The Effect of an Atherogenic Diet and Acute Hyperglycaemia on Endothelial Function in Rabbits Is Artery Specific

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethical Approval
2.2. Animal Model
2.3. Isometric Tension Myography
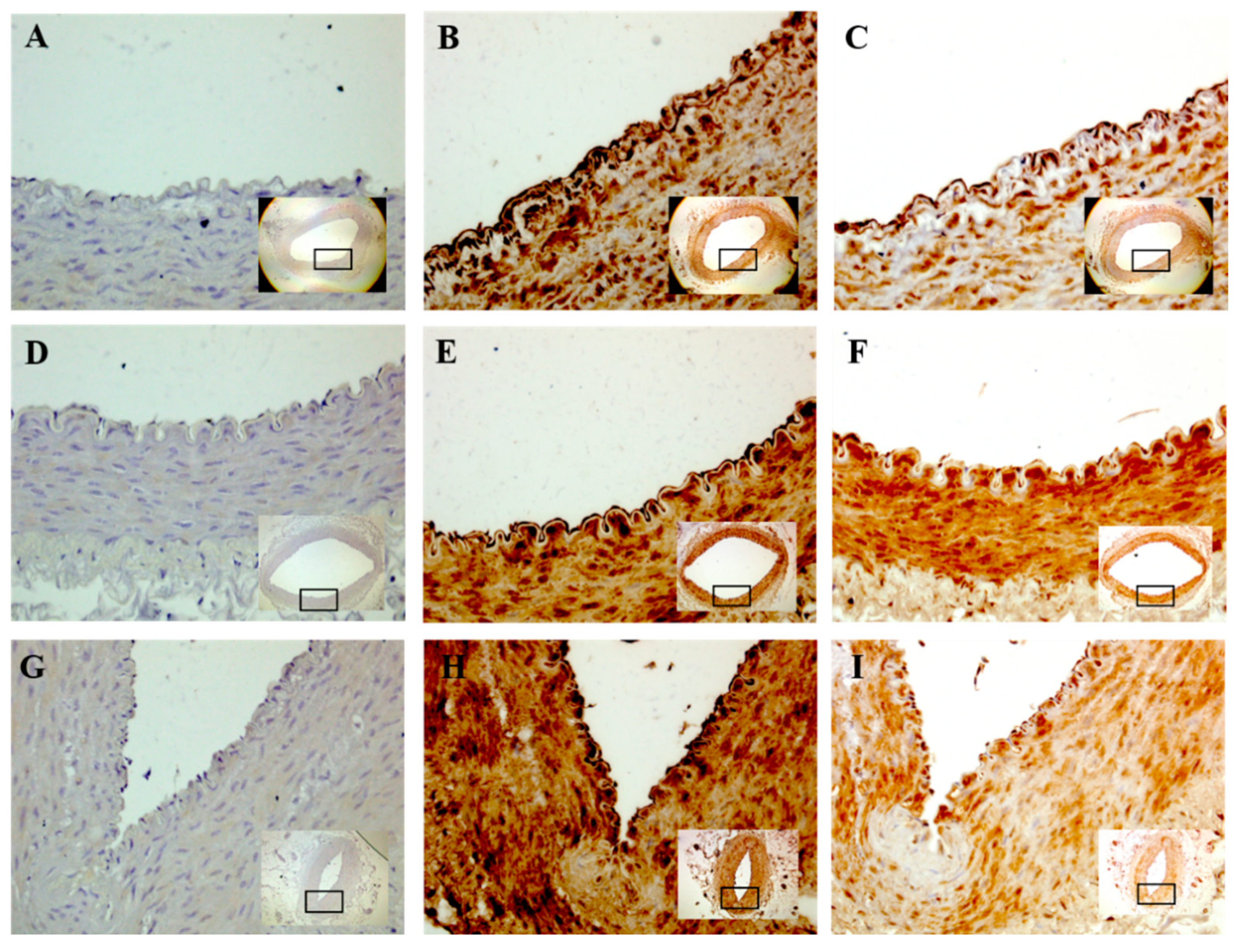
2.4. Immunohistochemistry (IHC)
2.5. IHC Semiquantification
2.6. Statistical Analysis
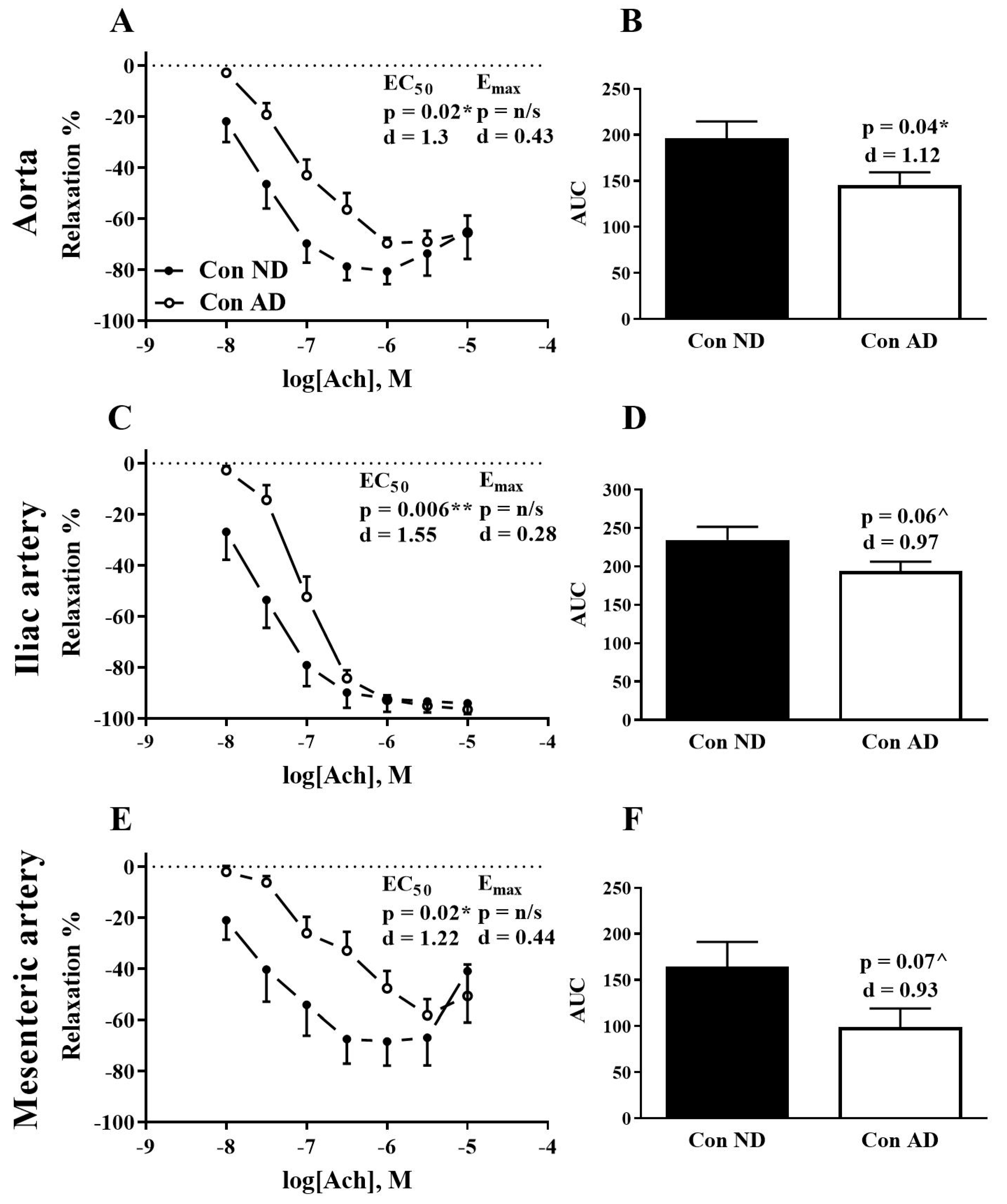
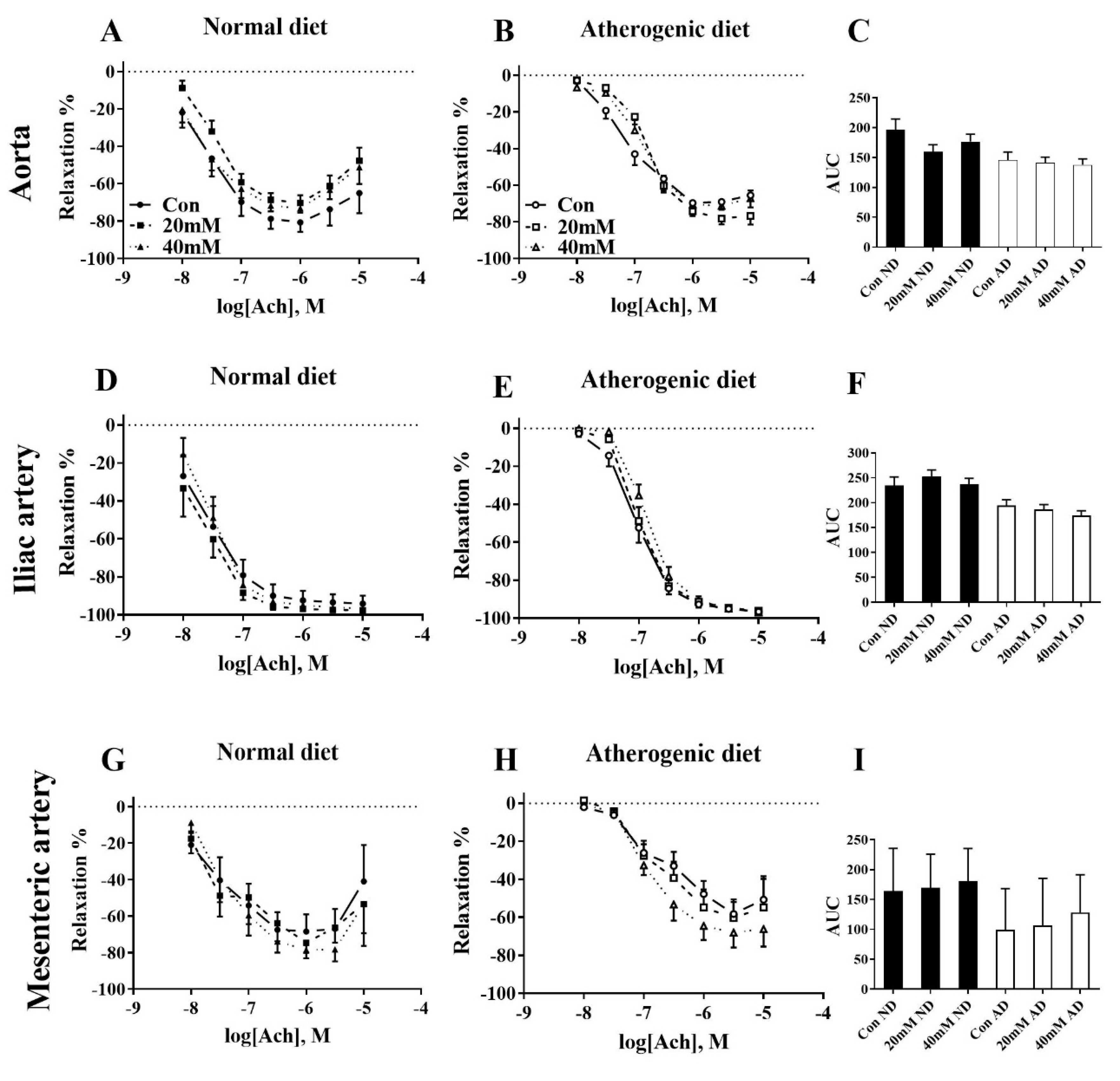
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Beckman, J.A.; Creager, M.A.; Libby, P. Diabetes and atherosclerosis: Epidemiology, pathophysiology, and management. JAMA 2002, 287, 2570–2581. [Google Scholar] [CrossRef] [PubMed]
- Creager, M.A.; Luscher, T.F.; Cosentino, F.; Beckman, J.A. Diabetes and vascular disease: Pathophysiology, clinical consequences, and medical therapy: Part I. Circulation 2003, 108, 1527–1532. [Google Scholar] [CrossRef] [PubMed]
- Bornfeldt, K.E.; Tabas, I. Insulin resistance, hyperglycemia, and atherosclerosis. Cell Metab. 2011, 14, 575–585. [Google Scholar] [CrossRef]
- Saad, M.I.; Abdelkhalek, T.M.; Saleh, M.M.; Kamel, M.A.; Youssef, M.; Tawfik, S.H.; Dominguez, H. Insights into the molecular mechanisms of diabetes-induced endothelial dysfunction: Focus on oxidative stress and endothelial progenitor cells. Endocrine 2015, 50, 537–567. [Google Scholar] [CrossRef] [PubMed]
- Gimbrone, M.A., Jr.; Garcia-Cardena, G. Endothelial Cell Dysfunction and the Pathobiology of Atherosclerosis. Circ. Res. 2016, 118, 620–636. [Google Scholar] [CrossRef] [PubMed]
- Schachinger, V.; Zeiher, A.M. Atherosclerosis-associated endothelial dysfunction. Z. Kardiol. 1899, 89 (Suppl. 9), 1899–1906, IX/70–74. [Google Scholar] [CrossRef] [PubMed]
- Fiorentino, T.V.; Prioletta, A.; Zuo, P.; Folli, F. Hyperglycemia-induced oxidative stress and its role in diabetes mellitus related cardiovascular diseases. Curr. Pharm. Des. 2013, 19, 5695–5703. [Google Scholar] [CrossRef] [PubMed]
