
DANCR Mediates the Rescuing Effects of Sesamin on Postmenopausal Osteoporosis Treatment via Orchestrating Osteogenesis and Osteoclastogenesis


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Result
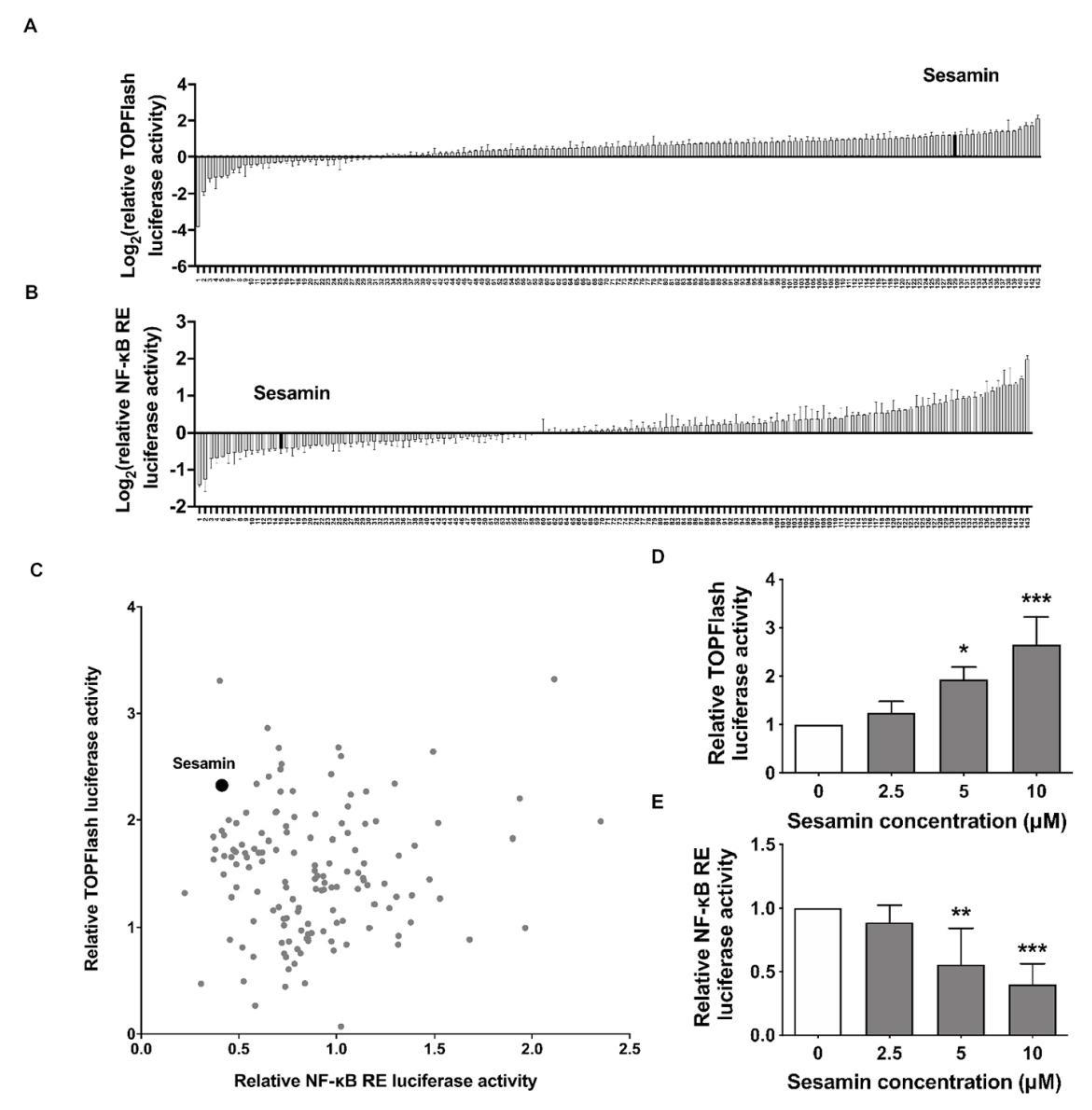
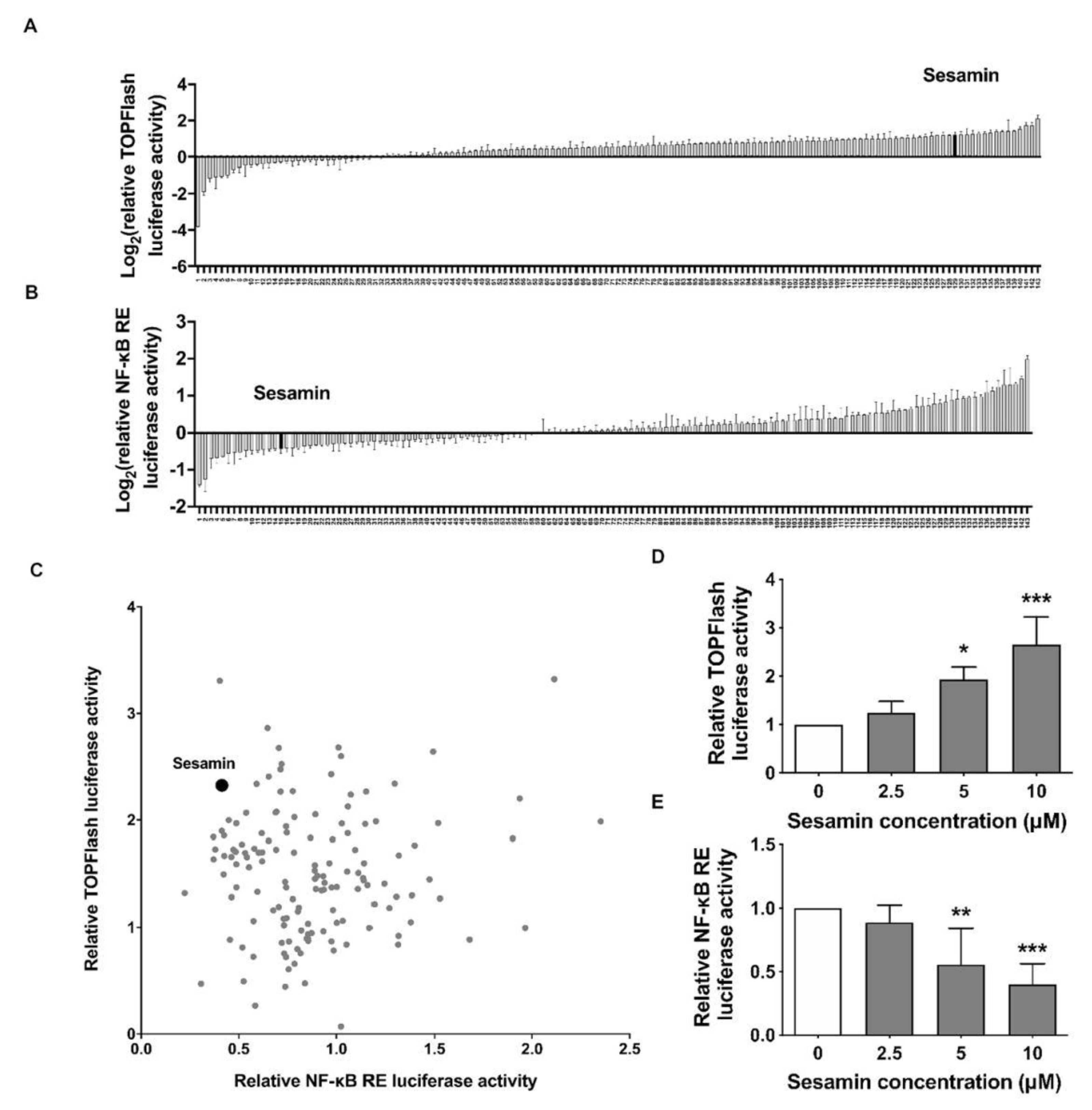
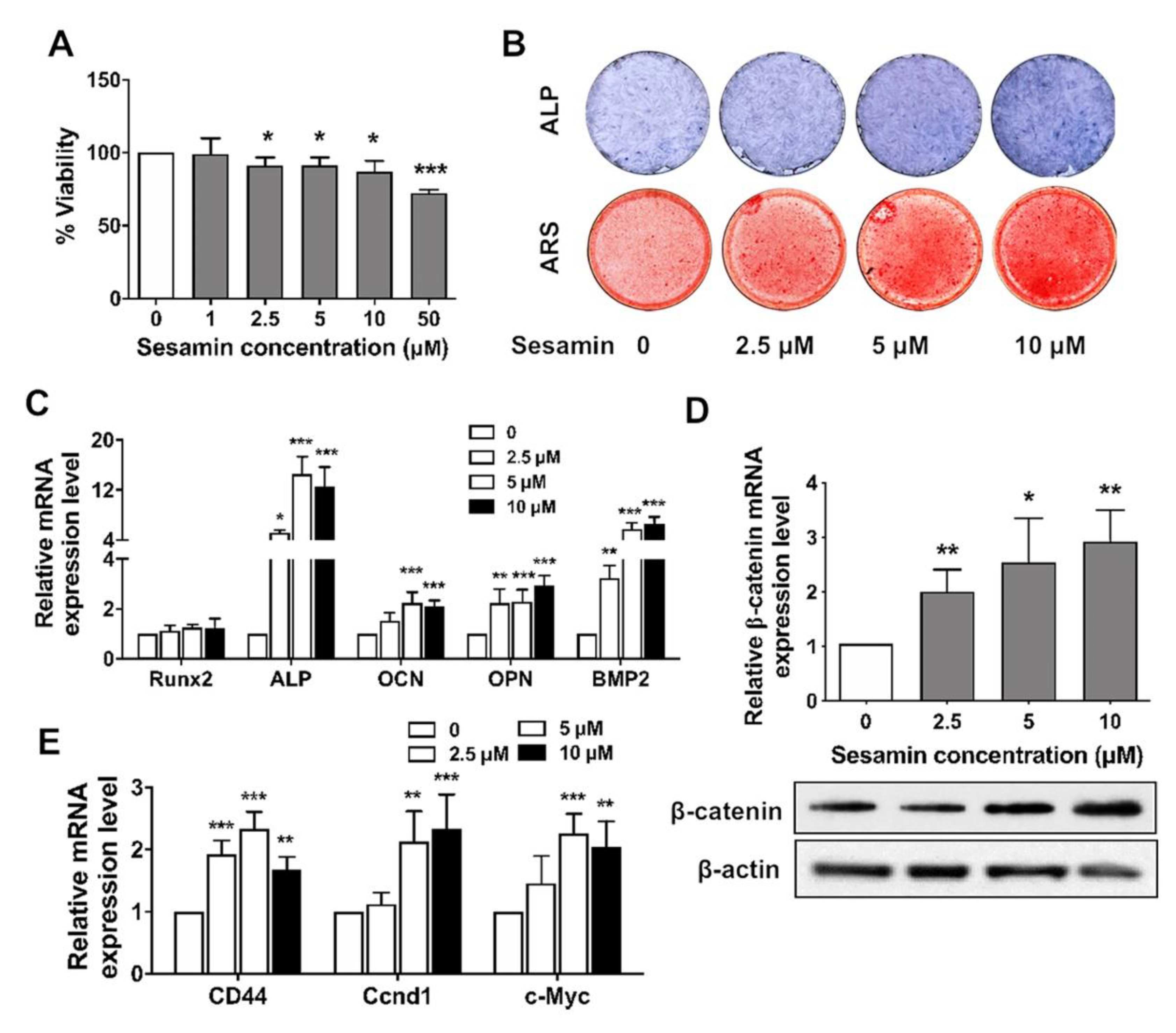
2.1. Identification of Sesamin as an Up-Regulator of Wnt/β-Catenin and Down-Regulator of NF-κB Signaling
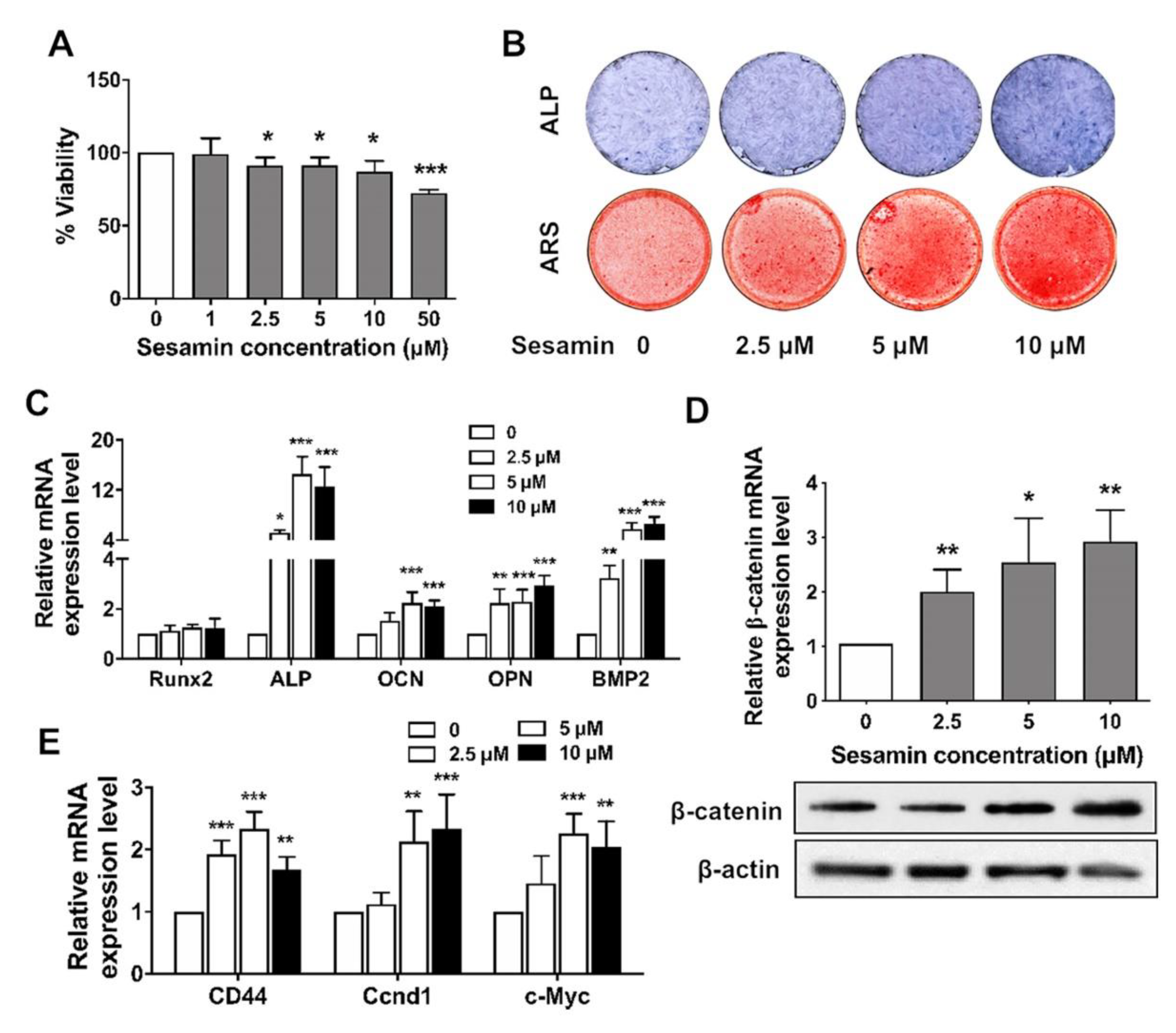
2.2. Sesamin Promotes Osteogenic Differentiation of BMSCs by Activating Wnt/β-Catenin Signaling
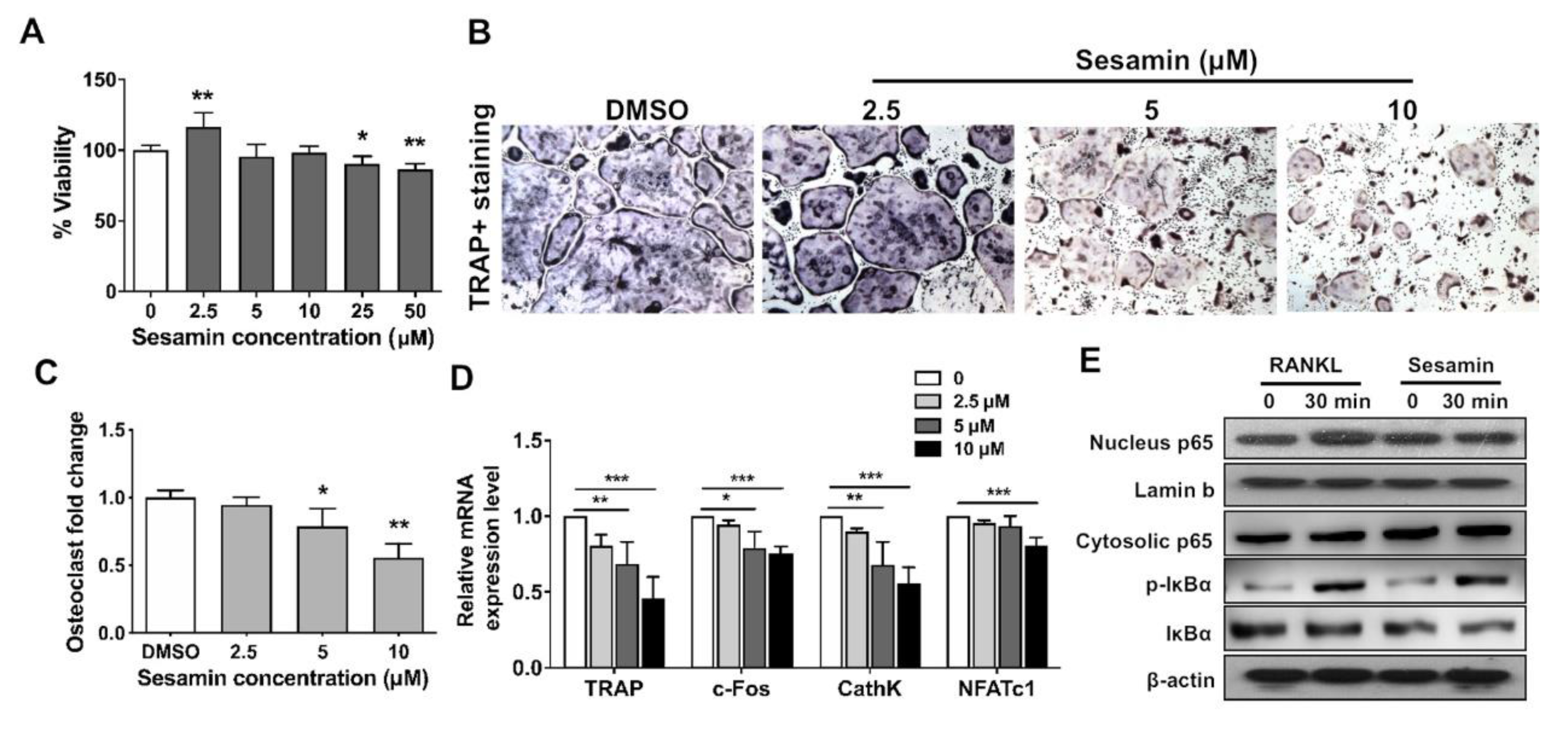
2.3. Sesamin Inhibits Osteoclastogenic Differentiation of RAW264.7 Cell by Deactivating NF-B Signaling Pathway
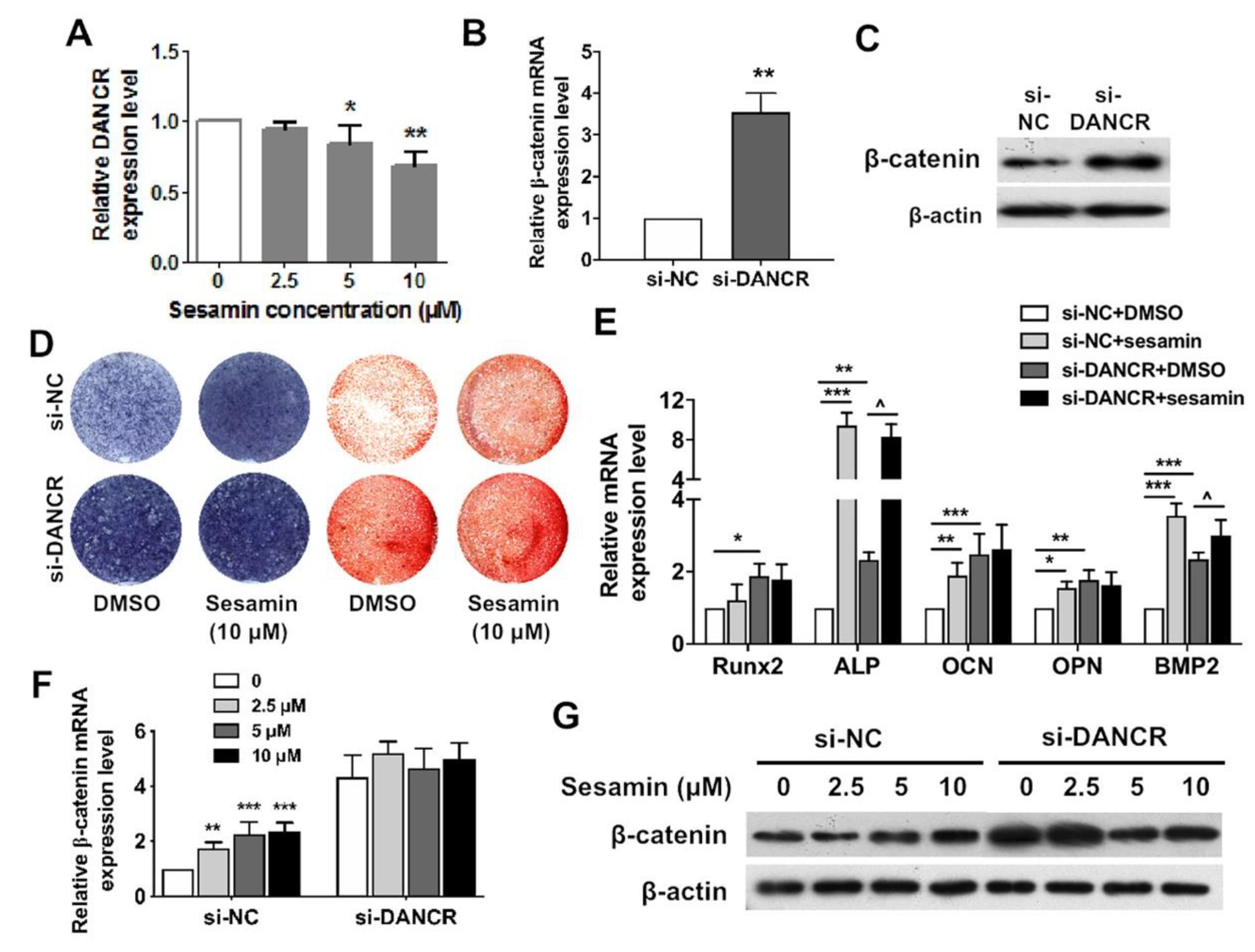
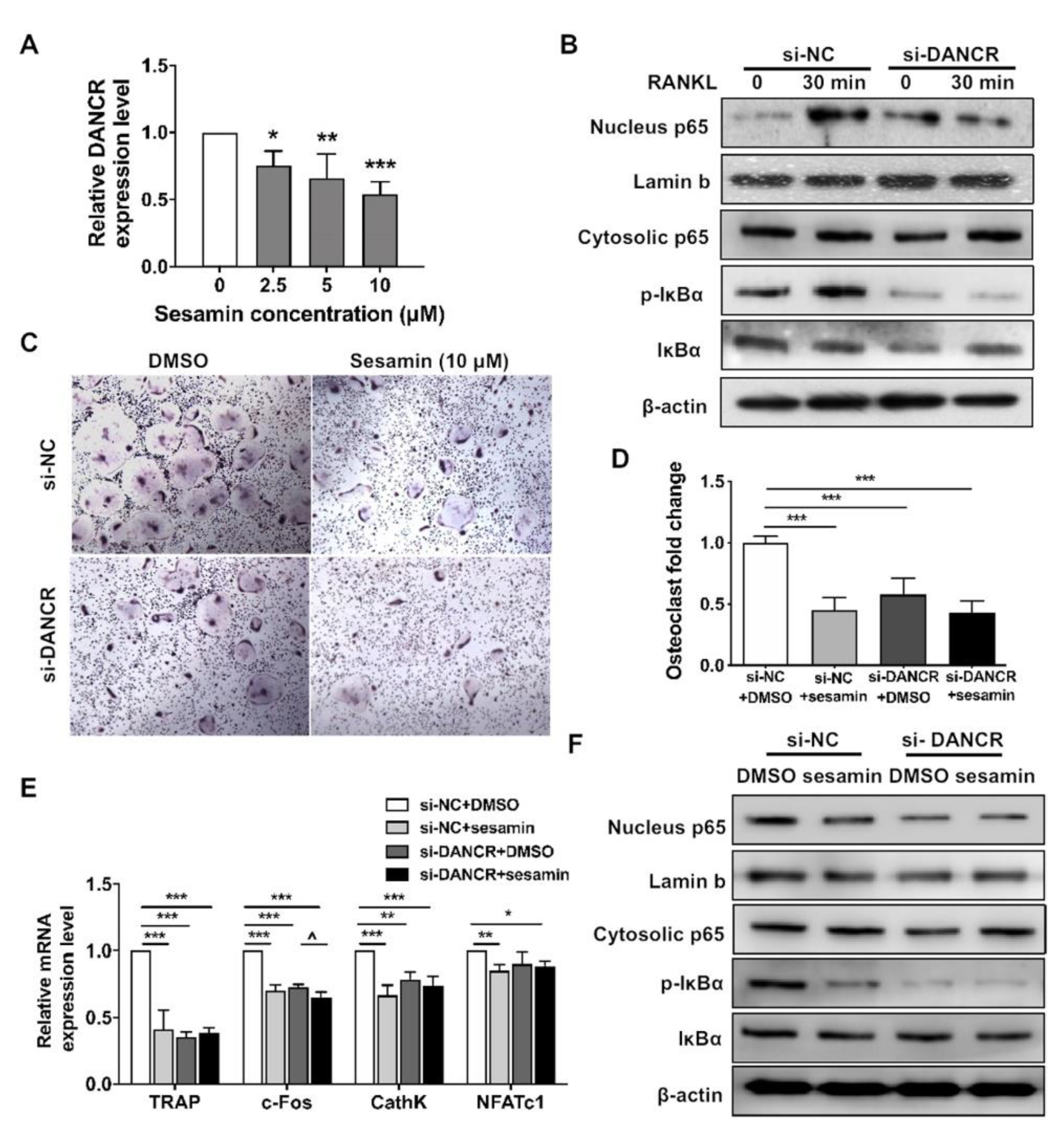
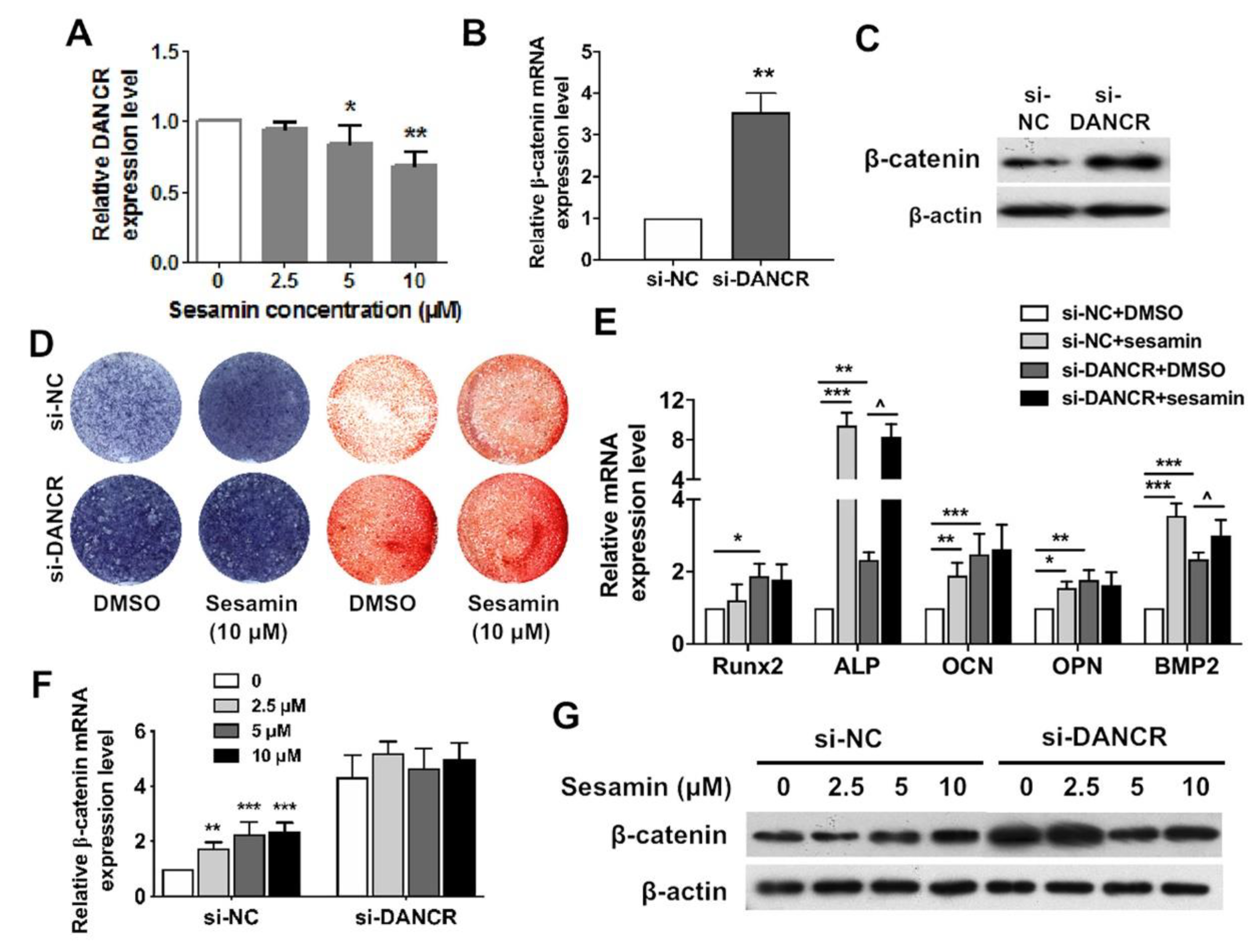
2.4. Osteogenic Role of Sesamin Is Dependent on DANCR Expression
2.5. DANCR Mediates the Sesamin Repressed Osteoclastogneesis
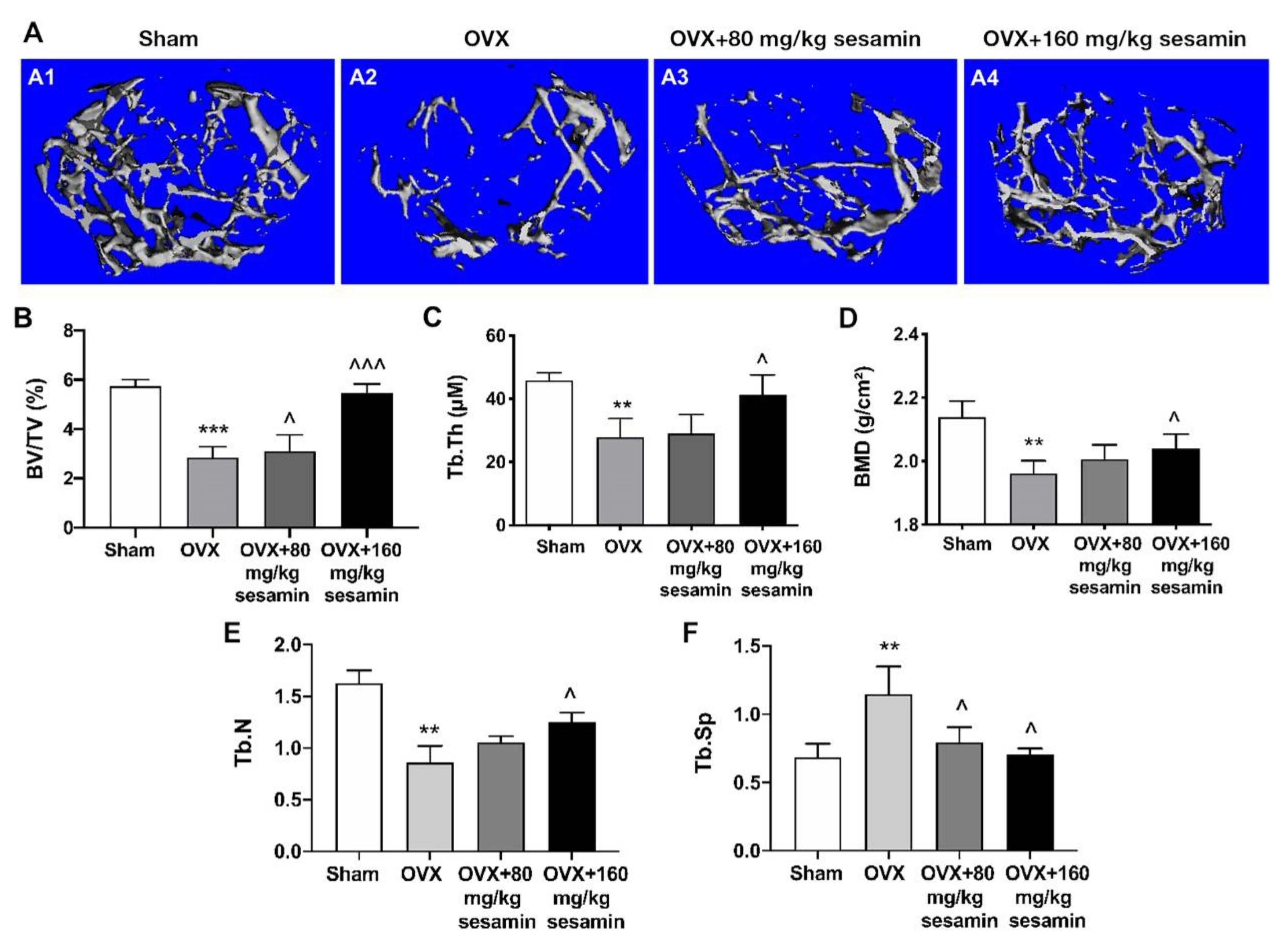
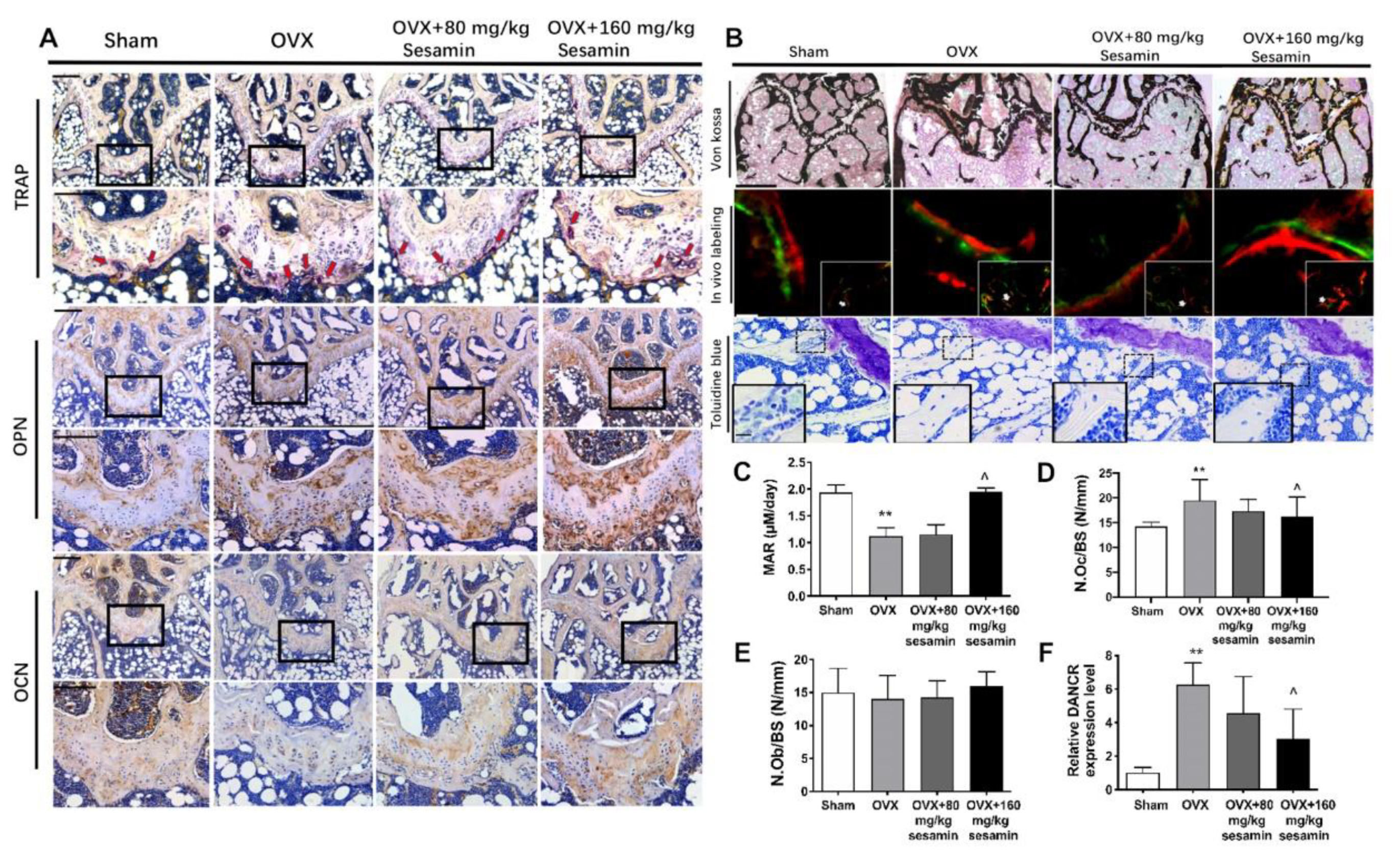
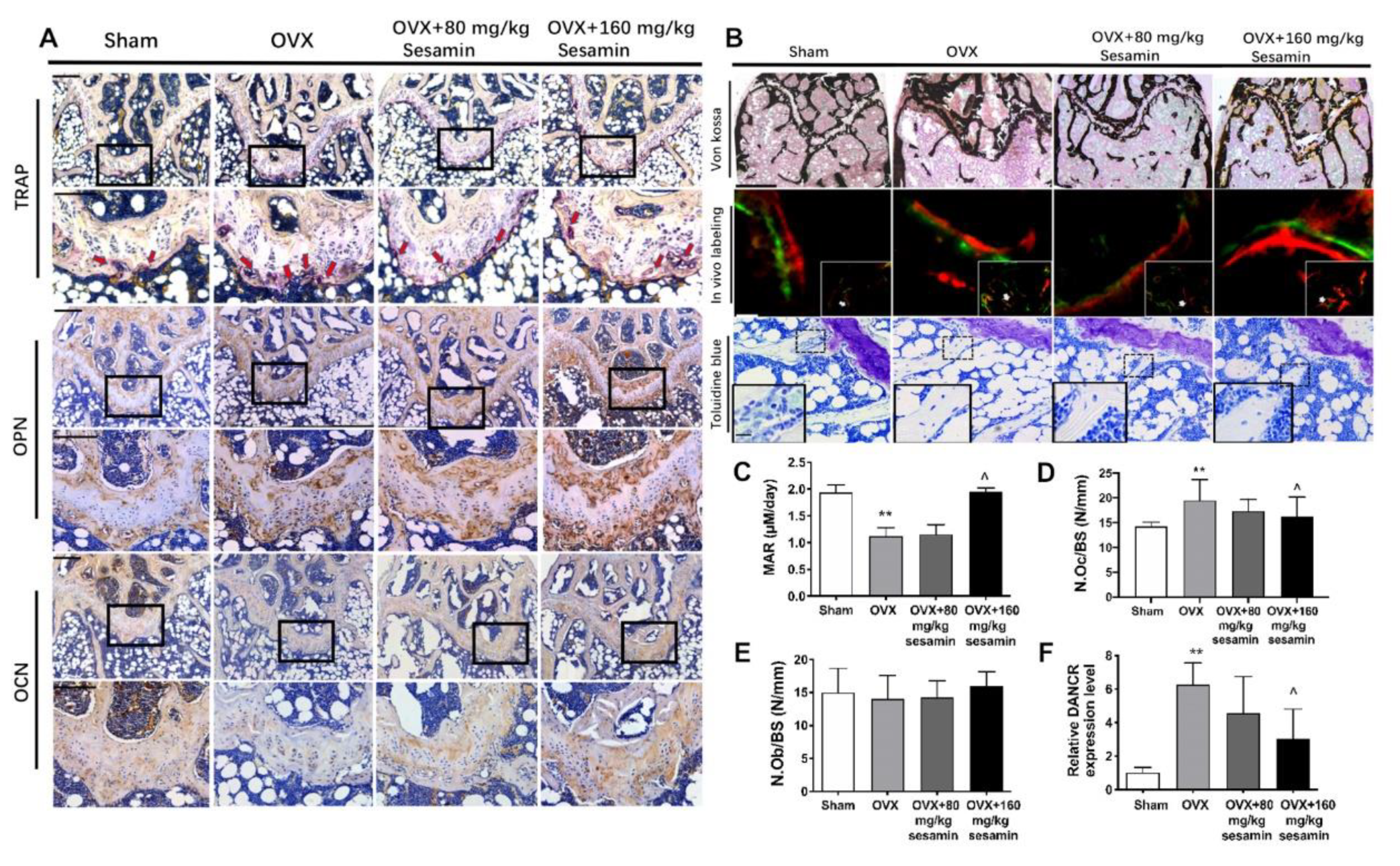
2.6. Sesamin Attenuates Bone Loss in OVX Mice Osteoporosis Model via Down-Regulating DANCR Expression
3. Discussion
4. Materials and Methods
4.1. Small Molecule Library
4.2. Cell Culture
4.3. Transient Transfection
4.4. Luciferase Activity Screening
4.5. Cell Viability Assay
4.6. Osteogenic Differentiation
4.7. Osteoclastogenesis
4.8. Mouse Ovariectomy (OVX)-Induced Osteoporosis Model
4.9. Micro-Computed Tomography (μCT) Analysis
4.10. Histology and Immunohistochemistry
