
Cyperus esculentus L. and Tetracarpidium conophorum Müll. Arg. Supplemented Diet Improved Testosterone Levels, Modulated Ectonucleotidases and Adenosine Deaminase Activities in Platelets from L-NAME-Stressed Rats

 and
and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Sample Preparation
2.3. Animals
2.4. Design of Experiments
2.5. Hormonal Assays
2.6. Platelet Preparation
2.7. Determination of E-NTPDase and Ecto-5′-Nucleotidase Activities
2.8. Adenosine Deaminase Activity Determination
2.9. Protein Determination
2.10. Statistical Evaluations
3. Results
3.1. Feed Intake Measurement and the Impact of TN and WN Supplementation in L-NAME-Stressed Rats
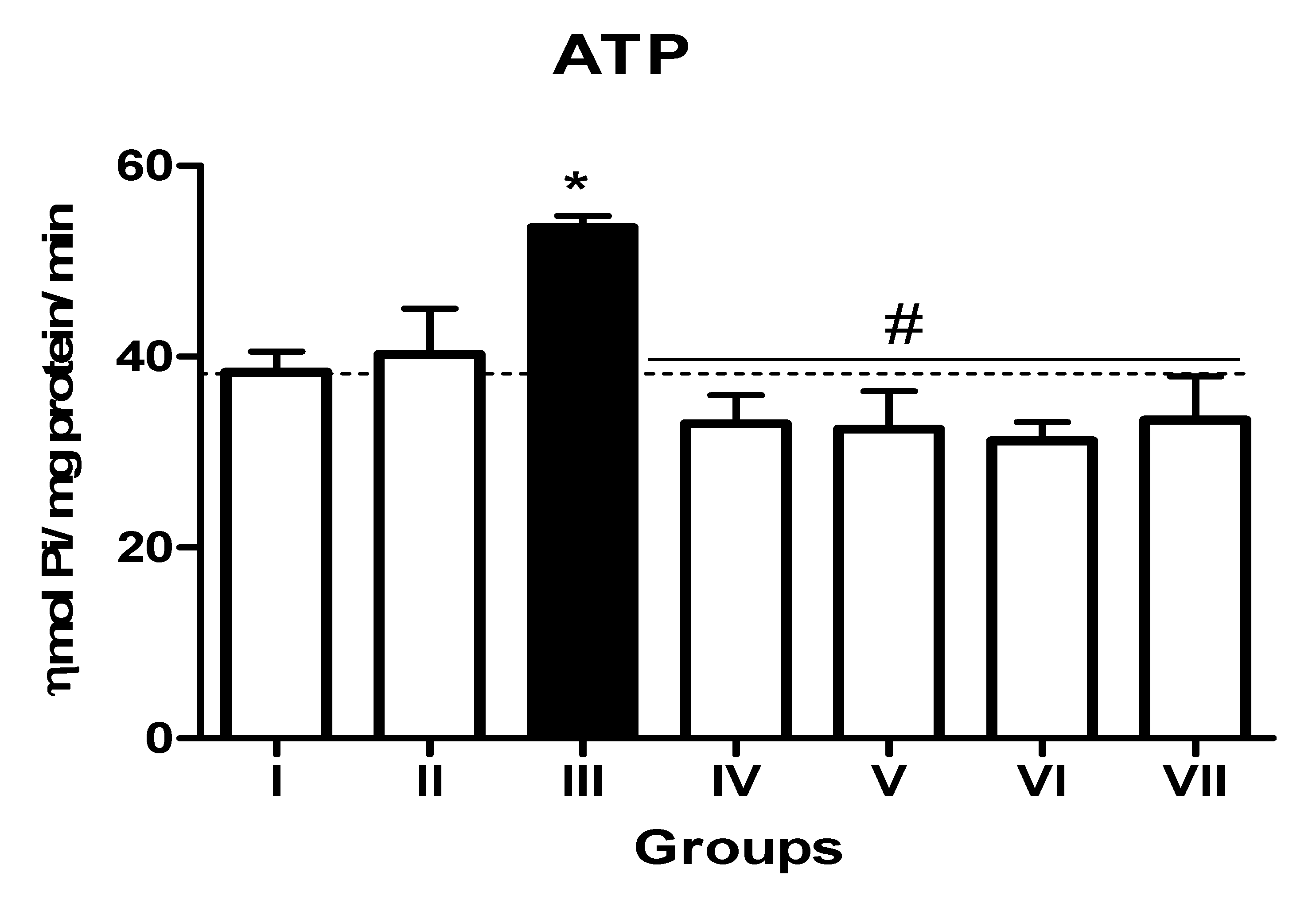
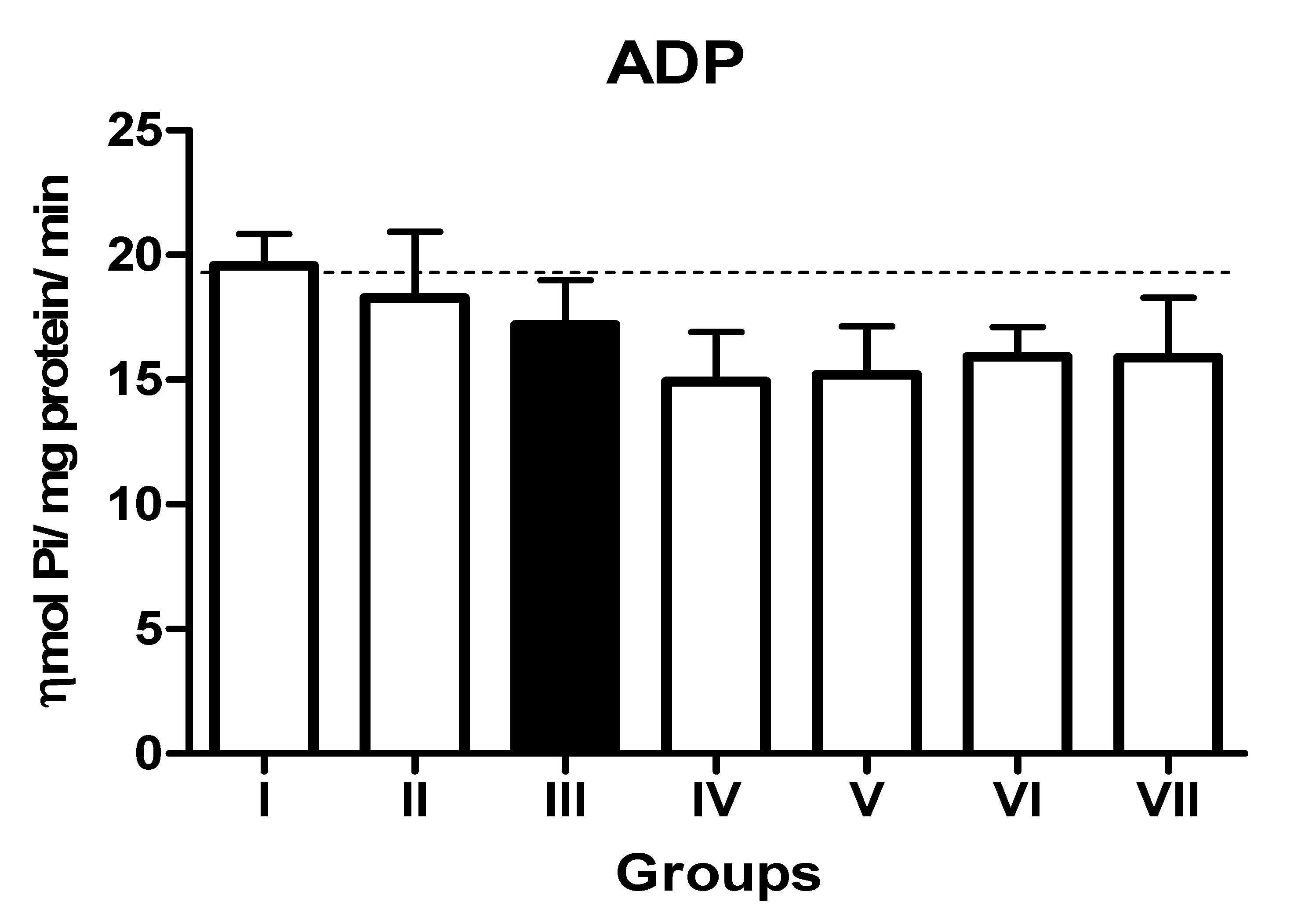
3.2. Impact of TN and WN Supplementation on the Activity of EC 3.6.1.5, EC 3.1.3.5, and ADA in the Platelets of L-NAME-Stressed Rats
3.3. Impact of TN and WN Supplementation on Testosterone and Luteinizing Hormone (LH) Levels in L-NAME-Stressed Rats
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ATP | adenosine triphosphate |
| ADP | adenosine diphosphate |
| AMP | adenosine monophosphate |
| ADA | adenosine deaminase |
| E-NTPdase | ectonucleotidases |
| ED | erectile dysfunction |
| L-NAME | Nω-nitro-L-arginine methyl ester hydrochloride |
| NOS | nitric oxide synthase |
| SIL | sildenafil |
| TN | tiger nut |
| WN | walnut |
References
- Vorchheimer, D.A.; Becker, R. Platelets in atherothrombosis. Mayo Clin. Proc. 2006, 81, 59–68. [Google Scholar] [CrossRef] [Green Version]
- Tousoulis, D.; Paroutoglou, L.P.; Papageorgiou, N.; Charakida, M.; Stefanadis, C. Recent therapeutic approaches to platelet activation in coronary artery disease. Pharmacol. Ther. 2010, 127, 108–120. [Google Scholar] [CrossRef]
- Vilahur, G.; Badimon, L. Antiplatelet properties of natural products. Vasc. Pharmacol. 2013, 59, 67–75. [Google Scholar] [CrossRef]
- Dockrell, M.E.C.; Walter, B.R.; Noon, J.P.; Watt, G.C.M.; Williams, B.C.; Webb, D.J. Hypertensive patients with carotid artery plaque exhibit increased platelet aggregability. Thromb. Res. 2006, 117, 615–622. [Google Scholar]
