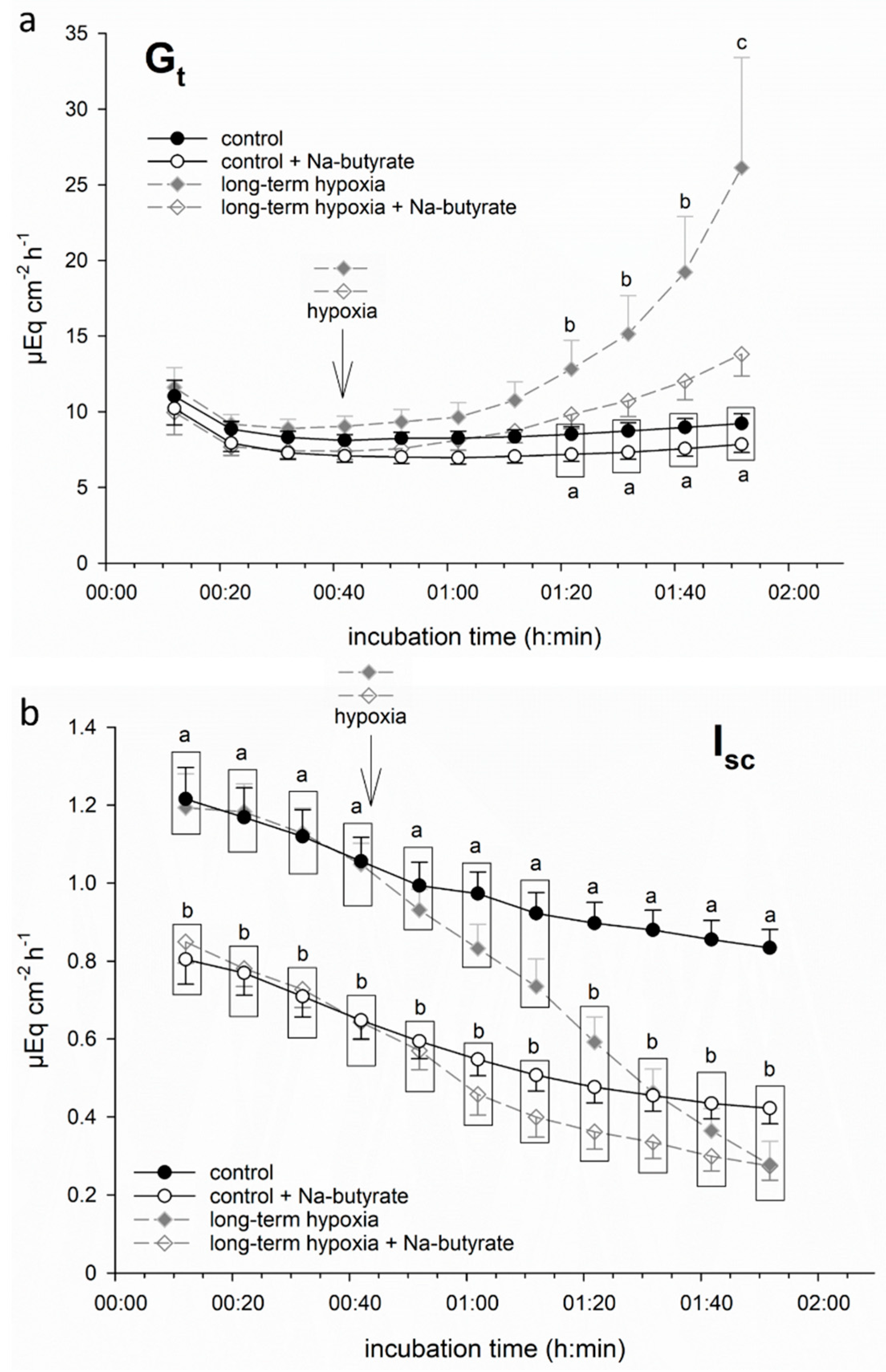
3.1. Hypoxia-Induced Damage Is Ameliorated by Butyrate Incubation
3.1.1. Long-Term but Not Short-Term Hypoxia Decreases Epithelial Integrity
3.1.2. Butyrate Incubation Ameliorates Hypoxia-Induced Changes
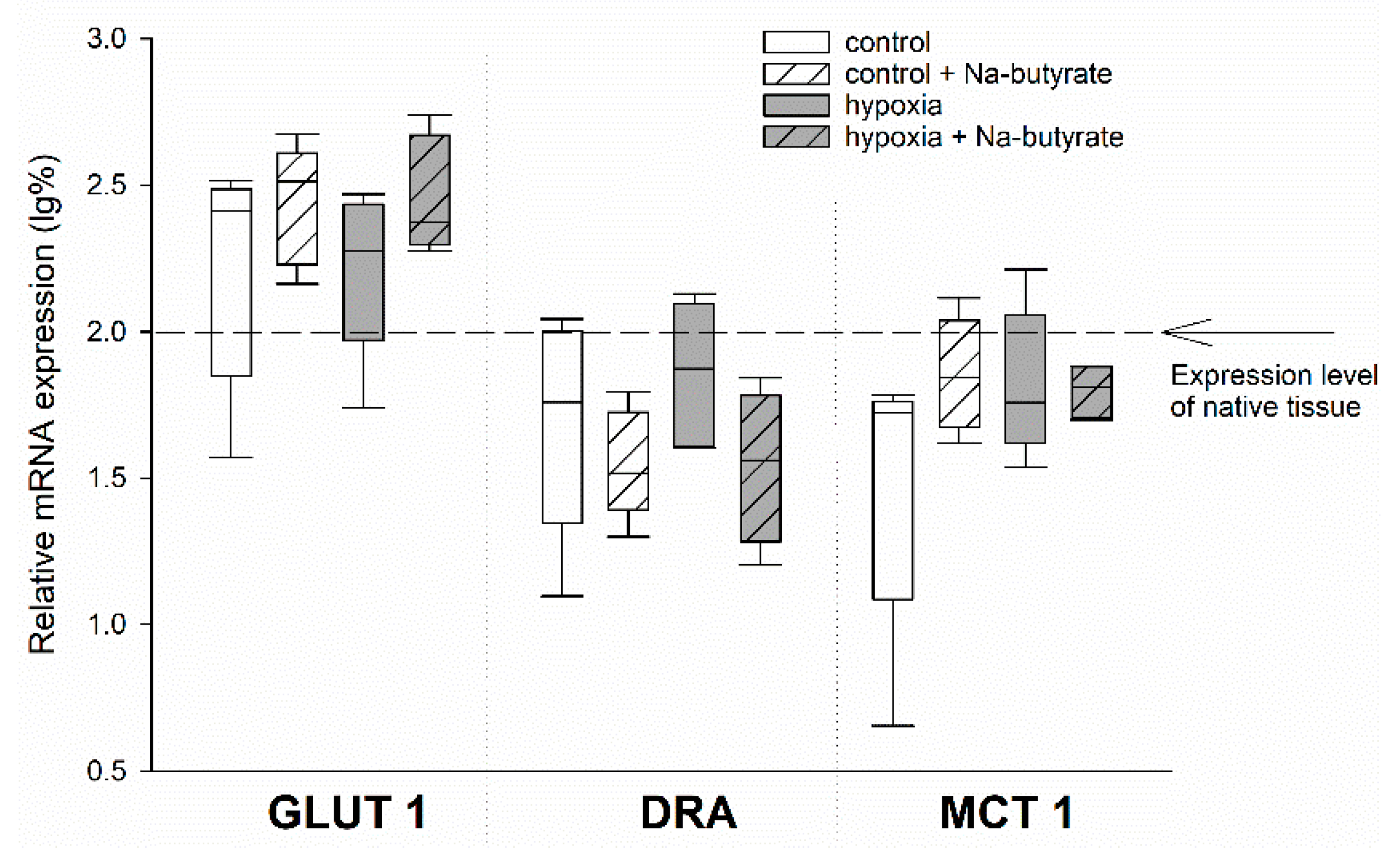
3.2. The Protective Effect of Butyrate Is Not Mediated on Gene Expression Level
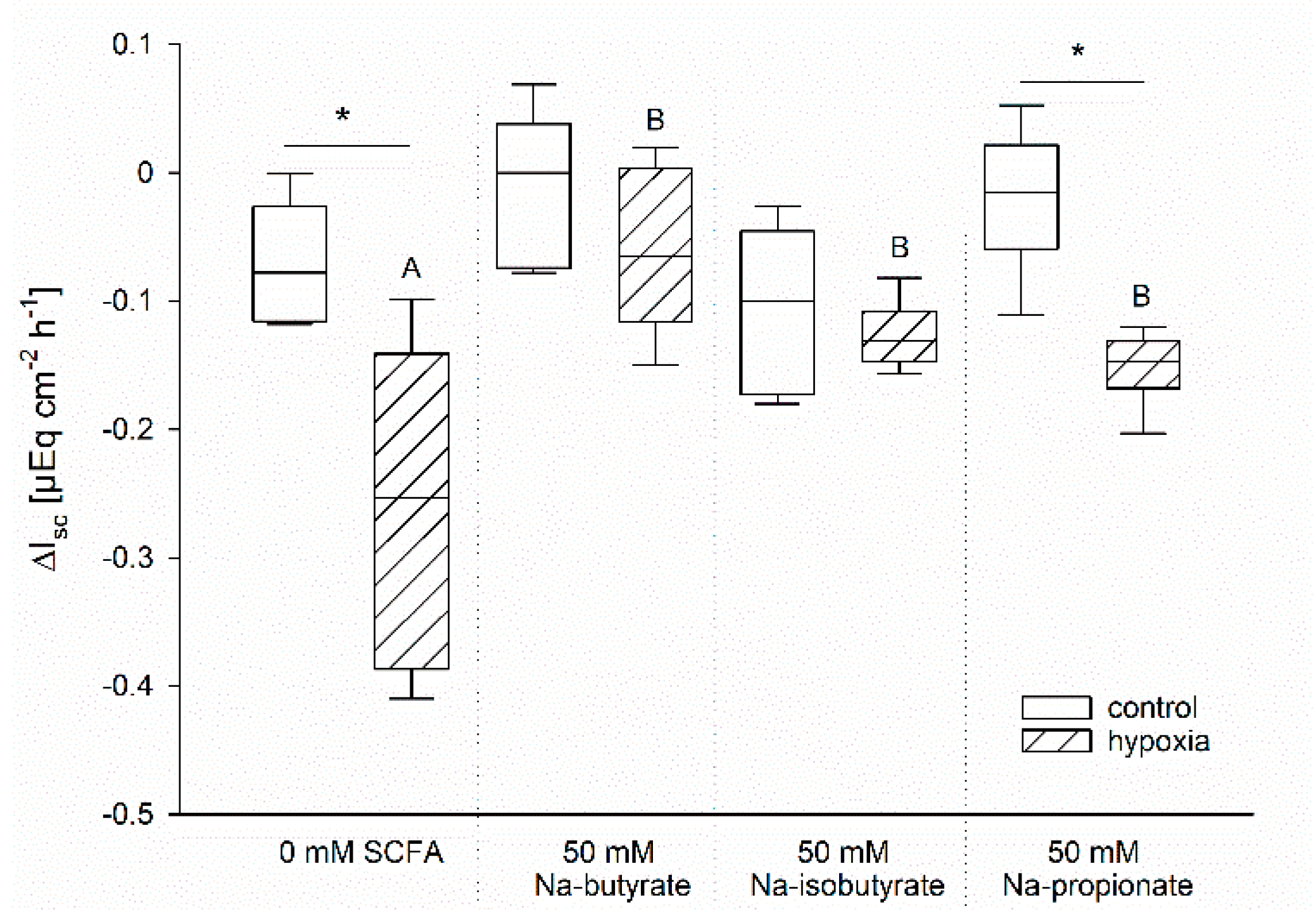
3.3. The Effect of Butyrate Is Not Nutritive
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bayley, H.S. Comparative physiology of the hindgut and its nutritional significance. J. Anim. Sci. 1978, 46, 1800–1802. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Newman, M.A.; Zebeli, Q.; Velde, K.; Grüll, D.; Molnar, T.; Kandler, W.; Metzler-Zebeli, B.U. Enzymatically Modified Starch Favorably Modulated Intestinal Transit Time and Hindgut Fermentation in Growing Pigs. PLoS ONE 2016, 11, e0167784. [Google Scholar] [CrossRef] [PubMed]
- Haenen, D.; Zhang, J.; Souza da Silva, C.; Bosch, G.; van der Meer, I.M.; van Arkel, J.; van den Borne, J.J.G.C.; Pérez Gutiérrez, O.; Smidt, H.; Kemp, B.; et al. A diet high in resistant starch modulates microbiota composition, SCFA concentrations, and gene expression in pig intestine. J. Nutr. 2013, 143, 274–283. [Google Scholar] [CrossRef]
- Evans, M.A.; Shronts, E.P. Intestinal fuels: Glutamine, short-chain fatty acids, and dietary fiber. J. Am. Diet. Assoc. 1992, 92, 1239–1246, 1249. [Google Scholar] [PubMed]
- Wong, J.M.W.; de Souza, R.; Kendall, C.W.C.; Emam, A.; Jenkins, D.J.A. Colonic health: Fermentation and short chain fatty acids. J. Clin. Gastroenterol. 2006, 40, 235–243. [Google Scholar] [CrossRef]
- Cook, S.I.; Sellin, J.H. Review article: Short chain fatty acids in health and disease. Aliment. Pharmacol. Ther. 1998, 12, 499–507. [Google Scholar] [CrossRef]
