
Nutrigenomic Effect of Hydroxytyrosol in Vascular Endothelial Cells: A Transcriptomic Profile Analysis

 ,
,  ,
,  ,
,  ,
,  ,
,  ,
,  ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Cell Culture and Treatment
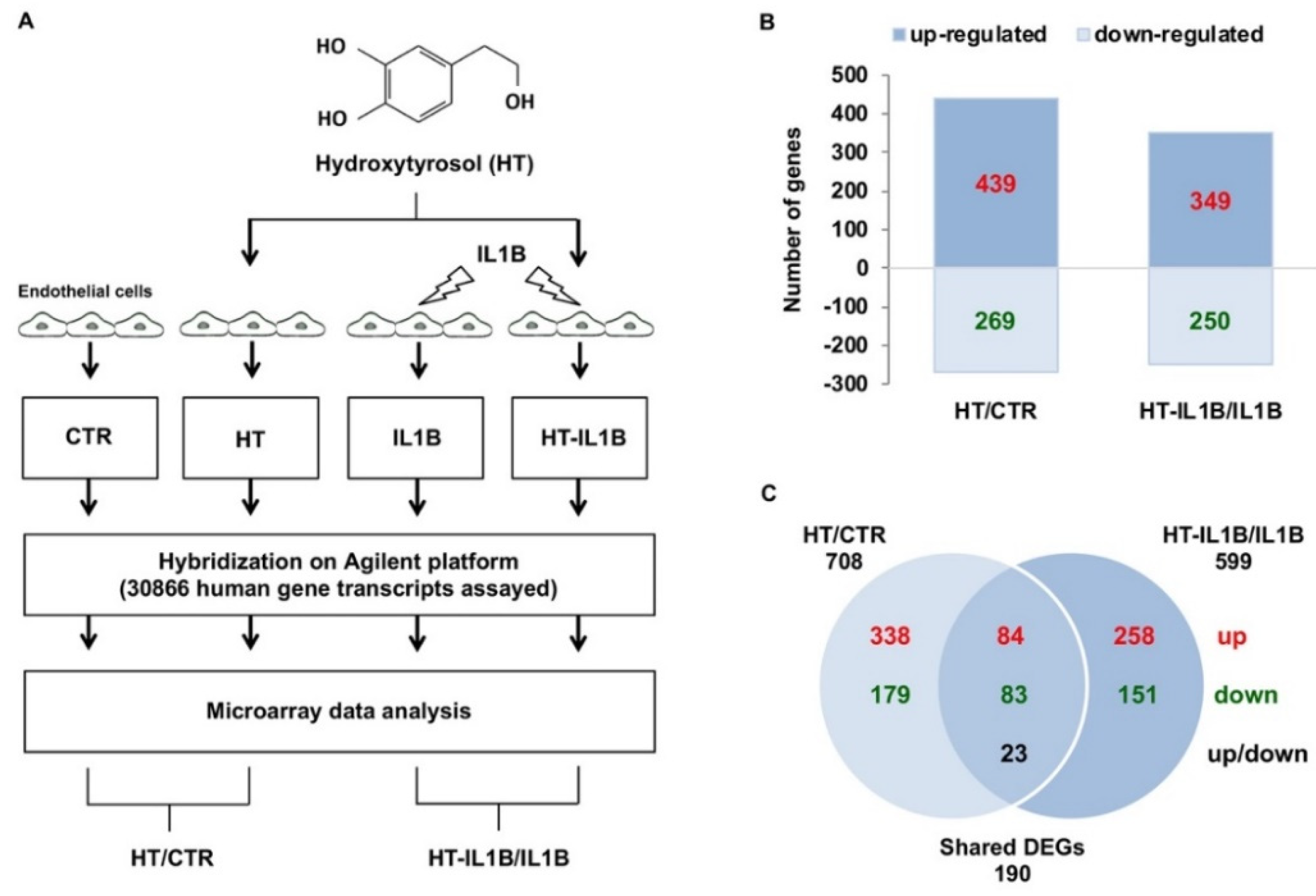
2.3. Experimental Design and RNA Isolation and Analysis
2.4. Microarray Analysis
2.5. Gene Ontology and Pathway Analysis of Transcriptomic Data
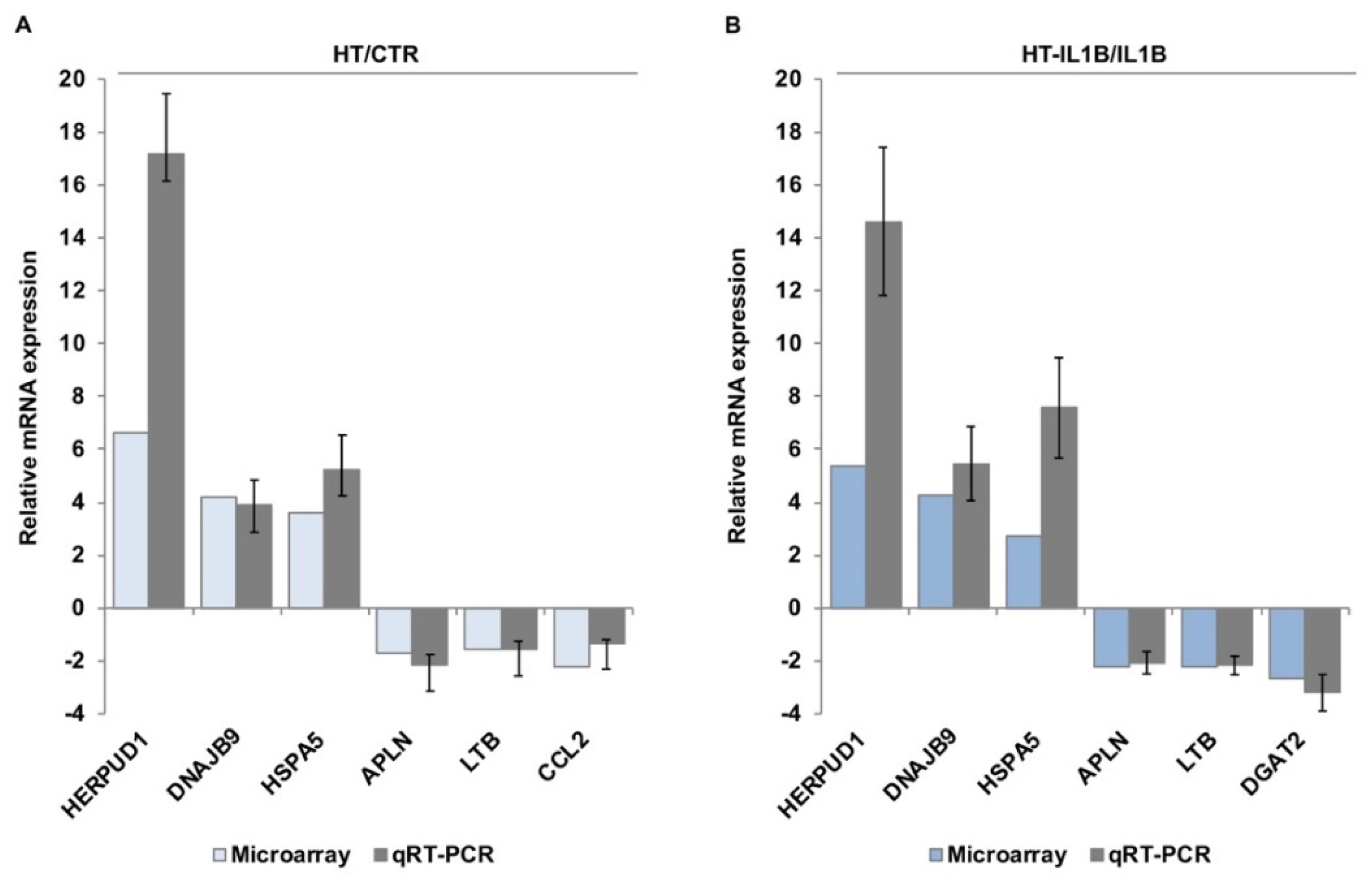
2.6. qRT-PCR of Candidate Target Genes
2.7. Statistical Analysis
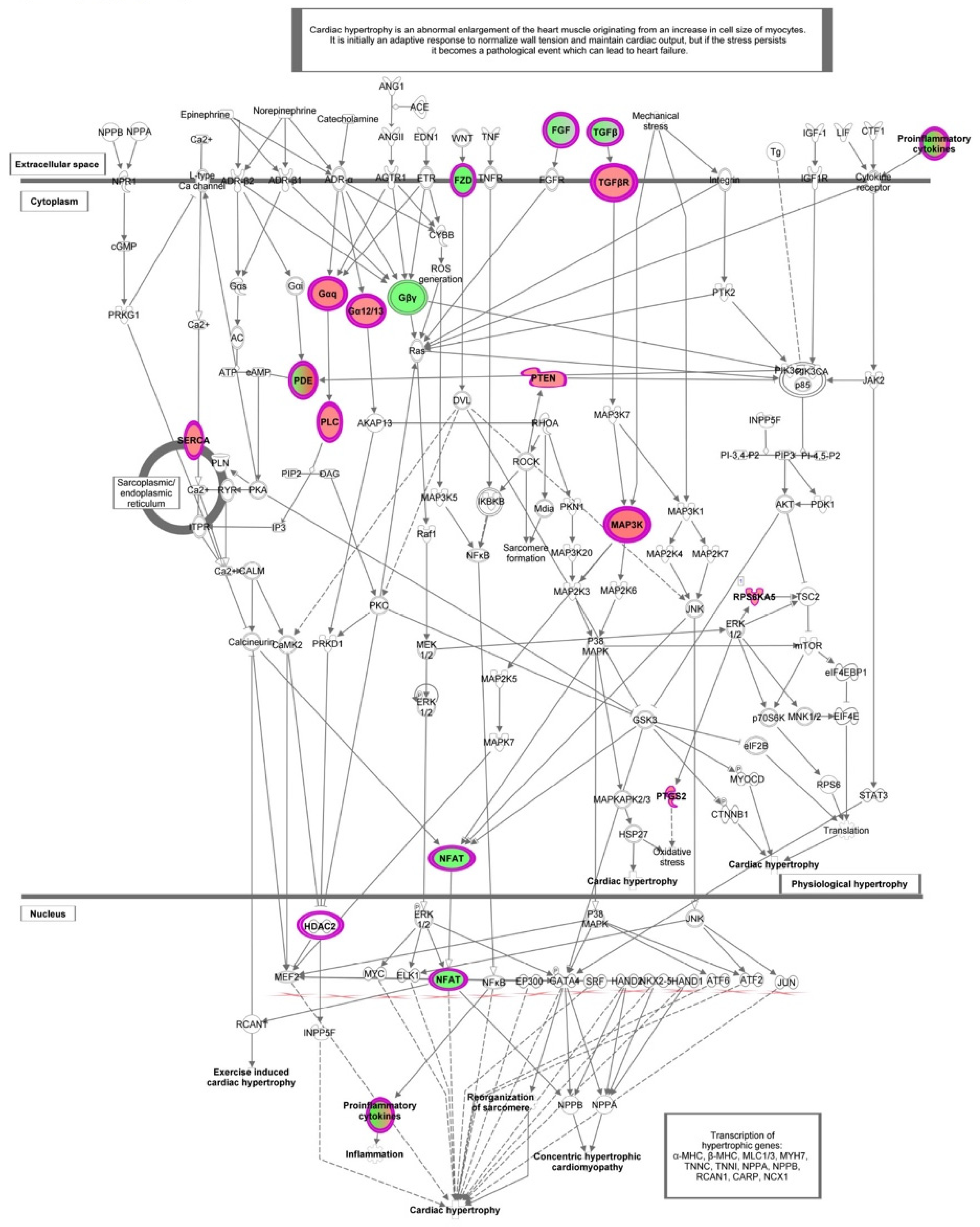
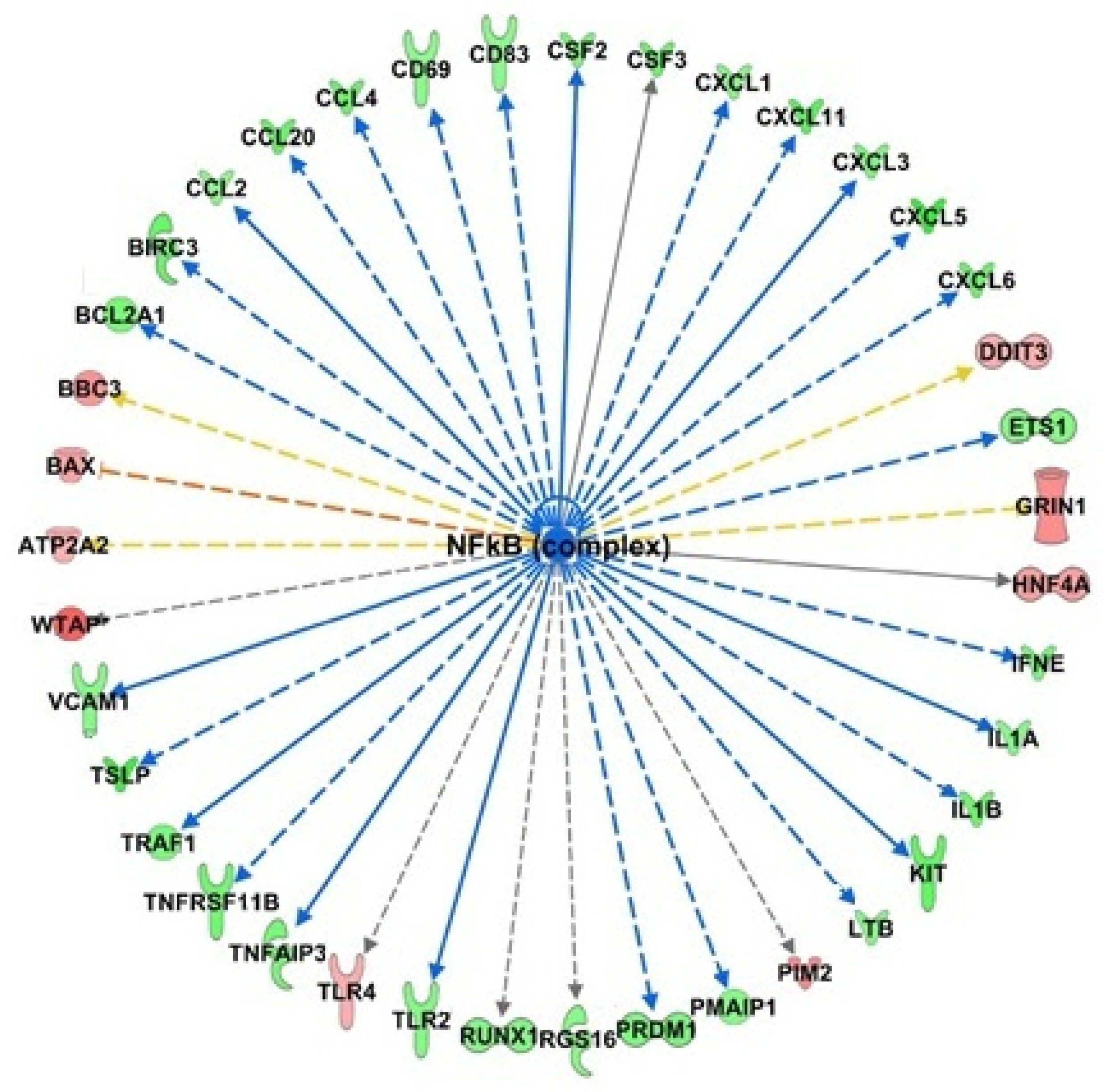
3. Results
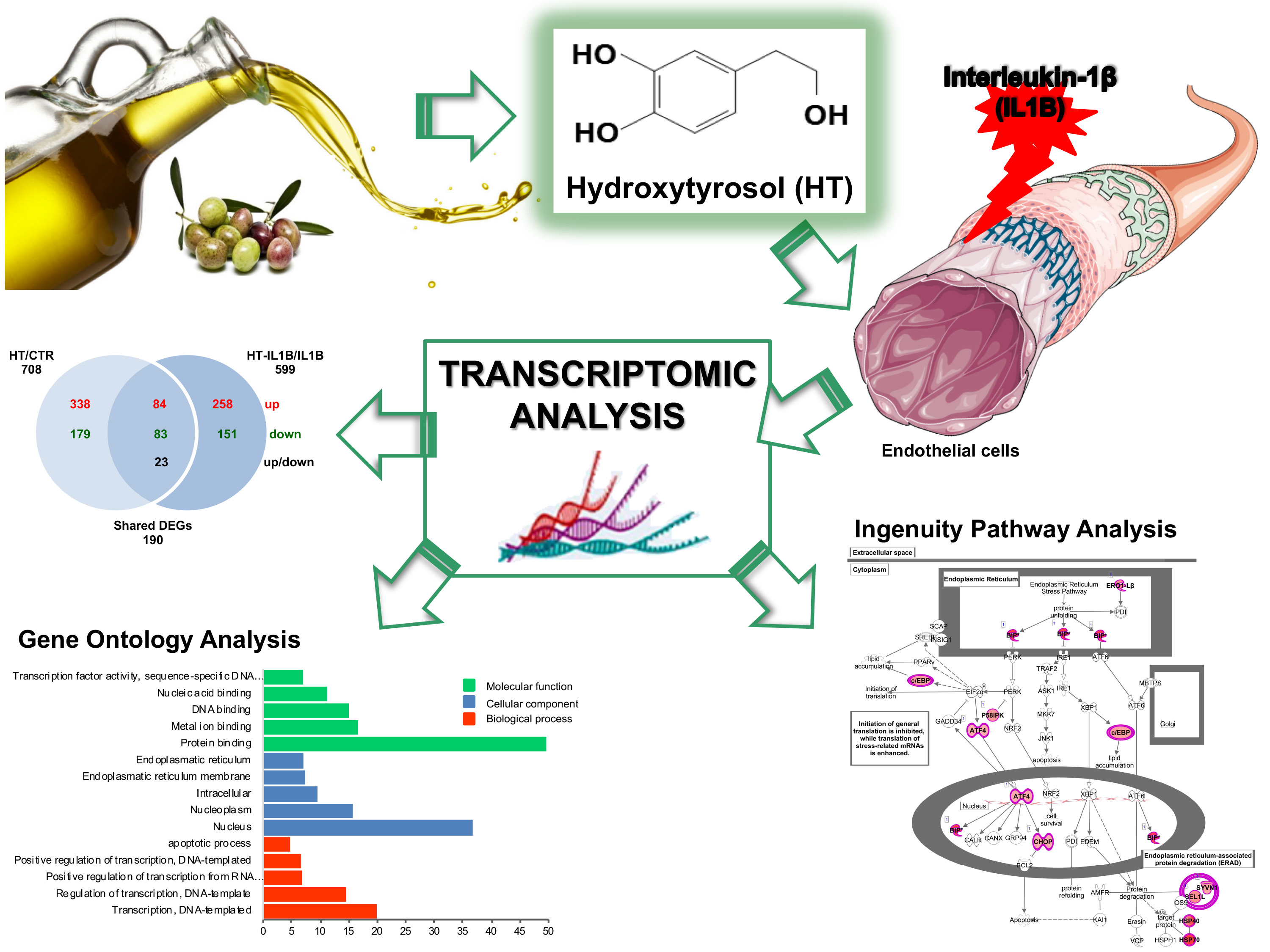
3.1. Gene Expression Profiling of HT Treated Endothelial Cells
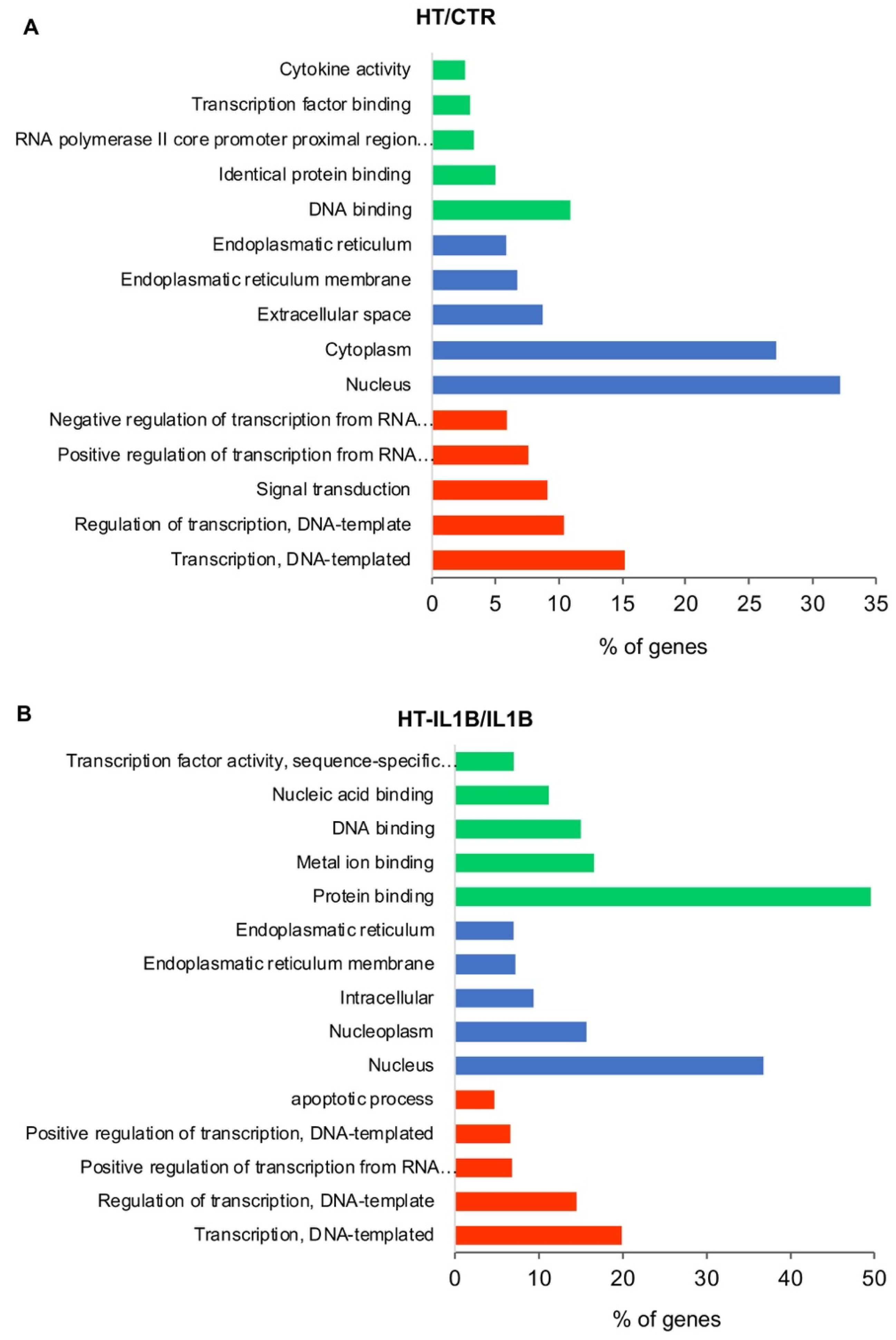
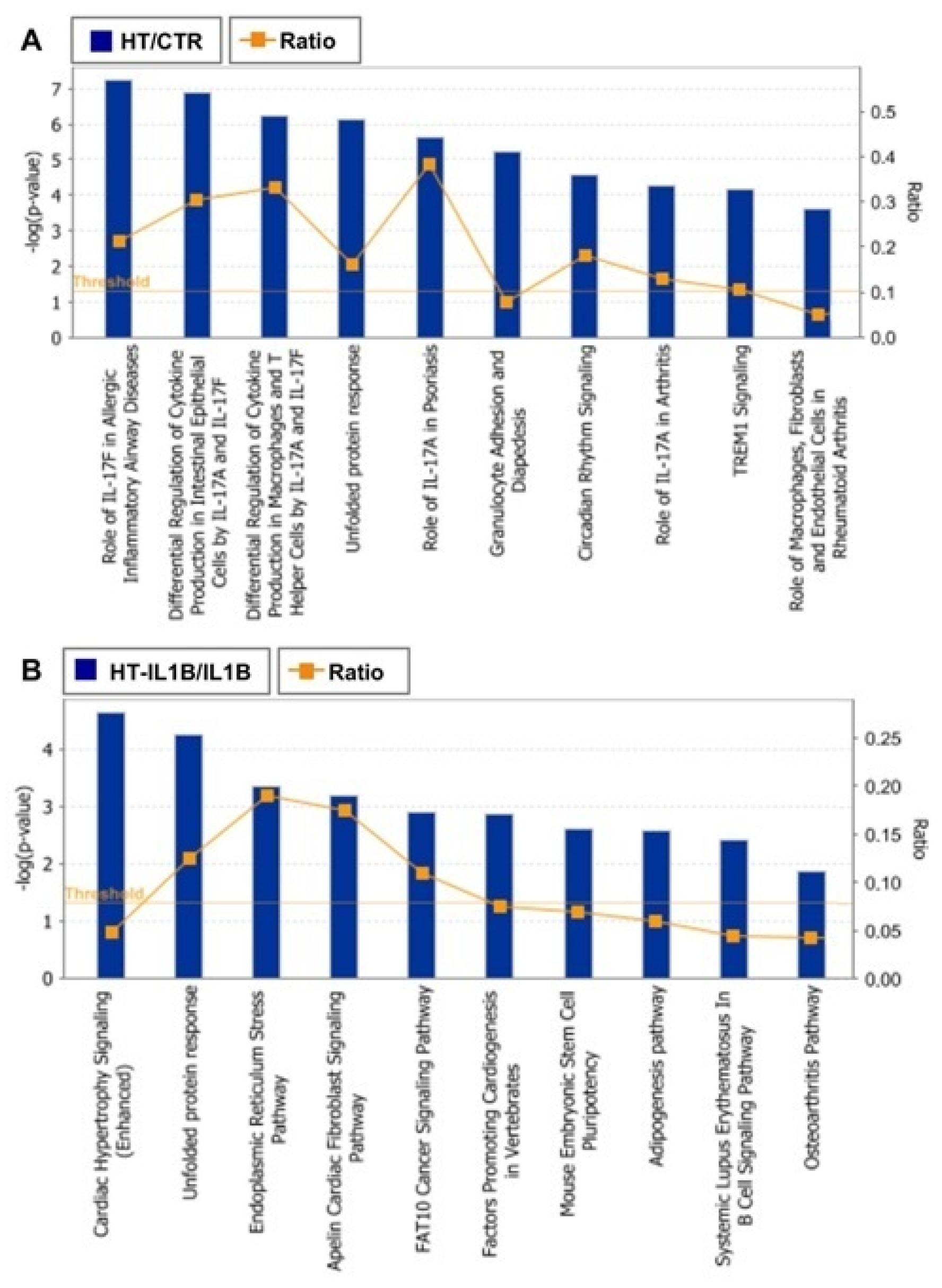
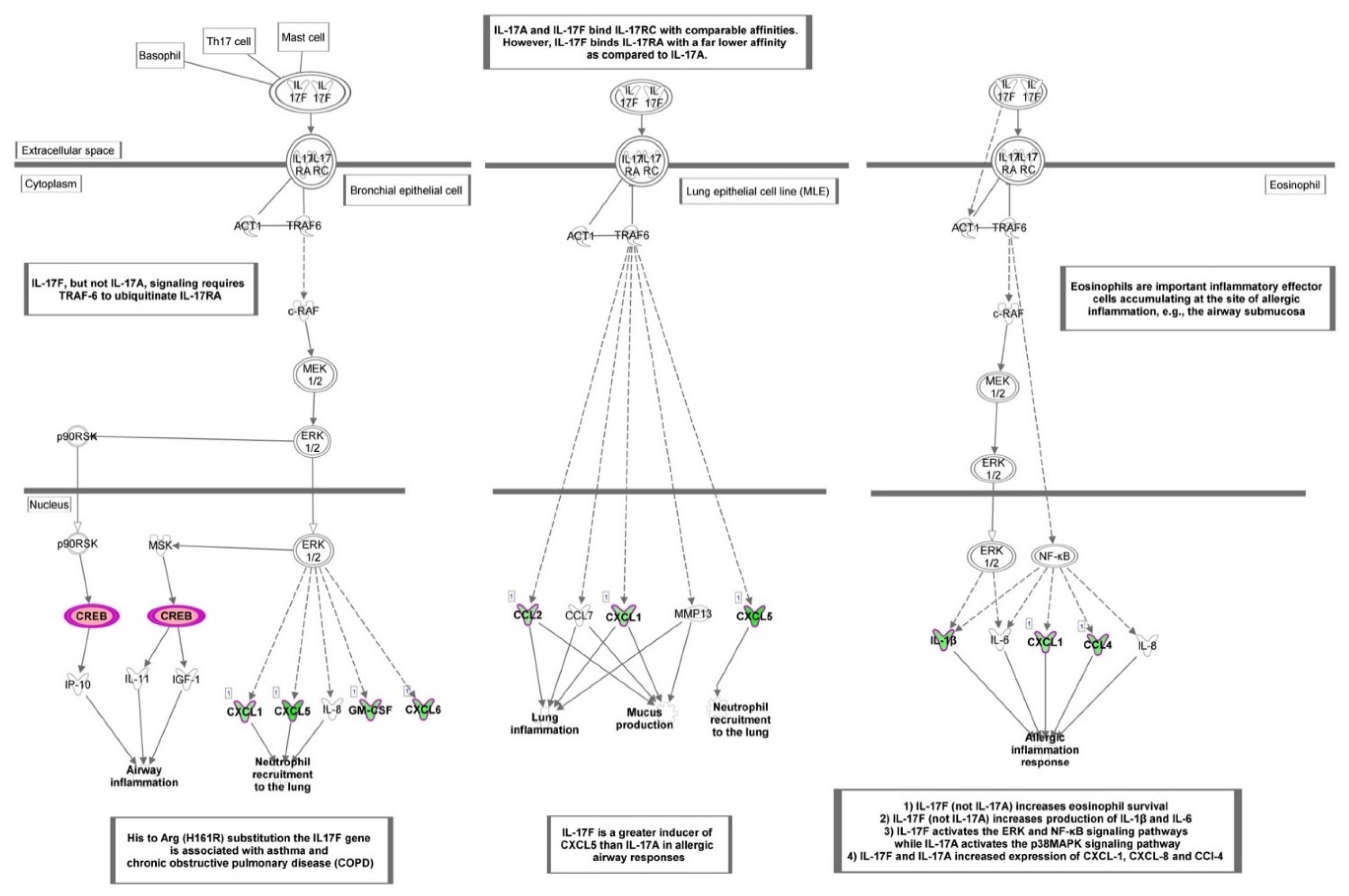
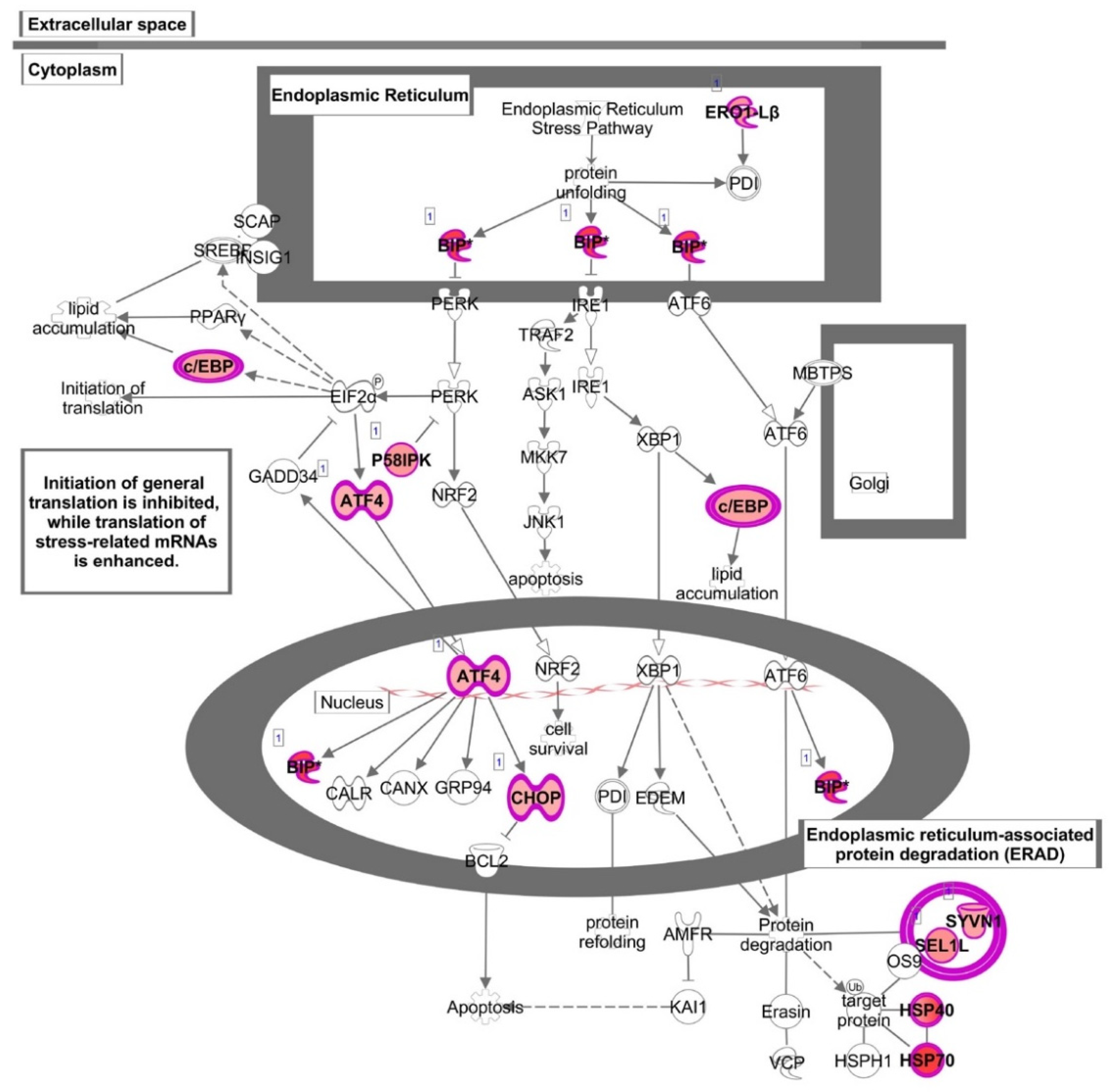
3.2. Functional Enrichment Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Estruch, R.; Ros, E.; Salas-Salvadó, J.; Covas, M.-I.; Corella, D.; Arós, F.; Gómez-Gracia, E.; Ruiz-Gutiérrez, V.; Fiol, M.; Lapetra, J.; et al. Primary Prevention of Cardiovascular Disease with a Mediterranean Diet Supplemented with Extra-Virgin Olive Oil or Nuts. N. Engl. J. Med. 2018, 378, e34. [Google Scholar] [CrossRef] [PubMed]
- Klimova, B.; Novotny, M.; Schlegel, P.; Valis, M. The Effect of Mediterranean Diet on Cognitive Functions in the Elderly Population. Nutrients 2021, 13, 2067. [Google Scholar] [CrossRef] [PubMed]
- Dominguez, L.; Di Bella, G.; Veronese, N.; Barbagallo, M. Impact of Mediterranean Diet on Chronic Non-Communicable Diseases and Longevity. Nutrients 2021, 13, 2028. [Google Scholar] [CrossRef] [PubMed]
- Dinu, M.; Pagliai, G.; Casini, A.; Sofi, F. Mediterranean diet and multiple health outcomes: An umbrella review of meta-analyses of observational studies and randomised trials. Eur. J. Clin. Nutr. 2017, 72, 30–43. [Google Scholar] [CrossRef]
- Marcelino, G.; Hiane, P.A.; Freitas, K.D.C.; Santana, L.F.; Pott, A.; Donadon, J.R.; Guimarães, R.D.C.A. Effects of Olive Oil and Its Minor Components on Cardiovascular Diseases, Inflammation, and Gut Microbiota. Nutrients 2019, 11, 1826. [Google Scholar] [CrossRef]
- D’Angelo, C.; Franceschelli, S.; Quiles, J.L.; Speranza, L. Wide Biological Role of Hydroxytyrosol: Possible Therapeutic and Preventive Properties in Cardio-vascular Diseases. Cells 2020, 9, 1932. [Google Scholar] [CrossRef] [PubMed]
- Covas, M.-I.; Konstantinidou, V.; Fitó, M. Olive Oil and Cardiovascular Health. J. Cardiovasc. Pharmacol. 2009, 54, 477–482. [Google Scholar] [CrossRef]
- EFSA Panel on Dietetic Products. Scientific Opinion on the substantiation of health claims related to polyphenols in olive and protection of LDL particles from oxidative damage (ID 1333, 1638, 1639, 1696, 2865), maintenance of normal blood HDL-cholesterol concentrations (ID 1639), maintenance of normal blood pressure (ID 3781), “anti-inflammatory properties” (ID 1882), “contributes to the upper respiratory tract health” (ID 3468), “can help to maintain a normal function of gastrointestinal tract” (3779), and “contributes to body defences against external agents” (ID 3467) pursuant to Article 13(1) of Regulation (EC) No 1924/2006. EFSA J. 2011, 9, 2033. [Google Scholar]