- Biancardi, V.C.; Bergamaschi, C.T.; Lopes, O.U.; Campos, R.R. Sympathetic activation in rats with L-NAME-induced hypertension. Braz. J. Med. Biol. Res. 2007, 40, 401–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lackland, D.T.; Weber, M.A. Global burden of cardiovascular disease and stroke: Hypertension at the core. Can. J. Cardiol. 2015, 31, 569–571. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, A.M.; Bagatini, M.D.; Martins, C.C.; Abdalla, F.H.; Zanini, D.; Schmatz, R.; Gutierres, J.; Pimentel, V.C.; Thomé, G.; Leal, C.A.M.; et al. Exercise training prevents ecto-nucleotidases alterations in platelets of hypertensive rats. Mol. Cell Biochem. 2012, 371, 147–156. [Google Scholar] [CrossRef]
- Akinyemi, A.J.; Thome, G.R.; Morsch, V.M.; Stefanello, N.; Goularte, J.F.; Belló-Klein, A.; Oboh, G.; Schetinger, M.R.C. Effect of dietary supplementation of ginger and turmeric rhizomes on angiotensin-1 converting enzyme (ACE) and arginase activities in L-NAME induced hypertensive rats. J. Funct. Foods 2015, 17, 792–801. [Google Scholar] [CrossRef]
- Olabiyi, A.A.; Carvalho, F.B.; Bottari, N.B.; Lopes, T.F.; da Costa, P.; Stefanelo, N.; Morsch, V.M.; Akindahunsi, A.A.; Oboh, G.; Schetinger, M.R. Dietary supplementation of tiger nut alters biochemical parameters relevant to erectile function after administration of L-NAME in rats. Food Res. Intern. 2018, 109, 358–367. [Google Scholar] [CrossRef]
- Godschalk, M.F.; Sison, A.; Mulligan, T. Diagnosis and therapy of erectile dysfunction. Drugs Today 1997, 33, 475–483. [Google Scholar]
- Jeremy, J.Y.; Ballard, S.; Naylor, A.; Miller, M.A.W.; Angelini, G.D. Effect of sildenafil (ViagraE), a specific inhibitor of cGMP phosphodiesterase on cAMP and cGMP formation by the rabbit corpus cavernosa, in vitro. Br. J. Urol. 1997, 79, 958–963. [Google Scholar] [CrossRef] [PubMed]
- Langtry, H.D.; Markham, A. Sildenafil: A review on its use in erectile dysfunction. Drugs 1999, 57, 967–989. [Google Scholar] [CrossRef] [PubMed]
- Anderson, K.E. Pharmacology of lower urinary tract smooth muscles and penile erectile tissues. Pharmacol. Rev. 1993, 45, 253–308. [Google Scholar] [PubMed]
- De Tejada, I.S.; Angulo, J.; Cellek, S.; González-Cadavid, N.; Heaton, J.; Pickard, R.; Simonsen, U. Pathophysiology of erectile dysfunction. J. Sex. Med. 2005, 2, 26–39. [Google Scholar] [CrossRef] [PubMed]
- Sobol, A.B.; Watala, C. The role of platelets in diabetes-related vascular complications. Diabetes Res. Clin. Pract. 2000, 50, 1–6. [Google Scholar] [CrossRef]