- Zeitouni, N.E.; Chotikatum, S.; von Köckritz-Blickwede, M.; Naim, H.Y. The impact of hypoxia on intestinal epithelial cell functions: Consequences for invasion by bacterial pathogens. Mol. Cell. Pediatr. 2016, 3, 14. [Google Scholar] [CrossRef]
- Ward, J.B.J.; Keely, S.J.; Keely, S.J. Oxygen in the regulation of intestinal epithelial transport. J. Physiol. 2014, 592, 2473–2489. [Google Scholar] [CrossRef]
- Carreau, A.; El Hafny-Rahbi, B.; Matejuk, A.; Grillon, C.; Kieda, C. Why is the partial oxygen pressure of human tissues a crucial parameter? Small molecules and hypoxia. J. Cell. Mol. Med. 2011, 15, 1239–1253. [Google Scholar] [CrossRef]
- Schofield, C.J.; Ratcliffe, P.J. Signalling hypoxia by HIF hydroxylases. Biochem. Biophys. Res. Commun. 2005, 338, 617–626. [Google Scholar] [CrossRef]
- Pugh, C.W.; Ratcliffe, P.J. New horizons in hypoxia signaling pathways. Exp. Cell Res. 2017, 356, 116–121. [Google Scholar] [CrossRef] [PubMed]
- Boidot, R.; Vegran, F.; Meulle, A.; Le Breton, A.; Dessy, C.; Sonveaux, P.; Lizard-Nacol, S.; Feron, O. Regulation of monocarboxylate transporter MCT1 expression by p53 mediates inward and outward lactate fluxes in tumors. Cancer Res. 2012, 72, 939–948. [Google Scholar] [CrossRef] [PubMed]
- Perez de Heredia, F.; Wood, I.S.; Trayhurn, P. Hypoxia stimulates lactate release and modulates monocarboxylate transporter (MCT1, MCT2, and MCT4) expression in human adipocytes. Pflügers Arch. 2010, 459, 509–518. [Google Scholar] [CrossRef] [PubMed]
- Rademakers, S.E.; Lok, J.; van der Kogel, A.J.; Bussink, J.; Kaanders, J.H. Metabolic markers in relation to hypoxia; staining patterns and colocalization of pimonidazole, HIF-1alpha, CAIX, LDH-5, GLUT-1, MCT1 and MCT4. BMC Cancer 2011, 11, 167. [Google Scholar] [CrossRef]
- Colgan, S.P. Targeting hypoxia in inflammatory bowel disease. J. Investig. Med. 2016, 64, 364–368. [Google Scholar] [CrossRef]