- Khatana, C.; Saini, N.K.; Chakrabarti, S.; Saini, V.; Sharma, A.; Saini, R.V.; Saini, A. Mechanistic Insights into the Oxidized Low-Density Lipoprotein-Induced Atherosclerosis. Oxid. Med. Cell. Longev. 2020, 2020, 1–14. [Google Scholar] [CrossRef]
- Konstantinidou, V.; Covas, M.-I.; Solà, R.; Fitó, M. Up-to date knowledge on the in vivo transcriptomic effect of the Mediterranean diet in humans. Mol. Nutr. Food Res. 2013, 57, 772–783. [Google Scholar] [CrossRef]
- Camargo, A.; Ruano, J.; Fernandez, J.M.; Parnell, L.D.; Jimenez, A.; Santos-Gonzalez, M.; Marin, C.; Perez-Martinez, P.; Uceda, M.; Lopez-Miranda, J.; et al. Gene expression changes in mononuclear cells in patients with metabolic syndrome after acute intake of phenol-rich virgin olive oil. BMC Genom. 2010, 11, 253. [Google Scholar] [CrossRef] [PubMed]
- Piroddi, M.; Albini, A.; Fabiani, R.; Giovannelli, L.; Luceri, C.; Natella, F.; Rosignoli, P.; Rossi, T.; Taticchi, A.; Servili, M.; et al. Nutrigenomics of extra-virgin olive oil: A review. BioFactors 2016, 43, 17–41. [Google Scholar] [CrossRef] [PubMed]
- Peng, Z.; Shu, B.; Zhang, Y.; Wang, M. Endothelial Response to Pathophysiological Stress. Arterioscler. Thromb. Vasc. Biol. 2019, 39, e233–e243. [Google Scholar] [CrossRef]
- Gimbrone, M.A.; García-Cardeña, G. Endothelial Cell Dysfunction and the Pathobiology of Atherosclerosis. Circ. Res. 2016, 118, 620–636. [Google Scholar] [CrossRef]
- Torres-Peña, J.D.; Rangel-Zuñiga, O.A.; Alcala-Diaz, J.F.; Lopez-Miranda, J.; Delgado-Lista, J. Mediterranean Diet and Endothelial Function: A Review of its Effects at Different Vascular Bed Levels. Nutrients 2020, 12, 2212. [Google Scholar] [CrossRef]
- Shannon, O.M.; Mendes, I.; Köchl, C.; Mazidi, M.; Ashor, A.W.; Rubele, S.; Minihane, A.-M.; Mathers, J.C.; Siervo, M. Mediterranean Diet Increases Endothelial Function in Adults: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. J. Nutr. 2020, 150, 1151–1159. [Google Scholar] [CrossRef] [PubMed]
- Carluccio, M.A.; Massaro, M.; Scoditti, E.; De Caterina, R. Vasculoprotective potential of olive oil components. Mol. Nutr. Food Res. 2007, 51, 1225–1234. [Google Scholar] [CrossRef] [PubMed]
- Carluccio, M.A.; Ancora, M.A.; Massaro, M.; Carluccio, M.; Scoditti, E.; Distante, A.; Storelli, C.; De Caterina, R. Homocysteine induces VCAM-1 gene expression through NF-kappaB and NAD(P)H oxidase activation: Protective role of Mediterranean diet polyphenolic antioxidants. Am. J. Physiol. Heart Circ. Physiol. 2007, 293, H2344–H2354. [Google Scholar] [CrossRef] [PubMed]
- Carluccio, M.A.; Siculella, L.; Ancora, M.A.; Massaro, M.; Scoditti, E.; Storelli, C.; Visioli, F.; Distante, A.; de Caterina, R. Olive oil and red wine antioxidant polyphenols inhibit endothelial activation: Antiatherogenic properties of Mediterranean diet phytochemicals. Arterioscler. Thromb. Vasc. Biol. 2003, 23, 622–629. [Google Scholar] [CrossRef]