- Son, S.M. Reactive oxygen and nitrogen species in pathogenesis of vascular complications of diabetes. Diabetes Metab. J. 2012, 36, 190–198. [Google Scholar] [CrossRef] [PubMed]
- Brownlee, M. Biochemistry and molecular cell biology of diabetic complications. Nature 2001, 414, 813–820. [Google Scholar] [CrossRef] [PubMed]
- Ceriello, A.; Genovese, S. Atherogenicity of postprandial hyperglycemia and lipotoxicity. Rev. Endocr. Metab. Disord. 2016, 17, 111–116. [Google Scholar] [CrossRef] [PubMed]
- Yamagishi, S.I.; Nakamura, K.; Matsui, T.; Ueda, S.I.; Imaizumi, T. Role of postprandial hyperglycaemia in cardiovascular disease in diabetes. Int. J. Clin. Pract. 2007, 61, 83–87. [Google Scholar] [CrossRef] [PubMed]
- Bonora, E.; Calcaterra, F.; Lombardi, S.; Bonfante, N.; Formentini, G.; Bonadonna, R.C.; Muggeo, M. Plasma glucose levels throughout the day and HbA(1c) interrelationships in type 2 diabetes: Implications for treatment and monitoring of metabolic control. Diabetes Care 2001, 24, 2023–2029. [Google Scholar] [CrossRef] [PubMed]
- Tesfamariam, B.; Brown, M.L.; Cohen, R.A. Elevated glucose impairs endothelium-dependent relaxation by activating protein kinase C. J. Clin. Investig. 1991, 87, 1643–1648. [Google Scholar] [CrossRef] [PubMed]
- Tesfamariam, B.; Brown, M.L.; Deykin, D.; Cohen, R.A. Elevated glucose promotes generation of endothelium-derived vasoconstrictor prostanoids in rabbit aorta. J. Clin. Investig. 1990, 85, 929–932. [Google Scholar] [CrossRef] [PubMed]
- Tesfamariam, B.; Cohen, R.A. Free radicals mediate endothelial cell dysfunction caused by elevated glucose. Am. J. Physiol. 1992, 263, H321–H326. [Google Scholar] [CrossRef] [PubMed]
