
Inhibitory Effects of Luteolin 7-Methyl Ether Isolated from Wikstroemia ganpi on Tnf-A/Ifn-Γ Mixture-Induced Inflammation in Human Keratinocyte


Abstract
:1. Introduction
2. Materials and Methods
2.1. General
2.2. Plant Material and Extraction
2.3. Isolation of Compounds from W. ganpi Extract
2.4. Quantitative Real-Time PCR Analysis of IL-4 mRNA Expression in RBL-2H Cells
2.5. Quantitative Real-Time PCR Analysis of the Expressions of TRPA1, TRPV1, IL-31, IL-6, GM-CSF, and G-CSF mRNA in HaCaT Cells
2.6. Statistical Analyses
3. Results
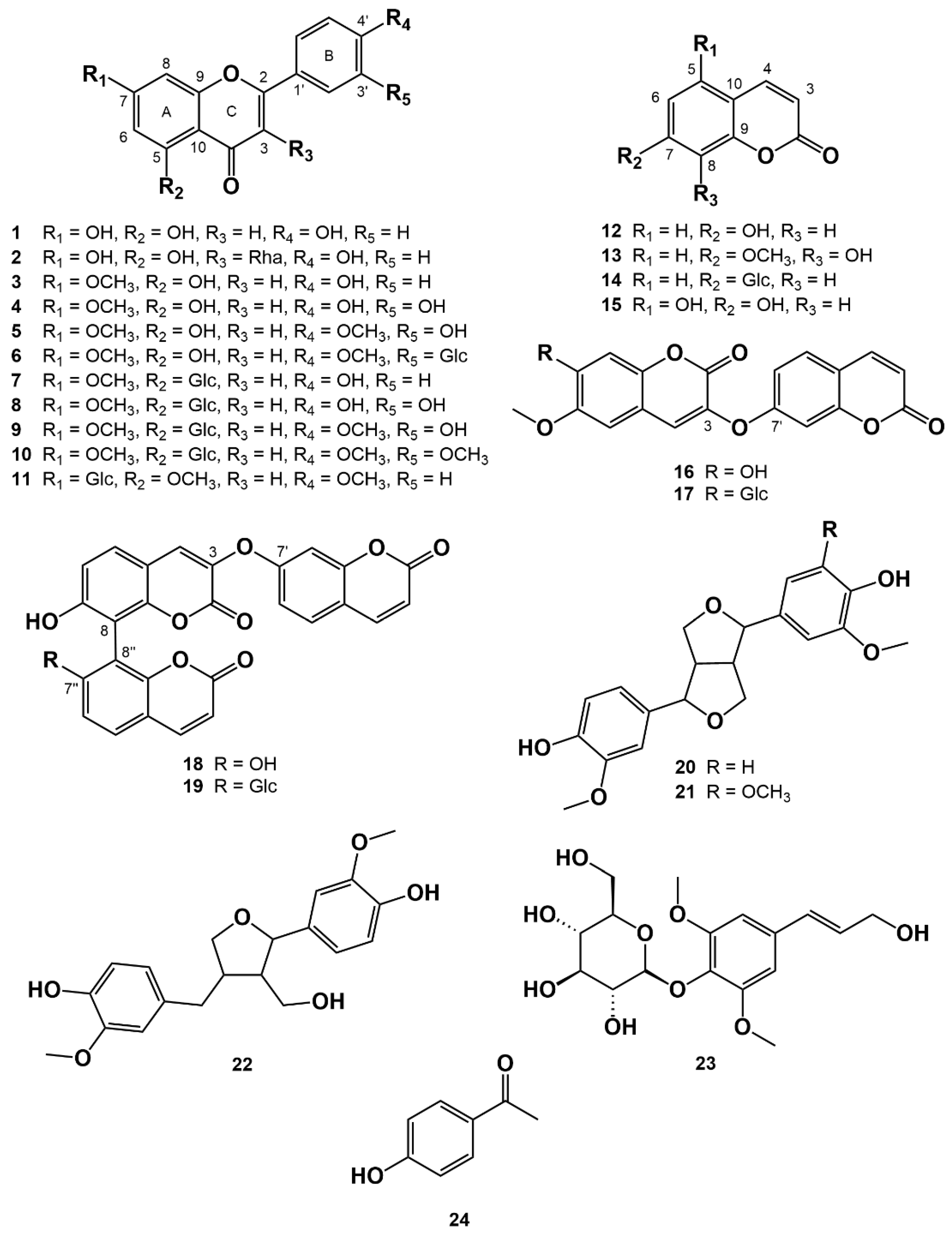
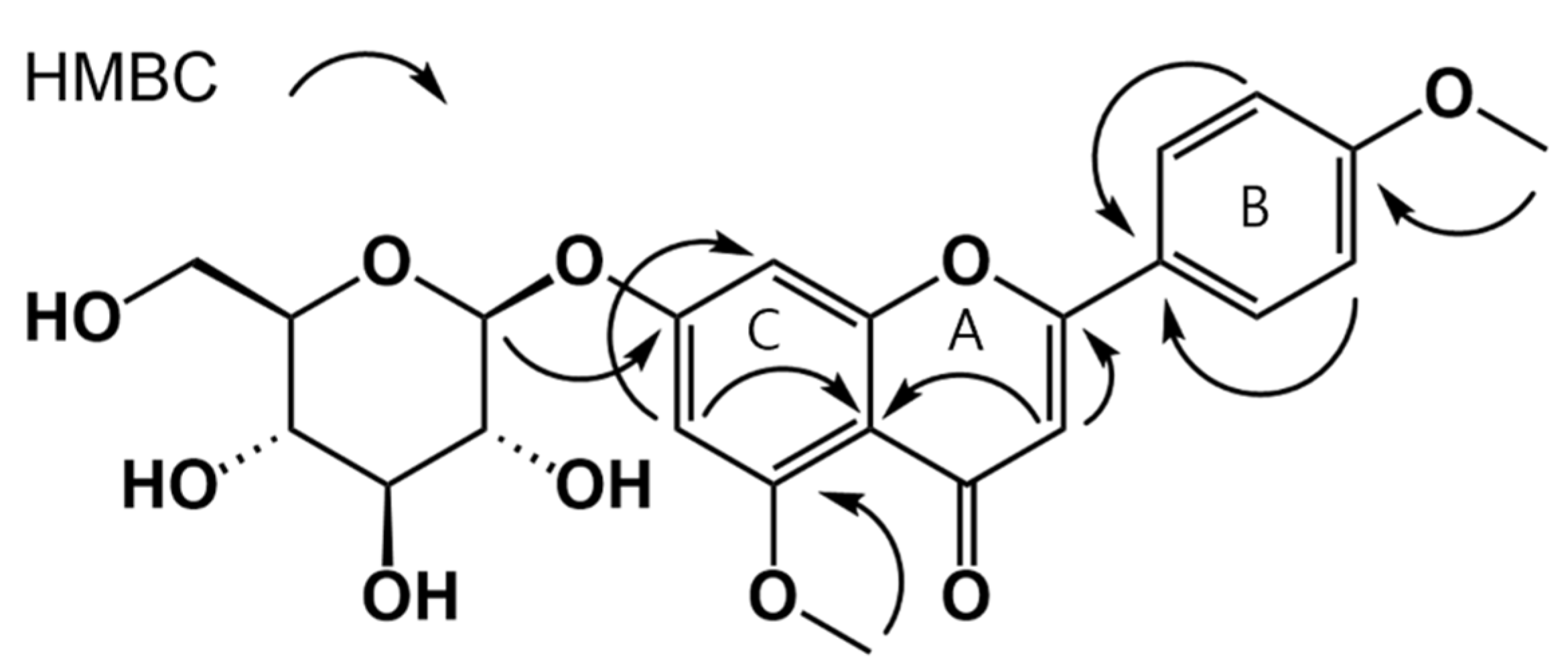
3.1. Isolation of Compounds from W. ganpi Extract and Structural Elucidation of Compound 11
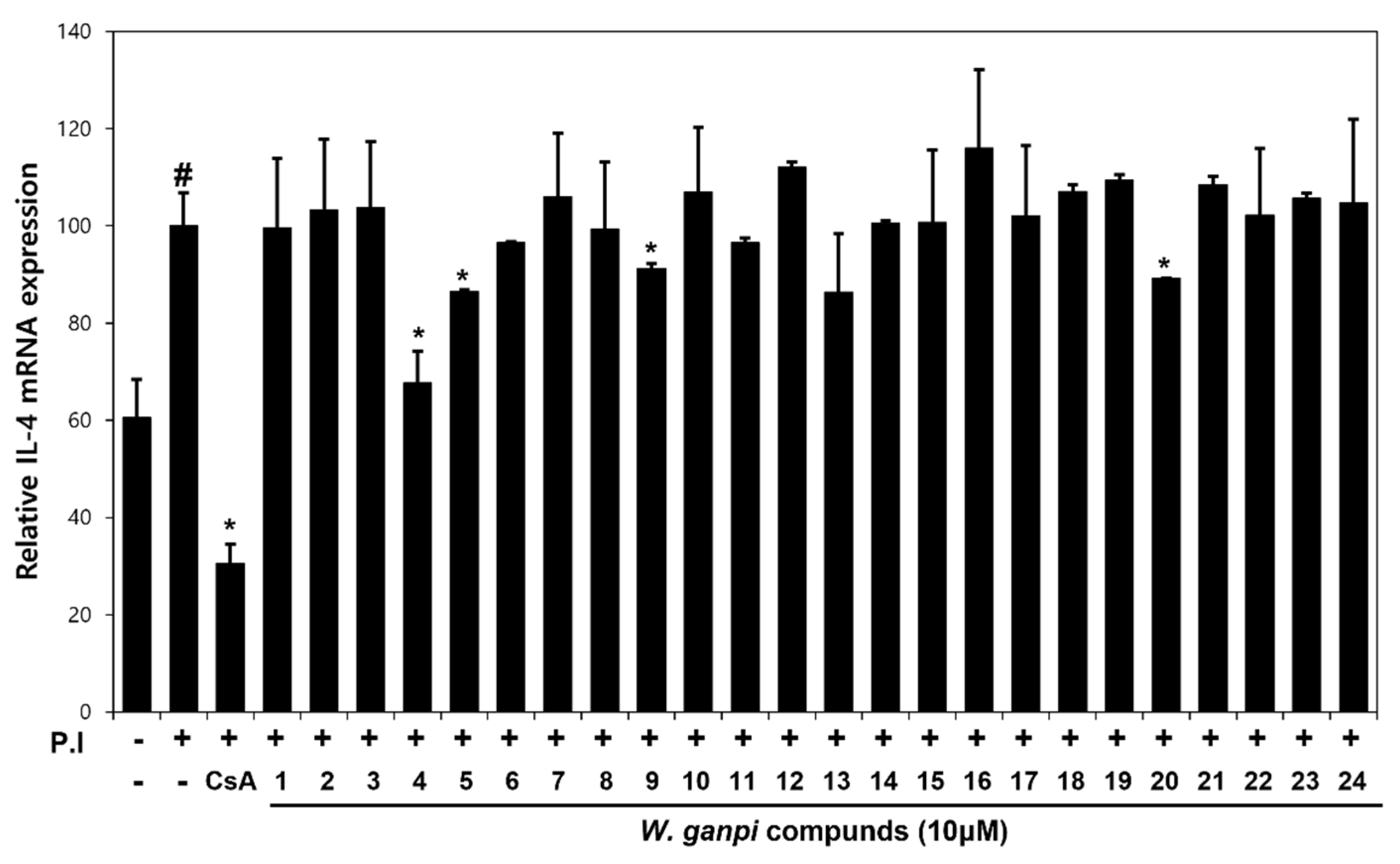
3.2. Screening of Anti-Atopic Activities of Compounds (1–24) Isolated from W. ganpi Extract
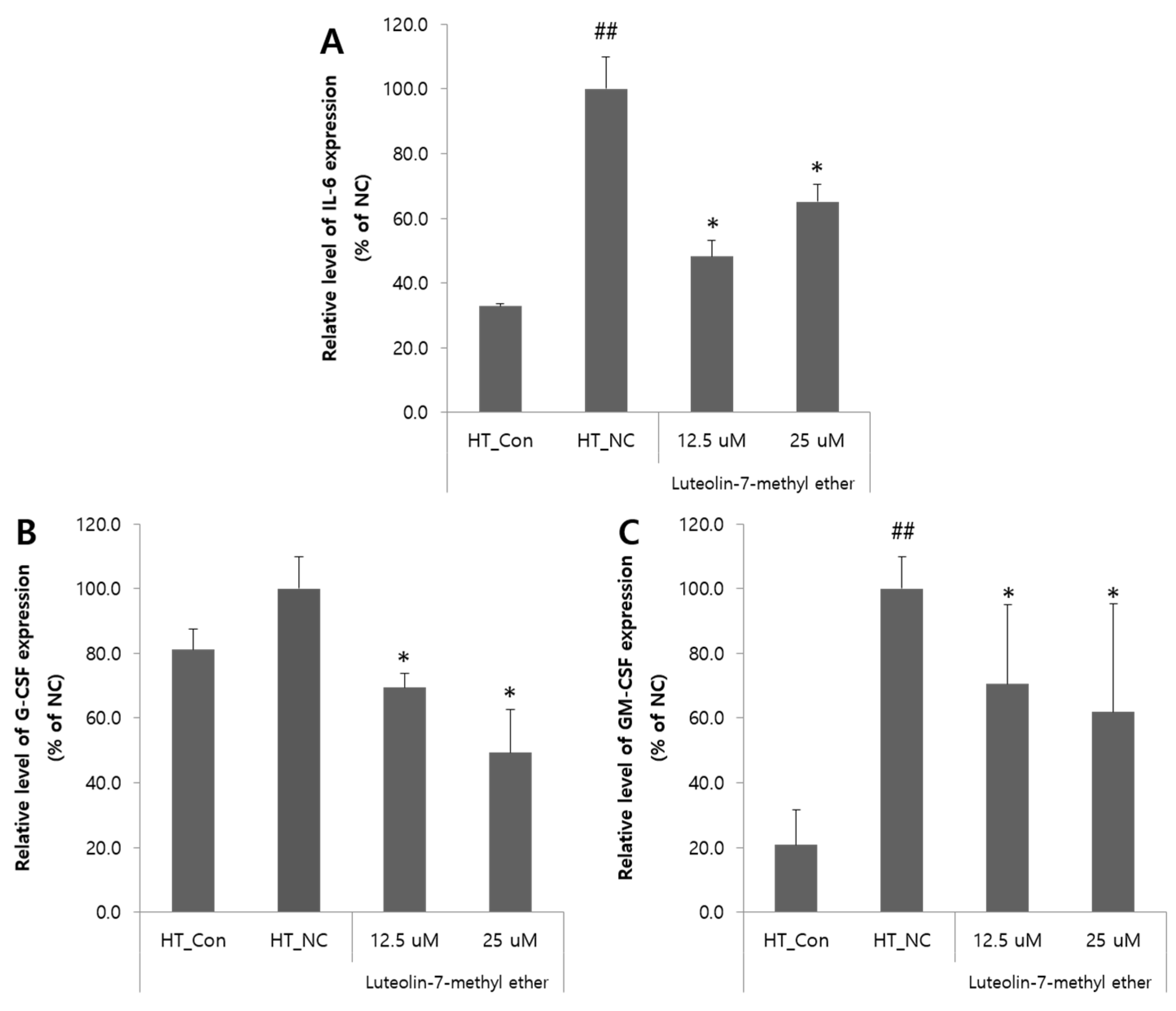
3.3. Effects of Luteolin 7-Methyl Ether (4) on the Expressions of IL-6, GM-CSF, and G-CSF mRNA in HaCaT Cells
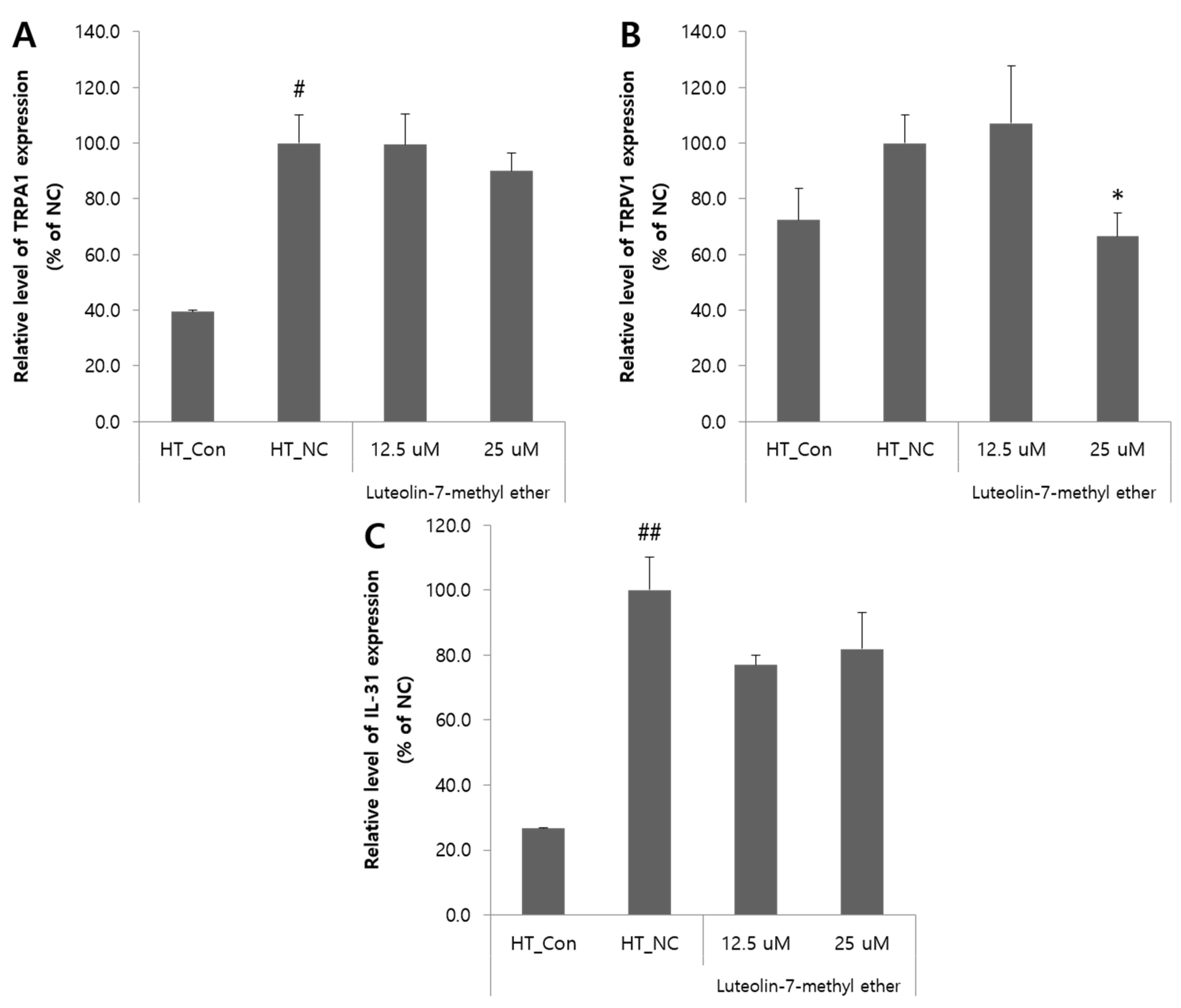
3.4. Effects of Luteolin 7-Methyl Ether (4) on the Pruritus-Related Inflammatory Mediators in HaCaT Cells
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Williams, H.C. Atopic Dermatitis. N. Engl. J. Med. 2005, 352, 2314–2324. [Google Scholar] [CrossRef]
- Nettis, E.; Ortoncelli, M.; Pellacani, G.; Foti, C.; Di Leo, E.; Patruno, C.; Rongioletti, F.; Argenziano, G.; Ferrucci, S.; Macchia, L. A multicenter study on the prevalence of clinical patterns and clinical phenotypes in adult atopic dermatitis. J. Investig. Allergol. Clin. Immunol. 2020, 30, 448–450. [Google Scholar] [CrossRef]
- Harrison, I.P.; Spada, F. Breaking the Itch–Scratch Cycle: Topical Options for the Management of Chronic Cutaneous Itch in Atopic Dermatitis. Medicines 2019, 6, 76. [Google Scholar] [CrossRef] [Green Version]
