Lemon Extract Reduces Angiotensin Converting Enzyme (ACE) Expression and Activity and Increases Insulin Sensitivity and Lipolysis in Mouse Adipocytes




Abstract
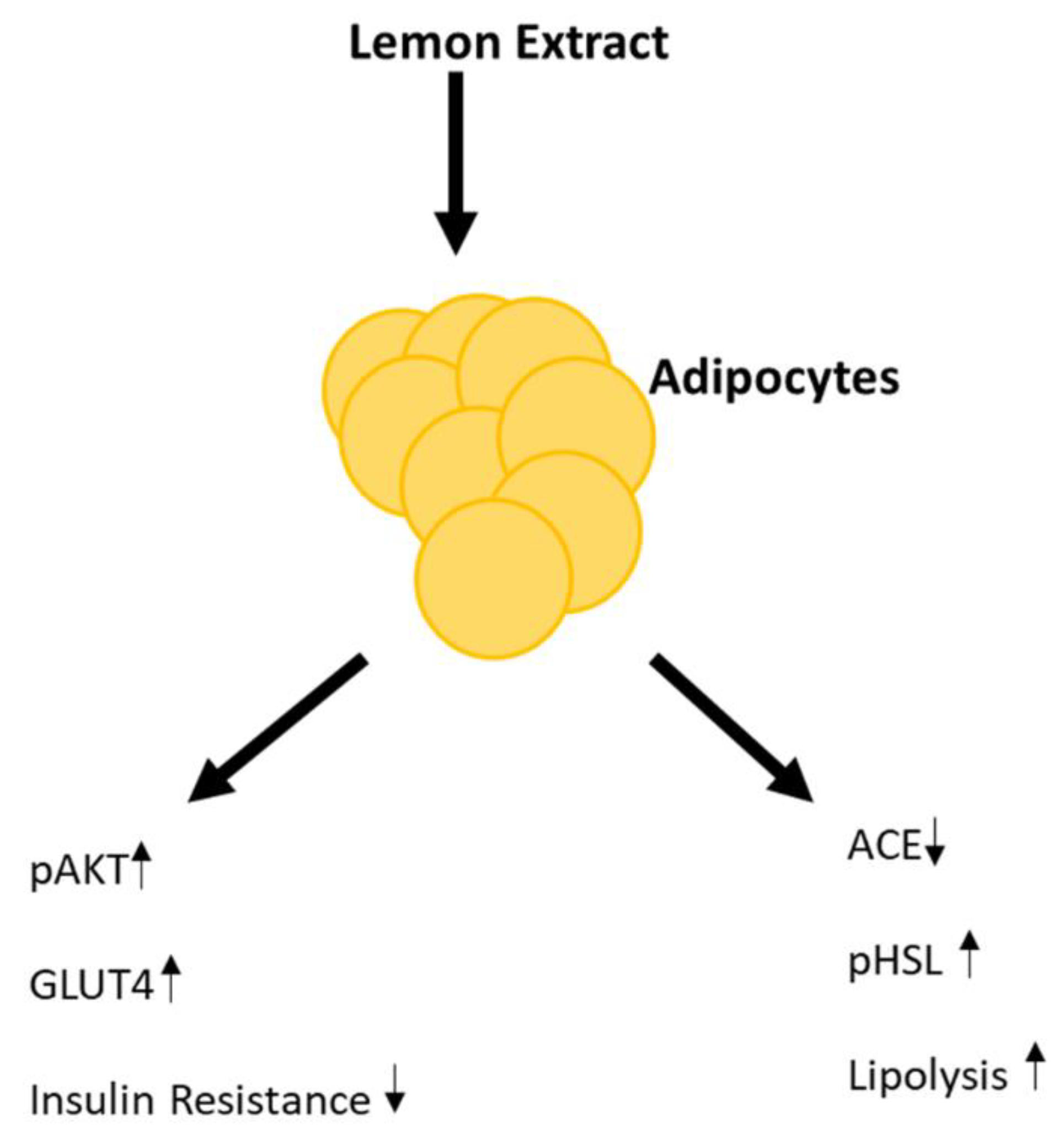
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Cell Culture
2.3. Preparation of Lemon Extract (LE)
2.4. RNA Isolation and Gene Expression
2.5. Protein Extraction
2.6. Western Blot Analysis
2.7. ACE Inhibitory Activity Assay
2.8. Free Glycerol Release Assay
2.9. Insulin Sensitivity Assay
2.10. Cell Viability Assay
2.11. Oil Red O Staining
2.12. High-Performance Liquid Chromatography (HPLC)
