
Infant Formula Based on Milk Fat Affects Immune Development in Both Normal Birthweight and Fetal Growth Restricted Neonatal Piglets

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Care and Housing
2.2. Diets
2.3. Blood Sampling and Evaluation of Immune Parameters
2.4. Statistics
3. Results
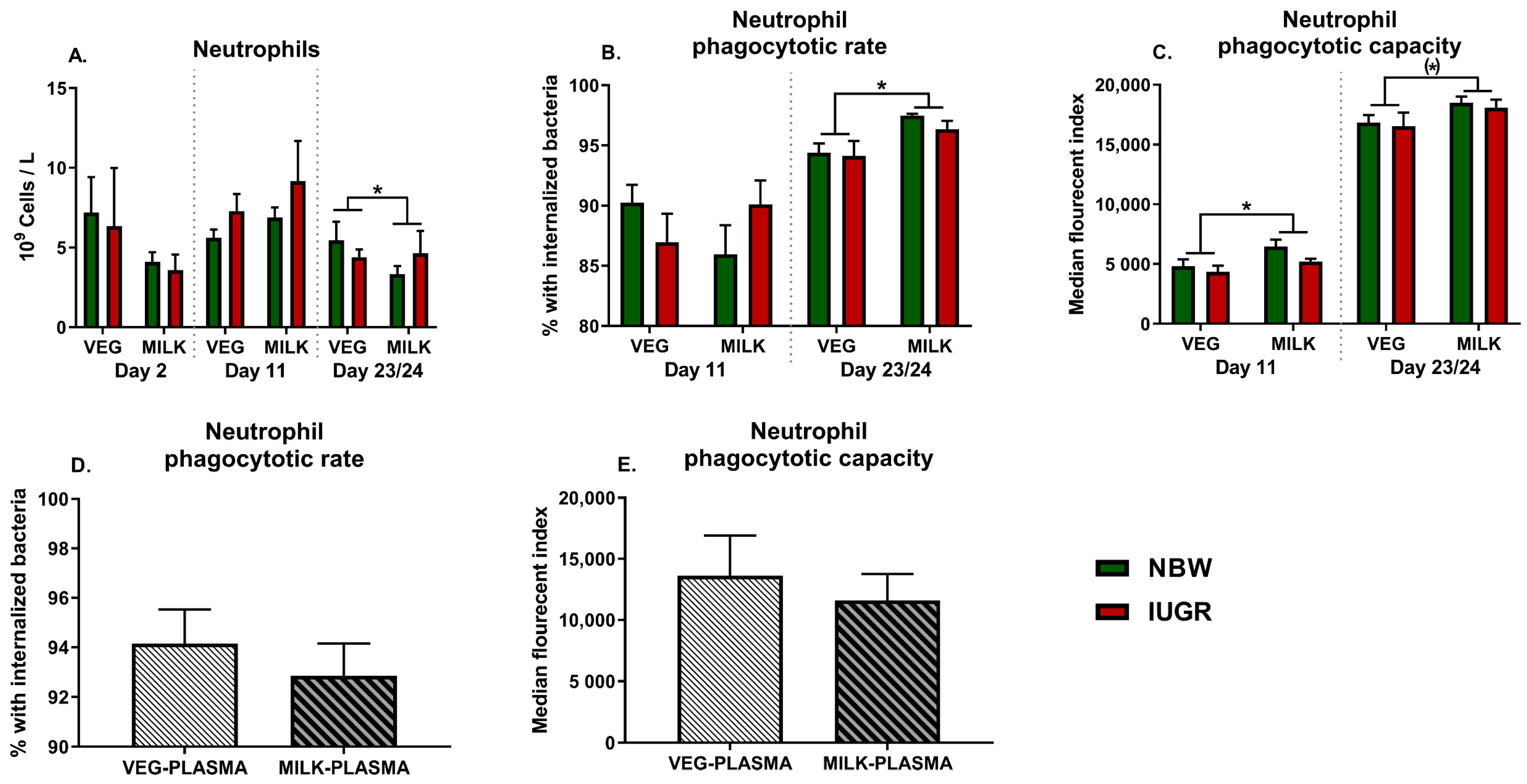
3.1. Neutrophil Development and Function
3.2. Innate Immune Parameters
3.3. Adaptive Immune Parameters
3.4. Cellular Metabolism
3.5. Other Hematological Parameters
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Jost, T.; Lacroix, C.; Braegger, C.P.; Chassard, C.; Zabielski, R. New Insights in Gut Microbiota Establishment in Healthy Breast Fed Neonates. PLoS ONE 2012, 7, e44595. [Google Scholar] [CrossRef] [PubMed]
- El Manouni el Hassani, S.; Berkhout, D.J.C.; Niemarkt, H.J.; Mann, S.; de Boode, W.P.; Cossey, V.; Hulzebos, C.V.; van Kaam, A.H.; Kramer, B.W.; van Lingen, R.A.; et al. Risk Factors for Late-Onset Sepsis in Preterm Infants: A Multicenter Case-Control Study. Neonatology 2019, 116, 42–51. [Google Scholar] [CrossRef] [PubMed]
- Ladomenou, F.; Moschandreas, J.; Kafatos, A.; Tselentis, Y.; Galanakis, E. Protective effect of exclusive breastfeeding against infections during infancy: A prospective study. Arch. Dis. Child. 2010, 95, 1004–1008. [Google Scholar] [CrossRef]
- Kristensen, S.; Salihu, H.M.; Keith, L.G.; Kirby, R.S.; Fowler, K.B.; Pass, M.A.B. SGA subtypes and mortality risk among singleton births. Early Hum. Dev. 2007, 83, 99–105. [Google Scholar] [CrossRef]
- Watts, T.; Roberts, I. Haematological abnormalities in the growth-restricted infant. Semin. Neonatol. 1999, 4, 41–54. [Google Scholar] [CrossRef]
- Li, J.; Li, H.; Mao, H.; Yu, M.; Yang, F.; Feng, T.; Fan, Y.; Lu, Q.; Shen, C.; Yin, Z.; et al. Impaired NK cell antiviral cytokine response against influenza virus in small-for-gestational-age neonates. Cell. Mol. Immunol. 2013, 10, 437–443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tröger, B.; Müller, T.; Faust, K.; Bendiks, M.; Bohlmann, M.K.; Thonnissen, S.; Herting, E.; Göpel, W.; Härtel, C. Intrauterine growth restriction and the innate immune system in preterm infants of ≤32 weeks gestation. Neonatology 2013, 103, 199–204. [Google Scholar] [CrossRef]
- Ferguson, A.C. Prolonged impairment of cellular immunity in children with intrauterine growth retardation. J. Pediatr. 1978, 93, 52–56. [Google Scholar] [CrossRef]
- McDade, T.W.; Beck, M.A.; Kuzawa, C.W.; Adair, L.S. Prenatal Undernutrition and Postnatal Growth Are Associated with Adolescent Thymic Function. J. Nutr. 2001, 131, 1225–1231. [Google Scholar] [CrossRef] [Green Version]