- Brandt, E.B.; Sivaprasad, U. Th2 Cytokines and Atopic Dermatitis. J. Clin. Cell Immunol. 2011, 2, 110. [Google Scholar] [CrossRef]
- Howell, M.D.; Kim, B.E.; Gao, P.; Grant, A.V.; Boguniewicz, M.; DeBenedetto, A.; Schneider, L.; Beck, L.A.; Barnes, K.C.; Leung, D.Y.M. Cytokine modulation of atopic dermatitis filaggrin skin expression. J. Allergy Clin. Immunol. 2009, 124, R7–R12. [Google Scholar] [CrossRef]
- Feld, M.; Garcia, R.; Buddenkotte, J.; Katayama, S.; Lewis, K.; Muirhead, G.; Hevezi, P.; Plesser, K.; Schrumpf, H.; Krjutskov, K.; et al. The pruritus-and TH2-associated cytokine IL-31 promotes growth of sensory nerves. J. Allergy Clin. Immunol. 2016, 138, 500–508.e524. [Google Scholar] [CrossRef] [Green Version]
- Leung, D.Y.M.; Boguniewicz, M.; Howell, M.D.; Nomura, I.; Hamid, Q.A. New insights into atopic dermatitis. J. Clin. Investig. 2004, 113, 651–657. [Google Scholar] [CrossRef]
- Park, K.-D.; Pak, S.C.; Park, K.-K. The Pathogenetic Effect of Natural and Bacterial Toxins on Atopic Dermatitis. Toxins 2017, 9, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sidbury, R.; Davis, D.M.; Cohen, D.E.; Cordoro, K.M.; Berger, T.G.; Bergman, J.N.; Chamlin, S.L.; Cooper, K.D.; Feldman, S.R.; Hanifin, J.M.; et al. Guidelines of care for the management of atopic dermatitis: Section 3. Management and treatment with phototherapy and systemic agents. J. Am. Acad. Dermatol. Int. 2014, 71, 327–349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fleischer, A.B. Treatment of atopic dermatitis: Role of tacrolimus ointment as a topical noncorticosteroidal therapy. J. Allergy Clin. Immunol. 1999, 104, S126–S130. [Google Scholar] [CrossRef]
