Dietary Fermented Soy Extract and Oligo-Lactic Acid Alleviate Chronic Kidney Disease in Mice via Inhibition of Inflammation and Modulation of Gut Microbiota



Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Animal Study
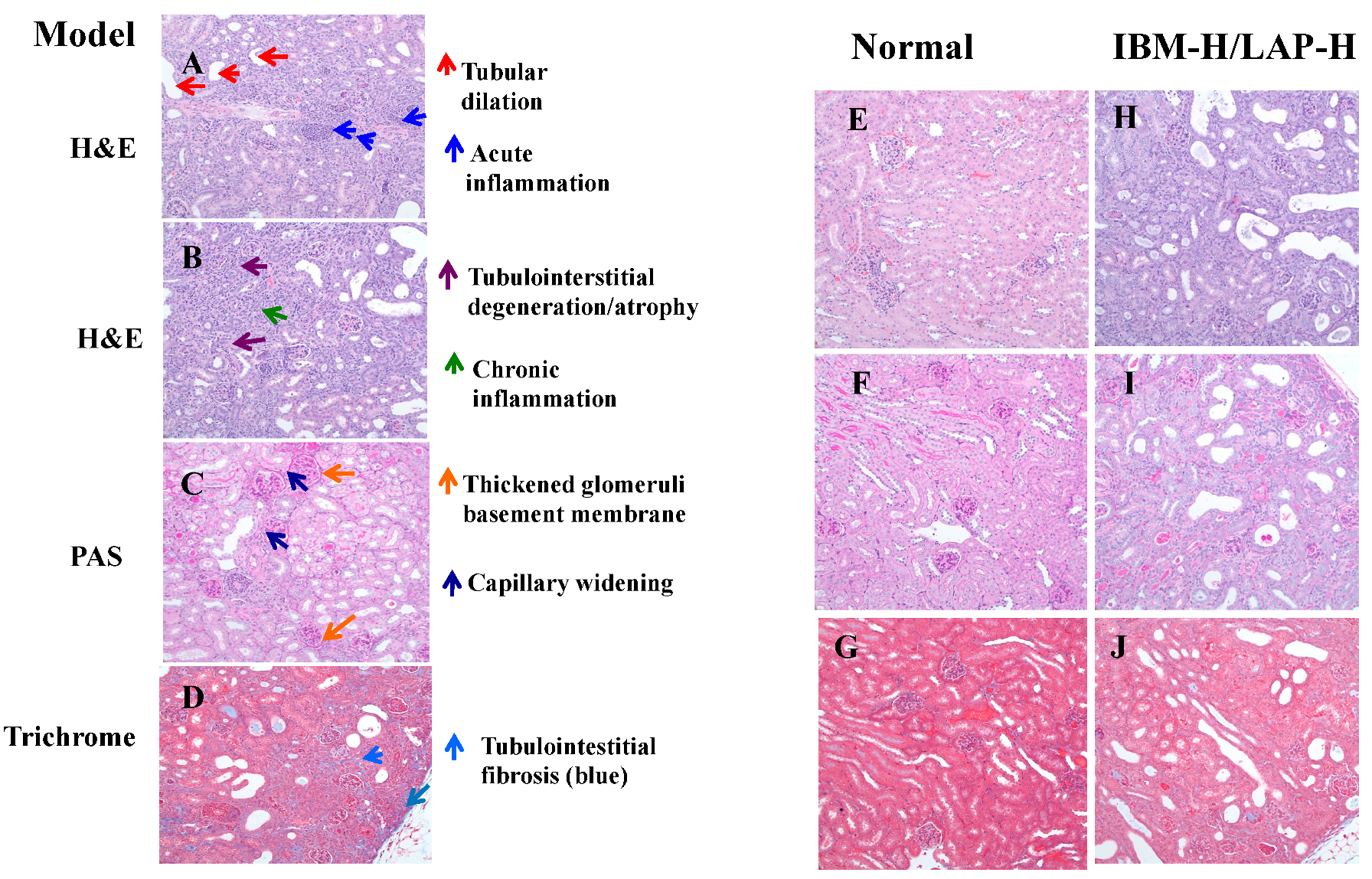
2.3. Histopathology Analysis
2.4. Multiplex Sandwich Immunoassays for Measurement of Blood Levels of Cytokines and Kidney Toxicity Biomarkers
2.5. Quantitative Polymerase Chain Reaction (qPCR) for Determination of Gut Microbiota
2.6. Quantitative Real-Time PCR (qRT-PCR) for Analysis of Inflammation- and Stem Cells-Related Genes in Kidney Tissues
2.7. Statistical Analysis
3. Results
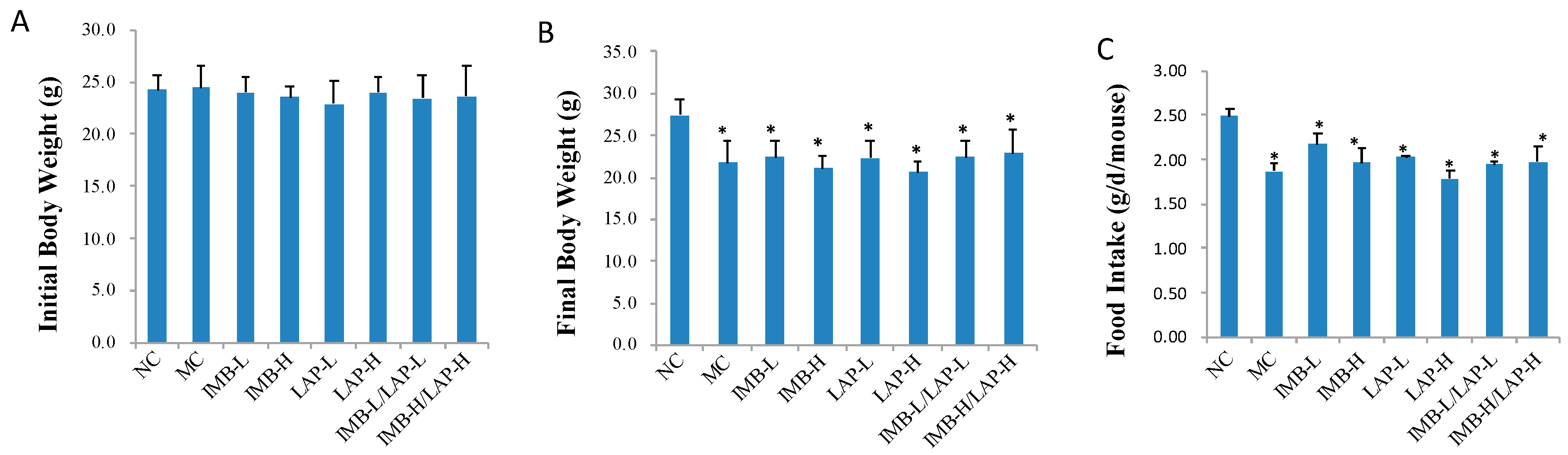
3.1. Effects of IMB and LAP Treatments on Body Weight
3.2. Effects of IMB and LAP Treatments on Kidney Inflammation and Damage
3.3. Effects of IMB and LAP Treatments on Circulating Levels of Cytokines and Kidney Injury Biomarkers
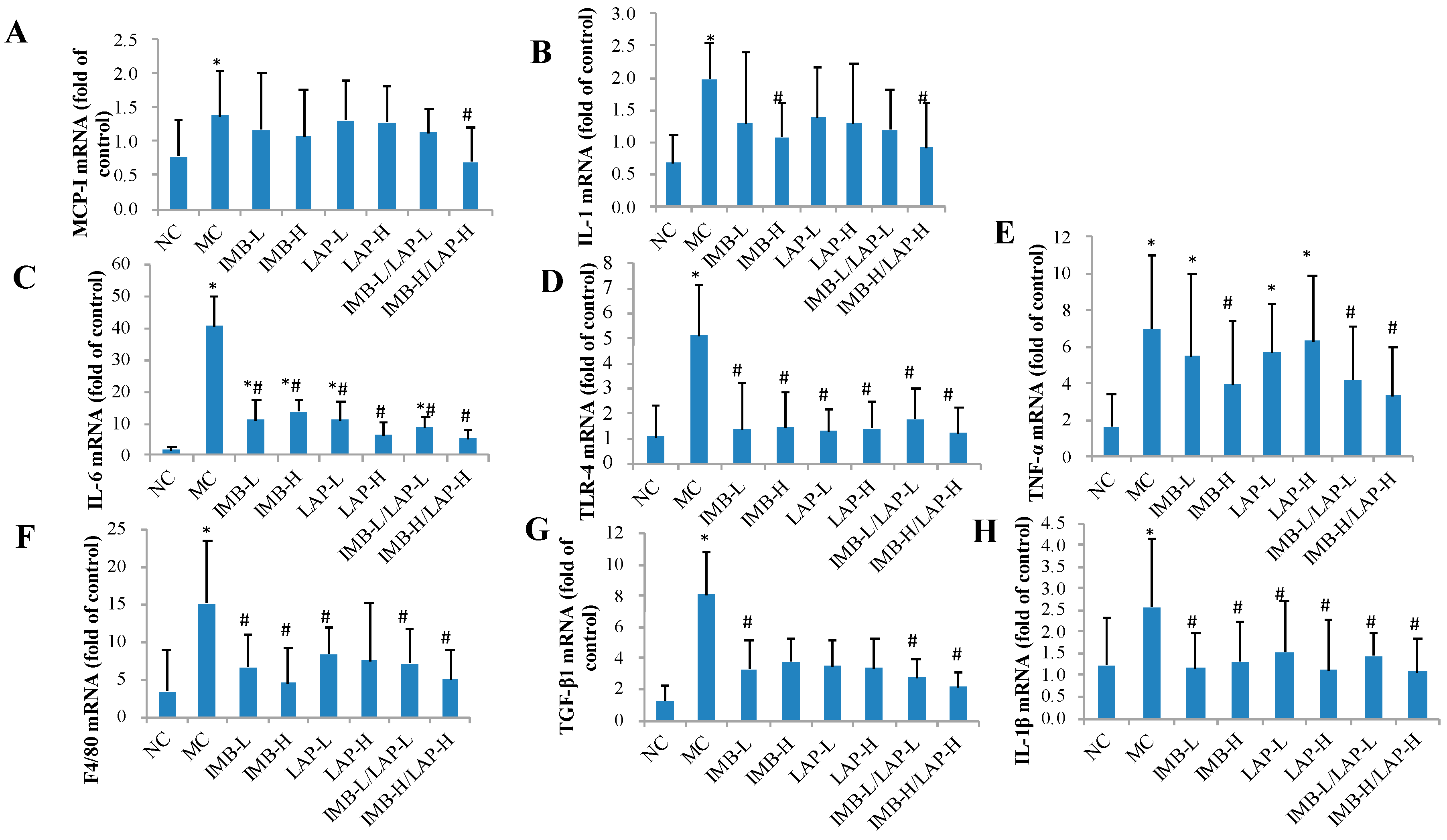
3.4. Effects of IMB and LAP Treatments on the Expression Levels of Inflammatory Cytokines in Kidney
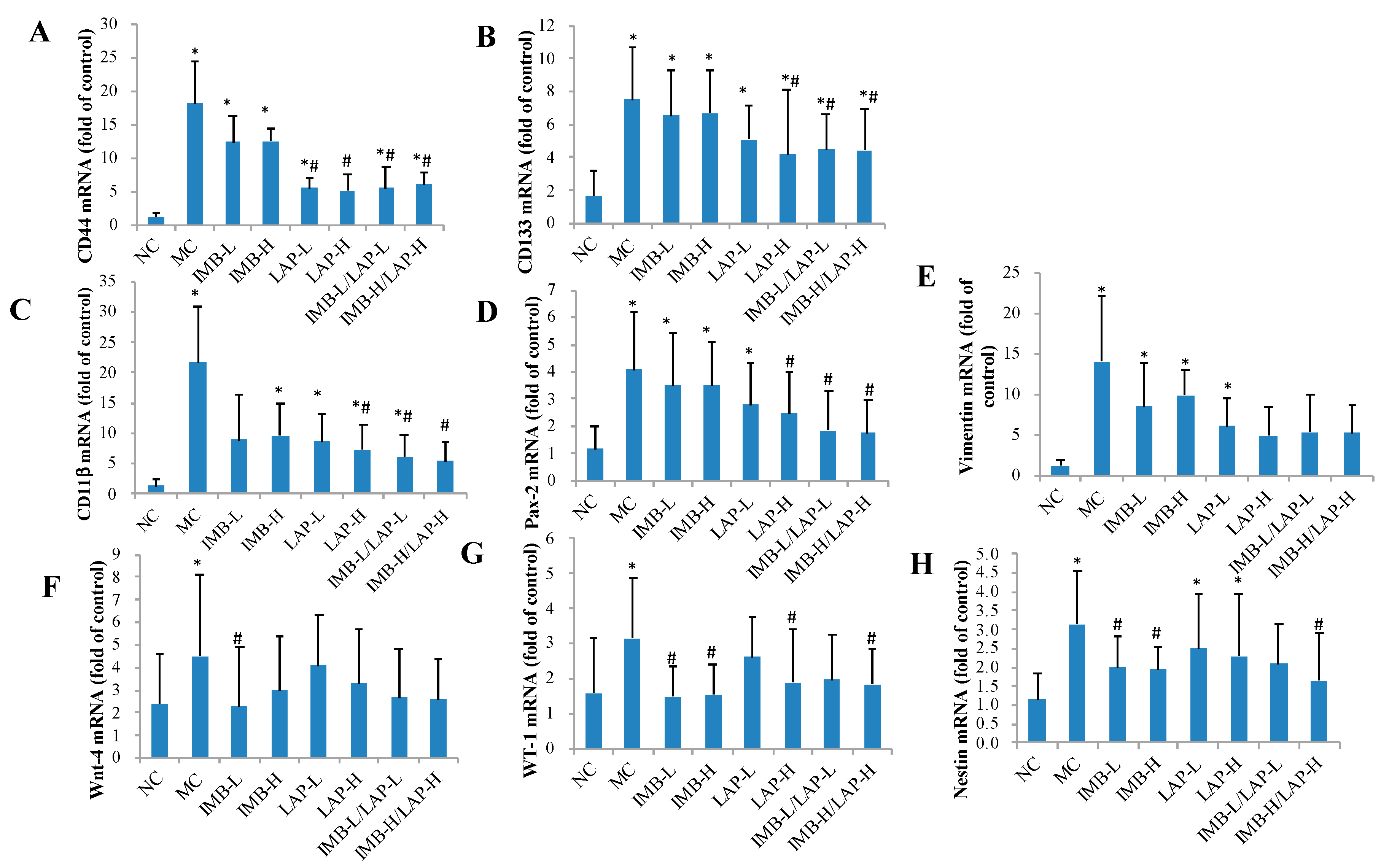
3.5. Effects of IMB and LAP Treatments on the Expression Levels of Stem Cell-Related Genes in Kidney
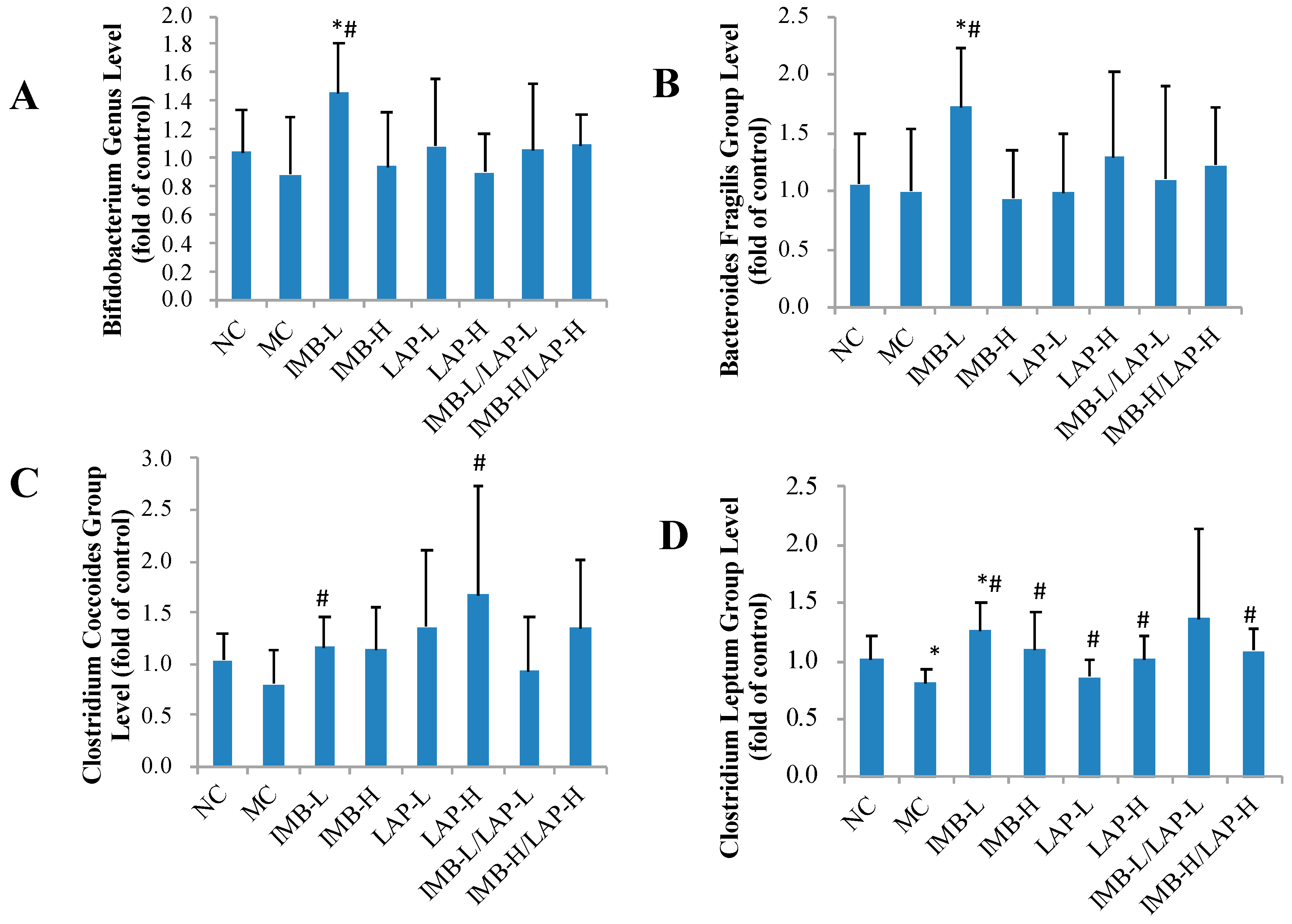
3.6. Effects of IMB and LAP Treatments on Gut Microbiota
3.7. Correlation Analysis between Histopathological and Metabolic Parameters
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Centers for Disease Control and Prevention. Chronic Kidney Disease in the United States. 2019. Available online: https://www.cdc.gov/kidneydisease/publications-resources/2019-national-facts.html (accessed on 8 June 2020).
- GBD Chronic Kidney Disease Collaboration. Global, regional, and national burden of chronic kidney disease, 1990–2017: A systematic analysis for the Global Burden of Disease Study 2017. Lancet 2020, 395, 709–733. [Google Scholar] [CrossRef]
- Peyster, E.; Chen, J.; Feldman, H.I.; Go, A.S.; Gupta, J.; Mitra, N.; Pan, Q.; Porter, A.; Rahman, M.; Raj, D.; et al. Inflammation and Arterial Stiffness in Chronic Kidney Disease: Findings From the CRIC Study. Am. J. Hypertens. 2017, 30, 400–408. [Google Scholar] [CrossRef]