2.13. Mass Spectroscopy
2.14. Statistics
3. Results
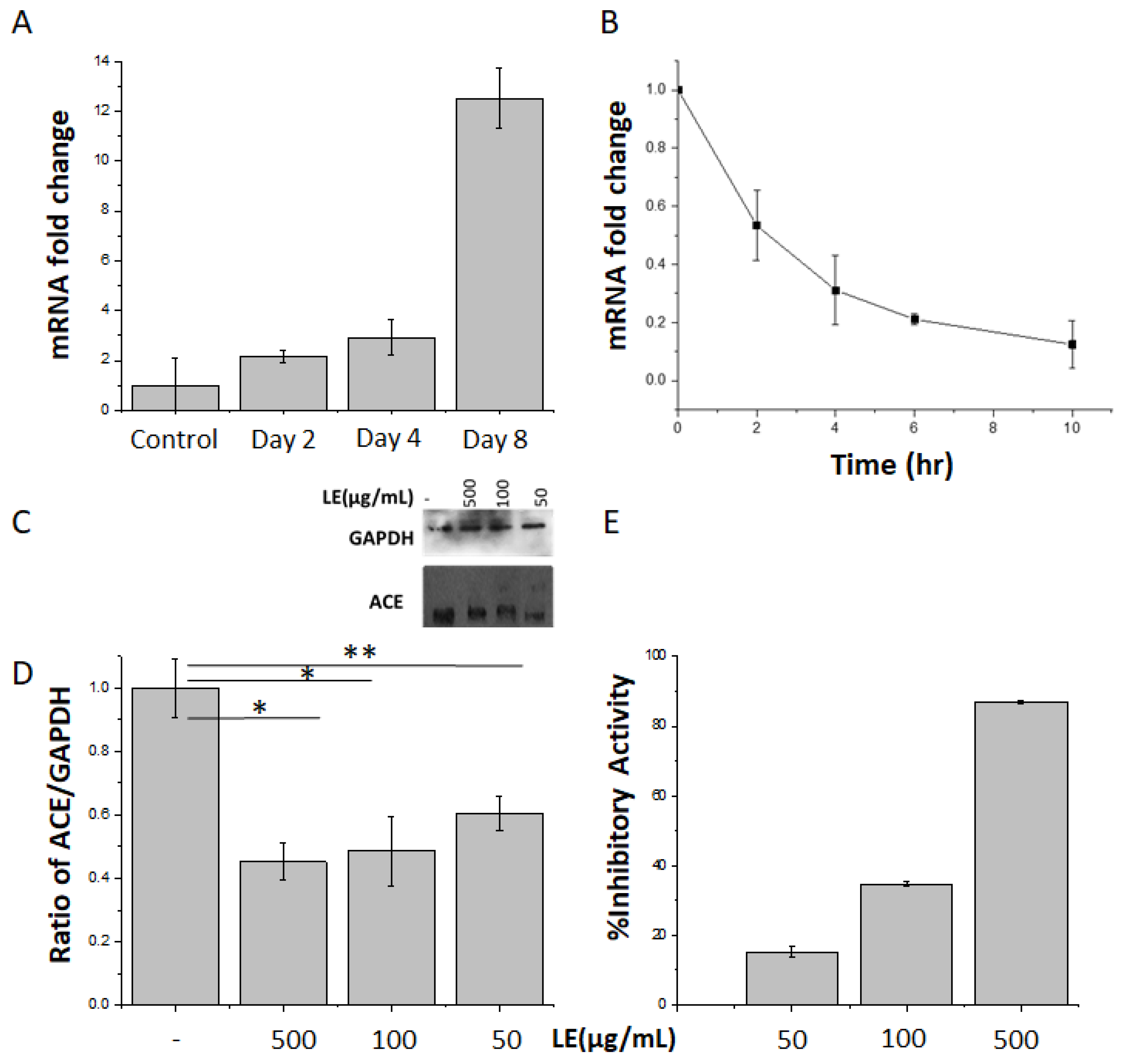
3.1. Lemon Extract Decreases ACE Gene and Protein Expression as Well as ACE Enzymatic Activity
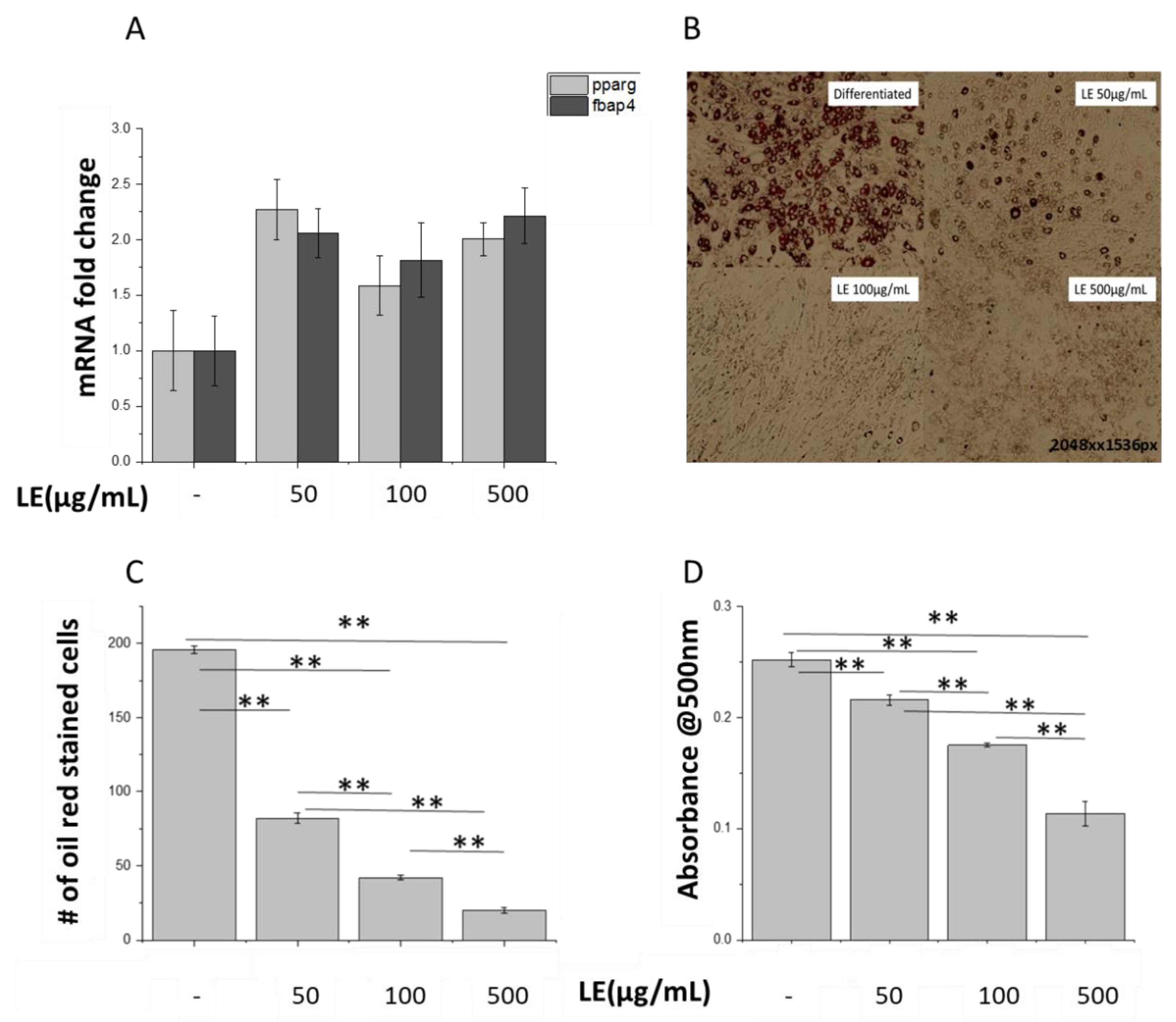
3.2. Lemon Extract Decreases Lipid Droplet Numbers in 3T3-L1 Adipocytes
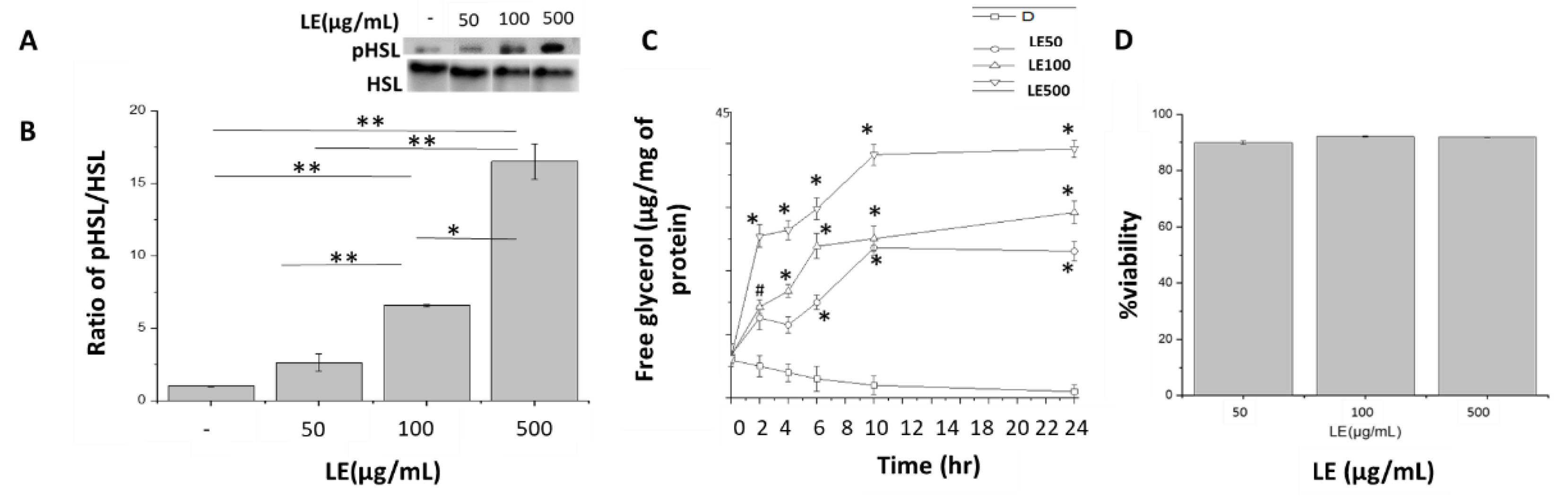
3.3. Stimulation of Lipolysis by Lemon Extract
3.4. Lemon Extract Increases Insulin Sensitivity
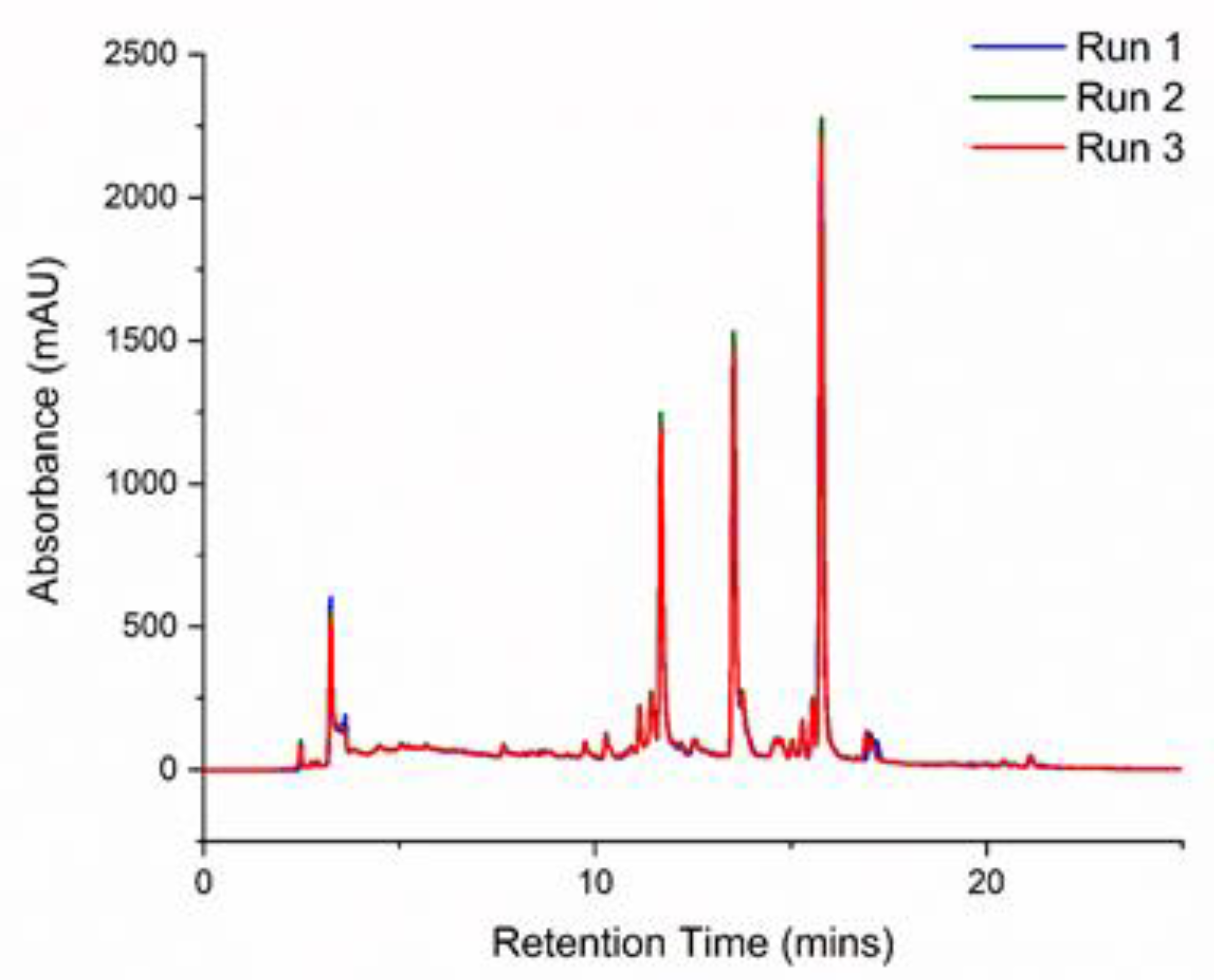
3.5. Identification of Lemon Constituents by HPLC and HPLC/MS
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Smith, K.B.; Smith, M.S. Obesity Statistics. Prim. Care 2016, 43, 121–135. [Google Scholar] [CrossRef] [PubMed]
- Schauer, P.R.; Kashyap, S.R.; Wolski, K.; Brethauer, S.A.; Kirwan, J.P.; Pothier, C.E.; Thomas, S.; Abood, B.; Nissen, S.E.; Bhatt, D.L. Bariatric surgery versus intensive medical therapy in obese patients with diabetes. N. Engl. J. Med. 2012, 366, 1567–1576. [Google Scholar] [CrossRef] [PubMed]
- National Health Services. Causes: Obesity: National Health Services (NHS). Available online: https://www.nhs.uk/conditions/obesity/causes/ (accessed on 6 June 2020).