- Berger, A.; Fleith, M.; Crozier, G. Nutritional implications of replacing bovine milk fat with vegetable oil in infant formulas. J. Pediatr. Gastroenterol. Nutr. 2000, 30, 115–130. [Google Scholar] [CrossRef] [PubMed]
- Ballard, O.; Morrow, A.L. Human Milk Composition. Nutrients and Bioactive Factors. Pediatr. Clin. N. Am. 2013, 60, 49–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jensen, R.G. The composition of bovine milk lipids: January 1995 to December 2000. J. Dairy Sci. 2002, 85, 295–350. [Google Scholar] [CrossRef]
- Zou, X.; Guo, Z.; Jin, Q.; Huang, J.; Cheong, L.; Xu, X.; Wang, X. Composition and microstructure of colostrum and mature bovine milk fat globule membrane. Food Chem. 2015, 185, 362–370. [Google Scholar] [CrossRef]
- Zanabria, R.; Tellez, A.M.; Griffiths, M.; Sharif, S.; Corredig, M. Modulation of immune function by milk fat globule membrane isolates. J. Dairy Sci. 2014, 97, 2017–2026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fontecha, J.; Brink, L.; Wu, S.; Pouliot, Y.; Visioli, F.; Jiménez-Flores, R. Sources, Production, and Clinical Treatments of Milk Fat Globule Membrane for Infant Nutrition and Well-Being. Nutrients 2020, 12, 1607. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Zavaleta, N.; Chen, S.-Y.; Lönnerdal, B.; Slupsky, C. Effect of bovine milk fat globule membranes as a complementary food on the serum metabolome and immune markers of 6-11-month-old Peruvian infants. NPJ Sci. Food 2018, 2, 6. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Peng, Y.; Li, Z.; Christensen, B.; Heckmann, A.B.; Lagerqvist, C.; Stenlund, H.; Lonnerdal, B.; Hernell, O.; West, C.E. Serum cytokine patterns are modulated in infants fed formula with probiotics or milk fat globule membranes: A randomized controlled trial. PLoS ONE 2021, 16, e0251293. [Google Scholar] [CrossRef]
- Timby, N.; Hernell, O.; Vaarala, O.; Melin, M.; Lönnerdal, B.; Domellöf, M. Infections in Infants Fed Formula Supplemented With Bovine Milk Fat Globule Membranes. J. Pediatr. Gastroenterol. Nutr. 2015, 60, 384–389. [Google Scholar] [CrossRef]
- El Alwani, M.; Wu, B.X.; Obeid, L.M.; Hannun, Y.A. Bioactive sphingolipids in the modulation of the inflammatory response. Pharmacol. Ther. 2006, 112, 171–183. [Google Scholar] [CrossRef]
- Park, E.J.; Suh, M.; Thomson, B.; Ma, D.W.L.; Ramanujam, K.; Thomson, A.B.R.; Clandinin, M.T. Dietary ganglioside inhibits acute inflammatory signals in intestinal mucosa and blood induced by systemic inflammation of Escherichia coli lipopolysaccharide. Shock 2007, 28, 112–117. [Google Scholar] [CrossRef]