- Dattola, A.; Bennardo, L.; Silvestri, M.; Nisticò, S.P. What’s new in the treatment of atopic dermatitis? Dermatol. Ther. (Heidelb) 2019, 32, e12787. [Google Scholar] [CrossRef] [PubMed]
- Middleton, J.M.D.E. Biological Properties of Plant Flavonoids: An Overview. Int. J. Pharmacogn. 1996, 34, 344–348. [Google Scholar] [CrossRef]
- Wang, T.-Y.; Li, Q.; Bi, K.-S. Bioactive flavonoids in medicinal plants: Structure, activity and biological fate. Asian J. Pharm. Sci. 2018, 13, 12–23. [Google Scholar] [CrossRef] [PubMed]
- Maleki, S.J.; Crespo, J.F.; Cabanillas, B. Anti-inflammatory effects of flavonoids. Food Chem. 2019, 299, 125124. [Google Scholar] [CrossRef]
- Serafini, M.; Peluso, I.; Raguzzini, A. Flavonoids as anti-inflammatory agents. Proc. Nutr. Soc. 2010, 69, 273–278. [Google Scholar] [CrossRef] [Green Version]
- Ginwala, R.; Bhavsar, R.; Chigbu, D.G.I.; Jain, P.; Khan, Z.K. Potential Role of Flavonoids in Treating Chronic Inflammatory Diseases with a Special Focus on the Anti-Inflammatory Activity of Apigenin. Antioxidants 2019, 8, 35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heratizadeh, A.; Werfel, T. Anti-inflammatory therapies in atopic dermatitis. Allergy 2016, 71, 1666–1675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, T.; Higa, S.; Hirano, T.; Kotani, M.; Matsumoto, M.; Fujita, A.; Kawase, I. Flavonoids as Potential Anti-Allergic Substances. Curr. Med. Chem.-Anti-Inflamm. Anti-Allergy Agents 2003, 2, 57–65. [Google Scholar] [CrossRef]
- Kim, H.P.; Son, K.H.; Chang, H.W.; Kang, S.S. Anti-inflammatory Plant Flavonoids and Cellular Action Mechanisms. J. Pharmacol. Sci. 2004, 96, 229–245. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.-H.; Park, J.-K.; Choi, J.; Jang, H.; Seol, J.-W. Anti-inflammatory effects of natural flavonoid diosmetin in IL-4 and LPS-induced macrophage activation and atopic dermatitis model. Int. Immunopharmacol. 