4.11. RNA Extraction and Real-Time PCR
4.12. Western Blot Analysis
4.13. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Zhu, Y.; Huang, Z.; Wang, Y.; Xu, W.; Chen, H.; Xu, J.; Luo, S.; Zhang, Y.; Zhao, D.; Hu, J. The efficacy and safety of denosumab in postmenopausal women with osteoporosis previously treated with bisphosphonates: A review. J. Orthop. Translat. 2020, 22, 7–13. [Google Scholar] [CrossRef]
- Bonaiuti, D.; Shea, B.; Iovine, R.; Negrini, S.; Robinson, V.; Kemper, H.C.; Wells, G.; Tugwell, P.; Cranney, A. Exercise for preventing and treating osteoporosis in postmenopausal women. Cochrane Database Syst. Rev. 2002, CD000333. [Google Scholar] [CrossRef]
- Sehmisch, S.; Hammer, F.; Christoffel, J.; Seidlova-Wuttke, D.; Tezval, M.; Wuttke, W.; Stuermer, K.M.; Stuermer, E.K. Comparison of the phytohormones genistein, resveratrol and 8-prenylnaringenin as agents for preventing osteoporosis. Planta Med. 2008, 74, 794–801. [Google Scholar] [CrossRef] [PubMed]
- Saad, F.A. Novel insights into the complex architecture of osteoporosis molecular genetics. Ann. N. Y. Acad. Sci. 2020, 1462, 37–52. [Google Scholar] [CrossRef] [PubMed]
- Felber, K.; Elks, P.M.; Lecca, M.; Roehl, H.H. Expression of osterix Is Regulated by FGF and Wnt/beta-Catenin Signalling during Osteoblast Differentiation. PLoS ONE 2015, 10, e0144982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.H.; Lee, N.K.; Lee, S.Y. Current Understanding of RANK Signaling in Osteoclast Differentiation and Maturation. Mol. Cells 2017, 40, 706–713. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Peng, X.; Qin, Y.; Wang, R.; Tang, J.; Cui, X.; Wang, T.; Liu, W.; Pan, H.; Li, B. Acceleration of bone regeneration by activating Wnt/beta-catenin signalling pathway via lithium released from lithium chloride/calcium phosphate cement in osteoporosis. Sci. Rep. 2017, 7, 45204. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.L.; Chen, J.J.; Zhang, G.N.; Wang, Y.C.; Si, S.Y.; Chen, L.F.; Wang, Z. Small molecule T63 suppresses osteoporosis by modulating osteoblast differentiation via BMP and WNT signaling pathways. Sci. Rep. 2017, 7, 10397. [Google Scholar] [CrossRef] [Green Version]
- Hu, J.P.; Nishishita, K.; Sakai, E.; Yoshida, H.; Kato, Y.; Tsukuba, T.; Okamoto, K. Berberine inhibits RANKL-induced osteoclast formation and survival through suppressing the NF-kappaB and Akt pathways. Eur. J. Pharmacol. 2008, 580, 70–79. [Google Scholar] [CrossRef]
- Wattel, A.; Kamel, S.; Prouillet, C.; Petit, J.P.; Lorget, F.; Offord, E.; Brazier, M. Flavonoid quercetin decreases osteoclastic differentiation induced by RANKL via a mechanism involving NF kappa B and AP-1. J. Cell Biochem. 2004, 92, 285–295. [Google Scholar] [CrossRef] [PubMed]
- Thummuri, D.; Guntuku, L.; Challa, V.S.; Ramavat, R.N.; Naidu, V.G.M. Abietic acid attenuates RANKL induced osteoclastogenesis and inflammation associated osteolysis by inhibiting the NF-KB and MAPK signaling. J. Cell Physiol. 2018, 234, 443–453. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Wang, H.; Huang, M.; Zong, Z.; Wu, X.; Xu, J.; Lan, H.; Zheng, J.; Zhang, X.; Lee, Y.W.; et al. Asiatic Acid Attenuates Bone Loss by Regulating Osteoclastic Differentiation. Calcif. Tissue Int. 2019, 105, 531–545. [Google Scholar] [CrossRef]