- Grabulosa, C.C.; Manfredi, S.R.; Canziani, M.E.; Quinto, B.M.R.; Barbosa, R.B.; Rebello, J.F.; Batista, M.C.; Cendoroglo, M.; Dalboni, M.A. Chronic kidney disease induces inflammation by increasing Toll-like receptor-4, cytokine and cathelicidin expression in neutrophils and monocytes. Exp. Cell Res. 2018, 365, 157–162. [Google Scholar] [CrossRef]
- Mihai, S.; Codrici, E.; Popescu, I.D.; Enciu, A.M.; Albulescu, L.; Necula, L.G.; Mambet, C.; Anton, G.; Tanase, C. Inflammation-Related Mechanisms in Chronic Kidney Disease Prediction, Progression, and Outcome. J. Immunol. Res. 2018, 2018, 2180373. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Chen, X.; Song, Y.; Caballero, B.; Cheskin, L.J. Association between obesity and kidney disease: A systematic review and meta-analysis. Kidney Int. 2008, 73, 19–33. [Google Scholar] [CrossRef] [PubMed]
- Messina, M. Soy and Health Update: Evaluation of the Clinical and Epidemiologic Literature. Nutrients 2016, 8, 754. [Google Scholar] [CrossRef] [PubMed]
- Anderson, J.W. Beneficial effects of soy protein consumption for renal function. Asia Pac. J. Clin. Nutr. 2008, 17 (Suppl. 1), 324–328. [Google Scholar] [PubMed]
- Anderson, J.W.; Blake, J.E.; Turner, J.; Smith, B.M. Effects of soy protein on renal function and proteinuria in patients with type 2 diabetes. Am. J. Clin. Nutr. 1998, 68, 1347S–1353S. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Yuan, W.-J. Effects of soy protein containing isoflavones in patients with chronic kidney disease: A systematic review and meta-analysis. Clin. Nutr. 2016, 35, 117–124. [Google Scholar] [CrossRef]
- Sun, S.; Li, H.; Chen, J.; Qian, Q. Lactic Acid: No Longer an Inert and End-Product of Glycolysis. Physiology 2017, 32, 453–463. [Google Scholar] [CrossRef]
- Akchurin, O.; Sureshbabu, A.; Doty, S.B.; Zhu, Y.S.; Patino, E.; Cunningham-Rundles, S.; Choi, M.E.; Boskey, A.; Rivella, S. Lack of hepcidin ameliorates anemia and improves growth in an adenine-induced mouse model of chronic kidney disease. Am. J. Physiol. Ren. Physiol. 2016, 311, F877–F889. [Google Scholar] [CrossRef] [PubMed]