- Snow, D.R.; Ward, R.E.; Olsen, A.; Jimenez-Flores, R.; Hintze, K.J. Membrane-rich milk fat diet provides protection against gastrointestinal leakiness in mice treated with lipopolysaccharide. J. Dairy Sci. 2011, 94, 2201–2212. [Google Scholar] [CrossRef] [Green Version]
- Bæk, O.; Ren, S.; Brunse, A.; Sangild, P.T.; Nguyen, D.N. Impaired neonatal immunity and infection resistance following fetal growth restriction in preterm pigs. Front. Immunol. 2020, 11, 1808. [Google Scholar] [CrossRef] [PubMed]
- Bæk, O.; Sangild, P.T.; Thymann, T.; Nguyen, D.N. Growth Restriction and Systemic Immune Development in Preterm Piglets. Front. Immunol. 2019, 10, 2402. [Google Scholar] [CrossRef]
- Amdi, C.; Lynegaard, J.C.; Thymann, T.; Williams, A.R. Intrauterine growth restriction in piglets alters blood cell counts and impairs cytokine responses in peripheral mononuclear cells 24 days post-partum. Sci. Rep. 2020, 10, 4683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, S.; Hui, Y.; Obelitz-Ryom, K.; Brandt, A.B.; Kot, W.; Nielsen, D.S.; Thymann, T.; Sangild, P.T.; Nguyen, D.N. Neonatal gut and immune maturation is determined more by postnatal age than by postconceptional age in moderately preterm pigs. Am. J. Physiol. Liver Physiol. 2018, 315, G855–G867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heegaard, P.M.H.; Pedersen, H.G.; Jensen, A.L.; Boas, U. A robust quantitative solid phase immunoassay for the acute phase protein C-reactive protein (CRP) based on cytidine 5′-diphosphocholine coupled dendrimers. J. Immunol. Methods 2009, 343, 112–118. [Google Scholar] [CrossRef] [PubMed]
- Burrin, D.; Sangild, P.T.; Stoll, B.; Thymann, T.; Buddington, R.; Marini, J.; Olutoye, O.; Shulman, R.J. Translational Advances in Pediatric Nutrition and Gastroenterology: New Insights from Pig Models. Annu. Rev. Anim. Biosci. 2020, 8, 321–354. [Google Scholar] [CrossRef] [Green Version]
- Bæk, O.; Fabiansen, C.; Friis, H.; Ritz, C.; Koch, J.; Willesen, J.L.; Heegaard, P.M.H.; Lykkesfeldt, J.; Briend, A.; Golden, M.H.; et al. Malnutrition Predisposes to Endotoxin-Induced Edema and Impaired Inflammatory Response in Parenterally Fed Piglets. J. Parenter. Enter. Nutr. 2019, 44, 668–676. [Google Scholar] [CrossRef]
- Amdi, C.; Pedersen, M.L.M.; Klaaborg, J.; Myhill, L.J.; Engelsmann, M.N.; Williams, A.R.; Thymann, T. Pre-weaning adaptation responses in piglets fed milk replacer with gradually increasing amounts of wheat. Br. J. Nutr. 2020, 126, 375–382. [Google Scholar] [CrossRef]