- Mordes, J.P.; Liu, C.; Xu, S. Medications for weight loss. Curr. Opin. Endocrinol. Diabetes. Obes. 2015, 22, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.G.; Park, C.-Y. Anti-Obesity Drugs: A Review about Their Effects and Safety. Diabetes Metab. J. 2012, 36, 13–25. [Google Scholar] [CrossRef] [Green Version]
- Soeliman, F.A.; Azadbakht, L. Weight loss maintenance: A review on dietary related strategies. J. Res. Med. Sci. Off. J. Isfahan Univ. Med. Sci. 2014, 19, 268–275. [Google Scholar]
- Dansinger, M.L.; Gleason, J.A.; Griffith, J.L.; Selker, H.P.; Schaefer, E.J. Comparison of the Atkins, Ornish, Weight Watchers, and Zone diets for weight loss and heart disease risk reduction: A randomized trial. JAMA 2005, 293, 43–53. [Google Scholar] [CrossRef]
- Blackburn, G.L.; Phillips, J.C.; Morreale, S. Physician’s guide to popular low-carbohydrate weight-loss diets. Cleve. Clin. J. Med. 2001, 68, 761. [Google Scholar] [CrossRef] [Green Version]
- Wan, R.; Ahmet, I.; Brown, M.; Cheng, A.; Kamimura, N.; Talan, M.; Mattson, M.P. Cardioprotective effect of intermittent fasting is associated with an elevation of adiponectin levels in rats. J. Nutr. Biochem. 2010, 21, 413–417. [Google Scholar] [CrossRef] [Green Version]
- Anson, R.M.; Guo, Z.; De Cabo, R.; Iyun, T.; Rios, M.; Hagepanos, A.; Ingram, D.K.; Lane, M.A.; Mattson, M.P. Intermittent fasting dissociates beneficial effects of dietary restriction on glucose metabolism and neuronal resistance to injury from calorie intake. Proc. Natl. Acad. Sci. USA 2003, 100, 6216–6220. [Google Scholar] [CrossRef] [Green Version]
- Brehm, B.J.; Seeley, R.J.; Daniels, S.R.; D’Alessio, D.A. A randomized trial comparing a very low carbohydrate diet and a calorie-restricted low fat diet on body weight and cardiovascular risk factors in healthy women. J. Clin. Endocrinol. Metab. 2003, 88, 1617–1623. [Google Scholar] [CrossRef] [Green Version]
- Henry, R.R.; Wiest-Kent, T.A.; Scheaffer, L.; Kolterman, O.G.; Olefsky, J.M. Metabolic consequences of very-low-calorie diet therapy in obese non-insulin-dependent diabetic and nondiabetic subjects. Diabetes 1986, 35, 155–164. [Google Scholar] [CrossRef] [PubMed]
- Baker, S.; Jerums, G.; Proietto, J. Effects and clinical potential of very-low-calorie diets (VLCDs) in type 2 diabetes. Diabetes Res. Clin. Pract. 2009, 85, 235–242. [Google Scholar] [CrossRef]
- Delbridge, E.; Proietto, J. State of the science: VLED (Very Low Energy Diet) for obesity. Asia Pac. J. Clin. Nutr. 2006, 15, 49–54. [Google Scholar] [PubMed]
- Kim, M.J.; Hwang, J.H.; Ko, H.J.; Na, H.B.; Kim, J.H. Lemon detox diet reduced body fat, insulin resistance, and serum hs-CRP level without hematological changes in overweight Korean women. Nutr. Res. 2015, 35, 409–420. [Google Scholar] [CrossRef]
- Lu, Y.; Xi, W.; Ding, X.; Fan, S.; Zhang, Y.; Jiang, D.; Li, Y.; Huang, C.; Zhou, Z. Citrange fruit extracts alleviate obesity-associated metabolic disorder in high-fat diet-induced obese C57BL/6 mouse. Int. J. Mol. Sci. 2013, 14, 23736–23750. [Google Scholar] [CrossRef]
- Estaquio, C.; Castetbon, K.; Kesse-Guyot, E.; Bertrais, S.; Deschamps, V.; Dauchet, L.; Péneau, S.; Galan, P.; Hercberg, S. The French National Nutrition and Health Program score is associated with nutritional status and risk of major chronic diseases. J. Nutr. 2008, 138, 946–953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.; Serdula, M.; Janket, S.-J.; Cook, N.R.; Sesso, H.D.; Willett, W.C.; Manson, J.E.; Buring, J.E. A prospective study of fruit and vegetable intake and the risk of type 2 diabetes in women. Diabetes Care 2004, 27, 2993–2996. [Google Scholar] [CrossRef] [Green Version]
- Coelho, R.C.L.A.; Hermsdorff, H.H.M.; Bressan, J. Anti-inflammatory properties of orange juice: Possible favorable molecular and metabolic effects. Plant Foods Hum. Nutr. 2013, 68, 1–10. [Google Scholar] [CrossRef]
- Mollace, V.; Sacco, I.; Janda, E.; Malara, C.; Ventrice, D.; Colica, C.; Visalli, V.; Muscoli, S.; Ragusa, S.; Muscoli, C.; et al. Hypolipemic and hypoglycaemic activity of bergamot polyphenols: From animal models to human studies. Fitoterapia 2011, 82, 309–316. [Google Scholar] [CrossRef]
- Dow, C.A.; Going, S.B.; Chow, H.-H.S.; Patil, B.S.; Thomson, C.A. The effects of daily consumption of grapefruit on body weight, lipids, and blood pressure in healthy, overweight adults. Metabolism. 2012, 61, 1026–1035. [Google Scholar] [CrossRef]
- Aptekmann, N.P.; Cesar, T.B. Long-term orange juice consumption is associated with low LDL-cholesterol and apolipoprotein B in normal and moderately hypercholesterolemic subjects. Lipids Health Dis. 2013, 12, 119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oboh, G.; Bello, F.O.; Ademosun, A.O.; Akinyemi, A.J.; Adewuni, T.M. Antioxidant, hypolipidemic, and anti-angiotensin-1-converting enzyme properties of lemon (Citrus limon) and lime (Citrus aurantifolia) juices. Comp. Clin. Path. 2015, 24, 1395–1406. [Google Scholar] [CrossRef]
- Tejpal, S.; Sanghera, N.; Manoharan, V.; Planas-Iglesias, J.; Myler, K.; Klein-Seetharaman, J. Towards personalised molecular feedback for weight loss. BMC Obes. 2019, 6, 20. [Google Scholar] [CrossRef] [PubMed]