- Hummitzsch, L.; Zitta, K.; Berndt, R.; Wong, Y.L.; Rusch, R.; Hess, K.; Wedel, T.; Gruenewald, M.; Cremer, J.; Steinfath, M.; et al. Remote ischemic preconditioning attenuates intestinal mucosal damage: Insight from a rat model of ischemia-reperfusion injury. J. Transl. Med. 2019, 17, 136. [Google Scholar] [CrossRef]
- Sharma, D.; Maslov, L.N.; Singh, N.; Jaggi, A.S. Remote ischemic preconditioning-induced neuroprotection in cerebral ischemia-reperfusion injury: Preclinical evidence and mechanisms. Eur. J. Pharmacol. 2020, 173380. [Google Scholar] [CrossRef]
- Du, X.; Yang, J.; Liu, C.; Wang, S.; Zhang, C.; Zhao, H.; Du, H.; Geng, X. Hypoxia-Inducible Factor 1α and 2α Have Beneficial Effects in Remote Ischemic Preconditioning Against Stroke by Modulating Inflammatory Responses in Aged Rats. Front. Aging Neurosci. 2020, 12, 54. [Google Scholar] [CrossRef]
- Diao, H.; Jiao, A.R.; Yu, B.; Mao, X.B.; Chen, D.W. Gastric infusion of short-chain fatty acids can improve intestinal barrier function in weaned piglets. Genes Nutr. 2019, 14, 4. [Google Scholar] [CrossRef]
- Dengler, F.; Rackwitz, R.; Benesch, F.; Pfannkuche, H.; Gäbel, G. Both butyrate incubation and hypoxia upregulate genes involved in the ruminal transport of SCFA and their metabolites. J. Anim. Physiol. Anim. Nutr. 2015, 99, 379–390. [Google Scholar] [CrossRef]
- Bedford, A.; Gong, J. Implications of butyrate and its derivatives for gut health and animal production. Anim. Nutr. 2018, 4, 151–159. [Google Scholar] [CrossRef] [PubMed]
- Scheppach, W.; Müller, J.G.; Boxberger, F.; Dusel, G.; Richter, F.; Bartram, H.P.; Christl, S.U.; Dempfle, C.E.; Kasper, H. Histological changes in the colonic mucosa following irrigation with short-chain fatty acids. Eur. J. Gastroenterol. Hepatol. 1997, 9, 163–168. [Google Scholar] [CrossRef] [PubMed]
- Hamer, H.M.; Jonkers, D.; Venema, K.; Vanhoutvin, S.; Troost, F.J.; Brummer, R.-J. Review article: The role of butyrate on colonic function. Aliment. Pharmacol. Ther. 2008, 27, 104–119. [Google Scholar] [CrossRef] [PubMed]