- Haouari, M.E.; Rosado, J.A. Platelet signalling abnormalities in patients with type 2 diabetes mellitus: A review. Blood Cells Mol. Dis. 2008, 41, 119–123. [Google Scholar] [CrossRef]
- Fürstenau, C.R.; Trentin, D.D.S.; Gossenheimer, A.N.; Ramos, D.B.; Casali, E.A.; Barreto-Chaves, M.L.M.; Sarkis, J.J.F. Ectonucleotidase activities are altered in serum and platelets of L-NAME-treated rats. Blood Cells Mol. Dis. 2008, 41, 223–229. [Google Scholar] [CrossRef]
- Robson, S.C.; Sévigny, J.; Zimmermann, H. The E-NTPDase family of ectonucleotidases: Structure function relationships and pathophysiological significance. Purinergic Signal. 2000, 2, 409–430. [Google Scholar] [CrossRef] [Green Version]
- Sträter, N. Ecto-5′-nucleotidase: Structure function relationships. Purinergic Signal. 2006, 2, 343–350. [Google Scholar] [CrossRef] [Green Version]
- Blackburn, M.R.; Kellems, R.E. Adenosine deaminase deficiency: Metabolic basis of immune deficiency and pulmonary inflammation. Adv. Immunol. 2005, 86, 1–41. [Google Scholar]
- Yegutkin, G.G. Nucleotide- and nucleoside-converting ectoenzymes: Important modulators of purinergic signalling cascade. Biochim. Biophys. Acta-Bioenerg. 2008, 1783, 673–694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajfer, J. Relationship between testosterone and erectile dysfunction. Rev. Urol. 2000, 2, 122–128. [Google Scholar] [PubMed]
- Wu, F.C.; Tajar, A.; Beynon, J.M.; Pye, S.; Silman, A.J.; Finn, J.D.; O’Neill, T.; Bartfai, G.; Casanueva, F.F.; Forti, G.; et al. Identification of late-onset hypogonadism in middle-aged and elderly men. N. Engl. J. Med. 2010, 363, 123–135. [Google Scholar] [CrossRef]
- Hull, E.; Lorrain, D.S.; Du, J.; Matuszewich, L.; Lumley, L.A.; Putnam, S.K.; Moses, J. Hormone-neurotransmitter interactions in the control of sexual behavior. Behav. Brain Res. 1999, 105, 105–116. [Google Scholar] [CrossRef]
- Corona, G.; Isidori, A.M.; Buvat, J.; Aversa, A.; Ratrelli, G.; Hackett, G.; Rochira, V.; Lenzi, A.; Maggi, M. Testosterone supplementation and sexual function: A meta-analysis study. J. Sex. Med. 2014, 11, 1577–1592. [Google Scholar] [CrossRef]
- Adejuyitan, J.A.; Otunola, E.T.; Akande, E.A.; Bolarinwa, I.F.; Oladokun, F.M. Some physicochemical properties of flour obtained from fermentation of tigernut (Cyperus esculentus) sourced from a market in Ogbomoso, Nigeria. Afr. J. Food Sci. 2009, 3, 51–55. [Google Scholar]