- Tejpal, S.; Bastie, C.; Seetharaman, J.K. Lemon Juice: A potential source of Angiotensin Converting Enzyme antagonism for weight loss and insulin resistance. Proc. Nutr. Soc. 2018, 77, E213. [Google Scholar] [CrossRef]
- Reaven, G.M. Insulin resistance: The link between obesity and cardiovascular disease. Med. Clin. N. Am. 2011, 95, 875–892. [Google Scholar] [CrossRef]
- Hill, J.O.; Peters, J.C. Biomarkers and functional foods for obesity and diabetes. Br. J. Nutr. 2002, 88, S213–S218. [Google Scholar] [CrossRef]
- Wilcox, G. Insulin and insulin resistance. Clin. Biochem. Rev. 2005, 26, 19–39. [Google Scholar]
- Atkinson, B.J.; Griesel, B.A.; King, C.D.; Josey, M.A.; Olson, A.L. Moderate GLUT4 overexpression improves insulin sensitivity and fasting triglyceridemia in high-fat diet-fed transgenic mice. Diabetes 2013, 62, 2249–2258. [Google Scholar] [CrossRef] [Green Version]
- Garvey, W.T.; Maianu, L.; Huecksteadt, T.P.; Birnbaum, M.J.; Molina, J.M.; Ciaraldi, T.P. Pretranslational suppression of a glucose transporter protein causes insulin resistance in adipocytes from patients with non-insulin-dependent diabetes mellitus and obesity. J. Clin. Investig. 1991, 87, 1072–1081. [Google Scholar] [CrossRef] [Green Version]
- Ikemoto, S.; Thompson, K.S.; Takahashi, M.; Itakura, H.; Lane, M.D.; Ezaki, O. High fat diet-induced hyperglycemia: Prevention by low level expression of a glucose transporter (GLUT4) minigene in transgenic mice. Proc. Natl. Acad. Sci. USA 1995, 92, 3096–3099. [Google Scholar] [CrossRef] [Green Version]
- Abel, E.D.; Peroni, O.; Kim, J.K.; Kim, Y.-B.; Boss, O.; Hadro, E.; Minnemann, T.; Shulman, G.I.; Kahn, B.B. Adipose-selective targeting of the GLUT4 gene impairs insulin action in muscle and liver. Nature 2001, 409, 729–733. [Google Scholar] [CrossRef] [PubMed]
- Garin-Shkolnik, T.; Rudich, A.; Hotamisligil, G.S.; Rubinstein, M. FABP4 attenuates PPARγ and adipogenesis and is inversely correlated with PPARγ in adipose tissues. Diabetes 2014, 63, 900–911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Queipo-Ortuño, M.I.; Escoté, X.; Ceperuelo-Mallafré, V.; Garrido-Sanchez, L.; Miranda, M.; Clemente-Postigo, M.; Pérez-Pérez1, R.; Peral, B.; Cardona, F.; Fernández-Real, J.M.; et al. FABP4 dynamics in obesity: Discrepancies in adipose tissue and liver expression regarding circulating plasma levels. PLoS ONE 2012, 7, e48605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duncan, R.E.; Ahmadian, M.; Jaworski, K.; Sarkadi-Nagy, E.; Sul, H.S. Regulation of lipolysis in adipocytes. Annu. Rev. Nutr. 2007, 27, 79–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carmen, G.-Y.; Víctor, S.-M. Signalling mechanisms regulating lipolysis. Cell. Signal. 2006, 18, 401–408. [Google Scholar] [CrossRef]
- Miyoshi, H.; Perfield, J.W., II; Obin, M.S.; Greenberg, A.S. Adipose triglyceride lipase regulates basal lipolysis and lipid droplet size in adipocytes. J. Cell. Biochem. 2008, 105, 1430–1436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stasiakowa, L. Sodium excretion in obese adolescents. Pol. Med. J. 1968, 7, 1344–1356. [Google Scholar]
- Wang, P.; Holst, C.; Andersen, M.R.; Astrup, A.; Bouwman, F.G.; van Otterdijk, S.; Wodzig, W.K.W.H.; Van Baak, M.A.; Larsen, T.M.; Jebb, S.A.; et al. Blood profile of proteins and steroid hormones predicts weight change after weight loss with interactions of dietary protein level and glycemic index. PLoS ONE 2011, 6, e1677. [Google Scholar]
- Eriksson, U.; Danilczyk, U.; Penninger, J.M. Just the beginning: Novel functions for angiotensin-converting enzymes. Curr. Biol. 2002, 12, R745–R752. [Google Scholar] [CrossRef] [Green Version]
- Tejpal, S.; Sanghera, N.; Manoharan, V.; Planas-Iglesias, J.; Klein-Seetharaman, J. A marker for personalised feedback on dieting: Angiotensin Converting Enzyme. Proc. Nutr. Soc. 2019, 78, E44. [Google Scholar] [CrossRef] [Green Version]
- Pan, Y.-H.; Wang, M.; Huang, Y.-M.; Wang, Y.-H.; Chen, Y.-L.; Geng, L.-J.; Zhang, X.-X.; Zhao, H.-L. ACE Gene I/D Polymorphism and Obesity in 1,574 Patients with Type 2 Diabetes Mellitus. Dis. Markers 2016, 2016, 7420540. [Google Scholar] [CrossRef] [PubMed]
- Patel, V.B.; Parajuli, N.; Oudit, G.Y. Role of angiotensin-converting enzyme 2 (ACE2) in diabetic cardiovascular complications. Clin. Sci. 2014, 126, 471–482. [Google Scholar] [CrossRef]
- Perticone, F.; Ceravolo, R.; Iacopino, S.; Cloro, C.; Ventura, G.; Maio, R.; Gulletta, E.; Perrotti, N.; Mattioli, P.L. Relationship between Angiotensin-Converting Enzyme Gene Polymorphism and Insulin Resistance in Never-Treated Hypertensive Patients. J. Clin. Endocrinol. Metab. 2001, 86, 172–178. [Google Scholar] [CrossRef] [PubMed]
- Messerli, F.H.; Bangalore, S.; Bavishi, C.; Rimoldi, S.F. Angiotensin-Converting Enzyme Inhibitors in Hypertension: To Use or Not to Use? J. Am. Coll. Cardiol. 2018, 71, 1474–1482. [Google Scholar] [CrossRef] [PubMed]
- Jones, B.H.; Standridge, M.K.; Moustaid, N. Angiotensin II increases lipogenesis in 3T3-L1 and human adipose cells. Endocrinology 1997, 138, 1512–1519. [Google Scholar] [CrossRef] [PubMed]
- Darimont, C.; Vassaux, G.; Ailhaud, G.; Negrel, R. Differentiation of preadipose cells: Paracrine role of prostacyclin upon stimulation of adipose cells by angiotensin-II. Endocrinology 1994, 135, 2030–2036. [Google Scholar] [CrossRef]
