KiwiC for Vitality: Results of a Placebo-Controlled Trial Testing the Effects of Kiwifruit or Vitamin C Tablets on Vitality in Adults with Low Vitamin C Levels

 , , , and
, , , and 
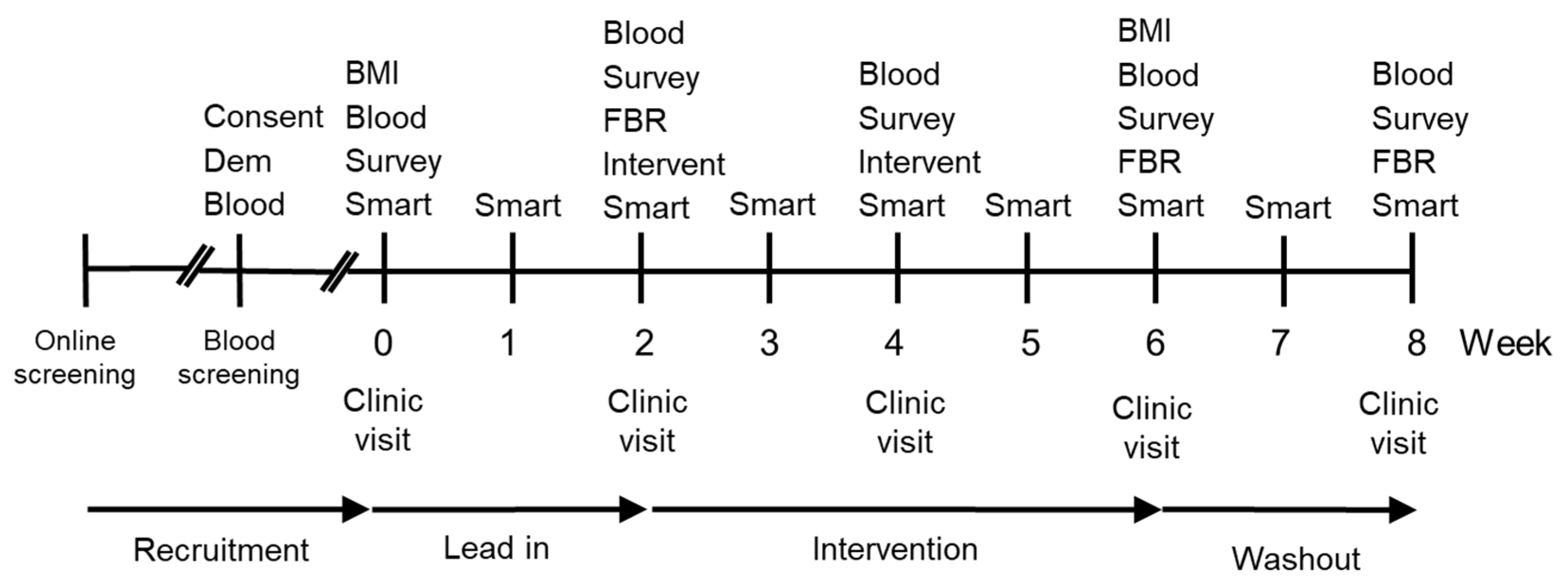{kind=link}
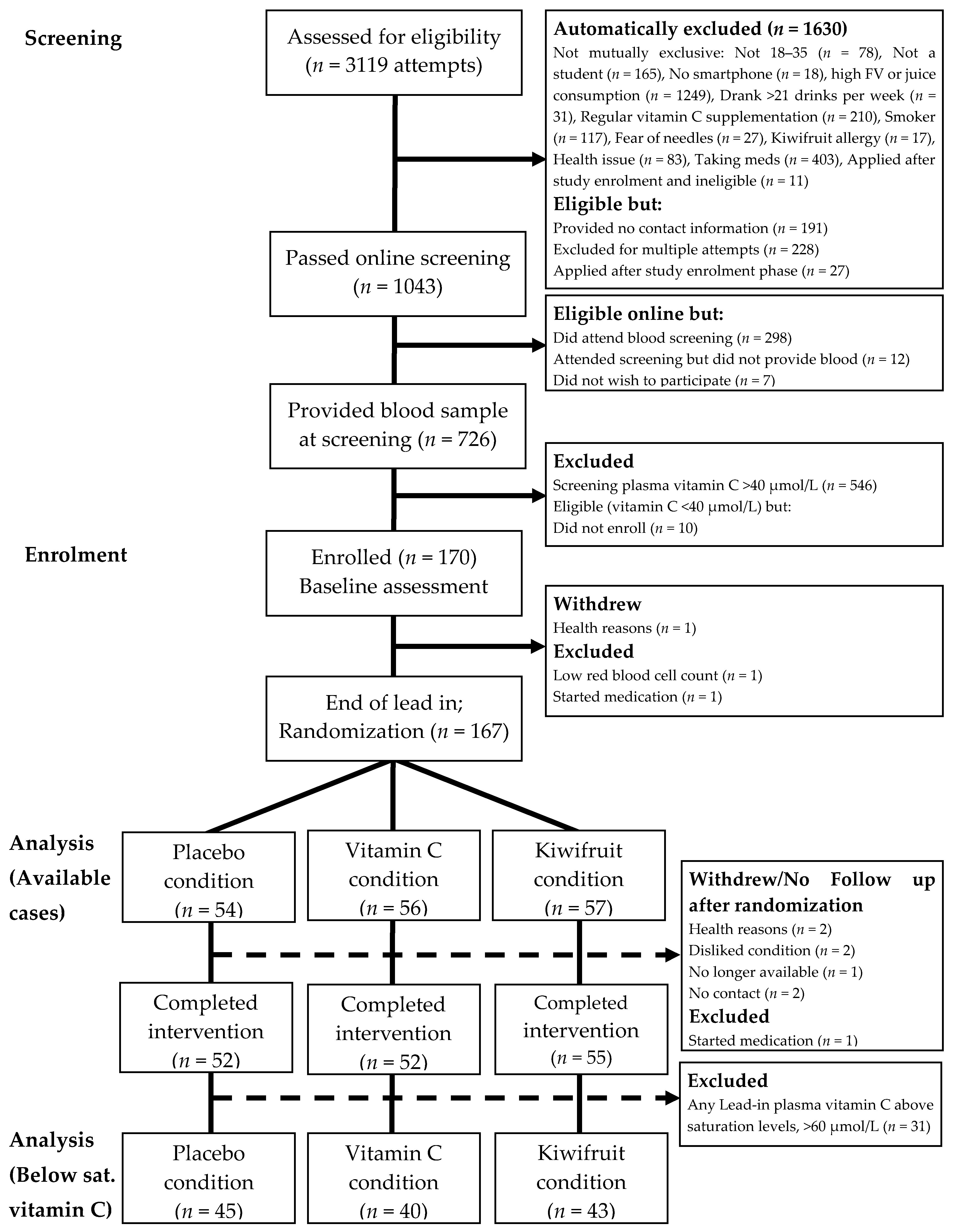{kind=link}
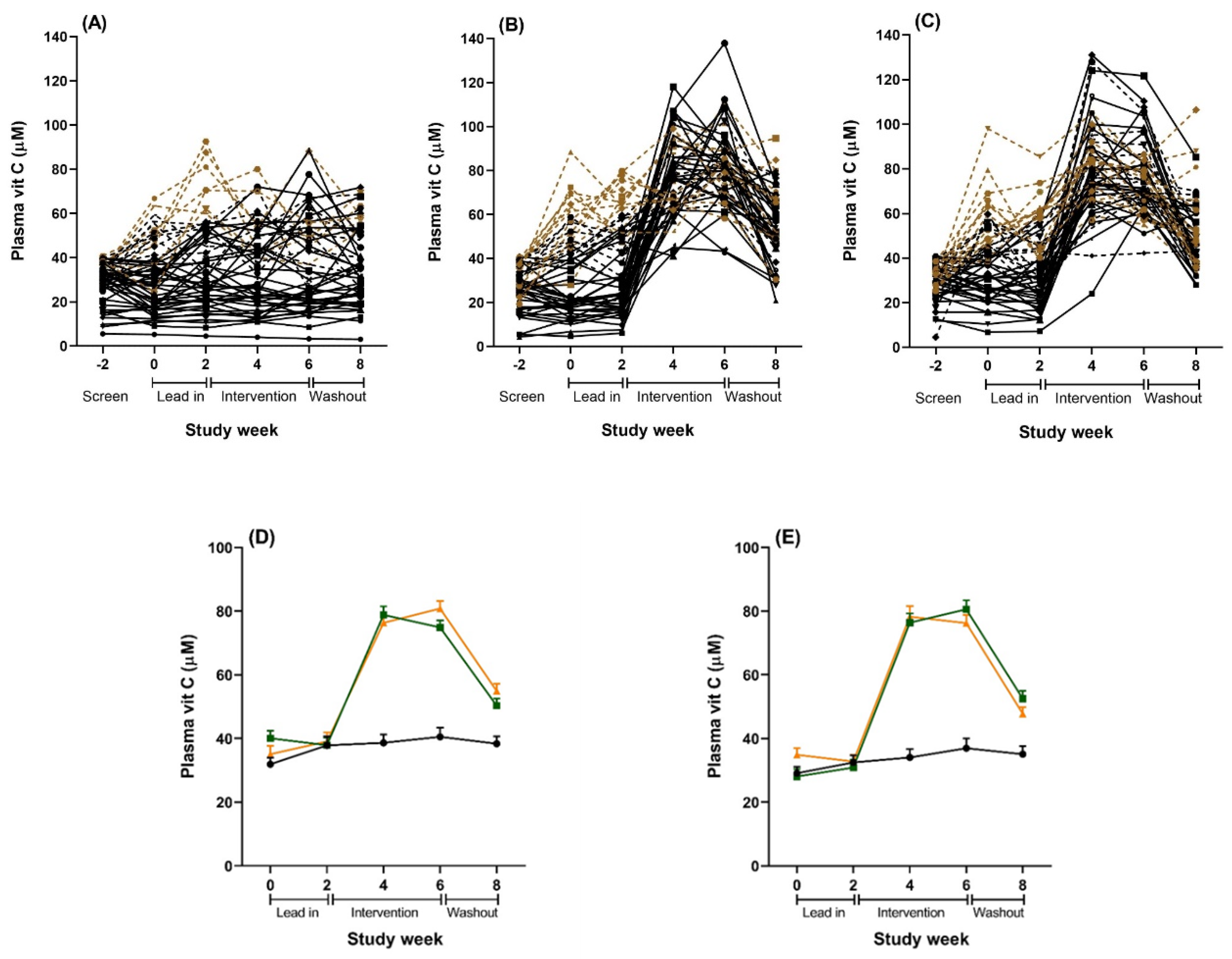{kind=link}
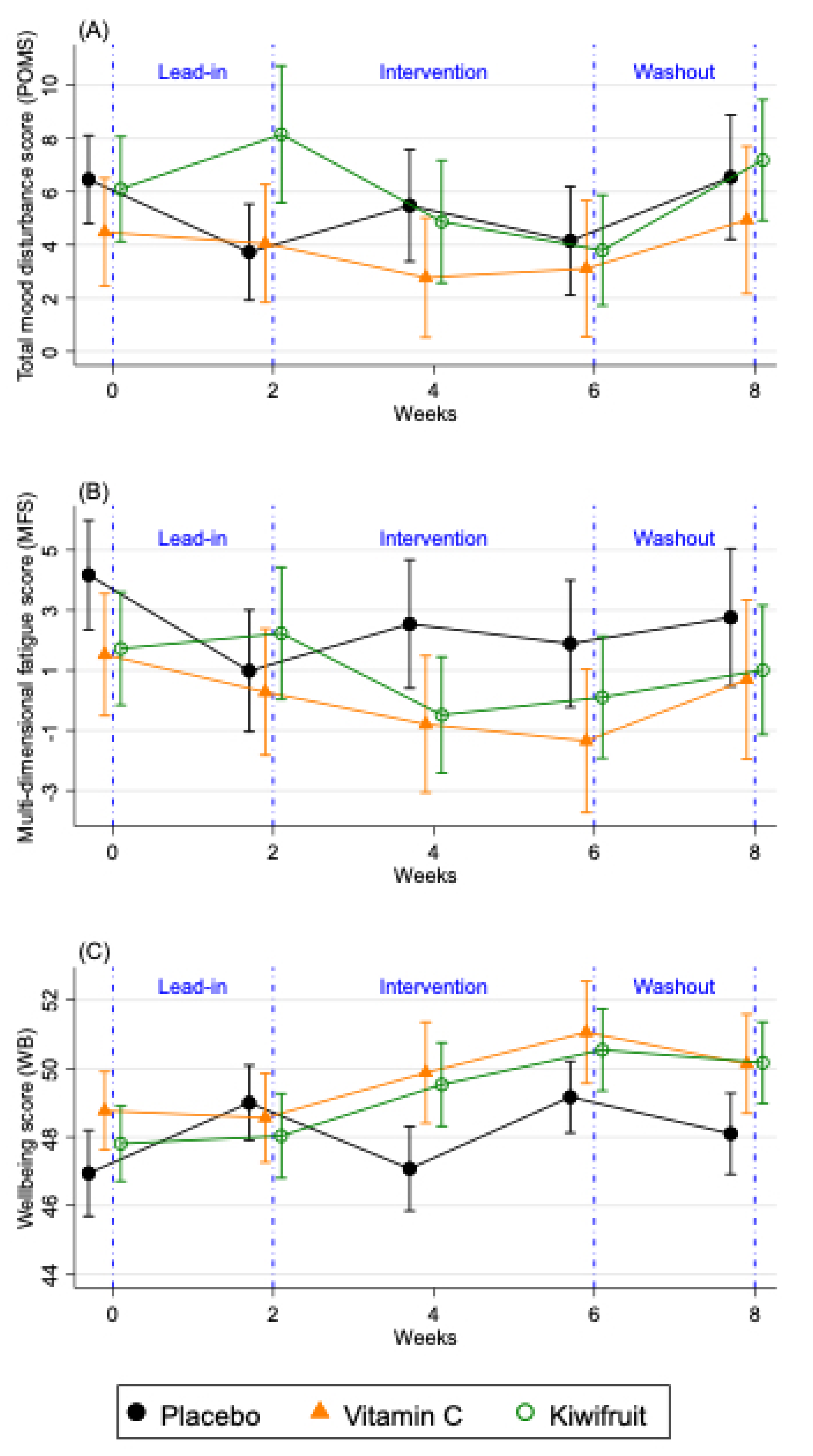{kind=link}
Abstract
Share and Cite