- Rahman, A.; Yamazaki, D.; Sufiun, A.; Kitada, K.; Hitomi, H.; Nakano, D.; Nishiyama, A. A novel approach to adenine-induced chronic kidney disease associated anemia in rodents. PLoS ONE 2018, 13, e0192531. [Google Scholar] [CrossRef] [PubMed]
- Kwok, L.Y.; Zhang, J.; Guo, Z.; Gesudu, Q.; Zheng, Y.; Qiao, J.; Huo, D.; Zhang, H. Characterization of fecal microbiota across seven Chinese ethnic groups by quantitative polymerase chain reaction. PLoS ONE 2014, 9, e93631. [Google Scholar] [CrossRef] [PubMed]
- Silverstein, D.M. Inflammation in chronic kidney disease: Role in the progression of renal and cardiovascular disease. Pediatric Nephrol. 2009, 24, 1445–1452. [Google Scholar] [CrossRef]
- Ali, B.H.; Adham, S.A.; Al Za’abi, M.; Waly, M.I.; Yasin, J.; Nemmar, A.; Schupp, N. Ameliorative effect of chrysin on adenine-induced chronic kidney disease in rats. PLoS ONE 2015, 10, e0125285. [Google Scholar] [CrossRef]
- Al Za’abi, M.; Al Busaidi, M.; Yasin, J.; Schupp, N.; Nemmar, A.; Ali, B.H. Development of a new model for the induction of chronic kidney disease via intraperitoneal adenine administration, and the effect of treatment with gum acacia thereon. Am. J. Transl. Res. 2015, 7, 28–38. [Google Scholar]
- Tummalapalli, L.; Nadkarni, G.N.; Coca, S.G. Biomarkers for predicting outcomes in chronic kidney disease. Curr. Opin. Nephrol. Hypertens 2016, 25, 480–486. [Google Scholar] [CrossRef]
- Kim, K.; Park, B.H.; Ihm, H.; Kim, K.M.; Jeong, J.; Chang, J.W.; Cho, Y.M. Expression of stem cell marker CD133 in fetal and adult human kidneys and pauci-immune crescentic glomerulonephritis. Histol. Histopathol. 2011, 26, 223–232. [Google Scholar] [CrossRef]
- Ward, H.H.; Romero, E.; Welford, A.; Pickett, G.; Bacallao, R.; Gattone, V.H., 2nd; Ness, S.A.; Wandinger-Ness, A.; Roitbak, T. Adult human CD133/1(+) kidney cells isolated from papilla integrate into developing kidney tubules. Biochim. Biophys. Acta 2011, 1812, 1344–1357. [Google Scholar] [CrossRef]
- Brossa, A.; Papadimitriou, E.; Collino, F.; Incarnato, D.; Oliviero, S.; Camussi, G.; Bussolati, B. Role of CD133 Molecule in Wnt Response and Renal Repair. Stem Cells Transl. Med. 2018, 7, 283–294. [Google Scholar] [CrossRef]