- Zebisch, K.; Voigt, V.; Wabitsch, M.; Brandsch, M. Protocol for effective differentiation of 3T3-L1 cells to adipocytes. Anal. Biochem. 2012, 425, 88–90. [Google Scholar] [CrossRef]
- Sentandreu, M.A.; Toldrá, F. A fluorescence-based protocol for quantifying angiotensin-converting enzyme activity. Nat. Protoc. 2006, 1, 2423–2427. [Google Scholar] [CrossRef]
- Guerrero, L.; Castillo, J.; Quiñones, M.; Garcia-Vallve, S.; Arola, L.; Pujadas, G.; Muguerza, B. Inhibition of Angiotensin-Converting Enzyme Activity by Flavonoids: Structure-Activity Relationship Studies. PLoS ONE 2012, 7, e49493. [Google Scholar] [CrossRef] [Green Version]
- Storka, A.; Vojtassakova, E.; Mueller, M.; Kapiotis, S.; Haider, D.G.; Jungbauer, A.; Wolzt, M. Angiotensin inhibition stimulates PPARγ and the release of visfatin. Eur. J. Clin. Invest. 2008, 38, 820–826. [Google Scholar] [CrossRef]
- Fujimura, A.; Ushijima, K.; Ando, H. Does the PPAR-γ-activating property of telmisartan provide a benefit in clinical practice? Hypertens. Res. 2013, 36, 183. [Google Scholar] [CrossRef] [PubMed]
- Mackenzie, R.W.; Elliott, B.T. Akt/PKB activation and insulin signaling: A novel insulin signaling pathway in the treatment of type 2 diabetes. Diabetes. Metab. Syndr. Obes. 2014, 7, 55–64. [Google Scholar] [CrossRef] [Green Version]
- Proestos, C.; Chorianopoulos, N.; Nychas, G.-J.E.; Komaitis, M. RP-HPLC analysis of the phenolic compounds of plant extracts. investigation of their antioxidant capacity and antimicrobial activity. J. Agric. Food Chem. 2005, 53, 1190–1195. [Google Scholar] [CrossRef] [PubMed]
- Cimpan, G.; Gocan, S. Analysis of Medicinal Plants by Hplc: Recent Approaches. J. Liq. Chromatogr. Relat. Technol. 2002, 25, 2225–2292. [Google Scholar] [CrossRef]
- Zhou, S.; Song, L.; Masschelein, J.; Sumang, F.A.M.; Papa, I.A.; Zulaybar, T.O.; Custodio, A.B.; Zabala, D.; Alcantara, E.P.; Santos, E.L.C.D.L.; et al. Pentamycin Biosynthesis in Philippine Streptomyces sS816: Cytochrome P450-Catalyzed Installation of the C-14 Hydroxyl Grou. ACS Chem. Biol. 2019, 14, 1305–1309. [Google Scholar]
- Eisenmann, J.C.; Sarzynski, M.A.; Glenn, K.; Rothschild, M.; Heelan, K.A. ACE I/D genotype, adiposity, and blood pressure in children. Cardiovasc. Diabetol. 2009, 8, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cooper, R.; McFarlane-Anderson, N.; Bennett, F.I.; Wilks, R.; Puras, A.; Tewksbury, D.; Ward, R.; Forrester, T. ACE, angiotensinogen and obesity: A potential pathway leading to hypertension. J. Hum. Hypertens. 1997, 11, 107–111. [Google Scholar] [CrossRef] [Green Version]
- Yvan-Charvet, L.; Quignard-Boulangé, A. Role of adipose tissue renin-angiotensin system in metabolic and inflammatory diseases associated with obesity. Kidney Int. 2011, 79, 162–168. [Google Scholar] [CrossRef]
- Mulvihill, E.E.; Burke, A.C.; Huff, M.W. Citrus Flavonoids as Regulators of Lipoprotein Metabolism and Atherosclerosis. Annu. Rev. Nutr. 2016, 36, 275–299. [Google Scholar] [CrossRef]
- Choi, J.S.; Yokozawa, T.; Oura, H. Improvement of hyperglycemia and hyperlipemia in streptozotocin-diabetic rats by a methanolic extract of Prunus davidiana stems and its main component, prunin. Planta Med. 1991, 57, 208–211. [Google Scholar] [CrossRef]
- Kern, P.A.; di Gregorio, G.; Lu, T.; Rassouli, N.; Ranganathan, G. Perilipin expression in human adipose tissue is elevated with obesity. J. Clin. Endocrinol. Metab. 2004, 89, 1352–1358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Der Merwe, M.-T.; Schlaphoff, G.P.; Crowther, N.J.; Boyd, I.H.; Gray, I.P.; Joffe, B.I.; Lonnroth, P.N. Lactate and Glycerol Release from Adipose Tissue in Lean, Obese, and Diabetic Women from South Africa. J. Clin. Endocrinol. Metab. 2001, 86, 3296–3303. [Google Scholar] [CrossRef] [Green Version]
- Watt, M.J.; Holmes, A.G.; Pinnamaneni, S.K.; Garnham, A.P.; Steinberg, G.R.; Kemp, B.E.; Febbraio, M.A. Regulation of HSL serine phosphorylation in skeletal muscle and adipose tissue. Am. J. Physiol. Metab. 2006, 290, E500–E508. [Google Scholar] [CrossRef] [PubMed]
- Gholamnezhad, Z.; Mégarbane, B.; Rezaee, R. Molecular Mechanisms Mediating Adaptation to Exercise BT. In Physical Exercise for Human Health; Xiao, J., Ed.; Springer: Singapore, 2020; pp. 45–61. [Google Scholar]
- Jayasooriya, A.P.; Mathai, M.L.; Walker, L.L.; Begg, D.P.; Denton, D.A.; Cameron-Smith, D.; Egan, G.; McKinley, M.J.; Rodger, P.D.; Sinclair, A.J.; et al. Mice lacking angiotensin-converting enzyme have increased energy expenditure, with reduced fat mass and improved glucose clearance. Proc. Natl. Acad. Sci. USA 2008, 105, 6531–6536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Premaratna, S.D.; Manickam, E.; Begg, D.P.; Rayment, D.J.; Hafandi, A.; Jois, M.; Cameron-Smith, D.; Weisinger, R.S. Angiotensin-converting enzyme inhibition reverses diet-induced obesity, insulin resistance and inflammation in C57BL/6J mice. Int. J. Obes. 2012, 36, 233–243. [Google Scholar] [CrossRef] [Green Version]
- Velkoska, E.; Warner, F.J.; Cole, T.J.; Smith, I.; Morris, M.J. Metabolic effects of low dose angiotensin converting enzyme inhibitor in dietary obesity in the rat. Nutr. Metab. Cardiovasc. Dis. 2010, 20, 49–55. [Google Scholar] [CrossRef]