- Salem, L.M.; Zommara, M.; Imaizumi, K. Dietary supplementation with Cyperus esculentus L. (tiger nut) tubers attenuated atherosclerotic lesion in apolip-oprotein E knockout mouse associated with inhibition of inflammatory cell responses. Am. J. Immunol. 2005, 1, 60–67. [Google Scholar]
- Chukwuma, E.R.; Obioma, N.; Christophe, O.I. The phytochemical composition and some biochemical effects of Nigerian tigernut (Cyperus esculentus L.) tuber. Pak. J. Nutr. 2010, 9, 709–715. [Google Scholar] [CrossRef] [Green Version]
- Amaeze, O.U.; Ayoola, G.A.; Sofidiya, M.O.; Adepoju-Bello, A.A.; Adegoke, A.O.; Coker, H.A.B. Evaluation of antioxidant activity of Tetracarpidium conophorum (Müll. Arg) Hutch & Dalziel leaves. Oxid. Med. Cell Longev. 2011, 2011, 976701. [Google Scholar]
- Olabiyi, A.A.; Oboh, G.; Adefegha, S.A. Effect of dietary supplementation of tiger nut (Cyperus esculentus L.) and walnut (Tetracarpidium conophorum Müll. Arg.) on sexual behavior, hormonal level, and antioxidant status in male rats. J. Food Biochem. 2016, 41, e12351. [Google Scholar] [CrossRef]
- Olabiyi, A.A.; Oboh, G.; Akinyemi, A.J.; Ademiluyi, A.O.; Boligon, A.A.; de Campos, M.M.A. Tiger nut (Cyperus esculentus L.) supplemented diet modulate key biochemical indices relevant to erectile function in male rats. J. Funct. Foods 2017, 34, 152–158. [Google Scholar] [CrossRef]
- Olabiyi, A.A.; Carvalho, F.B.; Bottari, N.B.; Morsch, V.M.; Morel, A.F.; Oboh, G.; Schetinger, M.R. Tiger nut and walnut extracts modulate extracellular metabolism of ATP and adenosine through the NOS/cGMP/PKG signaling pathway in kidney slices. Phytomedicine 2018, 43, 140–149. [Google Scholar] [CrossRef] [PubMed]
- Lunkes, G.I.; Lunkes, D.S.; Morsch, V.M.; Mazzanti, C.M.; Morsch, A.L.; Miron, V.R.; Schetinger, M.R. NTPDase and 5′-nucleotidase activities in rats alloxan-induced diabetes. Diabetes Res. Clin. Pract. 2004, 65, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Schetinger, M.R.C.; Morsch, V.M.; Bonan, C.D.; Wyse, A.T. NTPDase and 5′-nucleotidase activities in physiological and disease conditions: New perspectives for human health. BioFactors 2007, 31, 77–98. [Google Scholar] [CrossRef] [PubMed]
- Heymann, D.; Reddington, M.; Kreutzberg, G.W. Subcellular localization of 5′-nucleotidase in rat brain. J. Neurochem. 1984, 43, 971–978. [Google Scholar] [CrossRef] [PubMed]