- Qian, L.B.; Wang, H.P.; Chen, Y.; Chen, F.X.; Ma, Y.Y.; Bruce, I.C.; Xia, Q. Luteolin reduces high glucose-mediated impairment of endothelium-dependent relaxation in rat aorta by reducing oxidative stress. Pharmacol. Res. 2010, 61, 281–287. [Google Scholar] [CrossRef]
- Qian, L.B.; Wang, H.P.; Qiu, W.L.; Huang, H.; Bruce, I.C.; Xia, Q. Interleukin-2 protects against endothelial dysfunction induced by high glucose levels in rats. Vascul. Pharmacol. 2006, 45, 374–382. [Google Scholar] [CrossRef]
- Salheen, S.M.; Panchapakesan, U.; Pollock, C.A.; Woodman, O.L. The DPP-4 inhibitor linagliptin and the GLP-1 receptor agonist exendin-4 improve endothelium-dependent relaxation of rat mesenteric arteries in the presence of high glucose. Pharmacol. Res. 2015, 94, 26–33. [Google Scholar] [CrossRef]
- Taylor, P.D.; Poston, L. The effect of hyperglycaemia on function of rat isolated mesenteric resistance artery. Br. J. Pharmacol. 1994, 113, 801–808. [Google Scholar] [CrossRef]
- Habiyakare, B.; Alsaadon, H.; Mathai, M.L.; Hayes, A.; Zulli, A. Reduction of angiotensin A and alamandine vasoactivity in the rabbit model of atherogenesis: Differential effects of alamandine and Ang(1-7). Int. J. Exp. Pathol. 2014, 95, 290–295. [Google Scholar] [CrossRef]
- Zulli, A.; Hare, D.L. High dietary methionine plus cholesterol stimulates early atherosclerosis and late fibrous cap development which is associated with a decrease in GRP78 positive plaque cells. Int. J. Exp. Pathol. 2009, 90, 311–320. [Google Scholar] [CrossRef] [PubMed]
- El-Hawli, A.; Qaradakhi, T.; Hayes, A.; Rybalka, E.; Smith, R.; Caprnda, M.; Opatrilova, R.; Gazdikova, K.; Benckova, M.; Kruzliak, P.; et al. IRAP inhibition using HFI419 prevents moderate to severe acetylcholine mediated vasoconstriction in a rabbit model. Biomed. Pharmacother. 2017, 86, 23–26. [Google Scholar] [CrossRef] [PubMed]