- Santos, E.L.; Souza, K.D.P.; Da Silva, E.D.; Batista, E.C.; Martins, P.J.F.; D’Almeida, V.; Pesquero, J.B. Long term treatment with ACE inhibitor enalapril decreases body weight gain and increases life span in rats. Biochem. Pharmacol. 2009, 78, 951–958. [Google Scholar] [CrossRef]
- Jordan, J.; Stinkens, R.; Jax, T.; Engeli, S.; Blaak, E.; May, M.; Havekes, B.; Schindler, C.; Albrecht, D.; Pal, P.; et al. Improved Insulin Sensitivity With Angiotensin Receptor Neprilysin Inhibition in Individuals With Obesity and Hypertension. Clin. Pharmacol. Ther. 2017, 101, 254–263. [Google Scholar] [CrossRef]
- Moro, C.; Galitzky, J.; Sengenes, C.; Crampes, F.; Lafontan, M.; Berlan, M. Functional and Pharmacological Characterization of the Natriuretic Peptide-Dependent Lipolytic Pathway in Human Fat Cells. J. Pharmacol. Exp. Ther. 2004, 308, 984–992. [Google Scholar] [CrossRef] [Green Version]
- Sengenès, C.; Berlan, M.; De Glisezinski, I.; Lafontan, M.; Galitzky, J. Natriuretic peptides: A new lipolytic pathway in human adipocytes. FASEB J. 2000, 14, 1345–1351. [Google Scholar] [CrossRef]
- Sengenès, C.; Bouloumié, A.; Hauner, H.; Berlan, M.; Busse, R.; Lafontan, M.; Galitzky, J. Involvement of a cGMP-dependent Pathway in the Natriuretic Peptide-mediated Hormone-sensitive Lipase Phosphorylation in Human Adipocytes. J. Biol. Chem. 2004, 278, 48617–48626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willenbrock, R.; Scheuermann, M.; Thibault, G.; Haass, M.; Höhnel, K.; Bohlender, J.; Luft, F.C.; Dietz, R. Angiotensin inhibition and atrial natriuretic peptide release after acute volume expansion in rats with aortocaval shunt. Cardiovasc. Res. 1999, 42, 733–742. [Google Scholar] [CrossRef] [Green Version]
- Liu, D.; Ceddia, R.P.; Collins, S. Cardiac natriuretic peptides promote adipose ‘browning’ through mTOR complex-1. Mol. Metab. 2018, 9, 192–198. [Google Scholar] [CrossRef] [PubMed]
- Kaisanlahti, A.; Glumoff, T. Browning of white fat: Agents and implications for beige adipose tissue to type 2 diabetes. J. Physiol. Biochem. 2019, 75, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carpentier, A.C.; Blondin, D.P.; Virtanen, K.A.; Richard, D.; Haman, F.; Turcotte, É.E. Brown Adipose Tissue Energy Metabolism in Humans. Front. Endocrinol. 2018, 9, 447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braun, K.; Oeckl, J.; Westermeier, J.; Li, Y.; Klingenspor, M. Non-adrenergic control of lipolysis and thermogenesis in adipose tissues. J. Exp. Biol. 2018, 221, jeb.165381. [Google Scholar] [CrossRef] [Green Version]
- Tejpal, S.; Sanghera, N.; Manoharan, V.; Planas-Iglesias, J.; Bastie, C.C.; Klein-Seetharaman, J. Angiotensin Converting Enzyme (ACE): A Marker for Personalized Feedback on Dieting. Nutrients 2020, 12, 660. [Google Scholar] [CrossRef] [Green Version]
- Han, L.-K.; Sumiyoshi, M.; Zheng, Y.-N.; Okuda, H.; Kimura, Y. Anti-obesity action of Salix matsudana leaves (Part 2). Isolation of anti-obesity effectors from polyphenol fractions of Salix matsudana. Phytother. Res. 2003, 17, 1195–1198. [Google Scholar] [CrossRef] [PubMed]
- Zang, Y.; Zhang, L.; Igarashi, K.; Yu, C. The anti-obesity and anti-diabetic effects of kaempferol glycosides from unripe soybean leaves in high-fat-diet mice. Food Funct. 2015, 6, 834–841. [Google Scholar] [CrossRef] [PubMed]
- Jorge, A.P.; Horst, H.; De Sousa, E.; Pizzolatti, M.G.; Silva, F.R.M.B. Insulinomimetic effects of kaempferitrin on glycaemia and on 14C-glucose uptake in rat soleus muscle. Chem. Biol. Interact. 2004, 149, 89–96. [Google Scholar] [CrossRef]
- Borradaile, N.M.; De Dreu, L.E.; Huff, M.W. Inhibition of net HepG2 cell apolipoprotein B secretion by the citrus flavonoid naringenin involves activation of phosphatidylinositol 3-kinase, independent of insulin receptor substrate-1 phosphorylation. Diabetes 2003, 52, 2554–2561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.H.; Park, Y.; Bae, K.; Bok, S.H.; Kwon, Y.; Lee, E.; Choi, M.-S. Cholesterol-lowering activity of naringenin via inhibition of 3-hydroxy-3-methylglutaryl coenzyme A reductase and acyl coenzyme A: Cholesterol acyltransferase in rats. Ann. Nutr. Metab. 1999, 43, 173–180. [Google Scholar] [CrossRef] [PubMed]
- Mulvihill, E.E.; Assini, J.M.; Lee, J.K.; Allister, E.M.; Sutherland, B.G.; Koppes, J.B.; Sawyez, C.G.; Edwards, J.Y.; Telford, D.E.; Charbonneau, A.; et al. Nobiletin attenuates VLDL overproduction, dyslipidemia, and atherosclerosis in mice with diet-induced insulin resistance. Diabetes 2011, 60, 1446–1457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burke, A.C.; Sutherland, B.G.; Telford, D.E.; Morrow, M.R.; Sawyez, C.G.; Edwards, J.Y.; Drangova, M.; Huff, M.W. Intervention with citrus flavonoids reverses obesity and improves metabolic syndrome and atherosclerosis in obese Ldlr(-/-) mice. J. Lipid Res. 2018, 59, 1714–1728. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.K.; Jeong, T.-S.; Lee, M.-K.; Park, Y.B.; Choi, M.-S. Lipid-lowering efficacy of hesperetin metabolites in high-cholesterol fed rats. Clin. Chim. Acta. 2003, 327, 129–137. [Google Scholar] [CrossRef]