- Calabriso, N.; Gnoni, A.; Stanca, E.; Cavallo, A.; Damiano, F.; Siculella, L.; Carluccio, M.A. Hydroxytyrosol Ameliorates Endothelial Function under Inflammatory Conditions by Preventing Mitochondrial Dysfunction. Oxid. Med. Cell. Longev. 2018, 2018, 1–14. [Google Scholar] [CrossRef]
- Calabriso, N.; Massaro, M.; Scoditti, E.; D’Amore, S.; Gnoni, A.; Pellegrino, M.; Storelli, C.; De Caterina, R.; Palasciano, G.; Carluccio, M.A. Extra virgin olive oil rich in polyphenols modulates VEGF-induced angiogenic responses by preventing NADPH oxidase activity and expression. J. Nutr. Biochem. 2015, 28, 19–29. [Google Scholar] [CrossRef]
- Carpi, S.; Scoditti, E.; Massaro, M.; Polini, B.; Manera, C.; Digiacomo, M.; Esposito Salsano, J.; Poli, G.; Tuccinardi, T.; Doccini, S.; et al. The Extra-Virgin Olive Oil Polyphenols Oleocanthal and Oleacein Counteract Inflammation-Related Gene and miRNA Expression in Adipocytes by Attenuating NF-κB Activation. Nutrients 2019, 11, 2855. [Google Scholar] [CrossRef] [PubMed]
- Scoditti, E.; Calabriso, N.; Massaro, M.; Pellegrino, M.; Storelli, C.; Martines, G.; De Caterina, R.; Carluccio, M.A. Mediterranean diet polyphenols reduce inflammatory angiogenesis through MMP-9 and COX-2 inhibition in human vascular endothelial cells: A potentially protective mechanism in atherosclerotic vascular disease and cancer. Arch. Biochem. Biophys. 2012, 527, 81–89. [Google Scholar] [CrossRef]
- Scoditti, E.; Capurso, C.; Capurso, A.; Massaro, M. Vascular effects of the Mediterranean diet—Part II: Role of omega-3 fatty acids and olive oil polyphenols. Vasc. Pharmacol. 2014, 63, 127–134. [Google Scholar] [CrossRef]
- Scoditti, E.; Nestola, A.; Massaro, M.; Calabriso, N.; Storelli, C.; De Caterina, R.; Carluccio, M.A. Hydroxytyrosol suppresses MMP-9 and COX-2 activity and expression in activated human monocytes via PKCα and PKCβ1 inhibition. Atherosclerosis 2014, 232, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Herrera-Marcos, L.V.; Lou-Bonafonte, J.M.; Arnal, C.; Navarro, M.A.; Osada, J. Transcriptomics and the Mediterranean Diet: A Systematic Review. Nutrients 2017, 9, 472. [Google Scholar] [CrossRef]
- D’Amore, S.; Vacca, M.; Cariello, M.; Graziano, G.; D’Orazio, A.; Salvia, R.; Sasso, R.C.; Sabbà, C.; Palasciano, G.; Moschetta, A. Genes and miRNA expression signatures in peripheral blood mononuclear cells in healthy subjects and patients with metabolic syndrome after acute intake of extra virgin olive oil. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2016, 1861, 1671–1680. [Google Scholar] [CrossRef]
- De Gonzalo-Calvo, D.; Vea, A.; Bär, C.; Fiedler, J.; Couch, L.S.; Brotons, C.; Llorente-Cortes, V.; Thum, T. Circulating non-coding RNAs in biomarker-guided cardiovascular therapy: A novel tool for personalized medicine? Eur. Heart J. 2019, 40, 1643–1650. [Google Scholar] [CrossRef] [PubMed]
- De Santis, S.; Cariello, M.; Piccinin, E.; Sabbà, C.; Moschetta, A. Extra Virgin Olive Oil: Lesson from Nutrigenomics. Nutrients 2019, 11, 2085. [Google Scholar] [CrossRef]