- Gabel, G.; Vogler, S.; Martens, H. Short-chain fatty acids and CO2 as regulators of Na+ and Cl− absorption in isolated sheep rumen mucosa. J. Comp. Physiol. B 1991, 161, 419–426. [Google Scholar] [CrossRef]
- Pfaffl, M.W.; Horgan, G.W.; Dempfle, L. Relative expression software tool (REST) for group-wise comparison and statistical analysis of relative expression results in real-time PCR. Nucleic Acids Res. 2002, 30, e36. [Google Scholar] [CrossRef]
- Dengler, F.; Rackwitz, R.; Pfannkuche, H.; Gäbel, G. Coping with Hypoxia. J. Equine Vet. Sci. 2018, 69, 1–10. [Google Scholar] [CrossRef]
- Carra, G.E.; Ibáñez, J.E.; Saraví, F.D. Electrogenic transport, oxygen consumption, and sensitivity to acute hypoxia of human colonic epithelium. Int. J. Colorectal Dis. 2011, 26, 1205–1210. [Google Scholar] [CrossRef]
- Chu, S.; Montrose, M.H. Extracellular pH regulation in microdomains of colonic crypts: Effects of short-chain fatty acids. Proc. Natl. Acad. Sci. USA 1995, 92, 3303–3307. [Google Scholar] [CrossRef]
- Jaskiewicz, J.; Zhao, Y.; Hawes, J.W.; Shimomura, Y.; Crabb, D.W.; Harris, R.A. Catabolism of isobutyrate by colonocytes. Arch. Biochem. Biophys. 1996, 327, 265–270. [Google Scholar] [CrossRef]
- Herrmann, J.; Hermes, R.; Breves, G. Transepithelial transport and intraepithelial metabolism of short-chain fatty acids (SCFA) in the porcine proximal colon are influenced by SCFA concentration and luminal pH. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2011, 158, 169–176. [Google Scholar] [CrossRef]
- Pfannkuche, H.; Mauksch, A.; Gäbel, G. Modulation of electrogenic transport processes in the porcine proximal colon by enteric neurotransmitters. J. Anim. Physiol. Anim. Nutr. 2012, 96, 482–493. [Google Scholar] [CrossRef] [PubMed]
- Dengler, F.; Gäbel, G. The Fast Lane of Hypoxic Adaptation: Glucose Transport Is Modulated via A HIF-Hydroxylase-AMPK-Axis in Jejunum Epithelium. Int. J. Mol. Sci. 2019, 20, 4993. [Google Scholar] [CrossRef] [PubMed]