- Chan, K.-M.; Delfert, D.; Junger, K.D. A direct colorimetric assay for Ca2+-stimulated ATPase activity. Anal. Biochem. 1986, 157, 375–380. [Google Scholar] [CrossRef]
- Guisti, G.; Galanti, B. Colorimetric method. In Methods of Enzymatic Analysis; Bergmeyer, H.U., Ed.; Academic Press: New York, NY, USA, 1974; pp. 315–323. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Lippi, G.; Mattiuzzi, C.; Comelli, I.; Cervellin, G. Mean platelet volume in patients with ischemic heart disease: Meta-analysis of diagnostic studies. Blood Coag. Fibrinol. 2013, 24, 216–219. [Google Scholar] [CrossRef]
- Jeremy, J.; Angelini, G.; Khan, M.; Mikhailidis, D.; Morgan, R.; Thompson, C.; Bruckdorfer, K.; Naseem, K. Platelets, oxidant stress and erectile dysfunction: An hypothesis. Cardiovasc. Res. 2000, 46, 50–54. [Google Scholar] [CrossRef] [Green Version]
- Kouidrat, Y.; Pizzol, D.; Cosco, T.; Thompson, T.; Carnaghi, M.; Bertoldo, A.; Solmi, M.; Stubbs, B.; Veronese, N. High prevalence of erectile dysfunction in diabetes: A systematic review and meta-analysis of 145 studies. Diabet. Med. 2017, 34, 1185–1192. [Google Scholar] [CrossRef]
- Esposito, K.; Maiorino, M.I.; Bellastella, G. Diabetes and sexual dysfunction; Current perspective. Diabetes Metab. Syndr. Obes. Targets Ther. 2014, 7, 95–105. [Google Scholar] [CrossRef] [Green Version]
- Agrawal, J.; Singh, V.B.; Meena, B.; Yadav, R.; Shareef, M.; Saini, V. Mean paltelet volume (MPV) the neglected marker of cardiovascular risk and glycosylated hemoglobin in diabetic patient. Diabetes Metab. Syndr. Clin. Res. Rev. 2016, 11, 109–112. [Google Scholar] [CrossRef] [PubMed]
- Lunkes, G.I.; Lunkes, D.; Stefanello, F.; Morsch, A.; Morsch, V.M.; Mazzanti, C.M.; Schetinger, M.R.C. Enzymes that hydrolyze adenine nucleotides in diabetes and associated pathologies. Thromb. Res. 2003, 2183, 189–194. [Google Scholar] [CrossRef]
- Leal, C.A.M.; Schetinger, M.R.C.; Leal, D.B.R.; Bauchspiess, K.; Schrekker, C.M.L.; Maldonado, P.A.; Morsch, V.M.; Da Silva, J.E.P. NTPDase and 5′-nucleotidase activities in platelets of human pregnant with a normal or high risk from thrombosis. Mol. Cell. Biochem. 2007, 304, 325–330. [Google Scholar] [CrossRef]
- Duarte, M.M.F.; Loro, V.L.; Rocha, J.B.T.; Leal, D.B.R.; de Bem, A.F.; Dorneles, A.; Morsch, V.M.; Schetinger, M.R.C. Enzymes that hydrolyze adenine nucleotides of patients with hypercholesterolemia and inflammatory processes. FEBS J. 2007, 274, 2707–2714. [Google Scholar] [CrossRef]