- Sorensen, N.S.; Skovgaard, K.; Heegaard, P.M. Porcine blood mononuclear cell cytokine responses to PAMP molecules: Comparison of mRNA and protein production. Vet. Immunol. Immunopathol. 2011, 139, 296–302. [Google Scholar] [CrossRef]
- Meisenberg, G. Principles of Medical Biochemistry, 2nd ed.; Elsevier: Amsterdam, The Netherlands, 2006. [Google Scholar]
- Mwaniki, M.K.; Atieno, M.; Lawn, J.E.; Newton, C.R.J.C. Long-term neurodevelopmental outcomes after intrauterine and neonatal insults: A systematic review. Lancet 2012, 379, 445–452. [Google Scholar] [CrossRef] [Green Version]
- Adams-Chapman, I. Long-Term Impact of Infection on the Preterm Neonate. Semin. Perinatol. 2012, 36, 462–470. [Google Scholar] [CrossRef] [PubMed]
- Lausten-Thomsen, U.; Olsen, M.; Greisen, G.; Schmiegelow, K. Inflammatory markers in umbilical cord blood from small-for-gestational-age newborns. Fetal Pediatr. Pathol. 2014, 33, 114–118. [Google Scholar] [CrossRef] [PubMed]
- Amarilyo, G.; Oren, A.; Mimouni, F.B.; Ochshorn, Y.; Deutsch, V.; Mandel, D. Increased cord serum inflammatory markers in small-for-gestational-age neonates. J. Perinatol. 2011, 31, 30–32. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein | Gene | Forward Sequence (5′ to 3′) | Reverse Sequence (5′ to 3′) |
|---|---|---|---|
| C-X-C motif chemokine ligand 9 | CXCL9 | GAAAAGCAGTGTTGCCTTGCT | TGATGCAGGAACAACGTCCAT |
| C-X-C motif chemokine ligand 10 | CXCL10 | ATCATCCCGAGCTGTTGAGC | CCAGGACTTGGCACATTCAC |
| GATA binding protein 3 | GATA3 | ACCCCTTATTAAGCCCAAGC | TCCAGAGAGTCGTCGTTGTG |
| Hypoxia inducible factor 1 alpha | HIF1A | TGTGTTATCTGTCGCTTTGAGTC | TTTCGCTTTCTCTGAGCATTC |
| Hexokinase 1 | HK1 | TTTCCCTTGTCGGCAATCCA | CCTCCACTCCGCTTGCTTTA |
| Hypoxanthine phosphoribosyltransferase 1 | HPRT1 | TATGGACAGGACTGAACGGC | ACACAGAGGGCTACGATGTG |
| Interferon gamma | IFNG | AGCTTTGCGTGACTTTGTGT | ATGCTCCTTTGAATGGCCTG |
| Interleukin 2 | IL2 | AAGCTCTGGAGGGAGTGCTA | CAACAGCAGTTACTGTCTCATCA |
| Interleukin 4 | IL4 | GTACCAGCAACTTCGTCCAC | CCTTCTCCGTCGTGTTCTCT |
| Interleukin 6 | IL6 | TGCCACCTCAGACAAAATGC | AGGTTCAGGTTGTTTTCTGCC |
| Interleukin 10 | IL10 | GTCCGACTCAACGAAGAAGG | GCCAGGAAGATCAGGCAATA |
| Interleukin 17 | IL17 | GCACACGGGCTGCATCAACG | TGCAACCAACAGTGACCCGCA |
| Myeloperoxidase | MPO | CCCGAGTTGCTTTCCTCACT | AAGAAGGGGATGCAGTCACG |
| Pyruvate dehydrogenase α1 | PDHA1 | GTCAGGAAGCTTGTTGCGTG | GGTAAAGCCATGAGCTCGGT |
| Pyruvate kinase | PKM | GCCCTGGACACTAAAGGACC | CAGCCACAGGACATTCTCGT |
| Peroxisome proliferator-activated receptor gamma | PPARG | TGACCATGGTTGACACCGAG | GATCAGCTCTCGGGAATGGG |
| RAR-related orphan receptor alpha | RORA | CAGCGCTCCAACATCTTCTC | GACCAGCACCACTTCCATTG |