- Collins, D.; Kopic, S.; Bachlechner, J.; Ritter, M.; Winter, D.C.; Geibel, J.P. Hypoxia inhibits colonic ion transport via activation of AMP kinase. Ann. Surg. 2011, 254, 957–963. [Google Scholar] [CrossRef]
- Schindele, S.; Pouokam, E.; Diener, M. Hypoxia/Reoxygenation Effects on Ion Transport across Rat Colonic Epithelium. Front. Physiol. 2016, 7, 247. [Google Scholar] [CrossRef] [PubMed]
- Gäbel, G.; Garz, B.; Ahrens, F.; Aschenbach, J.R. Effect of nitric oxide on electrolyte transport across the porcine proximal colon. J. Comp. Physiol. B 2003, 173, 177–186. [Google Scholar] [CrossRef]
- Alper, S.L.; Chernova, M.N.; Stewart, A.K. Regulation of Na+-independent Cl-/HCO3- exchangers by pH. JOP 2001, 2, 171–175. [Google Scholar]
- Jacob, P.; Rossmann, H.; Lamprecht, G.; Kretz, A.; Neff, C.; Lin-Wu, E.; Gregor, M.; Groneberg, D.A.; Kere, J.; Seidler, U. Down-regulated in adenoma mediates apical Cl-/HCO3- exchange in rabbit, rat, and human duodenum. Gastroenterology 2002, 122, 709–724. [Google Scholar] [CrossRef]
- Borthakur, A.; Saksena, S.; Gill, R.K.; Alrefai, W.A.; Ramaswamy, K.; Dudeja, P.K. Regulation of monocarboxylate transporter 1 (MCT1) promoter by butyrate in human intestinal epithelial cells: Involvement of NF-kappaB pathway. J. Cell. Biochem. 2008, 103, 1452–1463. [Google Scholar] [CrossRef]
- Rius, J.; Guma, M.; Schachtrup, C.; Akassoglou, K.; Zinkernagel, A.S.; Nizet, V.; Johnson, R.S.; Haddad, G.G.; Karin, M. NF-kappaB links innate immunity to the hypoxic response through transcriptional regulation of HIF-1alpha. Nature 2008, 453, 807–811. [Google Scholar] [CrossRef]
- Inan, M.S.; Rasoulpour, R.J.; Yin, L.; Hubbard, A.K.; Rosenberg, D.W.; Giardina, C. The luminal short-chain fatty acid butyrate modulates NF-kappaB activity in a human colonic epithelial cell line. Gastroenterology 2000, 118, 724–734. [Google Scholar] [CrossRef]
- Lin, S.-C.; Hardie, D.G. AMPK: Sensing Glucose as well as Cellular Energy Status. Cell Metab. 2018, 27, 299–313. [Google Scholar] [CrossRef] [PubMed]