- Hourani, S.M.O. Purinoceptors and platelet aggregation. J. Auton. Pharmacol. 1996, 16, 349–352. [Google Scholar] [CrossRef]
- Ansari, H.R.; Nadeem, A.; Talukder, M.H.; Sakhalkar, S.; Mustafa, S.J. Evidence for the involvement of nitric oxide in A2B receptor-mediated vasorelaxation of mouse aorta. Am. J. Physiol. Heart Circ. Physiol. 2007, 292, 719–725. [Google Scholar] [CrossRef]
- Preli, R.B.; Klein, K.P.; Herrington, D.M. Vascular effects of dietary L-arginine supplementation. Atherosclerosis 2002, 162, 1–15. [Google Scholar] [CrossRef]
- Iyú, D.; Atucha, N.M.; Martínez-Prieto, C.; Ortiz, M.C.; García-Estañ, J. Altered calcium signaling in platelets from nitric oxide-deficient hypertensive. Cell Commun. Signal. 2004, 2, 1. [Google Scholar] [CrossRef] [Green Version]
- Gudmundsdóttir, I.J.; McRobbie, S.J.; Robinson, S.D.; Newby, D.E.; Megson, I.L. Sildenafil potentiates nitric oxide mediated inhibition of human platelet aggregation. Biochem. Biophys. Res. Commun. 2005, 337, 382–385. [Google Scholar] [CrossRef] [PubMed]
- Batlouni, M. Ativação plaquetária e trombose arterial. Arq. Bras. Cardiol. 1993, 60, 425–431. [Google Scholar] [PubMed]
- Vasta, V.; Meacci, E.; Farnararo, M.; Bruni, P. Identification of a specific transport system for L-arginine in human platelets. Biochem. Biophys. Res. Commun. 1995, 206, 878–884. [Google Scholar] [CrossRef] [PubMed]
- De Meirelles, L.R.; Mendes-Ribeiro, A.C.; Santoro, M.M.; Mendes, M.A.; Da Silva, M.N.; Mann, G.E.; Brunini, T.M. Inhibitory effects of endogenous L-arginine analogues on nitric oxide synthesis in platelets: Role in pletelet hyperaggregability in hypertension. Clinic. Exp. Pharmacol. Physiol. 2007, 34, 1267–1271. [Google Scholar] [CrossRef] [PubMed]
- McNairn, A.J.; Chuang, C.H.; Bloom, J.C.; Wallace, M.D.; Schimenti, J.C. Femalebiased embryonic death from inflammation induced by genomic instability. Nature 2019, 567, 105–108. [Google Scholar] [CrossRef] [PubMed]
- Banihani, S.A. Testosterone in males as enhanced by onion (Allium Cepa L.). Biomolecules 2019, 9, 75. [Google Scholar] [CrossRef] [Green Version]
- Banihani, S.A. Ginger and testosterone. Biomolecules 2018, 8, 119. [Google Scholar] [CrossRef]