- Smith, R.M.; Rai, S.; Kruzliak, P.; Hayes, A.; Zulli, A. Putative Nox2 inhibitors worsen homocysteine-induced impaired acetylcholine-mediated relaxation. Nutr. Metab. Cardiovasc. Dis. 2019, 29, 856–864. [Google Scholar] [CrossRef] [PubMed]
- Arora, R.; Hare, D.L.; Zulli, A. Simvastatin reduces endothelial NOS: Caveolin-1 ratio but not the phosphorylation status of eNOS in vivo. J. Atheroscler. Thromb. 2012, 19, 705–711. [Google Scholar] [CrossRef]
- Zulli, A.; Buxton, B.F.; Black, M.J.; Ming, Z.; Cameron, A.; Hare, D.L. The immunoquantification of caveolin-1 and eNOS in human and rabbit diseased blood vessels. J. Histochem. Cytochem. 2006, 54, 151–159. [Google Scholar] [CrossRef]
- Qaradakhi, T.; Matsoukas, M.T.; Hayes, A.; Rybalka, E.; Caprnda, M.; Rimarova, K.; Sepsi, M.; Busselberg, D.; Kruzliak, P.; Matsoukas, J.; et al. Alamandine reverses hyperhomocysteinemia-induced vascular dysfunction via PKA-dependent mechanisms. Cardiovasc. Ther. 2017, 35, e12306. [Google Scholar] [CrossRef]
- Zulli, A.; Lau, E.; Wijaya, B.P.; Jin, X.; Sutarga, K.; Schwartz, G.D.; Learmont, J.; Wookey, P.J.; Zinellu, A.; Carru, C.; et al. High dietary taurine reduces apoptosis and atherosclerosis in the left main coronary artery: Association with reduced CCAAT/enhancer binding protein homologous protein and total plasma homocysteine but not lipidemia. Hypertension 2009, 53, 1017–1022. [Google Scholar] [CrossRef]
- Zulli, A.; Hare, D.L.; Buxton, B.F.; Widdop, R.E. Vasoactive role for angiotensin II type 2 receptors in human radial artery. Int. J. Immunopathol. Pharmacol. 2014, 27, 79–85. [Google Scholar] [CrossRef] [PubMed]
- Zulli, A.; Burrell, L.M.; Widdop, R.E.; Black, M.J.; Buxton, B.F.; Hare, D.L. Immunolocalization of ACE2 and AT2 receptors in rabbit atherosclerotic plaques. J. Histochem. Cytochem. 2006, 54, 147–150. [Google Scholar] [CrossRef]