- Dengler, F.; Rackwitz, R.; Pfannkuche, H.; Gäbel, G. Glucose transport across lagomorph jejunum epithelium is modulated by AMP-activated protein kinase (AMPK) under hypoxia. J. Appl. Physiol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Hardie, D.G. AMPK as a direct sensor of long-chain fatty acyl-CoA esters. Nat. Metab. 2020. [Google Scholar] [CrossRef]
- Wang, F.; Wu, H.; Fan, M.; Yu, R.; Zhang, Y.; Liu, J.; Zhou, X.; Cai, Y.; Huang, S.; Hu, Z.; et al. Sodium butyrate inhibits migration and induces AMPK-mTOR pathway-dependent autophagy and ROS-mediated apoptosis via the miR-139-5p/Bmi-1 axis in human bladder cancer cells. FASEB J. 2020, 34, 4266–4282. [Google Scholar] [CrossRef] [PubMed]
- Kumar, J.; Rani, K.; Datt, C. Molecular link between dietary fibre, gut microbiota and health. Mol. Biol. Rep. 2020. [Google Scholar] [CrossRef] [PubMed]
- Gill, P.A.; van Zelm, M.C.; Muir, J.G.; Gibson, P.R. Review article: Short chain fatty acids as potential therapeutic agents in human gastrointestinal and inflammatory disorders. Aliment. Pharmacol. Ther. 2018, 48, 15–34. [Google Scholar] [CrossRef]
- Borthakur, A.; Priyamvada, S.; Kumar, A.; Natarajan, A.A.; Gill, R.K.; Alrefai, W.A.; Dudeja, P.K. A novel nutrient sensing mechanism underlies substrate-induced regulation of monocarboxylate transporter-1. Am. J. Physiol. Gastrointest. Liver Physiol. 2012, 303, G1126–G1133. [Google Scholar] [CrossRef]
- Zhang, J.; Cheng, S.; Wang, Y.; Yu, X.; Li, J. Identification and characterization of the free fatty acid receptor 2 (FFA2) and a novel functional FFA2-like receptor (FFA2L) for short-chain fatty acids in pigs: Evidence for the existence of a duplicated FFA2 gene (FFA2L) in some mammalian species. Domest. Anim. Endocrinol. 2014, 47, 108–118.e1. [Google Scholar] [CrossRef]