2020, 89, 107046. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.N.; Shin, S.A.; Choo, G.S.; Kim, H.J.; Park, Y.S.; Kim, B.S.; Kim, S.K.; Cho, S.D.; Nam, J.S.; Choi, C.S.; et al. Anti-inflammatory effect of quercetin and galangin in LPS-stimulated RAW264.7 macrophages and DNCB-induced atopic dermatitis animal models. Int. J. Mol. Med. 2018, 41, 888–898. [Google Scholar] [CrossRef] [PubMed]
- Jung-Won, Y.; Myung-Hoon, Y.; Yong-Shik, K. Growth Environment and Vegetation Structure of Native Habitats of Wikstroemia ganpi (Sieb. et Zucc.) Maxim.1. Korean J. Environ. Ecol. 2014, 28, 331–341. [Google Scholar] [CrossRef]
- Lu, C.-L.; Zhu, L.; Piao, J.-H.; Jiang, J.-G. Chemical compositions extracted from Wikstroemia indica and their multiple activities. Pharm. Biol. 2012, 50, 225–231. [Google Scholar] [CrossRef] [Green Version]
- Borris, R.P.; Blaskó, G.; Cordell, G.A. Ethnopharmacologic and phytochemical studies of the Thymelaeaceae. J. Ethnopharmacol. 1988, 24, 41–91. [Google Scholar] [CrossRef]
- Li, Y.-M.; Zhu, L.; Jiang, J.-G.; Yang, L.; Wang, D.-Y. Bioactive Components and Pharmacological Action of Wikstroemia indica (L.) C. A. Mey and Its Clinical Application. Curr. Pharm. Biotechnol. 2009, 10, 743–752. [Google Scholar] [CrossRef]
- Jegal, J.; Park, N.-J.; Jo, B.-G.; Kim, T.-Y.; Bong, S.-K.; Choi, S.; Paik, J.-H.; Kim, J.-W.; Kim, S.-N.; Yang, M.H. Wikstroemia ganpi Extract Improved Atopic Dermatitis-Like Skin Lesions via Suppression of Interleukin-4 in 2,4-Dinitrochlorobenzene-Induced SKH-1 Hairless Mice. Molecules 2021, 26, 2016. [Google Scholar] [CrossRef] [PubMed]
- Van Loo, P.; De Bruyn, A.; Buděšínský, M. Reinvestigation of the structural assignment of signals in the 1H and 13C NMR spectra of the flavone apigenin. Magn. Reson. Chem. 1986, 24, 879–882. [Google Scholar] [CrossRef]