- Chotiyarnwong, P.; McCloskey, E.V. Pathogenesis of glucocorticoid-induced osteoporosis and options for treatment. Nat. Rev. Endocrinol. 2020, 16, 437–447. [Google Scholar] [CrossRef] [PubMed]
- Rezende, E.; Bradaschia-Correa, V.; Siviero, F.; Ambrosio, L.M.B.; Arana-Chavez, V.E. Effects of bisphosphonates on osteogenesis and osteoclastogenesis signaling during the endochondral ossification of growing rats. Cell Tissue Res. 2017, 368, 287–300. [Google Scholar] [CrossRef] [PubMed]
- Vahle, J.L.; Sato, M.; Long, G.G.; Young, J.K.; Francis, P.C.; Engelhardt, J.A.; Westmore, M.S.; Linda, Y.; Nold, J.B. Skeletal changes in rats given daily subcutaneous injections of recombinant human parathyroid hormone (1–34) for 2 years and relevance to human safety. Toxicol. Pathol. 2002, 30, 312–321. [Google Scholar] [CrossRef]
- Chavassieux, P.; Chapurlat, R.; Portero-Muzy, N.; Roux, J.P.; Garcia, P.; Brown, J.P.; Libanati, C.; Boyce, R.W.; Wang, A.; Grauer, A. Bone-Forming and Antiresorptive Effects of Romosozumab in Postmenopausal Women With Osteoporosis: Bone Histomorphometry and Microcomputed Tomography Analysis After 2 and 12 Months of Treatment. J. Bone Miner. Res. Off. J. Am. Soc. Bone Mineral. Res. 2019, 34, 1597–1608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chapurlat, R.D. Treatment of postmenopausal osteoporosis with odanacatib. Expert Opin. Pharmacother. 2014, 15, 559–564. [Google Scholar] [CrossRef]