- Carluccio, M.A.; Massaro, M.; Bonfrate, C.; Siculella, L.; Maffia, M.; Nicolardi, G.; Distante, A.; Storelli, C.; De Caterina, R. Oleic acid inhibits endothelial activation: A direct vascular antiatherogenic mechanism of a nutritional component in the mediterranean diet. Arterioscler. Thromb. Vasc. Biol. 1999, 19, 220–228. [Google Scholar] [CrossRef]
- Massaro, M.; Martinelli, R.; Gatta, V.; Scoditti, E.; Pellegrino, M.; Carluccio, M.A.; Calabriso, N.; Buonomo, T.; Stuppia, L.; Storelli, C.; et al. Correction: Transcriptome-Based Identification of New Anti-Anti-Inflammatory and Vasodilating Properties of the n-3 Fatty Acid Docosahexaenoic Acid in Vascular Endothelial Cell Under Proinflammatory Conditions. PLoS ONE 2016, 11, e0154069. [Google Scholar] [CrossRef]
- Buonomo, T.; Carraresi, L.; Rossini, M.; Martinelli, R. Involvement of aryl hydrocarbon receptor signaling in the development of small cell lung cancer induced by HPV E6/E7 oncoproteins. J. Transl. Med. 2011, 9, 2. [Google Scholar] [CrossRef]
- Hazas, M.-C.L.D.L.; Martin-Hernández, R.; Crespo, M.C.; Tomé-Carneiro, J.; del Pozo-Acebo, L.; Ruiz-Roso, M.B.; Escola-Gil, J.C.; Osada, J.; Portillo, M.P.; Martinez, J.A.; et al. Identification and validation of common molecular targets of hydroxytyrosol. Food Funct. 2019, 10, 4897–4910. [Google Scholar] [CrossRef] [PubMed]
- Figueira, I.; Tavares, L.; Jardim, C.E.C.G.; Costa, I.; Terrasso, A.; Almeida, A.F.; Govers, C.; Mes, J.J.; Gardner, R.; Becker, J.; et al. Blood–brain barrier transport and neuroprotective potential of blackberry-digested polyphenols: An in vitro study. Eur. J. Nutr. 2017, 58, 113–130. [Google Scholar] [CrossRef] [PubMed]
- McGeachy, M.J.; Cua, D.J.; Gaffen, S.L. The IL-17 Family of Cytokines in Health and Disease. Immunity 2019, 50, 892–906. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Crother, T.R.; Arditi, M. Emerging Role of IL-17 in Atherosclerosis. J. Innate Immun. 2010, 2, 325–333. [Google Scholar] [CrossRef]
- Collins, T.; Read, M.; Neish, A.S.; Whitley, M.Z.; Thanos, D.; Maniatis, T. Transcriptional regulation of endothelial cell adhesion molecules: NF-kappa B and cytokine-inducible enhancers. FASEB J. 1995, 9, 899–909. [Google Scholar] [CrossRef]
- Echeverría, F.; Ortiz, M.; Valenzuela, R.; Videla, L.A. Hydroxytyrosol and Cytoprotection: A Projection for Clinical Interventions. Int. J. Mol. Sci. 2017, 18, 930. [Google Scholar] [CrossRef]
- LaRussa, T.; Oliverio, M.; Suraci, E.; Greco, M.; Placida, R.; Gervasi, S.; Marasco, R.; Imeneo, M.; Paolino, D.; Tucci, L.; et al. Oleuropein Decreases Cyclooxygenase-2 and Interleukin-17 Expression and Attenuates Inflammatory Damage in Colonic Samples from Ulcerative Colitis Patients. Nutrients 2017, 9, 391. [Google Scholar] [CrossRef] [PubMed]
- Acosta, E.H.; Pérez, J.A.S.; Arjona, J.A.; Visioli, F. An olive polyphenol-based nutraceutical improves cutaneous manifestations of psoriasis in humans. Pharmanutrition 2018, 6, 45. [Google Scholar] [CrossRef]