- Fukuchi, Y.; Hiramitsu, M.; Okada, M.; Hayashi, S.; Nabeno, Y.; Osawa, T.; Naito, M. Lemon Polyphenols Suppress Diet-induced Obesity by Up-Regulation of mRNA Levels of the Enzymes Involved in beta-Oxidation in Mouse White Adipose Tissue. J. Clin. Biochem. Nutr. 2008, 43, 201–209. [Google Scholar] [CrossRef] [Green Version]
- Assini, J.M.; Mulvihill, E.E.; Huff, M.W. Citrus flavonoids and lipid metabolism. Curr. Opin. Lipidol. 2013, 24, 34–40. [Google Scholar] [CrossRef]
- Rizza, S.; Muniyappa, R.; Iantorno, M.; Kim, J.-A.; Chen, H.; Pullikotil, P.; Senese, N.; Tesauro, M.; Lauro, D.; Cardillo, C.; et al. Citrus polyphenol hesperidin stimulates production of nitric oxide in endothelial cells while improving endothelial function and reducing inflammatory markers in patients with metabolic syndrome. J. Clin. Endocrinol. Metab. 2011, 96, E782–E792. [Google Scholar] [CrossRef]
- Ghanim, H.; Sia, C.L.; Upadhyay, M.; Upadhyay, M.; Korzeniewski, K.; Viswanathan, P.; Abuaysheh, S.; Mohanty, P.; Dandona, P. Orange juice neutralizes the proinflammatory effect of a high-fat, high-carbohydrate meal and prevents endotoxin increase and Toll-like receptor expression. Am. J. Clin. Nutr. 2010, 91, 940–949. [Google Scholar] [CrossRef] [Green Version]
- Nakajima, V.M.; Macedo, G.A.; Macedo, J.A. Citrus bioactive phenolics: Role in the obesity treatment. LWT Food Sci. Technol. 2014, 59, 1205–1212. [Google Scholar] [CrossRef] [Green Version]
- Ono, E.; Inoue, J.; Hashidume, T.; Shimizu, M.; Sato, R. Anti-obesity and anti-hyperglycemic effects of the dietary citrus limonoid nomilin in mice fed a high-fat diet. Biochem. Biophys. Res. Commun. 2011, 410, 677–681. [Google Scholar] [CrossRef] [PubMed]
- Maruyama, T.; Miyamoto, Y.; Nakamura, T.; Tamai, Y.; Okada, H.; Sugiyama, E.; Nakamura, T.; Itadani, H.; Tanaka, K. Identification of membrane-type receptor for bile acids (M-BAR). Biochem. Biophys. Res. Commun. 2002, 298, 714–719. [Google Scholar] [CrossRef]
- Kwon, E.-Y.; Choi, M.-S. Dietary Eriodictyol Alleviates Adiposity, Hepatic Steatosis, Insulin Resistance, and Inflammation in Diet-Induced Obese Mice. Int. J. Mol. Sci. 2019, 20, 1227. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time (min) | A: Water | B: Acetonitrile | Flow Rate (mL/min) |
|---|---|---|---|
| 0 | 95 | 5 | 1 |
| 10 | 75 | 25 | 1 |
| 20 | 60 | 40 | 1 |
| 30 | 50 | 50 | 1 |
| 35 | 5 | 95 | 1 |
| 40 | 5 | 95 | 1 |
| 50 | 95 | 5 | 1 |
| 60 | 95 | 5 | 1 |
| RT (min) | [M-H]- | Mol. Formula | MS Fragments (m/z) | Name | Class |
|---|---|---|---|---|---|
| 1.3 | 191.02 | C6H7O7 | 133, 111 | (iso)citric acid | Organic acid |
| 5.9 | 771.201 | C33H39O21 | 695, 547, 415, 375, 353, 285, 191 | Kaempferol-Osophoroside-Oglucoside | Flavonol acylated glycoside |
| 13.1 | 711.28 | C34H47O16 | 693, 549, 341 | Nomilinic acid-17-O-glucoside | Limonoid |
| 14.7 | 693.2775 | C34H45O15 | 531, 443, 341 | Nomilinic acid-4-Oglucoside | Limonoid |
| 2.6 | 651. | C29H31O17 | 507, 417, 341 | Kaempferol acetyl dihexoside | Flavonoid-3-o-glycosides |
| 13.1 | 649.2513 | C32H41O14 | 413, 341 | Limonin glucoside | Limonoid |
| 8.9 | 625.17 | C28H33O16 | 383, 312 | Diosmetin-6, 8-di-C-hexoside (Lucenin-2,4′-methyl ether) | flavonoid-7-o-glycosides |
| 2.3 | 623.1622 | C28H31O16 | 605, 533, 503, 579, 443 | 6,8-C, CDiglucosyldiosmetin isomer | C-flavone glycoside |
| 2.7 | 623.1565 | C28H31O16 | 605, 533, 503, 413, 329 | C,CDiglucosyldiosmetin | C-Flavone glycoside |
| 14.9 | 609.1825 | C28H33O15 | 301.07 | Diosmetin-7-O-rutinoside (diosmin) | Flavonoid-7-o-glycosides |
| 15.7 | 609.1824 | C28H33O15 | Chrysoeriol 7-rutinoside | flavonoid-7-o-glycosides | |
| 13.6 | 607.1668 | C25H31O15 | 299,284 | Diosmetin 7-O-neohesperidoside Neodiosmin | Flavone |
| 14.7 | 607.1668 | C25H31O15 | 299,283.8 | Chrysoeriol-7-O-neohesperidoside | Flavonoid-7-o-glycosides |
| 13.3 | 595.1668 | C27H31O15 | 505, 457, 427, 421, 409, 391, 379, 355, 337, 325, 307, 295 | Apigenin-6,8-di-C-glucoside | Flavonoid |
| 13.0 | 287.0556 | C15H11O6 | 151,135,125,107 | (2S)-Eriodictyol | Flavone |
| 36.7 | 269.24 | C17H37O2 | 225,201,151 | Apigenin | Flavone |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tejpal, S.; Wemyss, A.M.; Bastie, C.C.; Klein-Seetharaman, J. Lemon Extract Reduces Angiotensin Converting Enzyme (ACE) Expression and Activity and Increases Insulin Sensitivity and Lipolysis in Mouse Adipocytes. Nutrients 2020, 12, 2348. https://doi.org/10.3390/nu12082348
Tejpal S, Wemyss AM, Bastie CC, Klein-Seetharaman J. Lemon Extract Reduces Angiotensin Converting Enzyme (ACE) Expression and Activity and Increases Insulin Sensitivity and Lipolysis in Mouse Adipocytes. Nutrients. 2020; 12(8):2348. https://doi.org/10.3390/nu12082348
Chicago/Turabian StyleTejpal, Shilpa, Alan M. Wemyss, Claire C. Bastie, and Judith Klein-Seetharaman. 2020. "Lemon Extract Reduces Angiotensin Converting Enzyme (ACE) Expression and Activity and Increases Insulin Sensitivity and Lipolysis in Mouse Adipocytes" Nutrients 12, no. 8: 2348. https://doi.org/10.3390/nu12082348
APA StyleTejpal, S., Wemyss, A. M., Bastie, C. C., & Klein-Seetharaman, J. (2020). Lemon Extract Reduces Angiotensin Converting Enzyme (ACE) Expression and Activity and Increases Insulin Sensitivity and Lipolysis in Mouse Adipocytes. Nutrients, 12(8), 2348. https://doi.org/10.3390/nu12082348