Conner, T.S.; Fletcher, B.D.; Haszard, J.J.; Pullar, J.M.; Spencer, E.; Mainvil, L.A.; Vissers, M.C.M. KiwiC for Vitality: Results of a Placebo-Controlled Trial Testing the Effects of Kiwifruit or Vitamin C Tablets on Vitality in Adults with Low Vitamin C Levels. Nutrients 2020, 12, 2898. https://doi.org/10.3390/nu12092898
Conner TS, Fletcher BD, Haszard JJ, Pullar JM, Spencer E, Mainvil LA, Vissers MCM. KiwiC for Vitality: Results of a Placebo-Controlled Trial Testing the Effects of Kiwifruit or Vitamin C Tablets on Vitality in Adults with Low Vitamin C Levels. Nutrients. 2020; 12(9):2898. https://doi.org/10.3390/nu12092898
Chicago/Turabian StyleConner, Tamlin S., Benjamin D. Fletcher, Jillian J. Haszard, Juliet M. Pullar, Emma Spencer, Louise A. Mainvil, and Margreet C. M. Vissers. 2020. "KiwiC for Vitality: Results of a Placebo-Controlled Trial Testing the Effects of Kiwifruit or Vitamin C Tablets on Vitality in Adults with Low Vitamin C Levels" Nutrients 12, no. 9: 2898. https://doi.org/10.3390/nu12092898
APA StyleConner, T. S., Fletcher, B. D., Haszard, J. J., Pullar, J. M., Spencer, E., Mainvil, L. A., & Vissers, M. C. M. (2020). KiwiC for Vitality: Results of a Placebo-Controlled Trial Testing the Effects of Kiwifruit or Vitamin C Tablets on Vitality in Adults with Low Vitamin C Levels. Nutrients, 12(9), 2898. https://doi.org/10.3390/nu12092898