- Hetz, C.; Zhang, K.; Kaufman, R.J. Mechanisms, regulation and functions of the unfolded protein response. Nat. Rev. Mol. Cell Biol. 2020, 21, 421–438. [Google Scholar] [CrossRef]
- Lin, J.H.; Walter, P.; Yen, T.S. Endoplasmic reticulum stress in disease pathogenesis. Annu. Rev. Pathol. 2008, 3, 399–425. [Google Scholar] [CrossRef] [PubMed]
- Ren, J.; Bi, Y.; Sowers, J.R.; Hetz, C.; Zhang, Y. Endoplasmic reticulum stress and unfolded protein response in cardiovascular diseases. Nat. Rev. Cardiol. 2021, 18, 499–521. [Google Scholar] [CrossRef]
- Hetz, C.; Papa, F.R. The Unfolded Protein Response and Cell Fate Control. Mol. Cell 2018, 69, 169–181. [Google Scholar] [CrossRef]
- Amodio, G.; Moltedo, O.; Faraonio, R.; Remondelli, P. Targeting the Endoplasmic Reticulum Unfolded Protein Response to Counteract the Oxidative Stress-Induced Endothelial Dysfunction. Oxid. Med. Cell. Longev. 2018, 2018, 1–13. [Google Scholar] [CrossRef]
- Birukova, A.A.; Singleton, P.A.; Gawlak, G.; Tian, X.; Mirzapoiazova, T.; Mambetsariev, B.; Dubrovskyi, O.; Oskolkova, O.V.; Bochkov, V.N.; Birukov, K.G. GRP78 is a novel receptor initiating a vascular barrier protective response to oxidized phospholipids. Mol. Biol. Cell 2014, 25, 2006–2016. [Google Scholar] [CrossRef]
- Sarcinelli, C.; Dragic, H.; Piecyk, M.; Barbet, V.; Duret, C.; Barthelaix, A.; Ferraro-Peyret, C.; Fauvre, J.; Renno, T.; Chaveroux, C.; et al. ATF4-Dependent NRF2 Transcriptional Regulation Promotes Antioxidant Protection during Endoplasmic Reticulum Stress. Cancers 2020, 12, 569. [Google Scholar] [CrossRef] [PubMed]
- Zrelli, H.; Matsuoka, M.; Kitazaki, S.; Araki, M.; Kusunoki, M.; Zarrouk, M.; Miyazaki, H. Hydroxytyrosol Induces Proliferation and Cytoprotection against Oxidative Injury in Vascular Endothelial Cells: Role of Nrf2 Activation and HO-1 Induction. J. Agric. Food Chem. 2011, 59, 4473–4482. [Google Scholar] [CrossRef] [PubMed]
- Putics, A.; Végh, E.M.; Csermely, P.; Sőti, C. Resveratrol Induces the Heat-Shock Response and Protects Human Cells from Severe Heat Stress. Antioxid. Redox Signal. 2008, 10, 65–76. [Google Scholar] [CrossRef]
- Valdés, A.; Sullini, G.; Ibáñez, E.; Cifuentes, A.; García-Cañas, V. Rosemary polyphenols induce unfolded protein response and changes in cholesterol metabolism in colon cancer cells. J. Funct. Foods 2015, 15, 429–439. [Google Scholar] [CrossRef]
- Barabutis, N. Unfolded Protein Response supports endothelial barrier function. Biochimie 2019, 165, 206–209. [Google Scholar] [CrossRef] [PubMed]
- Giordano, E.; Davalos, A.; Nicod, N.; Visioli, F. Hydroxytyrosol attenuates tunicamycin-induced endoplasmic reticulum stress in human hepatocarcinoma cells. Mol. Nutr. Food Res. 2013, 58, 954–962. [Google Scholar] [CrossRef]
- Leitman, J.; Shenkman, M.; Gofman, Y.; Shtern, N.O.; Ben-Tal, N.; Hendershot, L.M.; Lederkremer, G.Z. Herp coordinates compartmentalization and recruitment of HRD1 and misfolded proteins for ERAD. Mol. Biol. Cell 2014, 25, 1050–1060. [Google Scholar] [CrossRef] [PubMed]