- Jeng, K.C.; Hou, R.C.; Wang, J.C.; Ping, L.I. Sesamin inhibits lipopolysaccharide-induced cytokine production by suppression of p38 mitogen-activated protein kinase and nuclear factor-kappaB. Immunol. Lett. 2005, 97, 101–106. [Google Scholar] [CrossRef]
- Phitak, T.; Pothacharoen, P.; Settakorn, J.; Poompimol, W.; Caterson, B.; Kongtawelert, P. Chondroprotective and anti-inflammatory effects of sesamin. Phytochemistry 2012, 80, 77–88. [Google Scholar] [CrossRef] [PubMed]
- Wanachewin, O.; Boonmaleerat, K.; Pothacharoen, P.; Reutrakul, V.; Kongtawelert, P. Sesamin stimulates osteoblast differentiation through p38 and ERK1/2 MAPK signaling pathways. BMC Complement. Altern Med. 2012, 12, 71. [Google Scholar] [CrossRef] [Green Version]
- Ma, Z.P.; Zhang, Z.F.; Yang, Y.F.; Yang, Y. Sesamin Promotes Osteoblastic Differentiation and Protects Rats from Osteoporosis. Med. Sci. Monit. 2019, 25, 5312–5320. [Google Scholar] [CrossRef] [PubMed]
- Wanachewin, O.; Pothacharoen, P.; Kongtawelert, P.; Phitak, T. Inhibitory effects of sesamin on human osteoclastogenesis. Arch. Pharm. Res. 2017, 40, 1186–1196. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.Q.; Hong, Z.L.; Zhan, L.B.; Chu, H.Y.; Zhang, X.Z.; Li, G.H. Wedelolactone enhances osteoblastogenesis by regulating Wnt/beta-catenin signaling pathway but suppresses osteoclastogenesis by NF-kappaB/c-fos/NFATc1 pathway. Sci. Rep. 2016, 6, 32260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sikora, M.; Marycz, K.; Smieszek, A. Small and Long Non-coding RNAs as Functional Regulators of Bone Homeostasis, Acting Alone or Cooperatively. Mol. Ther. Nucleic Acids 2020, 21, 792–803. [Google Scholar] [CrossRef]