- Naiman, N.; Fujioka, K.; Fujino, M.; Valerius, M.T.; Potter, S.S.; McMahon, A.P.; Kobayashi, A. Repression of Interstitial Identity in Nephron Progenitor Cells by Pax2 Establishes the Nephron-Interstitium Boundary during Kidney Development. Dev. Cell 2017, 41, 349–365.e3. [Google Scholar] [CrossRef] [PubMed]
- Eng, D.G.; Sunseri, M.W.; Kaverina, N.V.; Roeder, S.S.; Pippin, J.W.; Shankland, S.J. Glomerular parietal epithelial cells contribute to adult podocyte regeneration in experimental focal segmental glomerulosclerosis. Kidney Int. 2015, 88, 999–1012. [Google Scholar] [CrossRef] [PubMed]
- Lewington, A.J.; Padanilam, B.J.; Martin, D.R.; Hammerman, M.R. Expression of CD44 in kidney after acute ischemic injury in rats. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2000, 278, R247–R254. [Google Scholar] [CrossRef] [PubMed]
- Rouschop, K.M.; Sewnath, M.E.; Claessen, N.; Roelofs, J.J.; Hoedemaeker, I.; van der Neut, R.; Aten, J.; Pals, S.T.; Weening, J.J.; Florquin, S. CD44 deficiency increases tubular damage but reduces renal fibrosis in obstructive nephropathy. J. Am. Soc. Nephrol. 2004, 15, 674–686. [Google Scholar] [CrossRef]
- Yang, G.; Cheng, Q.L.; Li, C.L.; Jia, Y.L.; Wang, J.X.; Yue, W.; Pei, X.T. Isolation and Identification of Rat Kidney Stem Cells. J. Sichuan Univ. 2015, 46, 667–672. [Google Scholar]
- Kreidberg, J.A. WT1 and kidney progenitor cells. Organogenesis 2010, 6, 61–70. [Google Scholar] [CrossRef]
- Cosola, C.; Rocchetti, M.T.; Sabatino, A.; Fiaccadori, E.; Di Iorio, B.R.; Gesualdo, L. Microbiota issue in CKD: How promising are gut-targeted approaches? J. Nephrol. 2019, 32, 27–37. [Google Scholar] [CrossRef]
- Wing, M.R.; Patel, S.S.; Ramezani, A.; Raj, D.S. Gut microbiome in chronic kidney disease. Exp. Physiol. 2016, 101, 471–477. [Google Scholar] [CrossRef]
- Kikuchi, M.; Ueno, M.; Itoh, Y.; Suda, W.; Hattori, M. Uremic Toxin-Producing Gut Microbiota in Rats with Chronic Kidney Disease. Nephron 2017, 135, 51–60. [Google Scholar] [CrossRef]