- Schulze, A.; Standera, S.; Buerger, E.; Kikkert, M.; van Voorden, S.; Wiertz, E.; Koning, F.; Kloetzel, P.-M.; Seeger, M. The Ubiquitin-domain Protein HERP forms a Complex with Components of the Endoplasmic Reticulum Associated Degradation Pathway. J. Mol. Biol. 2005, 354, 1021–1027. [Google Scholar] [CrossRef] [PubMed]
- Yan, S.; Zheng, C.; Chen, Z.-Q.; Liu, R.; Li, G.-G.; Hu, W.-K.; Pei, H.; Li, B. Expression of Endoplasmic Reticulum Stress-Related Factors in the Retinas of Diabetic Rats. Exp. Diabetes Res. 2011, 2012, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Giblin, M.J.; Smith, T.E.; Winkler, G.; Pendergrass, H.A.; Kim, M.J.; Capozzi, M.E.; Yang, R.; McCollum, G.W.; Penn, J.S. Nuclear factor of activated T-cells (NFAT) regulation of IL-1beta-induced retinal vascular inflammation. Biochim. Biophys. Acta Mol. Basis Dis. 2021, 1867, 166238. [Google Scholar] [CrossRef]
- Cai, Y.; Yao, H.; Sun, Z.; Wang, Y.; Zhao, Y.; Wang, Z.; Li, L. Role of NFAT in the Progression of Diabetic Atherosclerosis. Front. Cardiovasc. Med. 2021, 8, 140. [Google Scholar] [CrossRef]
- Dolinsky, V.W.; Soltys, C.-L.M.; Rogan, K.J.; Chan, A.Y.M.; Nagendran, J.; Wang, S.; Dyck, J.R.B. Resveratrol prevents pathological but not physiological cardiac hypertrophy. J. Mol. Med. 2014, 93, 413–425. [Google Scholar] [CrossRef]
- Motojima, H.; Villareal, M.O.; Iijima, R.; Han, J.; Isoda, H. Acteoside inhibits type Iota allergy through the down-regulation of Ca/NFAT and JNK MAPK signaling pathways in basophilic cells. J. Nat. Med. 2013, 67, 790–798. [Google Scholar] [CrossRef]
- Sung, S.; Kwon, D.; Um, E.; Kim, B. Could Polyphenols Help in the Control of Rheumatoid Arthritis? Molecules 2019, 24, 1589. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Jackson-Weaver, O.; Xu, J. The TGFbeta superfamily in cardiac dysfunction. Acta Biochim. Biophys. Sin. 2018, 50, 323–335. [Google Scholar] [CrossRef]
- Cho, J.G.; Lee, A.; Chang, W.; Lee, M.-S.; Kim, J. Endothelial to Mesenchymal Transition Represents a Key Link in the Interaction between Inflammation and Endothelial Dysfunction. Front. Immunol. 2018, 9, 294. [Google Scholar] [CrossRef] [PubMed]
- Di Gregorio, J.; Robuffo, I.; Spalletta, S.; Giambuzzi, G.; De Iuliis, V.; Toniato, E.; Martinotti, S.; Conti, P.; Flati, V. The Epithelial-to-Mesenchymal Transition as a Possible Therapeutic Target in Fibrotic Disorders. Front. Cell Dev. Biol. 2020, 8, 607483. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Wei, W.-Y.; Li, L.-L.; Hu, C.; Tang, Q.-Z. Therapeutic Potential of Polyphenols in Cardiac Fibrosis. Front. Pharmacol. 2018, 9, 122. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Lozano, M.; González-González, A.; Marchal, J.A.; Muela, E.M.; Molina, M.P.; Cara, F.E.; Brown, A.M.; García-Rivas, G.; Hernández-Brenes, C.; Lorente, J.A.; et al. Hydroxytyrosol inhibits cancer stem cells and the metastatic capacity of triple-negative breast cancer cell lines by the simultaneous targeting of epithelial-to-mesenchymal transition, Wnt/β-catenin and TGFβ signaling pathways. Eur. J. Nutr. 2018, 58, 3207–3219. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Accession Number | Forward Primer | Reverse Primer | Size (bp) |