- Zhang, J.; Li, X.; Cai, Z.; Li, H.; Yang, B. Association between testosterone with type 2 diabetes in adult males, a meta-analysis and trial sequential analysis. Aging Male 2019, 23, 607–618. [Google Scholar] [CrossRef] [PubMed]
- Asih, P.R.; Tegg, M.L.; Sohrabi, H.; Carruthers, M.; Gandy, S.E.; Saad, F.; Verdile, G.; Ittner, L.M.; Martins, R.N. Multiple mechanisms linking type 2 diabetes and Alzheimer’s disease: Testosterone as a modifier. J. Alzheimers Dis. 2017, 59, 445–466. [Google Scholar] [CrossRef]
- Pike, C.J. Sex and the development of Alzheimer’s disease. J. Neurosci. Res. 2017, 95, 671–680. [Google Scholar] [CrossRef] [Green Version]
- Kloner, R.A.; Carson, C.; Dobs, A.; Kopecky, S.; Mohler, E.R. Testosterone and cardiovascular disease. J. Am. Coll. Cardiol. 2016, 67, 545–557. [Google Scholar] [CrossRef]
- Budoff, M.J.; Ellenberg, S.S.; Lewis, C.E.; Mohler, E.R.; Wenger, N.K.; Bhasin, S.; Barret-Connor, E.; Swerdloff, R.S.; Stephen-Shields, A.; Cauley, J.A.; et al. Testosterone treatment and coronary artery plaque volume in older men with low testosterone. J. Am. Med. Assoc. 2017, 317, 708–716. [Google Scholar] [CrossRef]
- Kaplan, A.L.; Hu, J.C.; Morgentaler, A.; Mulhall, J.P.; Schulman, C.C.; Montorsi, F. Testosterone therapy in men with prostate cancer. Eur. Urol. 2015, 69, 894–903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohamad, N.V.; Soelaiman, I.-N.; Chin, K.-Y. A concise review of testosterone and bone health. Clin. Interv. Aging 2015, 11, 1317–1324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, X.; Li, J.; Zhang, H.; Wang, H.; Yin, G.; Miao, D. Pyrroloquinoline quinone prevents testosterone deficiency-induced osteoporosis by stimulating osteoblastic bone formation and inhibiting osteoclastic bone resorption. Am. J. Transl. Res. 2017, 9, 1230–1242. [Google Scholar] [PubMed]
- El-Migdadi, F.; Banihani, I.; Banihani, S.A. Clinico-hormonal correlation of oligospermic patients in the below sea level environment (Jordan valley). Neuroendocrinol. Lett. 2005, 26, 13–18. [Google Scholar] [PubMed]
- Banihani, S.A. Effect of coenzyme q10 supplementation on testosterone. Biomolecules 2018, 8, 172. [Google Scholar] [CrossRef] [Green Version]

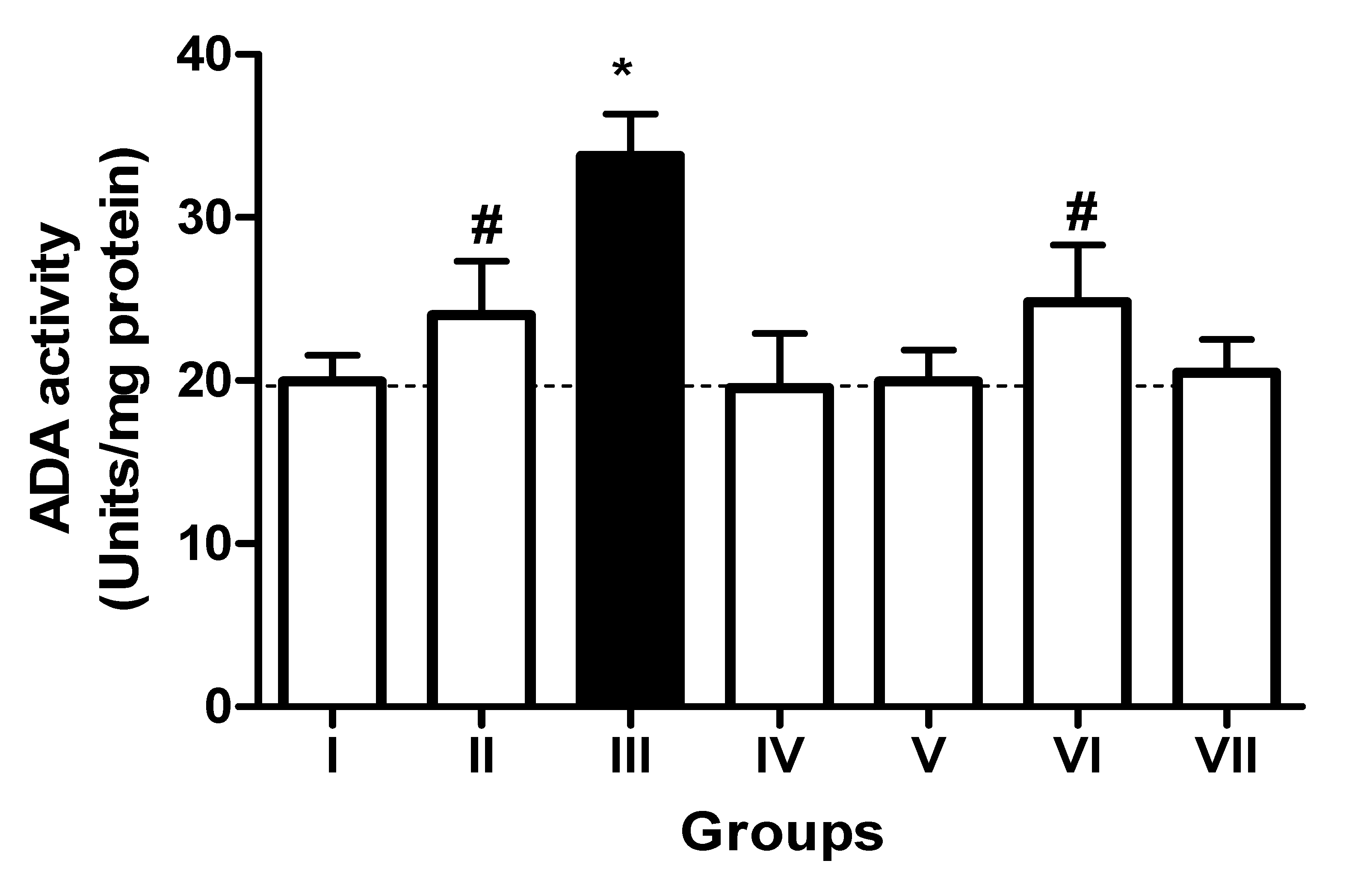

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | |||||||
|---|---|---|---|---|---|---|---|
| Materials | I | II | III | IV | V | VI | VII |