- Lau, W.L.; Kalantar-Zadeh, K.; Vaziri, N.D. The Gut as a Source of Inflammation in Chronic Kidney Disease. Nephron 2015, 130, 92–98. [Google Scholar] [CrossRef]
- Lau, W.L.; Vaziri, N.D. The Leaky Gut and Altered Microbiome in Chronic Kidney Disease. J. Ren. Nutr. Off. J. Counc. Ren. Nutr. Natl. Kidney Found. 2017, 27, 458–461. [Google Scholar] [CrossRef] [PubMed]
- Nallu, A.; Sharma, S.; Ramezani, A.; Muralidharan, J.; Raj, D. Gut microbiome in chronic kidney disease: Challenges and opportunities. Transl. Res. J. Lab. Clin. Med. 2017, 179, 24–37. [Google Scholar] [CrossRef] [PubMed]
- Sampaio-Maia, B.; Simoes-Silva, L.; Pestana, M.; Araujo, R.; Soares-Silva, I.J. The Role of the Gut Microbiome on Chronic Kidney Disease. Adv. Appl. Microbiol. 2016, 96, 65–94. [Google Scholar] [CrossRef] [PubMed]
- Yoshifuji, A.; Wakino, S.; Irie, J.; Tajima, T.; Hasegawa, K.; Kanda, T.; Tokuyama, H.; Hayashi, K.; Itoh, H. Gut Lactobacillus protects against the progression of renal damage by modulating the gut environment in rats. Nephrol. Dial. Transplant. 2016, 31, 401–412. [Google Scholar] [CrossRef]
- Lippi, I.; Perondi, F.; Ceccherini, G.; Marchetti, V.; Guidi, G. Effects of probiotic VSL#3 on glomerular filtration rate in dogs affected by chronic kidney disease: A pilot study. Can. Vet. J. 2017, 58, 1301–1305. [Google Scholar]
- Takagi, T.; Naito, Y.; Higashimura, Y.; Ushiroda, C.; Mizushima, K.; Ohashi, Y.; Yasukawa, Z.; Ozeki, M.; Tokunaga, M.; Okubo, T.; et al. Partially hydrolysed guar gum ameliorates murine intestinal inflammation in association with modulating luminal microbiota and SCFA. Br. J. Nutr. 2016, 116, 1199–1205. [Google Scholar] [CrossRef]
- Vrakas, S.; Mountzouris, K.C.; Michalopoulos, G.; Karamanolis, G.; Papatheodoridis, G.; Tzathas, C.; Gazouli, M. Intestinal Bacteria Composition and Translocation of Bacteria in Inflammatory Bowel Disease. PLoS ONE 2017, 12, e0170034. [Google Scholar] [CrossRef]
- Hirano, A.; Umeno, J.; Okamoto, Y.; Shibata, H.; Ogura, Y.; Moriyama, T.; Torisu, T.; Fujioka, S.; Fuyuno, Y.; Kawarabayasi, Y.; et al. Comparison of the microbial community structure between inflamed and non-inflamed sites in patients with ulcerative colitis. J. Gastroenterol. Hepatol. 2018, 33, 1590–1597. [Google Scholar] [CrossRef]