- Gao, D.-F.; Xu, M.; Yang, C.-R.; Xu, M.; Zhang, Y.-J. Phenolic Antioxidants from the Leaves of Camellia pachyandra Hu. J. Agric. Food Chem. 2010, 58, 8820–8824. [Google Scholar] [CrossRef] [PubMed]
- Talzhanov, N.A.; Sadyrbekov, D.T.; Smagulova, F.M.; Mukanov, R.M.; Raldugin, V.A.; Shakirov, M.M.; Tkachev, A.V.; Atazhanova, G.A.; Tuleuov, B.I.; Adekenov, S.M. Components of Artemisia pontica. Chem. Nat. Compd. 2005, 41, 178–181. [Google Scholar] [CrossRef]
- Saewan, N.; Koysomboon, S.; Chantrapromma, K. Anti-tyrosinase and anti-cancer activities of flavonoids from Blumea balsamifera DC. J. Med. Plant Res. 2011, 5, 1018–1025. [Google Scholar] [CrossRef]
- Devkota, H.P.; Yoshizaki, K.; Yahara, S. Pilloin 5-O-β-D-Glucopyranoside from the Stems of Diplomorpha ganpi. Biosci. Biotechnol. Biochem. 2012, 76, 1555–1557. [Google Scholar] [CrossRef] [Green Version]
- Parveen, N.; Khan, N.U. Luteolin 7,4’-dimethyl ether 3’-glucoside from Gelonium multiflorum. Phytochemistry 1987, 26, 2130–2131. [Google Scholar] [CrossRef]
- Lin, J.-H.; Lin, Y.-T.; Huang, Y.-J.; Wen, K.-C.; Chen, R.-M.; Ueng, T.-H.; Liao, C.-H. Isolation and cytotoxicity of flavonoids from Daphnis Genkwae Flos. J. Food Drug Anal. 2001, 9. [Google Scholar] [CrossRef]
- Xu, W.; Jin, H.; Zhang, W.; Hu, X.; Zhang, W.; Fu, J.; Su, J.; Yan, S.; Shen, Y. Chemical constituents from Daphne pedunculata. Chem. Nat. Compd. 2009, 45, 417–419. [Google Scholar] [CrossRef]
- Chen, D.; Bi, D.; Song, Y.-L.; Tu, P.-F. Flavanoids from the stems of Aquilaria sinensis. Chin. J. Nat. Med. 2012, 10, 287–291. [Google Scholar] [CrossRef]
- Liu, Z.-L.; Tian, X. The components of Cacalia tangutica. Bull. Korean Chem. Soc. 2004, 25, 1078–1080. [Google Scholar] [CrossRef]