- Zulli, A.; Burrell, L.M.; Buxton, B.F.; Hare, D.L. ACE2 and AT4R are present in diseased human blood vessels. Eur. J. Histochem. 2008, 52, 39–44. [Google Scholar] [CrossRef][Green Version]
- Rodevand, L.; Steen, N.E.; Elvsashagen, T.; Quintana, D.S.; Reponen, E.J.; Morch, R.H.; Lunding, S.H.; Vedal, T.S.J.; Dieset, I.; Melle, I.; et al. Cardiovascular risk remains high in schizophrenia with modest improvements in bipolar disorder during past decade. Acta Psychiatr. Scand. 2019, 139, 348–360. [Google Scholar] [CrossRef] [PubMed]
- Silva, A.S.E.; Lacerda, F.V.; da Mota, M.P.G. Effect of Strength Training on Plasma Levels of Homocysteine in Patients with Type 2 Diabetes. Int. J. Prev. Med. 2019, 10, 80. [Google Scholar] [CrossRef] [PubMed]
- Maylor, B.D.; Zakrzewski-Fruer, J.K.; Stensel, D.J.; Orton, C.J.; Bailey, D.P. Effects of Frequency and Duration of Interrupting Sitting on Cardiometabolic Risk Markers. Int. J. Sports Med. 2019, 40, 818–824. [Google Scholar] [CrossRef]
- Cohen, J. Statistical Power Analysis for the Behavioral Sciences; Routledge: Abingdon, UK, 2013. [Google Scholar]
- Guo, X.; Liu, W.L.; Chen, L.W.; Guo, Z.G. High glucose impairs endothelium-dependent relaxation in rabbit aorta. Acta Pharmacol. Sin. 2000, 21, 169–173. [Google Scholar] [PubMed]
- VanderLaan, P.A.; Reardon, C.A.; Getz, G.S. Site specificity of atherosclerosis: Site-selective responses to atherosclerotic modulators. Arterioscler. Thromb. Vasc. Biol. 2004, 24, 12–22. [Google Scholar] [CrossRef]
- Traub, O.; Berk, B.C. Laminar shear stress: Mechanisms by which endothelial cells transduce an atheroprotective force. Arterioscler. Thromb. Vasc. Biol. 1998, 18, 677–685. [Google Scholar] [CrossRef]
- Motwani, J.G.; Topol, E.J. Aortocoronary saphenous vein graft disease: Pathogenesis, predisposition, and prevention. Circulation 1998, 97, 916–931. [Google Scholar] [CrossRef]