- Sawin, E.A.; De Wolfe, T.J.; Aktas, B.; Stroup, B.M.; Murali, S.G.; Steele, J.L.; Ney, D.M. Glycomacropeptide is a prebiotic that reduces Desulfovibrio bacteria, increases cecal short-chain fatty acids, and is anti-inflammatory in mice. Am. J. Physiol. Gastrointest. Liver Physiol. 2015, 309, G590–G601. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Forward | Reverse |
|---|---|---|
| IL-6 | GAGGATACCACTCCCAACAGACC | AAGTGCATCATCGTTGTTCATACA |
| TNF-α | CATGAGCACAGAAAGCATGATCCG | AAGCAGGAATGAGAAGAGGCTGAG |
| TGF-β1 | TACCATGCCAACTTCTGTCTGGGA | TGTGTTGGTTGTAGAGGGCAAGGA |
| F4/80 | AAGCATCCGAGACACACACAGTCT | TGACTGTACCCACATGGCTGATGA |
| MCP-1 | CTGGTCCGAGTGAGACAAAG | AGATCAGGCTCTGATGGAGAA |
| TLR-4 | CTGCAATGGACAAGGACCA | TCCCACTCCAGGTAAGTG |
| IL-1β | TAATACGACTCACTATAGGG | ATTTAGGTGACACTATAG |
| IL-1 | CATCCGCAAAGTGGTACGA | AGAAAGACTCCACCAGCCCAGT |
| CD133 | GCCCAAGCTGGAAGAATATG | CAGCAGAAAGCAGACAATCAA |
| Six2 | TCAATGGCAGTGGCAAGTCG | TCAAGCACGGAAAGCAAGCG |
| Pax-2 | TCCCAGTGTCTCATCCATCA | GTTAGAGGCGCTGGAAACAG |
| Oct-4 | CACGAGTGGAAAGCAACTCA | AGATGGTGGTCTGGCTGAAC |
| CD11b | GACTCAGTGAGCCCCATCAT | AGATCGTCTTGGCAGATGCT |
| Wnt-4 | AGAACTGGAGAAGTGTGGCTGTGACC | TGTATGTGGCTTGAACTGTGCATTCCG |
| WT-1 | ACATCCGACTTCCAAGACAGCACAC | TTGCAGCCAGACCTCTGAAATTCTG |
| CD29 | TGGTCAGCAACGCATATCTGG | GATCCACAAACCGCAACCT |
| CD44 | TCGATTTGAATGTAACCTGCCG | CAGTCCGGGAGATACTGTAGC |
| Vimentin | CGGCTGCGAGAGAAATTGC | CCACTTTCCGTTCAAGGTCAAG |
| NANOG | TCTTCCTGGTCCCCACAGTTT | GCAAGAATAGTTCTCGGGATGAA |
| Nestin | CCCTGAAGTCGAGGAGCTG | CTGCTGCACCTCTAAGCGA |
| Histopathology | Grade | NC | MC | IMB-L | IMB-H | LAP-L | LAP-H | IMB-L/LAP-L | IMB-H/LAP-H |
|---|---|---|---|---|---|---|---|---|---|
| Tubular dilation | 0 | 8 | 0 | 0 | 0 | 0 | 0 | 0 | 2 |
| 1 | 0 | 0 | 0 | 1 | 4 | 1 | 3 | 4 | |
| 2 | 0 | 2 | 3 | 3 | 2 | 2 | 4 | 2 | |
| 3 | 0 | 6 | 3 | 4 | 2 | 5 | 1 | 0 | |
| p value (vs. MC) | <0.05 | <0.05 | <0.05 | <0.05 | |||||
| Tubulointerstitium degeneration/atrophy | 0 | 8 | 0 | 1 | 1 | 0 | 0 | 0 | 2 |