- Liang, S.-C.; Ge, G.-B.; Xia, Y.-L.; Zhang, J.-W.; Qi, X.-Y.; Tu, C.-X.; Yang, L. In Vitro Evaluation of the Effect of 7-Methyl Substitution on Glucuronidation of Daphnetin: Metabolic Stability, Isoform Selectivity, and Bioactivity Analysis. J. Pharm. Sci. 2015, 104, 3557–3564. [Google Scholar] [CrossRef]
- Bjerre, J.; Nielsen, E.H.; Bols, M. Hydrolysis of Toxic Natural Glucosides Catalyzed by Cyclodextrin Dicyanohydrins. Eur. J. Org. Chem. 2008, 2008, 745–752. [Google Scholar] [CrossRef]
- Lee, D.; Bhat, K.P.L.; Fong, H.H.S.; Farnsworth, N.R.; Pezzuto, J.M.; Kinghorn, A.D. Aromatase Inhibitors from Broussonetia papyrifera. J. Nat. Prod. 2001, 64, 1286–1293. [Google Scholar] [CrossRef] [PubMed]
- Chaya, N.; Terauchi, K.; Yamagata, Y.; Kinjo, J.; Okabe, H. Antiproliferative Constituents in Plants 14. Coumarins and Acridone Alkaloids from Boenninghausenia japonica NAKAI. Biol. Pharm. Bull. 2004, 27, 1312–1316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kicel, A.; Wolbis, M. Coumarins from the flowers of Trifolium repens. Chem. Nat. Compd. 2012, 48, 130–132. [Google Scholar] [CrossRef]
- Baba, K.; Tabata, Y.; Taniguti, M.; Kozawa, M. Coumarins from Edgeworthia chrysantha. Phytochemistry 1989, 28, 221–225. [Google Scholar] [CrossRef]
- Han, S. Coumarins and Biflavonoids from Edgeworthia chrysantha and Their Glucose Uptake Activity. Master’s Thesis, Seoul National University Graduate School, Seoul, Korea, February 2020. [Google Scholar]
- Páska, C.; Innocenti, G.; Ferlin, M.; Kunvári, M.; László, M. Pinoresinol from Ipomoea Cairica Cell Cultures. Nat. Prod. Lett. 2002, 16, 359–363. [Google Scholar] [CrossRef]
- Deyama, T.; Ikawa, T.; Nishibe, S. The constituents of Eucommia ulmoides OLIV. I. Isolation and structures of three new lignan glycosides. Chem. Pharm. Bull. 1985, 33, 3651–3657. [Google Scholar] [CrossRef] [Green Version]