- Ozkor, M.A.; Murrow, J.R.; Rahman, A.M.; Kavtaradze, N.; Lin, J.; Manatunga, A.; Quyyumi, A.A. Endothelium-derived hyperpolarizing factor determines resting and stimulated forearm vasodilator tone in health and in disease. Circulation 2011, 123, 2244–2253. [Google Scholar] [CrossRef]
- Jamwal, S.; Sharma, S. Vascular endothelium dysfunction: A conservative target in metabolic disorders. Inflamm. Res. 2018, 67, 391–405. [Google Scholar] [CrossRef]
- Sena, C.M.; Pereira, A.M.; Seica, R. Endothelial dysfunction - a major mediator of diabetic vascular disease. Biochim. Biophys. Acta 2013, 1832, 2216–2231. [Google Scholar] [CrossRef]
- Ceriello, A. Postprandial hyperglycemia and diabetes complications: Is it time to treat? Diabetes 2005, 54, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Rai, S.; Hare, D.L.; Zulli, A. A physiologically relevant atherogenic diet causes severe endothelial dysfunction within 4 weeks in rabbit. Int. J. Exp. Pathol. 2009, 90, 598–604. [Google Scholar] [CrossRef]
- Alarcon, G.; Roco, J.; Medina, M.; Medina, A.; Peral, M.; Jerez, S. High fat diet-induced metabolically obese and normal weight rabbit model shows early vascular dysfunction: Mechanisms involved. Int. J. Obes. 2018, 42, 1535–1543. [Google Scholar] [CrossRef] [PubMed]
- Parry, S.A.; Turner, M.C.; Woods, R.M.; James, L.J.; Ferguson, R.A.; Cocks, M.; Whytock, K.L.; Strauss, J.A.; Shepherd, S.O.; Wagenmakers, A.J.M.; et al. High-fat overfeeding impairs peripheral glucose metabolism and muscle microvascular eNOS Ser1177 phosphorylation. J. Clin. Endocrinol. Metab. 2019, 105, 65–77. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.W.; Benzie, I.F.; Ma, S.W.; Strain, J.J.; Hannigan, B.M. Acute hyperglycemia and oxidative stress: Direct cause and effect? Free Radic. Biol. Med. 2008, 44, 1217–1231. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Abdominal Aorta | n | Log EC50 ± SEM | p vs. Con | p vs. Con | Emax ± SEM | p vs. Con | d vs. Con | AUC ± SEM | p vs. Con | d vs. Con |