| Skimmed milk | 37.5 | 37.5 | 37.5 | 33.1 | 33.1 | 21.3 | 26.3 |
| Oil | 10.0 | 10.0 | 10.0 | 10.0 | 10.0 | 10.0 | 10.0 |
| Vitamin mix. | 4.0 | 4.0 | 4.0 | 4.0 | 4.0 | 4.0 | 4.0 |
| Corn Starch | 48.5 | 48.5 | 48.5 | 32.9 | 32.9 | 44.7 | 39.7 |
| Tig 1 | - | - | - | 20.0 | - | - | - |
| Tig 2 | - | - | - | - | 20.0 | - | - |
| Wal 1 | - | - | - | - | - | 20.0 | - |
| Wal 2 | - | - | - | - | - | - | 20.0 |
| VIAGRA | - | + | - | - | - | - | - |
| L-NAME | - | + | + | + | + | + | + |
| Total (g) | 100 | 100 | 100 | 100 | 100 | 100 | 100 |
| Groups | Testosterone Hormone | Luteinizing Hormone |
|---|---|---|
| (ng/dL) | (mIU/mL) | |
| I | 306.7 ± 5.7 a | 28.5 ± 0.1 a |
| II | 102.4 ± 7.4 d | 27.8 ± 0.9 a |
| III | 54.5 ± 2.2 f | 27.1 ± 0.2 |
| IV | 88.7 ± 0.7 e | 27.5 ± 0.6 a |
| V | 118.2 ± 9.3 c | 29.2 ± 0.7 a |
| VI | 145.5 ± 6.0 b | 29.8 ± 0.4 a |
| VII | 123.8 ± 5.7 c | 29.5 ± 0.3 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Olabiyi, A.A.; Morsch, V.M.; Oboh, G.; Schetinger, M.R.C. Cyperus esculentus L. and Tetracarpidium conophorum Müll. Arg. Supplemented Diet Improved Testosterone Levels, Modulated Ectonucleotidases and Adenosine Deaminase Activities in Platelets from L-NAME-Stressed Rats. Nutrients 2021, 13, 3529. https://doi.org/10.3390/nu13103529
Olabiyi AA, Morsch VM, Oboh G, Schetinger MRC. Cyperus esculentus L. and Tetracarpidium conophorum Müll. Arg. Supplemented Diet Improved Testosterone Levels, Modulated Ectonucleotidases and Adenosine Deaminase Activities in Platelets from L-NAME-Stressed Rats. Nutrients. 2021; 13(10):3529. https://doi.org/10.3390/nu13103529
Chicago/Turabian StyleOlabiyi, Ayodeji Augustine, Vera Maria Morsch, Ganiyu Oboh, and Maria Rosa Chitolina Schetinger. 2021. "Cyperus esculentus L. and Tetracarpidium conophorum Müll. Arg. Supplemented Diet Improved Testosterone Levels, Modulated Ectonucleotidases and Adenosine Deaminase Activities in Platelets from L-NAME-Stressed Rats" Nutrients 13, no. 10: 3529. https://doi.org/10.3390/nu13103529
APA StyleOlabiyi, A. A., Morsch, V. M., Oboh, G., & Schetinger, M. R. C. (2021). Cyperus esculentus L. and Tetracarpidium conophorum Müll. Arg. Supplemented Diet Improved Testosterone Levels, Modulated Ectonucleotidases and Adenosine Deaminase Activities in Platelets from L-NAME-Stressed Rats. Nutrients, 13(10), 3529. https://doi.org/10.3390/nu13103529