- Xie, L.-H.; Akao, T.; Hamasaki, K.; Deyama, T.; Hattori, M. Biotransformation of Pinoresinol Diglucoside to Mammalian Lignans by Human Intestinal Microflora, and Isolation of Enterococcus faecalis Strain PDG-1 Responsible for the Transformation of (+)-Pinoresinol to (+)-Lariciresinol. Chem. Pharm. Bull. 2003, 51, 508–515. [Google Scholar] [CrossRef] [Green Version]
- Yang, E.-J.; Kim, S.-I.; Ku, H.-Y.; Lee, D.-S.; Lee, J.-W.; Kim, Y.-S.; Seong, Y.-H.; Song, K.-S. Syringin from stem bark of Fraxinus rhynchophylla protects Aβ(25–35)-induced toxicity in neuronal cells. Arch Pharm. Res. 2010, 33, 531–538. [Google Scholar] [CrossRef]
- Li, J.; Kadota, S.; Kawata, Y.; Hattori, M.; Xu, G.-J.; Namba, T. Constituents of the Roots of Cynanchum bungei DECNE. Isolation and Structures of Four New Glucosides, Bunngeiside-A,-B,-C, and-D. Chem. Pharm. Bull. 1992, 40, 3133–3137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bíró, T.; Tóth, B.I.; Marincsák, R.; Dobrosi, N.; Géczy, T.; Paus, R. TRP channels as novel players in the pathogenesis and therapy of itch. Biochim. Biophys. Acta Mol. Basis Dis. 2007, 1772, 1004–1021. [Google Scholar] [CrossRef] [Green Version]
- Lotz, M.; Jirik, F.; Kabouridis, P.; Tsoukas, C.; Hirano, T.; Kishimoto, T.; Carson, D.A. B cell stimulating factor 2/interleukin 6 is a costimulant for human thymocytes and T lymphocytes. J. Exp. Med. 1988, 167, 1253–1258. [Google Scholar] [CrossRef] [PubMed]
- Girolomoni, G.; Pastore, S. The role of keratinocytes in the pathogenesis of atopic dermatitis. J. Am. Acad. Dermatol. 2001, 45, S25–S28. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound 11 | ||
|---|---|---|
| Position | δH (J in Hz) | δC |
| 2 | 161.02 | |
| 3 | 6.81 (s) | 106.53 |