|---|---|---|---|---|---|---|---|---|---|---|
| ND Con | 7 | −7.59 ± 0.12 | −81 ±13 | 196 ± 18 | ||||||
| ND 20 mM | 7 | −7.43 ± 0.09 | n/s | 0.54 | −70 ± 11 | n/s | 0.32 | 160 ± 11 | 0.08 ^ | 0.92 |
| ND 40 mM | 7 | −7.65 ± 0.1 | n/s | 0.18 | −73 ± 2 | n/s | 0.29 | 177 ± 12 | n/s | 0.48 |
| AD Con | 10 | −7.10 ± 0.13 | −70 ± 2 | 146 ± 13 | ||||||
| AD 20 mM | 11 | −6.81 ± 0.06 | 0.03 * | 0.88 | −78 ± 3 | 0.04 * | 0.97 | 141 ± 9 | n/s | 0.13 |
| AD 40 mM | 11 | −6.93 ± 0.07 | n/s | 0.52 | −71 ± 3 | n/s | 0.18 | 138 ± 10 | n/s | 0.21 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tacey, A.; Qaradakhi, T.; Smith, C.; Pittappillil, C.; Hayes, A.; Zulli, A.; Levinger, I. The Effect of an Atherogenic Diet and Acute Hyperglycaemia on Endothelial Function in Rabbits Is Artery Specific. Nutrients 2020, 12, 2108. https://doi.org/10.3390/nu12072108
Tacey A, Qaradakhi T, Smith C, Pittappillil C, Hayes A, Zulli A, Levinger I. The Effect of an Atherogenic Diet and Acute Hyperglycaemia on Endothelial Function in Rabbits Is Artery Specific. Nutrients. 2020; 12(7):2108. https://doi.org/10.3390/nu12072108
Chicago/Turabian StyleTacey, Alexander, Tawar Qaradakhi, Cassandra Smith, Chris Pittappillil, Alan Hayes, Anthony Zulli, and Itamar Levinger. 2020. "The Effect of an Atherogenic Diet and Acute Hyperglycaemia on Endothelial Function in Rabbits Is Artery Specific" Nutrients 12, no. 7: 2108. https://doi.org/10.3390/nu12072108
APA StyleTacey, A., Qaradakhi, T., Smith, C., Pittappillil, C., Hayes, A., Zulli, A., & Levinger, I. (2020). The Effect of an Atherogenic Diet and Acute Hyperglycaemia on Endothelial Function in Rabbits Is Artery Specific. Nutrients, 12(7), 2108. https://doi.org/10.3390/nu12072108