- Brown, A.J.; Goldsworthy, S.M.; Barnes, A.A.; Eilert, M.M.; Tcheang, L.; Daniels, D.; Muir, A.I.; Wigglesworth, M.J.; Kinghorn, I.; Fraser, N.J.; et al. The Orphan G protein-coupled receptors GPR41 and GPR43 are activated by propionate and other short chain carboxylic acids. J. Biol. Chem. 2003, 278, 11312–11319. [Google Scholar] [CrossRef]
- Chambers, E.S.; Morrison, D.J.; Frost, G. Control of appetite and energy intake by SCFA: What are the potential underlying mechanisms? Proc. Nutr. Soc. 2015, 74, 328–336. [Google Scholar] [CrossRef]
- Baaske, L.; Masur, F.; Dengler, F.; Rackwitz, R.; Kaiser, B.; Pfannkuche, H.; Gäbel, G. Possible influence of free fatty acid receptors on pH regulation in the ruminal epithelium of sheep. J. Anim. Physiol. Anim. Nutr. 2020, 104, 776–789. [Google Scholar] [CrossRef]
- Van Welden, S.; Selfridge, A.C.; Hindryckx, P. Intestinal hypoxia and hypoxia-induced signalling as therapeutic targets for IBD. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 596–611. [Google Scholar] [CrossRef] [PubMed]
- Shah, Y.M. The role of hypoxia in intestinal inflammation. Mol. Cell. Pediatr. 2016, 3, 1. [Google Scholar] [CrossRef] [PubMed]
- Colgan, S.P.; Taylor, C.T. Hypoxia: An alarm signal during intestinal inflammation. Nat. Rev. Gastroenterol. Hepatol. 2010, 7, 281–287. [Google Scholar] [CrossRef] [PubMed]
- Treem, W.R.; Ahsan, N.; Shoup, M.; Hyams, J.S. Fecal short-chain fatty acids in children with inflammatory bowel disease. J. Pediatr. Gastroenterol. Nutr. 1994, 18, 159–164. [Google Scholar] [CrossRef]
- Roediger, W.E. Colonic epithelial metabolism in ulcerative colitis. Gut 1993, 34, 1646. [Google Scholar] [CrossRef][Green Version]
- Scheppach, W.; Christl, S.U.; Bartram, H.P.; Richter, F.; Kasper, H. Effects of short-chain fatty acids on the inflamed colonic mucosa. Scand. J. Gastroenterol. Suppl. 1997, 222, 53–57. [Google Scholar] [CrossRef]