| 4 | 177.01 | |
| 5 | 6.91 (d, 2.4) | 163.67 |
| 6 | 103.46 | |
| 7 | 158.26 | |
| 8 | 7.09 (d, 2.4) | 96.61 |
| 9 | 158.55 | |
| 10 | 109.31 | |
| 1′ | 122.78 | |
| 2′, 6′ | 8.05 (d, 8.5) | 128.11 |
| 3′, 5′ | 7.12 (d, 8.5) | 114.61 |
| 4′ | 162.15 | |
| Glc 1 | 4.77 (d, 7.6) | 104.10 |
| 2 | 3.52–3.43 | 73.62 |
| 3 | 3.45–3.37 | 75.80 |
| 4 | 3.19–3.12 | 69.98 |
| 5 | 3.52–3.43 | 77.68 |
| 6 | 3.78–3.70 3.19–3.12 | 60.97 |
| 5-OCH3 | 3.90 | 56.14 |
| 4′-OCH3 | 3.86 | 55.60 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jegal, J.; Kim, T.-Y.; Park, N.-J.; Jo, B.-G.; Jo, G.-A.; Choi, H.-S.; Kim, S.-N.; Yang, M.H. Inhibitory Effects of Luteolin 7-Methyl Ether Isolated from Wikstroemia ganpi on Tnf-A/Ifn-Γ Mixture-Induced Inflammation in Human Keratinocyte. Nutrients 2021, 13, 4387. https://doi.org/10.3390/nu13124387
Jegal J, Kim T-Y, Park N-J, Jo B-G, Jo G-A, Choi H-S, Kim S-N, Yang MH. Inhibitory Effects of Luteolin 7-Methyl Ether Isolated from Wikstroemia ganpi on Tnf-A/Ifn-Γ Mixture-Induced Inflammation in Human Keratinocyte. Nutrients. 2021; 13(12):4387. https://doi.org/10.3390/nu13124387
Chicago/Turabian StyleJegal, Jonghwan, Tae-Young Kim, No-June Park, Beom-Geun Jo, Geon-A. Jo, Han-Seok Choi, Su-Nam Kim, and Min Hye Yang. 2021. "Inhibitory Effects of Luteolin 7-Methyl Ether Isolated from Wikstroemia ganpi on Tnf-A/Ifn-Γ Mixture-Induced Inflammation in Human Keratinocyte" Nutrients 13, no. 12: 4387. https://doi.org/10.3390/nu13124387
APA StyleJegal, J., Kim, T.-Y., Park, N.-J., Jo, B.-G., Jo, G.-A., Choi, H.-S., Kim, S.-N., & Yang, M. H. (2021). Inhibitory Effects of Luteolin 7-Methyl Ether Isolated from Wikstroemia ganpi on Tnf-A/Ifn-Γ Mixture-Induced Inflammation in Human Keratinocyte. Nutrients, 13(12), 4387. https://doi.org/10.3390/nu13124387