|---|---|---|---|---|
| HERPUD1 | NM_014685.4 | 5′- TATGGGTGGCTTCAGCTTTC-3′ | 5′-GGCTCCAGGATTAACAACCA-3′ | 228 |
| DNAJB9 | NM_012328.3 | 5′-TCGGAGGGTGCAGGATATTA-3′ | 5′-AGCATCCGGGCTCTTATTTT-3′ | 212 |
| HSPA5 | NM_005347.5 | 5′-TGCAGCAGGACATCAAGTTC-3′ | 5′-AGTTCCAGCGTCTTTGGTTG-3′ | 245 |
| DGAT2 | NM_032564.5 | 5′-GCTGACCTGGTTCCCATCTA-3′ | 5′-CAGGTGTCGGAGGAGAAGAG-3′ | 164 |
| FGF18 | NM_003862.3 | 5′-GGACCAGTGGGAAACACATC-3′ | 5′-CAGGGCCGTGTAGTTGTTCT-3′ | 245 |
| APLN | NM_017413.5 | 5′-CCAAGGAGCAGCATGAATCT-3′ | 5′-GAAAGGCATGGGTCCCTTAT-3′ | 243 |
| LTB | NM_002341.2 | 5′-AGGAGCCACTTCTCTGGTGA-3′ | 5′-CAGCTTCTGAAACCCCAGTC-3′ | 151 |
| CCL2 | NM_002982.3 | 5′-CCCCAGTCACCTGCTGTTAT-3 | 5′-TCCTGAACCCACTTCTGCTT-3′ | 166 |
| CXCL5 | NM_002994.5 | 5′-CCACTATGAGCCTCCTGTCC-3′ | 5′-CTATGGCGAACACTTGCAGA-3′ | 219 |
| CXCL11 | NM_005409.5 | 5′-AGAGGACGCTGTCTTTGCAT-3′ | 5′-TAAGCCTTGCTTGCTTCGAT-3′ | 175 |
| GAPDH | NM_002046.3 | 5′-ATCACTGCCACCCAGAAGAC-3′ | 5′-TTCTAGACGGCAGGTCAGGT-3′ | 210 |
| 18S rRNA | NR_003286.2 | 5′-AAACGGCTACCACATCCAAG-3′ | 5′-CCTCCAATGGATCCTCGTTA-3′ | 155 |
| Fold Change | HT/CTR | HT-IL1B/IL1B | ||||
|---|---|---|---|---|---|---|
| Total | Numbers | Regulation | Total | Numbers | Regulation | |
| >2.0 | 71 | 44 | up | 45 | 33 | up |
| 27 | down | 12 | down | |||
| 1.5<~<2.0 | 398 | 249 | up | 390 | 243 | up |
| 149 | down | 147 | down | |||
| Total | 469 | 293 | up | 435 | 276 | up |
| 176 | down | 159 | down | |||
| HT/CTR | |||||||
|---|---|---|---|---|---|---|---|
| Symbol | Entrez Gene Name | Function | Fold Change | Symbol | Entrez Gene Name | Function | Fold Change |
| HERPUD1 | homocysteine inducible ER protein with ubiquitin-like domain 1 | regulator of protein degradation | 6.603 | CXCL11 | C-X-C motif chemokine ligand 11 | chemotactic protein | −2.181 |
| DNAJB9 | DnaJ heat shock protein family (Hsp40) member B9 | co-chaperone | 4.179 | CCDC191 | coiled-coil domain containing 191 | uncharacterized | −2.215 |
| CDH15 | cadherin 15 | cell adhesion protein | 3.803 | SNHG8 | small nucleolar RNA host gene 8 | long noncoding RNA | −2.232 |
| SDF2L1 | stromal cell-derived factor 2 like 1 | chaperone binding | 3.617 | LOC730101 | uncharacterized LOC730101 | uncharacterized | −2.318 |
| HSPA5 | heat shock protein family A (Hsp70) member 5 | chaperone | 3.611 | FGF18 | fibroblast growth factor 18 | growth factor | −2.344 |
| WTAP | WT1 associated protein | splicing regulator | 2.992 | TSLP | thymic stromal lymphopoietin | hemopoietic cytokine | −2.562 |
| H1-5 | H1.5 linker histone. cluster member | chromatin DNA binding | 2.969 | CXCL5 | C-X-C motif chemokine ligand 5 | Chemotactic protein | −2.708 |
| NMRAL2P | NmrA-like redox sensor 2. Pseudo gene | uncharacterized | 2.907 | ZNF33B | zinc finger protein 33B | transcription regulator | −2.742 |
| NEUROG3 | neurogenin 3 | transcription regulator | 2.847 | CDADC1 | cytidine and dCMP deaminase domain containing 1 | DNA cytosine deamination | −2.758 |