- Liang, W.C.; Fu, W.M.; Wang, Y.B.; Sun, Y.X.; Xu, L.L.; Wong, C.W.; Chan, K.M.; Li, G.; Waye, M.M.; Zhang, J.F. H19 activates Wnt signaling and promotes osteoblast differentiation by functioning as a competing endogenous RNA. Sci. Rep. 2016, 6, 20121. [Google Scholar] [CrossRef] [PubMed]
- Feng, L.; Shi, L.; Lu, Y.F.; Wang, B.; Tang, T.; Fu, W.M.; He, W.; Li, G.; Zhang, J.F. Linc-ROR Promotes Osteogenic Differentiation of Mesenchymal Stem Cells by Functioning as a Competing Endogenous RNA for miR-138 and miR-145. Mol. Ther. Nucleic. Acids 2018, 11, 345–353. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Yujiao, W.; Fang, W.; Linhui, Y.; Ziqi, G.; Zhichen, W.; Zirui, W.; Shengwang, W. The roles of miRNA, lncRNA and circRNA in the development of osteoporosis. Biol. Res. 2020, 53, 40. [Google Scholar] [CrossRef]
- Sun, Y.; Cai, M.; Zhong, J.; Yang, L.; Xiao, J.; Jin, F.; Xue, H.; Liu, X.; Liu, H.; Zhang, Y.; et al. The long noncoding RNA lnc-ob1 facilitates bone formation by upregulating Osterix in osteoblasts. Nat. Metab. 2019, 1, 485–496. [Google Scholar] [CrossRef]
- Jin, F.; Li, J.; Zhang, Y.B.; Liu, X.; Cai, M.; Liu, M.; Li, M.; Ma, C.; Yue, R.; Zhu, Y.; et al. A functional motif of long noncoding RNA Nron against osteoporosis. Nat. Commun. 2021, 12, 3319. [Google Scholar] [CrossRef]