| S100 calcium binding protein A9 | S100A9 | GCCAAACTTTCTCAAGAAGCA | AGTGTCCAGGTCTTCCAGGAT |
| T-box transcription factor | TBET | CTGAGAGTCGCGCTCAACAA | ACCCGGCCACAGTAAATGAC |
| Transforming growth factor beta 1 | TGFB1 | GCAAGGTCCTGGCTCTGTA | TAGTACACGATGGGCAGTGG |
| Toll-like receptor 2 | TLR2 | CGTGTGCTATGACGCTTTCG | GTACTTGCACCACTCGCTCT |
| Toll-like receptor 4 | TLR4 | TGGTGTCCCAGCACTTCATA | CAACTTCTGCAGGACGATGA |
| Tumor necrosis factor alpha | TNFA | ATTCAGGGATGTGTGGCCTG | CCAGATGTCCCAGGTTGCAT |
| Day | VEG (n = 18) | MILK (n = 18) | NBW (n = 18) | IUGR (n = 18) | |
|---|---|---|---|---|---|
| Red blood cells (1012 cells/L) | 2 | 4.5 (0.3) | 3.8 (0.3) | 3.9 (0.2) | 4.4 (0.4) |
| 11 | 5.6 (0.2) | 5.6 (0.1) | 5.7 (0.1) | 5.5 (0.2) | |
| 23/24 | 5.8 (0.1) | 5.4 (0.1) | 5.9 (0.1) | 5.3 (0.1) *** | |
| Hemoglobin concentration (g/L) | 2 | 5.3 (0.2) | 4.7 (0.2) | 4.8 (0.2) | 5.2 (0.3) |
| 11 | 6.6 (0.1) | 6.6 (0.1) | 6.6 (0.1) | 6.5 (0.1) | |
| 23/24 | 6.9 (0.1) | 6.3 (0.1) * | 6.9 (0.1) | 6.4 (0.1) *** | |
| Hematocrit (%) | 2 | 27.8 (1.2) | 24.5 (1.6) | 25.5 (1.2) | 26.8 (1.9) |
| 11 | 36.7 (0.9) | 36.6 (0.6) | 36.8 (0.7) | 36.6 (0.8) *** | |
| 23/24 | 37.2 (0.7) | 34.1 (0.8) * | 37.2 (0.8) | 34.1 (0.8) *** | |
| Platelets (109 cells/L) | 2 | 176 (26) | 205 (58) | 194 (54) | 187 (36) |
| 1 | 364 (26) | 415 (38) | 404 (35) | 376 (31) | |
| 23/24 | 259 (23) | 317 (19) * | 315 (21) | 261 (22) (*) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bæk, O.; Skadborg, K.; Muk, T.; Amdi, C.; Heegaard, P.M.H.; Thymann, T.; Nguyen, D.N. Infant Formula Based on Milk Fat Affects Immune Development in Both Normal Birthweight and Fetal Growth Restricted Neonatal Piglets. Nutrients 2021, 13, 3310. https://doi.org/10.3390/nu13103310
Bæk O, Skadborg K, Muk T, Amdi C, Heegaard PMH, Thymann T, Nguyen DN. Infant Formula Based on Milk Fat Affects Immune Development in Both Normal Birthweight and Fetal Growth Restricted Neonatal Piglets. Nutrients. 2021; 13(10):3310. https://doi.org/10.3390/nu13103310
Chicago/Turabian StyleBæk, Ole, Karina Skadborg, Tik Muk, Charlotte Amdi, Peter M. H. Heegaard, Thomas Thymann, and Duc Ninh Nguyen. 2021. "Infant Formula Based on Milk Fat Affects Immune Development in Both Normal Birthweight and Fetal Growth Restricted Neonatal Piglets" Nutrients 13, no. 10: 3310. https://doi.org/10.3390/nu13103310
APA StyleBæk, O., Skadborg, K., Muk, T., Amdi, C., Heegaard, P. M. H., Thymann, T., & Nguyen, D. N. (2021). Infant Formula Based on Milk Fat Affects Immune Development in Both Normal Birthweight and Fetal Growth Restricted Neonatal Piglets. Nutrients, 13(10), 3310. https://doi.org/10.3390/nu13103310