| 1 | 0 | 1 | 1 | 3 | 4 | 0 | 3 | 3 | |
| 2 | 0 | 1 | 2 | 3 | 3 | 3 | 3 | 2 | |
| 3 | 0 | 6 | 4 | 1 | 1 | 5 | 2 | 1 | |
| p value (vs. MC) | <0.05 | <0.05 | <0.05 | <0.05 | |||||
| Interstitial chronic inflammation | 0 | 8 | 0 | 1 | 1 | 3 | 1 | 3 | 3 |
| 1 | 0 | 2 | 1 | 3 | 3 | 5 | 2 | 5 | |
| 2 | 0 | 2 | 4 | 3 | 1 | 0 | 3 | 0 | |
| 3 | 0 | 4 | 2 | 1 | 1 | 2 | 0 | 0 | |
| p value (vs. MC) | <0.05 | <0.05 | <0.05 | <0.05 | |||||
| Acute inflammation | 0 | 8 | 0 | 0 | 0 | 0 | 0 | 1 | 2 |
| 1 | 0 | 0 | 1 | 3 | 4 | 3 | 3 | 3 | |
| 2 | 0 | 2 | 5 | 1 | 1 | 0 | 1 | 2 | |
| 3 | 0 | 6 | 2 | 4 | 3 | 5 | 3 | 1 | |
| p value (vs. MC) | <0.05 | <0.05 | |||||||
| Indices | Tubular Dilation | Tubulointerstitial Degeneration/Atrophy | Interstitial Chronic Inflammation | Acute Inflammation | |||||
|---|---|---|---|---|---|---|---|---|---|
| R | p Value | R | p Value | R | p Value | R | p Value | ||
| Coefficient | KIM-1 | 0.517 | 0.000 | 0.341 | 0.007 | 0.565 | 0.000 | ||
| TIMP-1 | 0.708 | 0.000 | 0.631 | 0.000 | 0.493 | 0.000 | 0.759 | 0.000 | |
| Cystatin C | 0.807 | 0.000 | 0.811 | 0.000 | 0.687 | 0.000 | 0.780 | 0.000 | |
| Lipocalin 2 | 0.654 | 0.000 | 0.606 | 0.000 | 0.497 | 0.000 | 0.674 | 0.000 | |
| Clusterin | 0.290 | 0.041 | 0.242 | 0.090 | 0.293 | 0.039 | |||
| OPN | 0.659 | 0.000 | 0.634 | 0.000 | 0.467 | 0.000 | 0.735 | 0.000 | |
| IP-10 | 0.549 | 0.000 | 0.421 | 0.001 | 0.227 | 0.096 | 0.578 | 0.000 | |
| VEGF | 0.595 | 0.000 | 0.541 | 0.000 | 0.456 | 0.001 | 0.580 | 0.000 | |
| IL-6 | 0.461 | 0.001 | 0.371 | 0.011 | 0.334 | 0.023 | 0.532 | 0.000 | |
| IFN-γ | 0.477 | 0.000 | 0.398 | 0.003 | 0.498 | 0.000 | 0.391 | 0.004 | |
| Pathology | Tubular dilation | 0.822 | 0.000 | 0.768 | 0.000 | 0.843 | 0.000 | ||
| Tubulointerstitium degeneration/atrophy | 0.822 | 0.000 | 0.794 | 0.000 | 0.778 | 0.000 | |||
| Interstitial chronic inflammation | 0.768 | 0.000 | 0.794 | 0.000 | 0.697 | 0.000 | |||
| Acute inflammation | 0.843 | 0.000 | 0.778 | 0.000 | 0.697 | 0.000 | |||
| Kidney gene expression | IL-6 | 0.553 | 0.000 | 0.420 | 0.004 | 0.414 | 0.004 | 0.416 | 0.004 |