- Wang, C.G.; Hu, Y.H.; Su, S.L.; Zhong, D. LncRNA DANCR and miR-320a suppressed osteogenic differentiation in osteoporosis by directly inhibiting the Wnt/beta-catenin signaling pathway. Exp. Mol. Med. 2020, 52, 1310–1325. [Google Scholar] [CrossRef] [PubMed]
- Tong, X.; Gu, P.C.; Xu, S.Z.; Lin, X.J. Long non-coding RNA-DANCR in human circulating monocytes: A potential biomarker associated with postmenopausal osteoporosis. Biosci. Biotechnol. Biochem. 2015, 79, 732–737. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Zhou, G.; Li, M.; Hu, D.; Zhang, L.; Liu, P.; Lin, K. Long noncoding RNA DANCR mediates cisplatin resistance in glioma cells via activating AXL/PI3K/Akt/NF-kappaB signaling pathway. Neurochem. Int. 2018, 118, 233–241. [Google Scholar] [CrossRef] [PubMed]
- Feng, L.; Zhang, J.F.; Shi, L.; Yang, Z.M.; Wu, T.Y.; Wang, H.X.; Lin, W.P.; Lu, Y.F.; Lo, J.H.T.; Zhu, D.H.; et al. MicroRNA-378 Suppressed Osteogenesis of MSCs and Impaired Bone Formation via Inactivating Wnt/beta-Catenin Signaling. Mol. Ther. Nucleic Acids 2020, 21, 1017–1028. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.; Huang, J.; Zheng, L.; Liu, Y.; Liu, G.; Li, N.; Wang, K.; Zou, L.; Wu, T.; Qin, L.; et al. Glucocorticoid-induced osteoporosis in growing rats. Calcif. Tissue Int. 2014, 95, 362–373. [Google Scholar] [CrossRef]
- Feng, L.; Yang, Z.M.; Li, Y.C.; Wang, H.X.; Lo, J.H.T.; Zhang, X.T.; Li, G. Linc-ROR promotes mesenchymal stem cells chondrogenesis and cartilage formation via regulating SOX9 expression. Osteoarthr. Cartil. 2021, 29, 568–578. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Feng, L.; Huang, J.; Zhang, X.; Lin, W.; Wang, B.; Cui, L.; Lin, S.; Li, G. Asiatic acid protects articular cartilage through promoting chondrogenesis and inhibiting inflammation and hypertrophy in osteoarthritis. Eur. J. Pharmacol. 2021, 907, 174265. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, Z.; Feng, L.; Wang, H.; Li, Y.; Lo, J.H.T.; Zhang, X.; Lu, X.; Wang, Y.; Lin, S.; Tortorella, M.D.; et al. DANCR Mediates the Rescuing Effects of Sesamin on Postmenopausal Osteoporosis Treatment via Orchestrating Osteogenesis and Osteoclastogenesis. Nutrients 2021, 13, 4455. https://doi.org/10.3390/nu13124455
Yang Z, Feng L, Wang H, Li Y, Lo JHT, Zhang X, Lu X, Wang Y, Lin S, Tortorella MD, et al. DANCR Mediates the Rescuing Effects of Sesamin on Postmenopausal Osteoporosis Treatment via Orchestrating Osteogenesis and Osteoclastogenesis. Nutrients. 2021; 13(12):4455. https://doi.org/10.3390/nu13124455
Chicago/Turabian StyleYang, Zhengmeng, Lu Feng, Haixing Wang, Yucong Li, Jessica Hiu Tung Lo, Xiaoting Zhang, Xuan Lu, Yaofeng Wang, Sien Lin, Micky D. Tortorella, and et al. 2021. "DANCR Mediates the Rescuing Effects of Sesamin on Postmenopausal Osteoporosis Treatment via Orchestrating Osteogenesis and Osteoclastogenesis" Nutrients 13, no. 12: 4455. https://doi.org/10.3390/nu13124455
APA StyleYang, Z., Feng, L., Wang, H., Li, Y., Lo, J. H. T., Zhang, X., Lu, X., Wang, Y., Lin, S., Tortorella, M. D., & Li, G. (2021). DANCR Mediates the Rescuing Effects of Sesamin on Postmenopausal Osteoporosis Treatment via Orchestrating Osteogenesis and Osteoclastogenesis. Nutrients, 13(12), 4455. https://doi.org/10.3390/nu13124455