- Scheppach, W.; Sommer, H.; Kirchner, T.; Paganelli, G.-M.; Bartram, P.; Christl, S.; Richter, F.; Dusel, G.; Kasper, H. Effect of butyrate enemas on the colonic mucosa in distal ulcerative colitis. Gastroenterology 1992, 103, 51–56. [Google Scholar] [CrossRef]
- Zhou, C.; Li, L.; Li, T.; Sun, L.; Yin, J.; Guan, H.; Wang, L.; Zhu, H.; Xu, P.; Fan, X.; et al. SCFAs induce autophagy in intestinal epithelial cells and relieve colitis by stabilizing HIF-1α. J. Mol. Med. 2020. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene Bank Accession No. | Primer Sequence (5′–3′) | Annealing Temperature (°C) | Amplicon Length (bp) |
|---|---|---|---|---|
| DRA | NM_001130248.1 | F: CTTTGCTGTGGCCTTTTCTGTG R: ACCCGCCGCATATGTTACTCA | 60 | 110 |
| GLUT 1 | XM_021096908.1 | F: GGTTCATTGTGGCCGAACTC R: TACTGGAAGCACATGCCCAC | 60 | 108 |
| MCT 1 | NM_001128445.1 | F: TGATGGACCTTGTTGGACCTC R: GAGACGACCTAAAAGTGGTGG | 60 | 103 |
| RPL4 | XM_005659862.3 | F: GCACCACGCAAGAAGATTCA R: TGTCTTTGCATACGGGTTTAGC | 57 | 92 |
| TBP | XM_021085483.1 | F: GCGATTTGCTGCTGTAATCA R: GTCTGGACTGTTCTTCACTCT | 57 | 109 |
| YWHAZ | NM_001315726.1 | F: GGCCCTTAACTTCTCTGTGTT R: GGCTTCATCAAATGCTGTCT | 57 | 87 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dengler, F.; Kraetzig, A.; Gäbel, G. Butyrate Protects Porcine Colon Epithelium from Hypoxia-Induced Damage on a Functional Level. Nutrients 2021, 13, 305. https://doi.org/10.3390/nu13020305
Dengler F, Kraetzig A, Gäbel G. Butyrate Protects Porcine Colon Epithelium from Hypoxia-Induced Damage on a Functional Level. Nutrients. 2021; 13(2):305. https://doi.org/10.3390/nu13020305
Chicago/Turabian StyleDengler, Franziska, Anika Kraetzig, and Gotthold Gäbel. 2021. "Butyrate Protects Porcine Colon Epithelium from Hypoxia-Induced Damage on a Functional Level" Nutrients 13, no. 2: 305. https://doi.org/10.3390/nu13020305
APA StyleDengler, F., Kraetzig, A., & Gäbel, G. (2021). Butyrate Protects Porcine Colon Epithelium from Hypoxia-Induced Damage on a Functional Level. Nutrients, 13(2), 305. https://doi.org/10.3390/nu13020305