| FICD | FIC domain containing | Nucleotidyl transferase | 2.740 | ZNF594 | zinc finger protein 594 | transcription regulator | −3.310 |
| HT-IL1B/IL1B | |||||||
|---|---|---|---|---|---|---|---|
| Symbol | Entrez Gene Name | Function | Fold Change | Symbol | Entrez Gene Name | Function | Fold Change |
| HERPUD1 | homocysteine inducible ER protein with ubiquitin-like domain 1 | regulator of protein degradation | 5.383 | IP6K2 | inositol hexakisphosphate kinase 2 | kinase | −2.024 |
| DNAJB9 | DnaJ heat shock protein family (Hsp40) member B9 | co-chaperone | 4.232 | ZNF33B | zinc finger protein 33B | transcription regulator | −2.026 |
| MAT2A | methionine adenosyltransferase 2A | transferase | 2.866 | APLN | Apelin | protein binding | −2.233 |
| ZNF658 | zinc finger protein 658 | transcription regulator | 2.793 | LTB | lymphotoxin beta | cytokine | −2.238 |
| HSPA5 | heat shock protein family A (Hsp70) member 5 | chaperone | 2.754 | RBM12B | RNA binding motif protein 12B | RNA binding | −2.238 |
| ALPK1 | alpha kinase 1 | kinase | 2.668 | RGS16 | regulator of G protein signaling 16 | enzyme | −2.270 |
| ERO1B | endoplasmic reticulum oxidoreductase 1 beta | oxidoreductase | 2.647 | SRA1 | steroid receptor RNA activator 1 | transcription regulator | −2.340 |
| FDXACB1 | ferredoxin-fold anticodon binding domain containing 1 | uncharacterized | 2.592 | ZNF594 | zinc finger protein 594 | transcription regulator | −2.586 |
| SLC7A11 | solute carrier family 7 member 11 | transporter | 2.559 | DGAT2 | diacylglycerol O-acyltransferase 2 | acyltransferase | −2.648 |
| SLC10A7 | solute carrier family 10 member 7 | transporter | 2.465 | APOL6 | apolipoprotein L6 | lipid binding | −2.824 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carluccio, M.A.; Martinelli, R.; Massaro, M.; Calabriso, N.; Scoditti, E.; Maffia, M.; Verri, T.; Gatta, V.; De Caterina, R. Nutrigenomic Effect of Hydroxytyrosol in Vascular Endothelial Cells: A Transcriptomic Profile Analysis. Nutrients 2021, 13, 3990. https://doi.org/10.3390/nu13113990
Carluccio MA, Martinelli R, Massaro M, Calabriso N, Scoditti E, Maffia M, Verri T, Gatta V, De Caterina R. Nutrigenomic Effect of Hydroxytyrosol in Vascular Endothelial Cells: A Transcriptomic Profile Analysis. Nutrients. 2021; 13(11):3990. https://doi.org/10.3390/nu13113990
Chicago/Turabian StyleCarluccio, Maria Annunziata, Rosanna Martinelli, Marika Massaro, Nadia Calabriso, Egeria Scoditti, Michele Maffia, Tiziano Verri, Valentina Gatta, and Raffaele De Caterina. 2021. "Nutrigenomic Effect of Hydroxytyrosol in Vascular Endothelial Cells: A Transcriptomic Profile Analysis" Nutrients 13, no. 11: 3990. https://doi.org/10.3390/nu13113990
APA StyleCarluccio, M. A., Martinelli, R., Massaro, M., Calabriso, N., Scoditti, E., Maffia, M., Verri, T., Gatta, V., & De Caterina, R. (2021). Nutrigenomic Effect of Hydroxytyrosol in Vascular Endothelial Cells: A Transcriptomic Profile Analysis. Nutrients, 13(11), 3990. https://doi.org/10.3390/nu13113990