| TNF-α | 0.386 | 0.002 | 0.468 | 0.000 | 0.306 | 0.014 | 0.365 | 0.003 | |
| TGF-β1 | 0.370 | 0.008 | 0.403 | 0.003 | 0.301 | 0.032 | 0.351 | 0.012 | |
| TLR-4 | 0.372 | 0.030 | 0.386 | 0.024 | 0.410 | 0.016 | |||
| CD133 | 0.456 | 0.002 | 0.363 | 0.015 | 0.359 | 0.017 | 0.512 | 0.000 | |
| Pax-2 | 0.394 | 0.002 | 0.312 | 0.018 | 0.342 | 0.009 | 0.472 | 0.000 | |
| Six2 | 0.246 | 0.073 | 0.256 | 0.062 | 0.231 | 0.093 | |||
| CD11 β | 0.392 | 0.002 | 0.381 | 0.003 | 0.315 | 0.014 | 0.400 | 0.002 | |
| CD44 | 0.512 | 0.000 | 0.403 | 0.005 | 0.486 | 0.000 | 0.532 | 0.000 | |
| Vimentin | 0.365 | 0.008 | 0.325 | 0.019 | 0.388 | 0.005 | 0.410 | 0.003 | |
| Nestin | 0.352 | 0.005 | 0.481 | 0.000 | 0.351 | 0.005 | 0.330 | 0.009 | |
| Gut microbiota | Desulfovibrio genus | −0.451 | 0.002 | −0.375 | 0.012 | −0.430 | 0.004 | −0.410 | 0.006 |
| Prevotella genus | −0.321 | 0.034 | −0.284 | 0.061 | |||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, L.-X.; Abdolmaleky, H.M.; Yin, S.; Wang, Y.; Zhou, J.-R. Dietary Fermented Soy Extract and Oligo-Lactic Acid Alleviate Chronic Kidney Disease in Mice via Inhibition of Inflammation and Modulation of Gut Microbiota. Nutrients 2020, 12, 2376. https://doi.org/10.3390/nu12082376
He L-X, Abdolmaleky HM, Yin S, Wang Y, Zhou J-R. Dietary Fermented Soy Extract and Oligo-Lactic Acid Alleviate Chronic Kidney Disease in Mice via Inhibition of Inflammation and Modulation of Gut Microbiota. Nutrients. 2020; 12(8):2376. https://doi.org/10.3390/nu12082376
Chicago/Turabian StyleHe, Li-Xia, Hamid M. Abdolmaleky, Sheng Yin, Yihong Wang, and Jin-Rong Zhou. 2020. "Dietary Fermented Soy Extract and Oligo-Lactic Acid Alleviate Chronic Kidney Disease in Mice via Inhibition of Inflammation and Modulation of Gut Microbiota" Nutrients 12, no. 8: 2376. https://doi.org/10.3390/nu12082376
APA StyleHe, L.-X., Abdolmaleky, H. M., Yin, S., Wang, Y., & Zhou, J.-R. (2020). Dietary Fermented Soy Extract and Oligo-Lactic Acid Alleviate Chronic Kidney Disease in Mice via Inhibition of Inflammation and Modulation of Gut Microbiota. Nutrients, 12(8), 2376. https://doi.org/10.3390/nu12082376